Assessment of the Standardized Precipitation and Evaporation Index (SPEI) as a Potential Management Tool for Grasslands

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
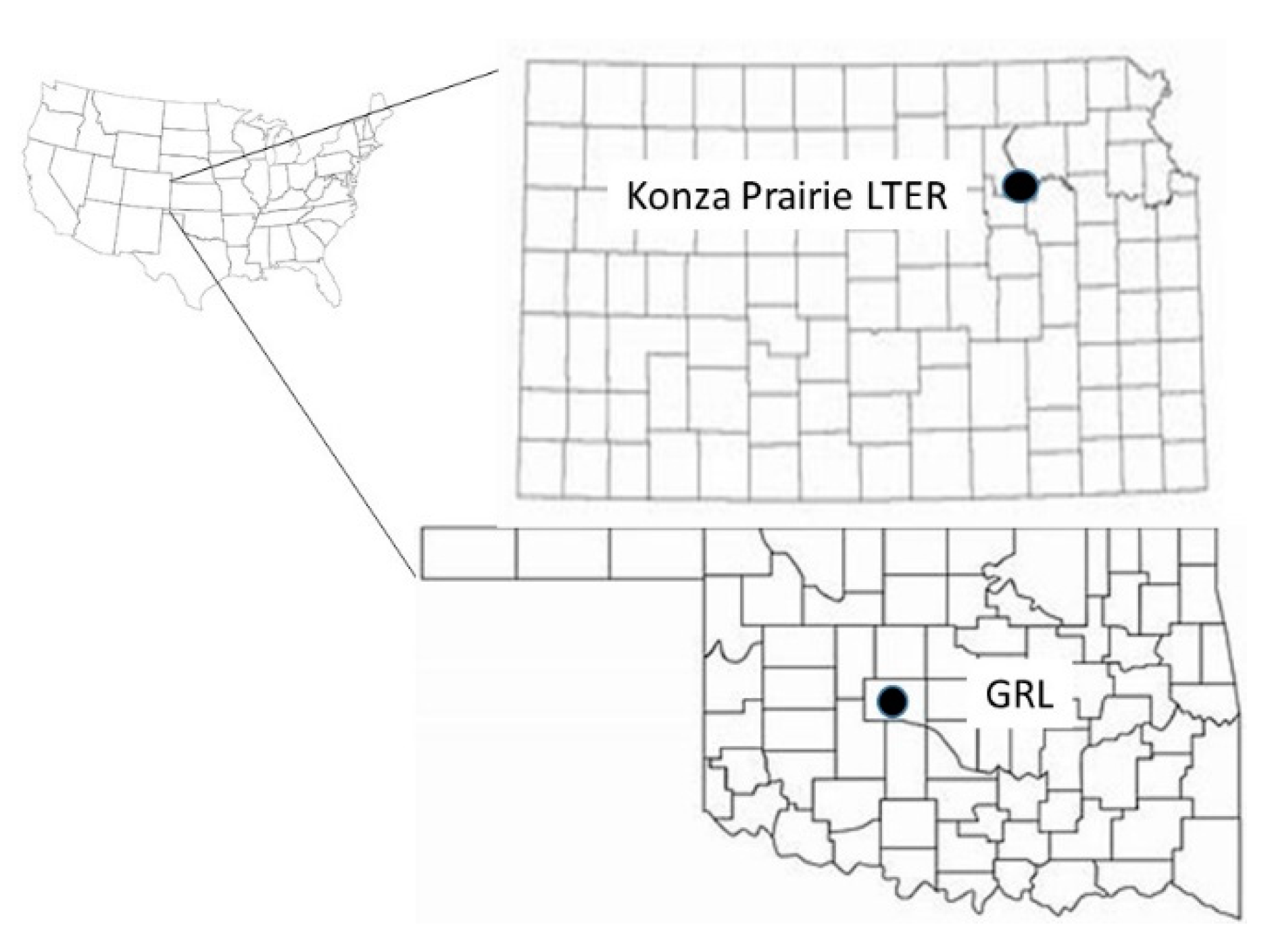
2.1. Study Sites
2.2. Above-Ground Forage Mass
2.3. Standardized Precipitation and Evaporation Index (SPEI)
2.4. Statistical Analysis
3. Results
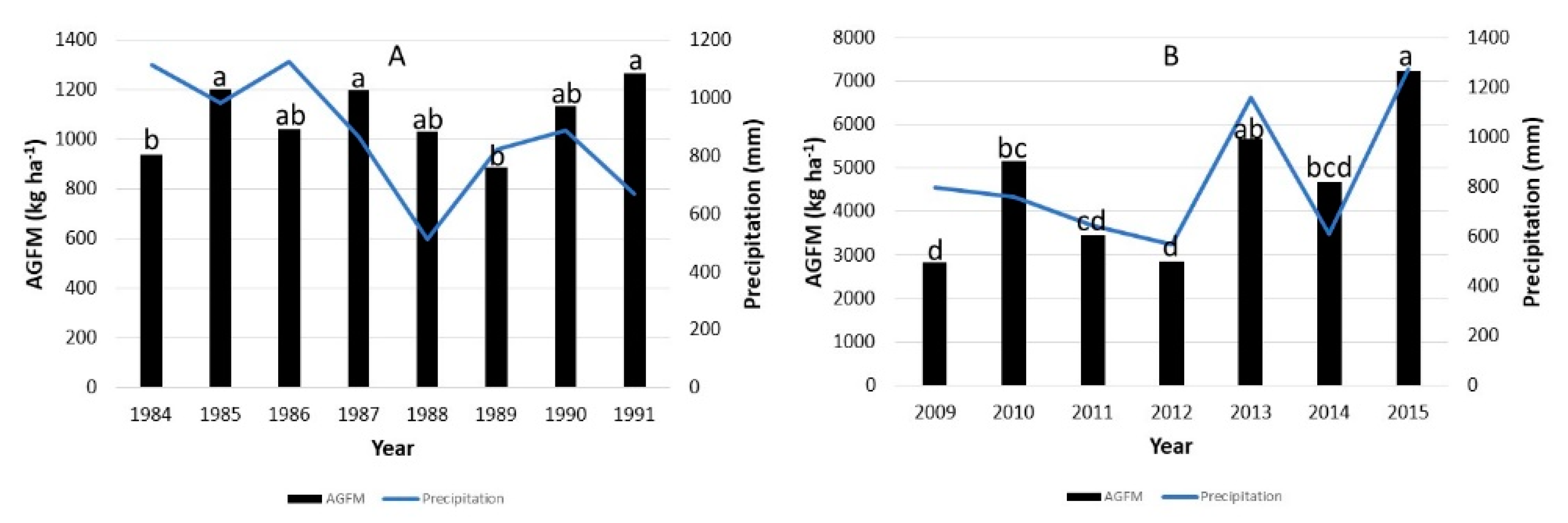
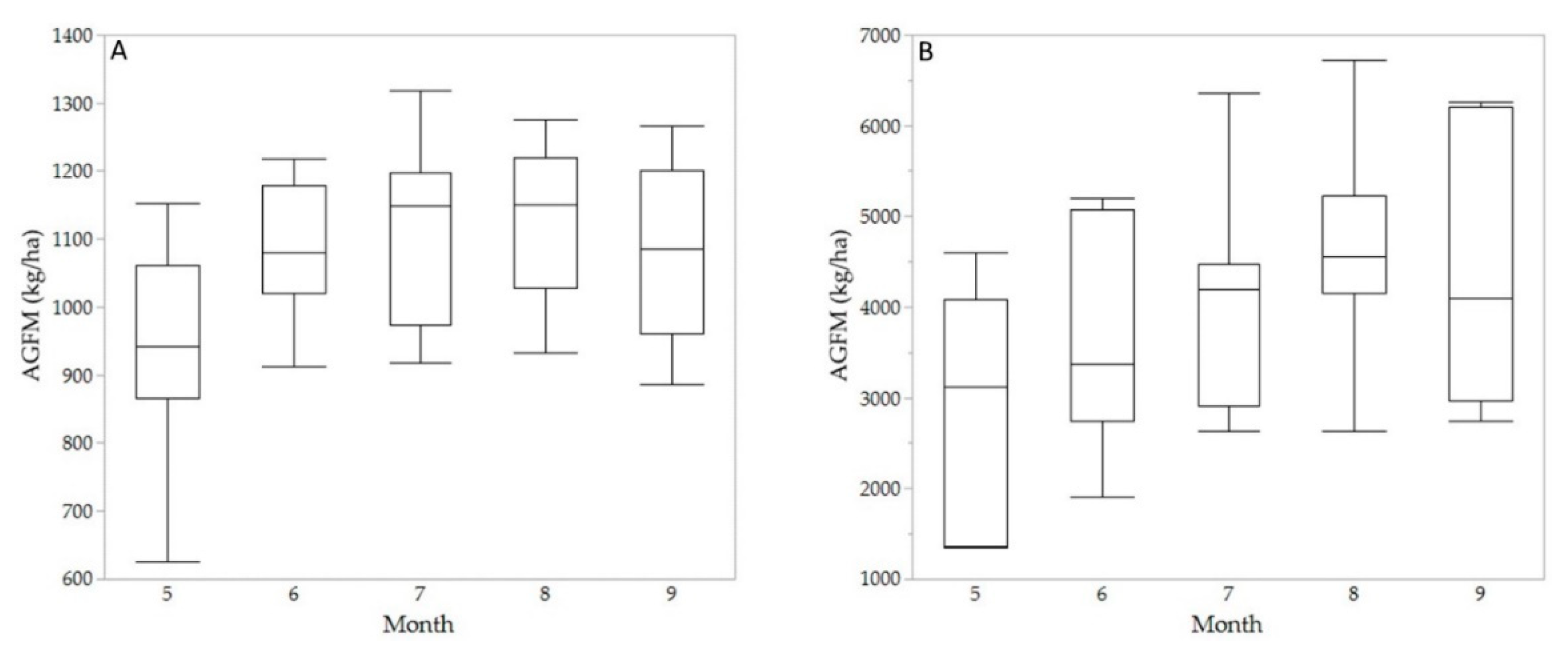
3.1. Above-Ground Forage Mass
3.2. Discriminant Analysis
3.3. Neural Networks
4. Discussion
4.1. Study Context
4.2. Discriminant Analysis
4.3. Artificial Neural Network
4.4. AGFM and AGFM Classes
4.5. SPEI
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Steiner, J.L.; Schneider, J.M.; Pope, C.; Pope, S.; Ford, P.; Steele, R.F. Southern Plains Assessment of Vulnerability and Preliminary Adaptation and Mitigation Strategies for Farmers, Ranchers, and Forest Land Owners; Anderson, T., Ed.; United States Department of Agriculture: Washington, DC, USA, 2017; 61p.
- National Agricultural Statistics Service. 2012 Census of Agriculture, Texas State and County Data; Geographic Area Series; Part 43A, AC-12-A-43A; United States Department of Agriculture, National Agricultural Statistics Service: Washington, DC, USA, 2014; Volume 1.
- National Agricultural Statistics Service. 2012 Census of Agriculture, Oklahoma State and County Data; Geographic Area Series; Part 36, AC-12-A-36; United States Department of Agriculture, National Agricultural Statistics Service: Washington, DC, USA, 2014; Volume 1.
- National Agricultural Statistics Service. 2012 Census of Agriculture, Oklahoma State and County Data; Geographic Area Series; Part 16, AC-12-A-16; United States Department of Agriculture, National Agricultural Statistics Service: Washington, DC, USA, 2014; Volume 1.
- Guerrero, B. The Impact of Agricultural Drought Losses on the Texas Economy. 2011. Available online: https://agecoext.tamu.edu/wp-content/uploads/2013/07/BriefingPaper09-01-11.pdf (accessed on 6 May 2019).
- Ranking of States with The Most Cattle. Available online: http://beef2live.com/story-ranking-states-cattle-0-108182 (accessed on 6 April 2019).
- Kunkel, K.E.; Stevens, L.E.; Stevens, S.E.; Sun, L.; Janssen, E.; Wuebbles, D.; Kruk, M.C.; Thomas, D.P.; Shulski, M.; Umphlett, N.; et al. Regional Climate Trends and Scenarios for the U.S. National Climate Assessment Part 4. Climate of the U.S. Great Plains; NOAA Technical Report; NESDIS: Washington, DC, USA, 2013; pp. 142–144.
- Dukes, J.S.; Chiariello, N.R.; Cleland, E.E.; Moore, L.A.; Shaw, M.R.; Thayer, S.; Tobeck, T.; Mooney, H.A.; Field, C.B. Responses of Grassland Production to Single and Multiple Global Environmental Changes. PLoS Biol. 2005, 3, e319. [Google Scholar] [CrossRef] [PubMed]
- Fay, P.A.; Carlisle, J.D.; Knapp, A.K.; Blair, J.M.; Collins, S.L. Productivity responses to altered rainfall patterns in a C4-dominated grassland. Oecologia 2003, 37, 245–251. [Google Scholar] [CrossRef]
- Knapp, A.K.; Smith, M.D. Variation among biomes in temporal dynamics of aboveground primary production. Science 2001, 291, 481–484. [Google Scholar] [CrossRef] [PubMed]
- Dale, R.F.; Shaw, R.H. Effect on corn yields of moisture stress and stand at two different fertility levels. Agron. J. 1965, 57, 475–479. [Google Scholar] [CrossRef]
- Boonjung, H.; Fukai, S. Effects of soil water deficit at different growth stages on rice growth and yield under upland conditions. 2. Phenology, biomass production and yield. Field Crops Res. 1996, 48, 47–55. [Google Scholar] [CrossRef]
- Earl, H.J.; Davis, R.F. Effect of drought stress on leaf and whole canopy maize radiation use efficiency and yield of maize. Agron. J. 2003, 95, 688–696. [Google Scholar] [CrossRef]
- Flanagan, L.B.; Johnson, B.G. Interacting effects of temperature, soil moisture, and plant biomass production on ecosystem respiration in a northern temperate grassland. Agric. For. Meteor. 2005, 130, 237–253. [Google Scholar] [CrossRef]
- Palmer, W.C. Meteorological Drought; Research Paper 45; US Weather Bureau, US Department of Commerce: Silver Spring, MD, USA, 1965; 58p.
- McKee, T.B.; Doesken, N.J.; Kleist, J. The Relationship of Drought Frequency and Duration to Time Scales. In Proceedings of the 8th Conference on Applied Climatology, Anaheim, CA, USA, 17–22 January 1993; American Meteorological Society: Boston, MA, USA, 1993. [Google Scholar]
- Bergman, K.H.; Sabol, P.; Miskus, D. Experimental indices for monitoring global drought conditions. In Proceedings of the 13th Annual Climate Diagnostics Workshop, Cambridge, MA, USA, 31 October–4 November 1988; US Department of Commerce: Washington, DC, USA. [Google Scholar]
- Modares, R. Streamflow drought time series forecasting. Stoch. Environ. Res. Risk Assess. 2007, 21, 223–233. [Google Scholar] [CrossRef]
- Anderson, M.C.; Hain, C.; Wardlow, B.; Pimstein, A.; Mecikalski, J.R.; Kustas, W.P. Evaluation of drought indices based on thermal remote sensing of evapotranspiration over the continental United States. J. Clim. 2011, 24, 2025–2044. [Google Scholar] [CrossRef]
- Svoboda, M.; Fuchs, B.A. Handbook of Drought Indicators and Indices, Integrated Drought Management Programme (IDMP), Integrated Drought Management Tools and Guidelines Series 2; WMO-No. 2273; World Meteorological Organization (WMO): Geneva, Switzerland, 2016; 45p. [Google Scholar]
- Vicente-Serrano, S.M.; Begueria, S.; Lopez-Moreno, J.I. A multi-scalar drought index sensitive to global warming: The standardized precipitation evapotranspiration index–SPEI. J. Clim. 2010, 23, 1696–1718. [Google Scholar] [CrossRef]
- Wilhite, D.; Glantz, M. Understanding the drought phenomenon: The role of definitions. Water Int. 1985, 10, 111–120. [Google Scholar] [CrossRef]
- Sivakumar, M.V.K.; Motha, R.P.; Wilhite, D.A.; Wood, D.A. Agricultural drought indices. In Proceedings of the A WMO/UNISDR Expert Group Meeting on Agricultural Drought Indices, Murcia, Spain, 1–4 June 2010. (AGM-11, WMO/TD No. 1572; WAOB-2011). [Google Scholar]
- Hunt, E.D.; Svoboda, M.; Wardlow, B.; Hubbard, K.; Hayes, M.; Arkebauer, T. Monitoring the effects of rapid onset of drought on non-irrigated maize with agronomic data and climate-based drought indices. Agric. For. Meteor. 2014, 191, 1–11. [Google Scholar] [CrossRef]
- Chen, T.; van der Werf, G.R.; de Ju, R.A.M.; Wang, G.; Dolman, A.J. A global analysis of the impact of drought on net primary productivity. Hydrol. Earth Syst. Sci. 2013, 17, 3885–3894. [Google Scholar] [CrossRef] [Green Version]
- Moorehead, J.E.; Gowda, P.H.; Singh, V.P.; Porter, D.O.; Marek, T.H.; Howell, T.A.; Stewar, B.A. Identifying and evaluating a suitable index for agricultural drought monitoring in the Texas High Plains. J. Am. Water Resour. Assoc. 2015, 51, 807–820. [Google Scholar] [CrossRef]
- Potop, V.; Mozny, M.; Soukup, J. Drought evolution at various time scales in the lowland regions and their impact on vegetable crops in the Czech Repubic. Agric. For. Meteor. 2012, 156, 121–133. [Google Scholar] [CrossRef]
- Potopova, V.; Stepanek, P.; Mozny, M.; Turkott, L.; Soukup, J. Performance of the standardised precipitation evapotranspiration index at various lags for agricultural drought risk assessment in the Czech Republic. Agric. For. Meteor. 2015, 202, 26–38. [Google Scholar] [CrossRef]
- Ogaya, R.; Barbeta, A.; Basnou, C.; Penuelas, J. Satellite data as indicators of tree biomass growth and forest dieback in a Mediterranean holm oak forest. Ann. For. Sci. 2015, 72, 135–144. [Google Scholar] [CrossRef]
- Klesse, S.; Ettold, S.; Frank, D. Integrating tree-ring and inventory-based measurements of aboveground biomass growth: Research opportunities and carbon cycle consequences from a large snow breakage event in the Swiss Alps. Eur. J. For. Res. 2016, 135, 297–311. [Google Scholar] [CrossRef]
- Liu, S.; Zhang, Y.; Cheng, F.; Hou, X.; Zhao, S. Response of grassland degradation to drought at different time-scales in Qinghai Province: Spatio-temporal characteristics, correlation, and implications. Rem. Sen. 2017, 9, 1329. [Google Scholar] [CrossRef]
- Barnes, M.L.; Moran, M.S.; Scott, R.L.; Kolb, T.E.; Ponce-Campos, G.E.; Moore, D.J.P.; Ross, M.A.; Mitra, B.; Dore, S. Vegetation productivity responds to sub-annual climate conditions across semiarid biomes. Ecosphere 2016, 7, e0.119. [Google Scholar] [CrossRef]
- Knapp, A.K.; Carroll, C.J.W.; Denton, E.M.; La Pierre, K.J.; Collins, S.L.; Smith, M.D. Differential sensitivity to regional-scale drought in six central US grasslands. Oecologia 2015, 177, 949–957. [Google Scholar] [CrossRef] [PubMed]
- Northup, B.K.; Daniel, J.A. Impact of climate and management on species composition of southern tallgrass prairie in Oklahoma. In Proceedings of the 1st National Conference on Grazing Lands, Las Vega, NV, USA, 5–8 December 2000. [Google Scholar]
- Norhtup, B.K.; Schneider, J.M.; Daniel, J.A. The effects of management and precipitation on forage composition of a southern tallgrass prairie. In Proceedings of the 15th Conference on Biometeorology and Aerobiology, Kansas City, MO, USA, 27 October–1 November 2002. [Google Scholar]
- Knapp, A. PAB02 Biweekly Measurement of Aboveground Net Primary Productivity on an Unburned and Annually Burned Watershed. Environ. Data Initiat. 2018. Available online: http://129.130.186.12/content/pab02-biweekly-measurement-aboveground-net-primary-productivity-unburned-and-annually-burned (accessed on 17 April 2019).
- Shapiro, S.S.; Wilk, M.B. An analysis of variance test for normality (complete samples). Biometrika 1965, 52, 591–611. [Google Scholar] [CrossRef]
- Slifker, J.F.; Shapiro, S.S. The Johnson system: Selection and parameter estimation. Technometrics 1980, 22, 239–246. [Google Scholar] [CrossRef]
- Tukey, J.W. The Problem of Multiple Comparisons. In Multiple Comparisons, 1948–1983; Volume 8 of The Collected Works of John W. Tukey. Unpublished manuscript; Braun, H.I., Ed.; Chapman & Hall: London, UK, 1994; pp. 1–300. [Google Scholar]
- Allen, R.G.; Pereira, L.S.; Raes, D.; Smith, M. Crop Evapotranspiration–Guidelines for Computing Crop Water Requirements; FAO Irrigation and Drainage Paper 56; Food and Agriculture Organization: Rome, Italy, 1998. [Google Scholar]
- Johnson, R.A.; Wichern, D.W. Applied Multivariate Statistical Analysis, 2nd ed.; Prentice-Hall Inc.: Englewood Cliffs, NJ, USA, 1988. [Google Scholar]
- Lawrence, J. Introduction to Neural Networks: Design, Theory, and Applications, 6th ed.; California Scientific Software: Nevada City, CA, USA, 1994. [Google Scholar]
- Abrams, M.D.; Knapp, A.K.; Hulbert, L.C. A ten-year record of aboveground biomass in a Kansas tallgrass prairie: Effects of fire and topographic position. Am. J. Bot. 1986, 73, 1509–1515. [Google Scholar] [CrossRef]
- Wiles, L.J.; Dunn, G.; Printz, J.; Patton, B.; Nyren, A. Spring precipitation as a predictor for peak standing crop of mixed-grass prairie. Rangel. Ecol. Manag. 2011, 64, 215–222. [Google Scholar] [CrossRef]
- Andales, A.A.; Derner, J.D.; Ahuja, L.R.; Hart, R.H. Strategic and tactical prediction of forage production in northern mixed-grass prairie. Rangel. Ecol. Manag. 2006, 59, 576–584. [Google Scholar] [CrossRef]
- Nippert, J.B.; Knapp, A.K.; Briggs, J.M. Intra-annual rainfall variability and grassland productivity: Can the past predict the future? Plant Ecol. 2006, 184, 65–74. [Google Scholar] [CrossRef]
- Vincente-Serrano, S.M.; Cuadrat-Prats, J.M.; Romo, A. Early prediction of crop production using drough indices at different time-scales and remote sensing data: Application in the Ebro Valley (north-east Spain). Int. J. Remote Sens. 2006, 27, 511–518. [Google Scholar] [CrossRef]
- Wang, J.; Rich, P.M.; Price, K.P.; Kettle, W.D. Relations between NDVI, grassland production, and crop yield in the central Great Plains. Geocarto Int. 2005, 20, 5–11. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| KP000a | Misclassification Rate 1 | GRL | Misclassification Rate | ||||||
|---|---|---|---|---|---|---|---|---|---|
| SPEI Months | AGFM Month | abv | avg | blw | SPEI Months | AGFM Month | abv | avg | blw |
| 1−4 | May | 0 | 17 | 0 | 1−4 | May | 0 | 0 | 0 |
| Jun | 0 | 0 | 0 | Jun | 0 | 0 | 0 | ||
| Jul | 0 | 0 | 0 | Jul | 0 | - | 0 | ||
| Aug | 0 | 0 | 0 | Aug | 0 | 0 | 0 | ||
| Sep | 0 | 0 | 0 | Sep | 33 | 0 | 0 | ||
| 1−5 | Jun | 0 | 0 | 0 | 1−5 | Jun | 0 | 0 | 0 |
| Jul | 0 | 0 | 0 | Jul | 0 | - | 0 | ||
| Aug | 0 | 0 | 0 | Aug | 33 | 0 | 0 | ||
| Sep | 0 | 0 | 0 | Sep | 33 | 0 | 33 | ||
| 1−6 | Jul | 0 | 0 | 0 | 1−6 | Jul | 33 | - | 0 |
| Aug | 0 | 0 | 0 | Aug | 33 | 50 | 0 | ||
| Sep | 0 | 0 | 0 | Sep | 33 | 0 | 0 | ||
| 1−7 | Aug | 0 | 0 | 0 | 1−7 | Aug | 33 | 50 | 50 |
| Sep | 0 | 0 | 0 | Sep | 33 | 0 | 0 | ||
| 1−8 | Sep | 0 | 25 | 0 | 1−8 | Sep | 66 | 0 | 0 |
| May | June | July | August | September | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Actual | abv | avg | blw | abv | avg | blw | abv | avg | blw | abv | avg | blw | abv | avg | blw | |
| Site = KP000a | abv | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| avg | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 0 | 1 | 0 | 1 | 0 | |
| blw | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | |
| TMR 1 | 100 | 50 | 100 | 66 | 0 | |||||||||||
| Site = GRL | abv | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 3 | 0 | 0 |
| avg | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | |
| blw | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 3 | |
| TMR | 50 | 50 | 50 | 50 | 0 | |||||||||||
| May | June | July | August | September | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Actual | abv | avg | blw | abv | avg | blw | abv | avg | blw | abv | avg | blw | abv | avg | blw | |
| Site KP000a Predicted by GRL | abv | 0 | 1 | 0 | 0 | 0 | 2 | 0 | 1 | 0 | 1 | 1 | 1 | 0 | 0 | 1 |
| avg | 3 | 0 | 3 | 1 | 1 | 3 | 3 | 2 | 0 | 0 | 1 | 2 | 3 | 0 | 1 | |
| blw | 0 | 0 | 1 | 0 | 1 | 0 | 1 | 1 | 0 | 2 | 0 | 0 | 0 | 0 | 1 | |
| TMR 1 | 88 | 88 | 75 | 75 | 83 | |||||||||||
| Site GRL Predicted by KP000a | abv | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 2 | 1 | 2 | 0 | 1 | 0 | 3 | 0 |
| avg | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 1 | |
| blw | 0 | 2 | 0 | 1 | 2 | 1 | 2 | 1 | 0 | 0 | 1 | 1 | 0 | 1 | 2 | |
| TMR | 67 | 75 | 86 | 43 | 71 | |||||||||||
| May | June | July | August | September | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Actual | abv | avg | blw | abv | avg | blw | abv | avg | blw | abv | avg | blw | abv | avg | blw | |
| ANN Calibration for Site = KP000a | abv | 1 | 0 | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 |
| avg | 0 | 5 | 0 | 0 | 3 | 0 | 0 | 5 | 0 | 0 | 3 | 0 | 0 | 2 | 1 | |
| blw | 0 | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 2 | 0 | 0 | 0 | |
| TMR 1 | 0 | 0 | 0 | 0 | 50 | |||||||||||
| Cross-Validation for Site = KP000a | abv | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 |
| avg | 0 | 1 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 1 | 0 | |
| blw | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 0 | |
| TMR | 0 | 33 | 0 | 0 | 50 | |||||||||||
| May | June | July | August | September | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Actual | abv | avg | blw | abv | avg | blw | abv | avg | blw | abv | avg | blw | abv | avg | blw | |
| ANN Calibration for Site = GRL | abv | 2 | 0 | 0 | 2 | 1 | 0 | 3 | -- | 0 | 2 | 0 | 0 | 2 | 1 | 0 |
| avg | 0 | 1 | 0 | 0 | 1 | 0 | -- | -- | -- | 0 | 2 | 0 | 0 | 0 | 0 | |
| blw | 0 | 0 | 1 | 0 | 0 | 2 | 2 | 0 | 0 | 0 | 0 | 2 | 0 | 3 | 0 | |
| TMR 1 | 0 | 17 | 40 | 0 | 67 | |||||||||||
| Cross-Validation for Site = GRL | abv | 2 | 0 | 0 | 0 | 0 | 0 | 0 | -- | 1 | 1 | 0 | 0 | 0 | 0 | 0 |
| avg | 0 | 0 | 0 | 0 | 0 | 0 | -- | -- | -- | 0 | 0 | 0 | 0 | 1 | 0 | |
| blw | 1 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | |
| TMR | 33 | 0 | 50 | 0 | 0 | |||||||||||
| May | June | July | August | September | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Actual | abv | avg | blw | abv | avg | blw | abv | avg | blw | abv | avg | blw | abv | avg | blw | |
| GRL ANN Prediction of Site KP000a | abv | 1 | 0 | 0 | 2 | 0 | 0 | 1 | 0 | 1 | 1 | 1 | 0 | 0 | 1 | 0 |
| avg | 1 | 5 | 0 | 2 | 3 | 0 | 1 | 4 | 0 | 0 | 4 | 0 | 0 | 4 | 0 | |
| blw | 1 | 0 | 0 | 0 | 1 | 0 | 1 | 1 | 0 | 0 | 2 | 0 | 0 | 1 | 0 | |
| TMR 1 | 25 | 38 | 38 | 38 | 33 | |||||||||||
| Site KP000a ANN Prediction of GRL | abv | 3 | 0 | 0 | 3 | 0 | 0 | 4 | 0 | 0 | 3 | 0 | 0 | 2 | 1 | 0 |
| avg | 0 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 1 | |
| blw | 1 | 1 | 0 | 2 | 1 | 0 | 3 | 0 | 0 | 1 | 1 | 0 | 1 | 0 | 2 | |
| TMR | 33 | 57 | 43 | 43 | 43 | |||||||||||
| May | June | July | August | September | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Actual | abv | avg | blw | abv | avg | blw | abv | avg | blw | abv | avg | blw | abv | avg | blw | |
| ANN Calibration for Combined Site Datasets | abv | 1 | 1 | 1 | 1 | 1 | 1 | 2 | 0 | 2 | 1 | 0 | 3 | 1 | 1 | 1 |
| avg | 1 | 3 | 0 | 3 | 0 | 0 | 1 | 4 | 0 | 0 | 4 | 1 | 1 | 3 | 1 | |
| blw | 0 | 2 | 0 | 2 | 1 | 0 | 1 | 0 | 3 | 1 | 1 | 1 | 0 | 0 | 3 | |
| TMR 1 | 56 | 89 | 31 | 50 | 36 | |||||||||||
| ANN Cross-Validation for Combined Site Datasets | abv | 1 | 0 | 0 | 2 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 0 |
| avg | 0 | 3 | 0 | 0 | 2 | 1 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | |
| blw | 0 | 1 | 0 | 1 | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 1 | |
| TMR | 20 | 33 | 0 | 33 | 0 | |||||||||||
| Statistic | 1SPEI1 | 1SPEI2 | 1SPEI3 | 1SPEI4 |
|---|---|---|---|---|
| Site = KP000a | ||||
| Maximum | 1.85 | 1.14 | 0.99 | 1.95 |
| Minimum | −0.89 | −1.69 | −1.66 | −1.20 |
| Range | 2.74 | 2.83 | 2.65 | 3.15 |
| Site = GRL | ||||
| Maximum | 0.80 | 1.09 | 1.3 | 1.49 |
| Minimum | −1.37 | −0.75 | −1.42 | −1.35 |
| Range | 2.17 | 1.84 | 2.72 | 2.84 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Starks, P.J.; Steiner, J.L.; Neel, J.P.S.; Turner, K.E.; Northup, B.K.; Gowda, P.H.; Brown, M.A. Assessment of the Standardized Precipitation and Evaporation Index (SPEI) as a Potential Management Tool for Grasslands. Agronomy 2019, 9, 235. https://doi.org/10.3390/agronomy9050235
Starks PJ, Steiner JL, Neel JPS, Turner KE, Northup BK, Gowda PH, Brown MA. Assessment of the Standardized Precipitation and Evaporation Index (SPEI) as a Potential Management Tool for Grasslands. Agronomy. 2019; 9(5):235. https://doi.org/10.3390/agronomy9050235
Chicago/Turabian StyleStarks, Patrick J., Jean L. Steiner, James P. S. Neel, Kenneth E. Turner, Brian K. Northup, Prasanna H. Gowda, and Michael A. Brown. 2019. "Assessment of the Standardized Precipitation and Evaporation Index (SPEI) as a Potential Management Tool for Grasslands" Agronomy 9, no. 5: 235. https://doi.org/10.3390/agronomy9050235