Climate Effects on Tallgrass Prairie Responses to Continuous and Rotational Grazing


 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
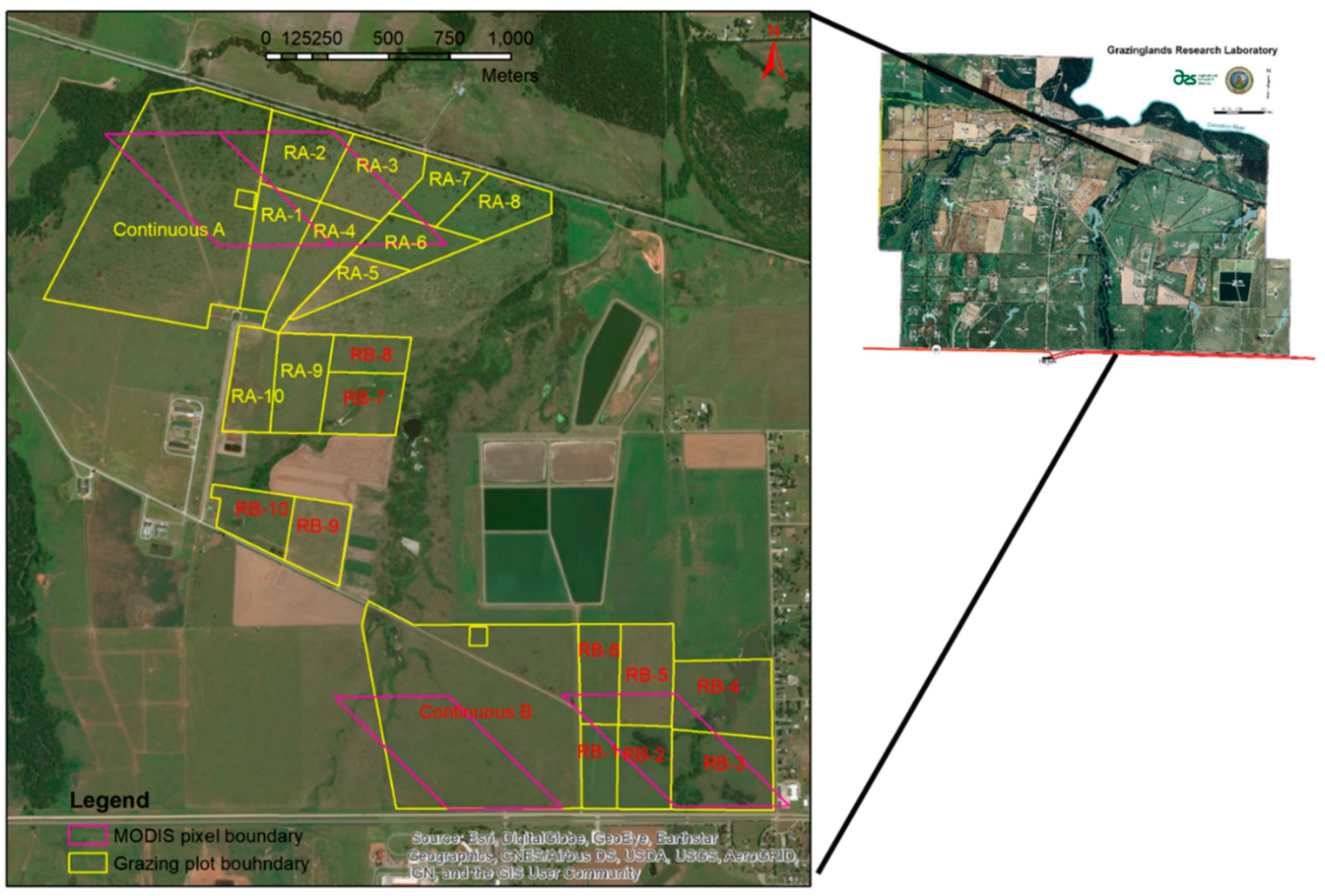
2.1. Study Area and Experiment Design
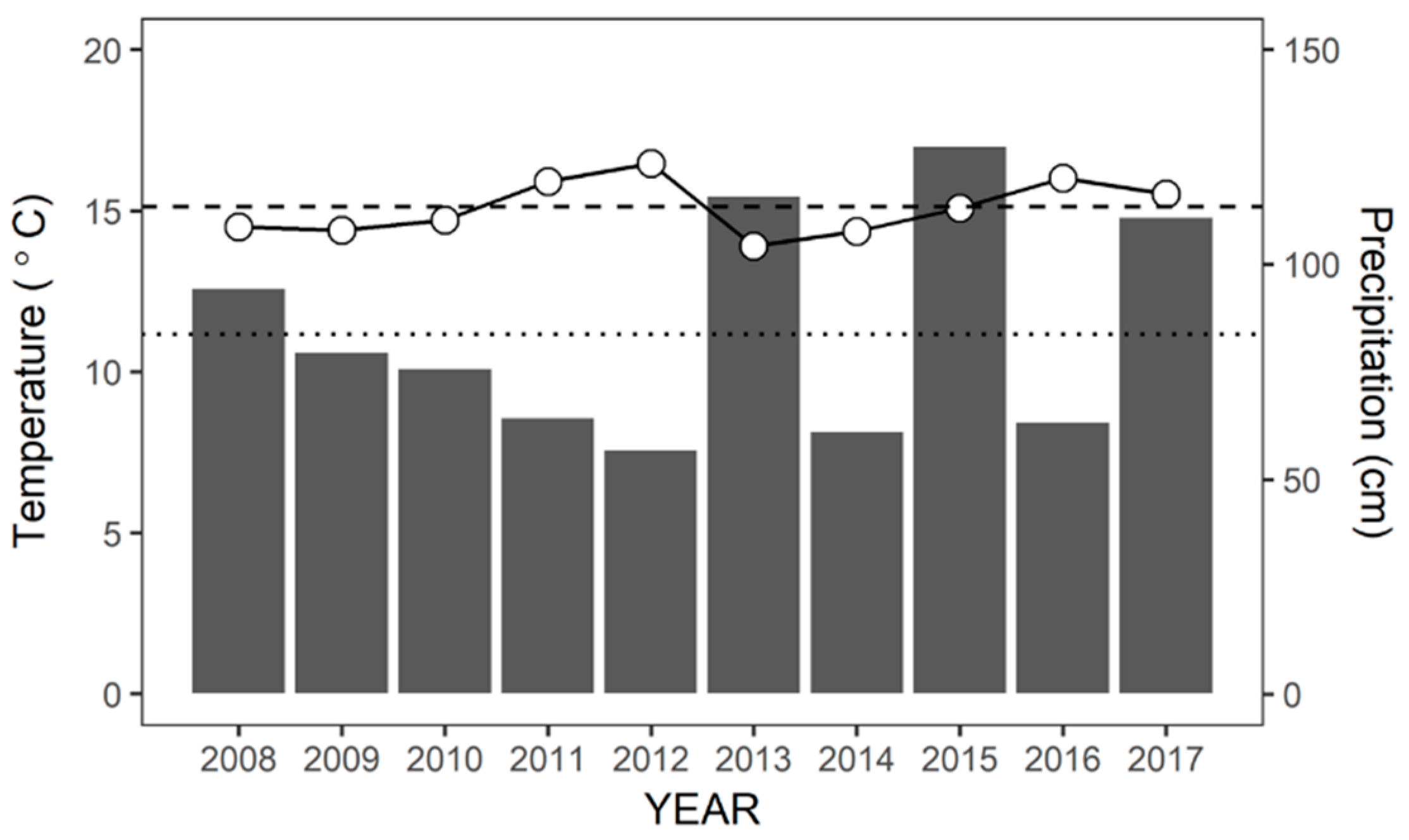
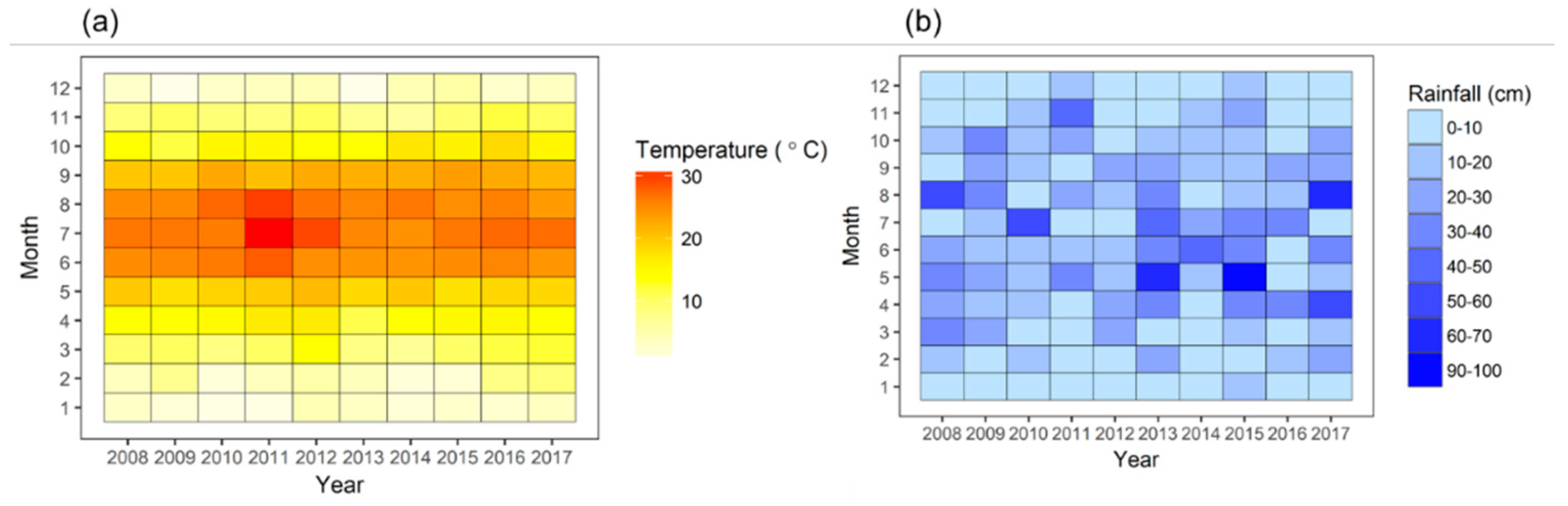
2.2. Climate Data
2.3. Landsat Images and the Enhanced Vegetation Index (EVI)
2.4. Gross Primary Production (GPP) from the Vegetation Photosynthesis Model
2.5. Statistical Analysis
3. Results
3.1. Annual and Seasonal Variations of Ta and P
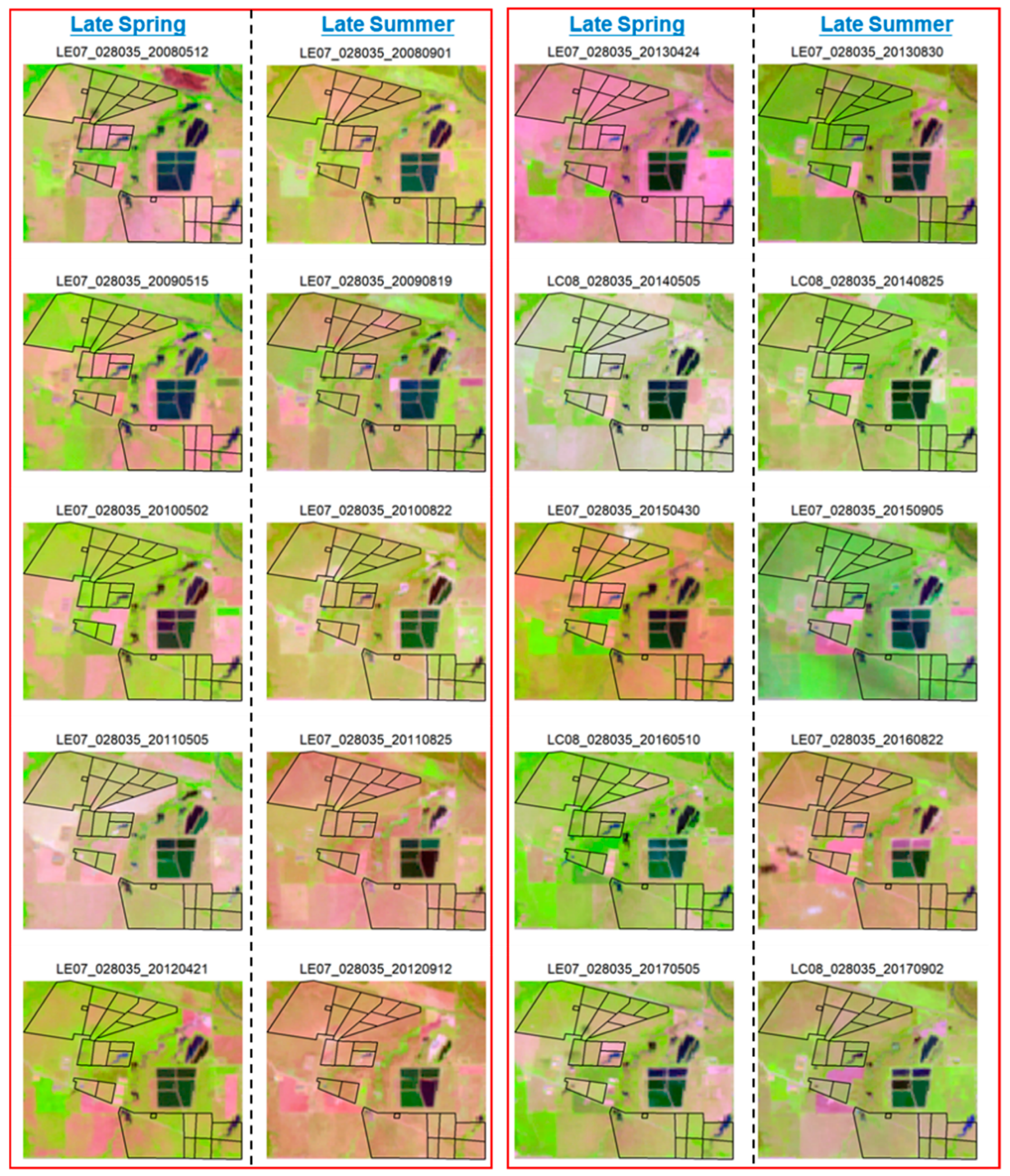
3.2. A glimpse of the Continuous and Rotational Grazing Paddocks in Late Spring and Late Summer
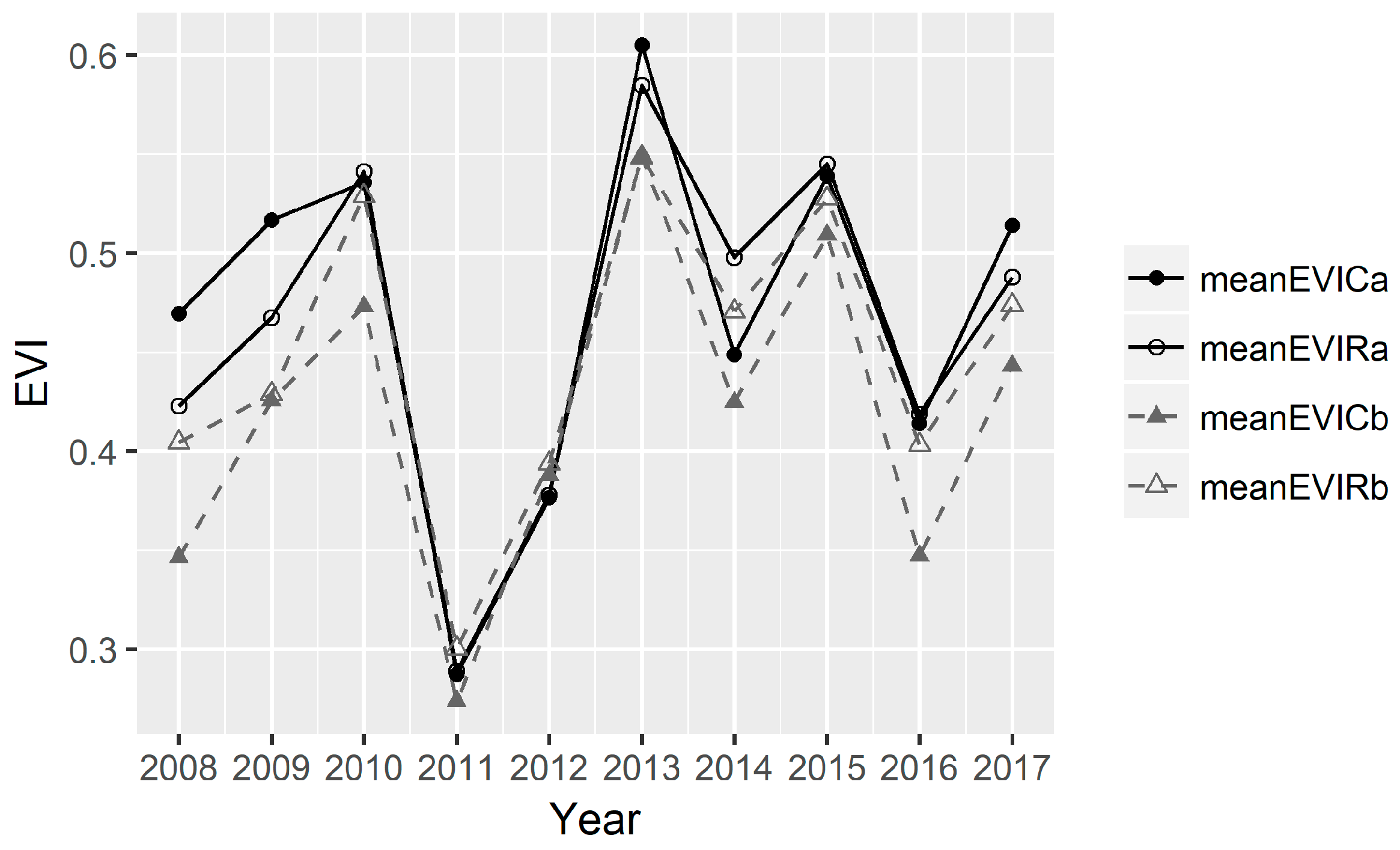
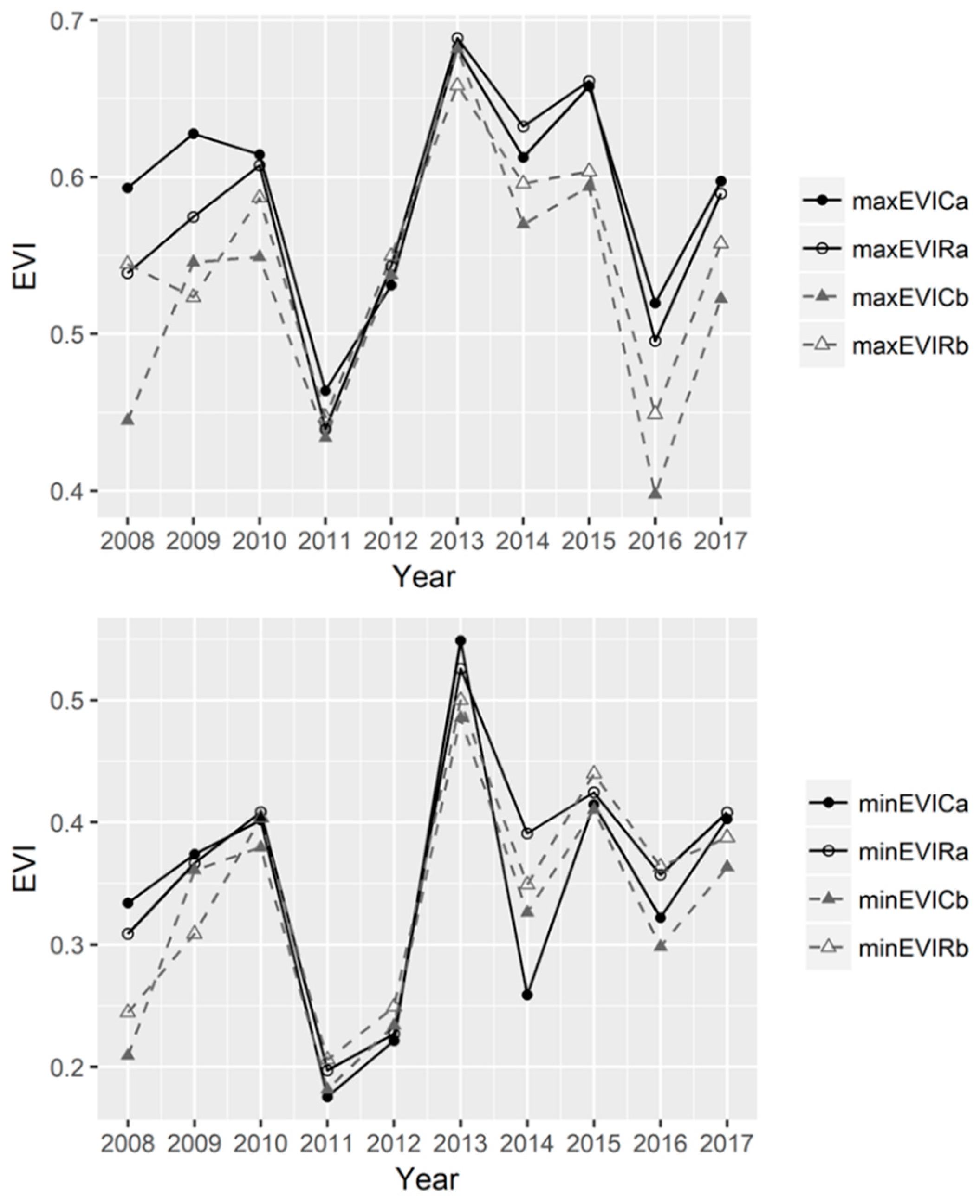
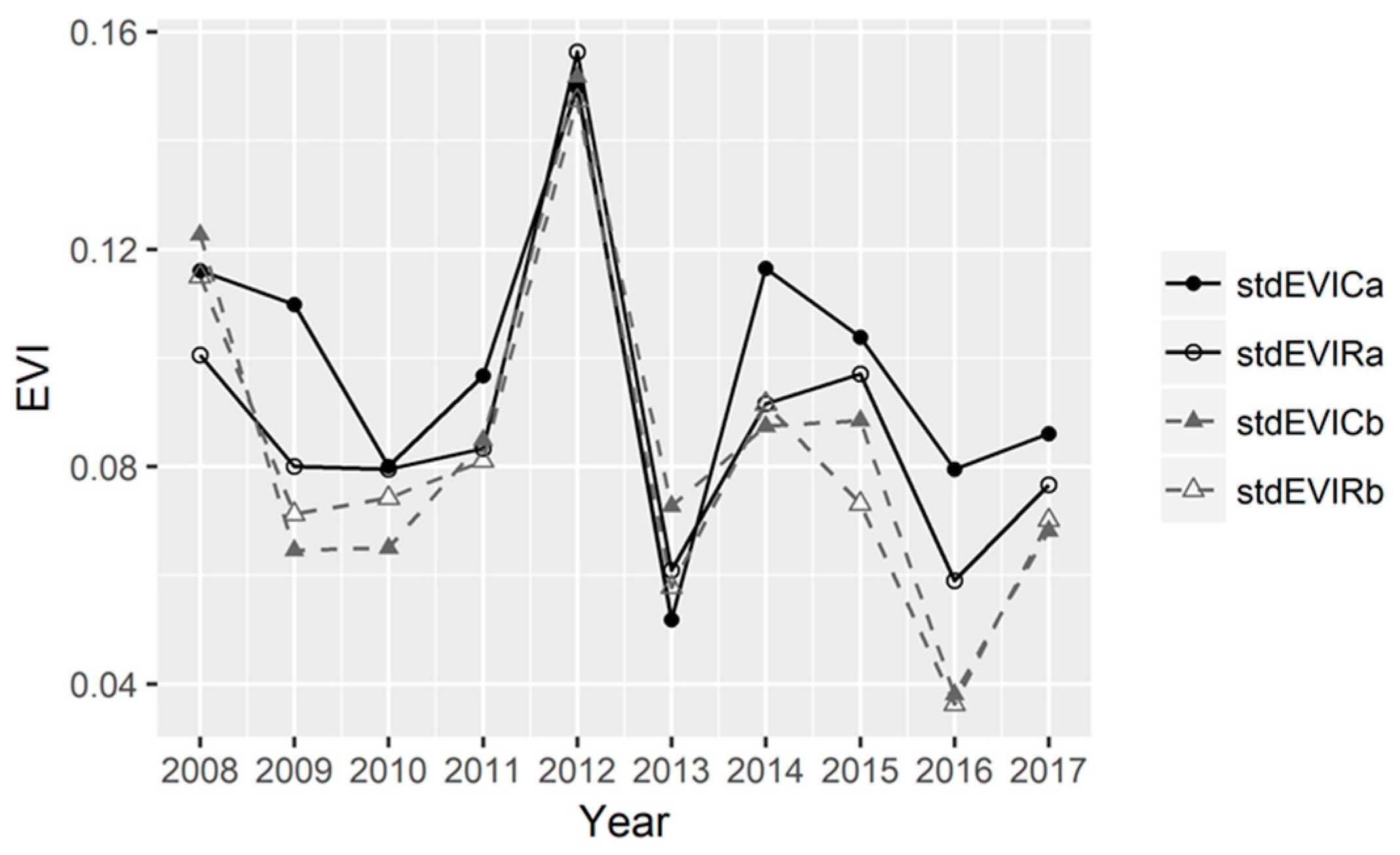
3.3. The Dynamics of EVI Values in the Continuous Versus Rotational Grazing Paddocks
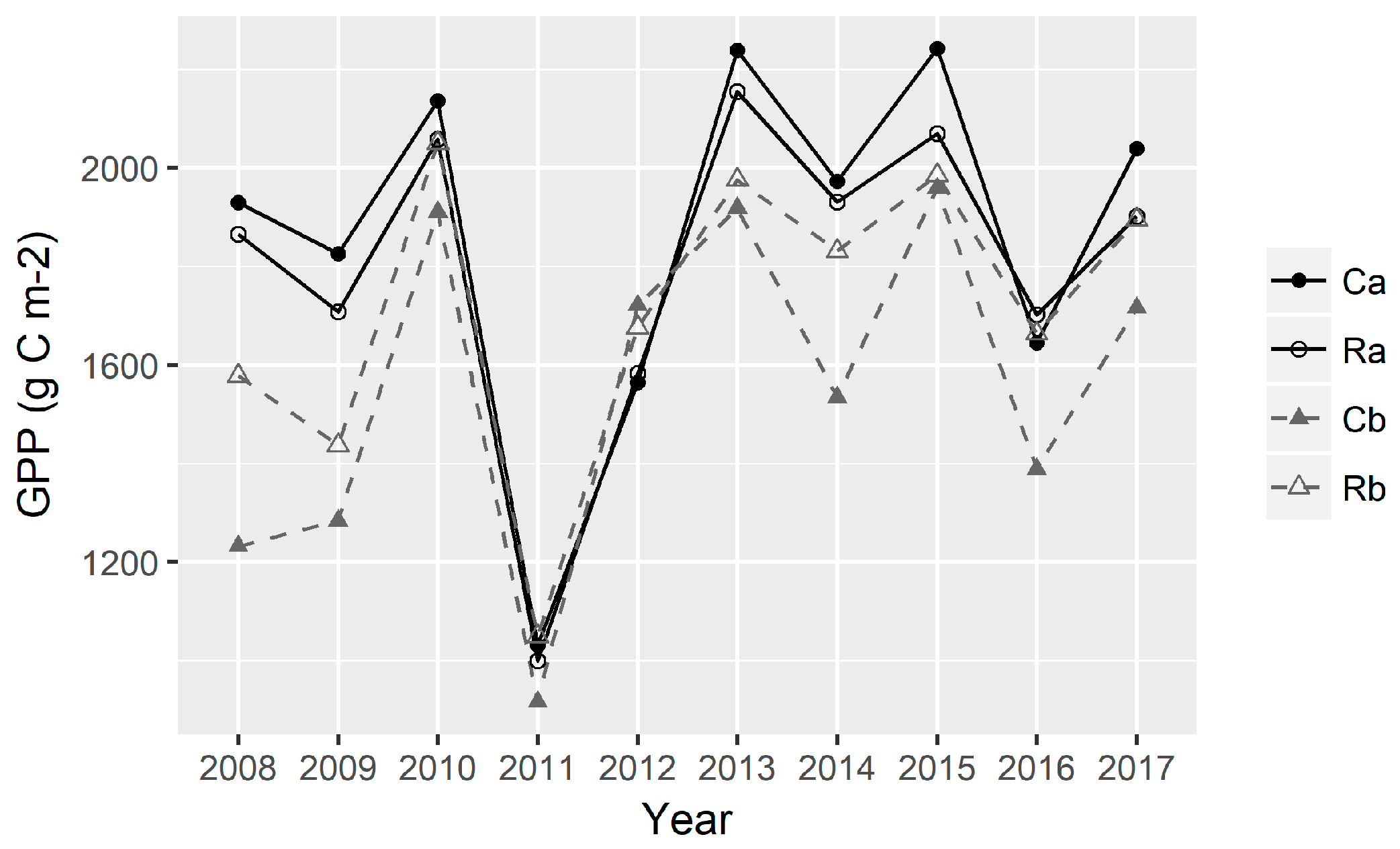
3.4. Dynamics of GPP in the Continuous Versus Rotational Grazing Paddocks
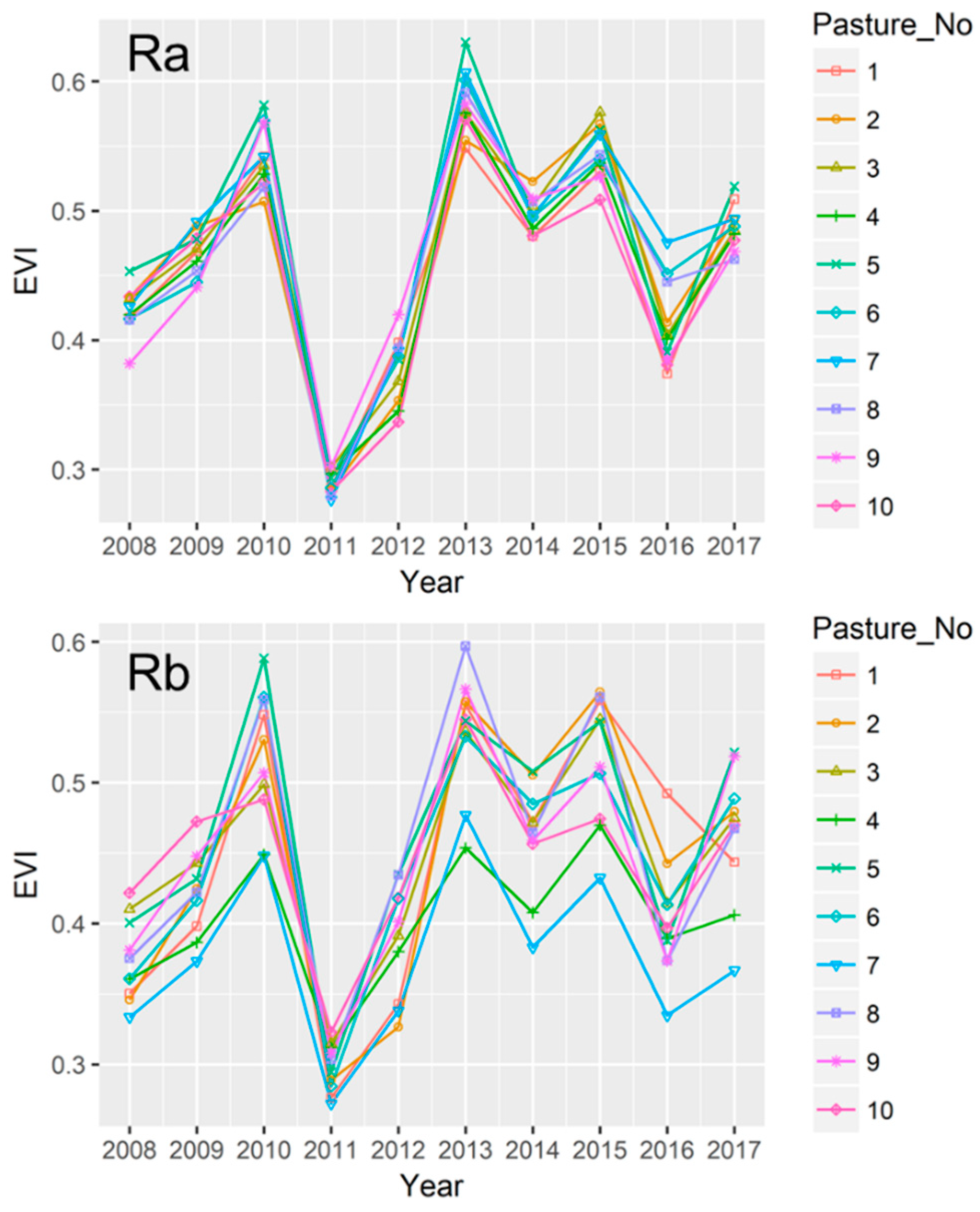
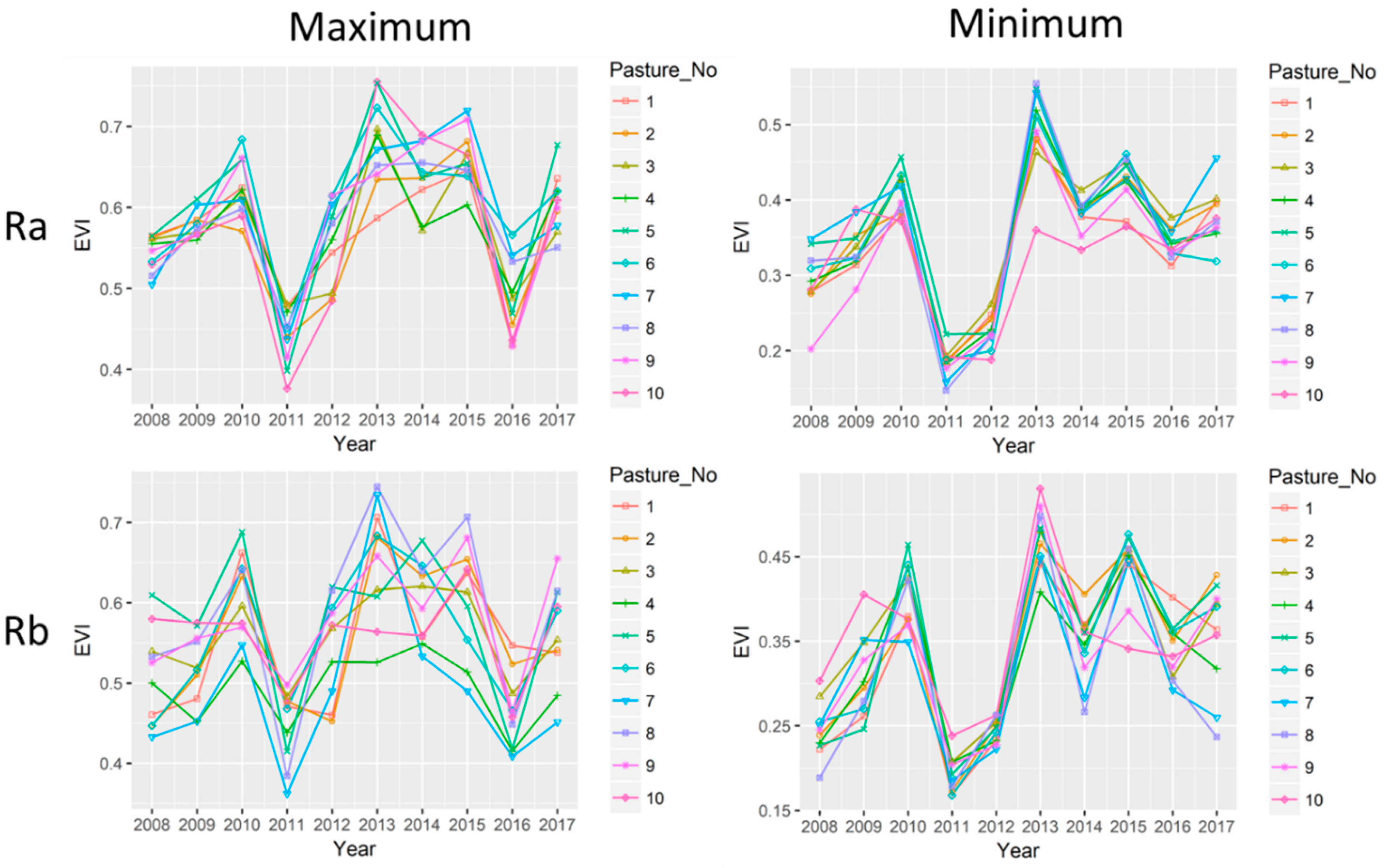
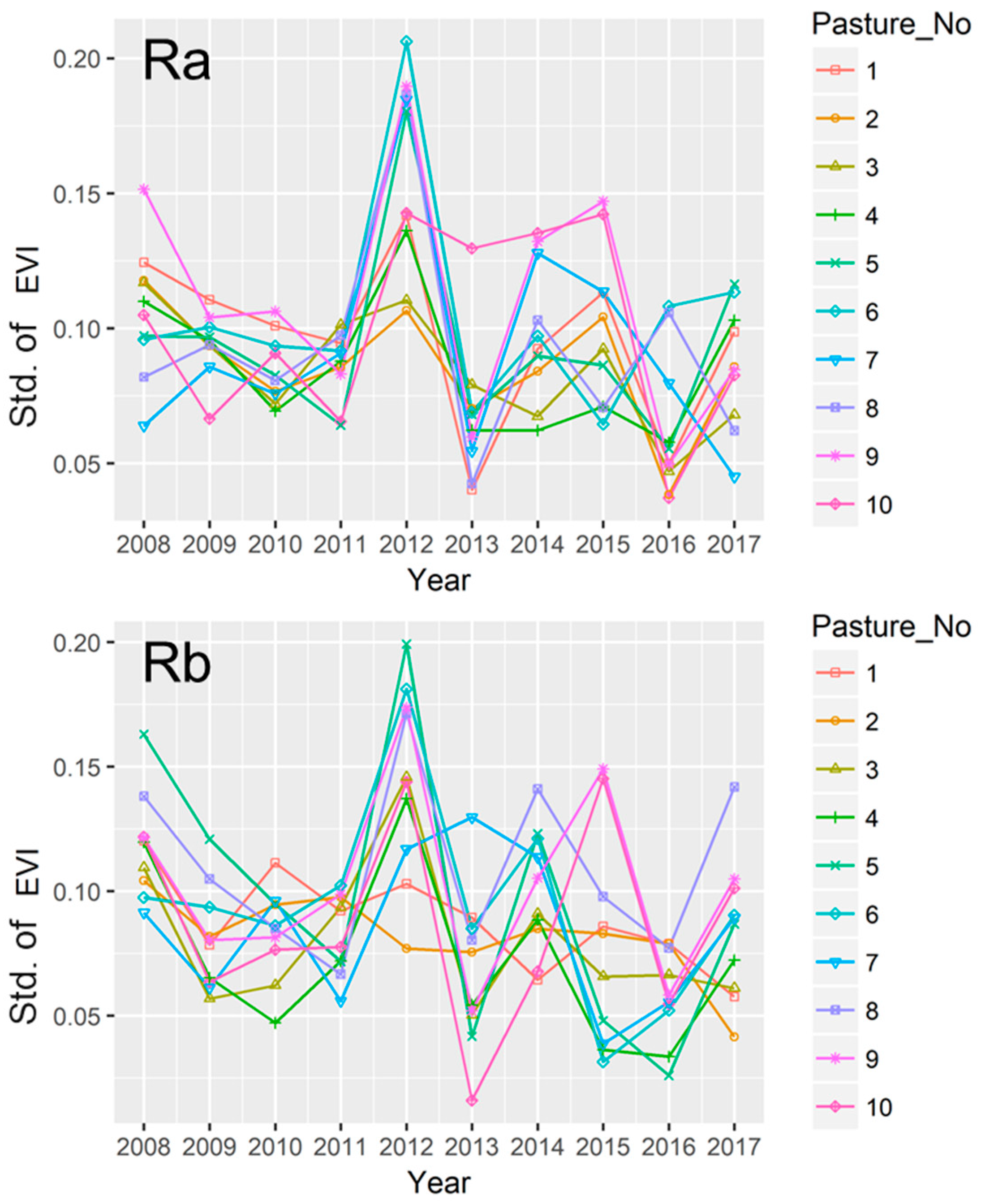
3.5. Variations in EVI among Paddocks in the Rotational Grazing Systems
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Samson, F.B.; Knopf, F.L.; Ostlie, W.R. Great Plains ecosystems: Past, present, and future. Wildl. Soc. Bull. 2004, 32, 6–15. [Google Scholar] [CrossRef]
- Cunfer, G. On the Great Plains: Agriculture and Environment; Texas A&M University Press: College Station, TX, USA, 2005. [Google Scholar]
- Westoby, M.; Walker, B.; Noy-Meir, I. Opportunistic management for rangelands not at equilibrium. J. Range Manag. 1989, 42, 266–274. [Google Scholar] [CrossRef]
- Fuhlendorf, S.D.; Engle, D.M. Restoring Heterogeneity on Rangelands: Ecosystem Management Based on Evolutionary Grazing Patterns: We propose a paradigm that enhances heterogeneity instead of homogeneity to promote biological diversity and wildlife habitat on rangelands grazed by livestock. BioScience 2001, 51, 625–632. [Google Scholar]
- Hubbard, W.A. Rotational grazing studies in western Canada. J. Range Manag. 1951, 4, 25–29. [Google Scholar] [CrossRef]
- Hyder, D.N.; Sawyer, W. Rotation-deferred grazing as compared to season-long grazing on sagebrush-bunchgrass ranges in Oregon. J. Range Manag. 1951, 4, 30–34. [Google Scholar] [CrossRef]
- McIlvain, E.; Savage, D. Eight-year comparisons of continuous and rotational grazing on the Southern Plains Experimental Range. J. Range Manag. 1951, 4, 42–47. [Google Scholar] [CrossRef]
- Rogler, G.A. A twenty-five year comparison of continuous and rotation grazing in the Northern Plains. J. Range Manag. 1951, 4, 35–41. [Google Scholar] [CrossRef]
- Sampson, A.W. A Symposium on Rotation Grazing in North America. J. Range Manag. 1951, 4, 19–24. [Google Scholar] [CrossRef]
- Sampson, A.W. Range Improvement by Deferred and Rotation Grazing; Series: Bulletin of the U.S. Department of Agriculture No. 34; U.S. Department of Agriculture: Washington, DC, USA, 1913.
- Leo, B.M. A Variation of Deferred Rotation Grazing for Use under Southwest Range Conditions. J. Range Manag. 1954, 7, 152–154. [Google Scholar]
- Heady, H.F. Continuous vs. specialized grazing systems: A review and application to the California annual type. J. Range Manag. 1961, 14, 182–193. [Google Scholar] [CrossRef]
- Bryant, F.C.; Dahl, B.E.; Pettit, R.D.; Britton, C.M. Does short-duration grazing work in arid and semiarid regions? J. Soil Water Conserv. 1989, 44, 290–296. [Google Scholar]
- White, M.R.; Pieper, R.D.; Donart, G.B.; Trifaro, L.W. Vegetational response to short-duration and continuous grazing in southcentral New Mexico. J. Range Manag. 1991, 44, 399–403. [Google Scholar] [CrossRef]
- Gillen, R.L.; Gillen, R.L.; McCollum III, F.T.; Tate, K.W.; Hodges, M.E. Tallgrass prairie response to grazing system and stocking rate. J. Range Manag. 1998, 51, 139–146. [Google Scholar] [CrossRef]
- Jacobo, E.J.; Rodríguez, A.M.; Bartoloni, N.; Deregibus, V.A. Rotational Grazing Effects on Rangeland Vegetation at a Farm Scale. Rangel. Ecol. Manag. 2006, 59, 249–257. [Google Scholar] [CrossRef]
- Briske, D.D.; Derner, J.D.; Brown, J.R.; Fuhlendorf, S.D.; Teague, W.R.; Havstad, K.M.; Gillen, R.L.; Ash, A.J.; Willms, W.D. Benefits of rotational grazing on rangelands: An evaluation of the experimental evidence. Rangel. Ecol. Manag. 2008, 61, 3–17. [Google Scholar] [CrossRef]
- Briske, D.D.; Derner, J.D.; Brown, J.R.; Fuhlendorf, S.D.; Teague, W.R.; Havstad, K.M.; Gillen, R.L.; Ash, A.J.; Willms, W.D. Rotational Grazing on Rangelands: Reconciliation of Perception and Experimental Evidence. Rangel. Ecol. Manag. 2008, 61, 3–17. [Google Scholar] [CrossRef]
- Budd, B.; Thorpe, J. Benefits of Managed Grazing: A Manager’s Perspective. Rangelands 2009, 31, 11–14. [Google Scholar] [CrossRef]
- Briske, D.D.; Sayre, N.F.; Huntsinger, L.; Fernández-Giménez, M.; Budd, B.; Derner, J.D. Origin, Persistence, and Resolution of the Rotational Grazing Debate: Integrating Human Dimensions into Rangeland Research. Rangel. Ecol. Manag. 2011, 64, 325–334. [Google Scholar] [CrossRef]
- Teague, R.; Provenza, F.; Kreuter, U.; Steffens, T.; Barnes, M. Multi-paddock grazing on rangelands: Why the perceptual dichotomy between research results and rancher experience? J. Environ. Manag. 2013, 128, 699–717. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Teague, W.R.; Park, S.C. Evaluation of Continuous and Multipaddock Grazing on Vegetation and Livestock Performance—A Modeling Approach. Rangel. Ecol. Manag. 2016, 69, 457–464. [Google Scholar] [CrossRef]
- Becker, W.; Kreuter, U.; Atkinson, S.; Teague, R. Whole-Ranch Unit Analysis of Multipaddock Grazing on Rangeland Sustainability in North Central Texas. Rangel. Ecol. Manag. 2017, 70, 448–455. [Google Scholar] [CrossRef]
- Norton, B.E.; Barnes, M.; Teague, R. Grazing Management Can Improve Livestock Distribution. Rangelands 2013, 35, 45–51. [Google Scholar] [CrossRef]
- Teague, W.; Dowhower, S.L.; Baker, S.A.; Haile, N.; DeLaune, P.B.; Conover, D.M. Grazing management impacts on vegetation, soil biota and soil chemical, physical and hydrological properties in tall grass prairie. Agric. Ecosyst. Environ. 2011, 141, 310–322. [Google Scholar] [CrossRef]
- Heitschmidt, R.; Dowhower, S.; Walker, J. Some effects of a rotational grazing treatment on quantity and quality of available forage and amount of ground litter. J. Range Manag. 1987, 40, 318–321. [Google Scholar] [CrossRef]
- Hart, R.H.; Samuel, M.J.; Test, P.S.; Smith, M.A. Cattle, vegetation, and economic responses to grazing systems and grazing pressure. J. Range Manag. 1988, 41, 282–286. [Google Scholar] [CrossRef]
- Hickman, K.R.; Hartnett, D.C.; Cochran, R.C.; Owensby, C.E. Grazing management effects on plant species diversity in tallgrass prairie. Rangel. Ecol. Manag. 2004, 57, 58–66. [Google Scholar] [CrossRef]
- Wood, M.K.; Blackburn, W.H. Vegetation and soil responses to cattle grazing systems in the Texas Rolling Plains. J. Range Manag. 1984, 37, 303–308. [Google Scholar] [CrossRef]
- Derner, J.D.; Hart, R.H. Grazing-induced modifications to peak standing crop in northern mixed-grass prairie. Rangel. Ecol. Manag. 2007, 60, 270–276. [Google Scholar]
- Derner, J.D.; Hart, R.H. Livestock and vegetation responses to rotational grazing in short-grass steppe. West. North Am. Nat. 2007, 67, 359–367. [Google Scholar] [CrossRef]
- Cassels, D.M.; Gillen, R.L.; Ted McCollum, F.; Tate, K.W.; Hodges, M.E. Effects of grazing management on standing crop dynamics in tallgrass prairie. J. Range Manag. 1995, 48, 81–84. [Google Scholar] [CrossRef]
- Martin, S.C.; Ward, D.E. Perennial grasses respond inconsistently to alternate year seasonal rest. Rangel. Ecol. Manag. J. Range Manag. Arch. 1976, 29, 346. [Google Scholar] [CrossRef]
- Reardon, P.O.; Merrill, L.B. Vegetative response under various grazing management systems in the Edwards Plateau of Texas. J. Range Manag. 1976, 29, 195–198. [Google Scholar] [CrossRef]
- Kawamura, K.; Akiyama, T.; Yokota, H.O.; Tsutsumi, M.; Yasuda, T.; Watanabe, O.; Wang, S. Quantifying grazing intensities using geographic information systems and satellite remote sensing in the Xilingol steppe region, Inner Mongolia, China. Agric. Ecosyst. Environ. 2005, 107, 83–93. [Google Scholar] [CrossRef]
- Numata, I.; Roberts, D.A.; Chadwick, O.A.; Schimel, J.; Sampaio, F.R.; Leonidas, F.C.; Soares, J.V. Characterization of pasture biophysical properties and the impact of grazing intensity using remotely sensed data. Remote Sens. Environ. 2007, 109, 314–327. [Google Scholar] [CrossRef]
- Ma, S.; Zhou, Y.; Gowda, P.; Chen, L.; Steiner, J.; Starks, P.; Neel, J. Evaluating the impacts of continuous and rotational grazing on tallgrass prairie landscape using high spatial resolution imagery. Agronomy 2019, in press. [Google Scholar]
- Starks, P.J.; Steiner, J.L.; Neel, J.P.S.; Turner, K.E.; Northup, B.K.; Brown, M.A.; Gowda, P.H. Assessment of the SPEI as a potential management tool for grasslands. Agronomy 2019, in press. [Google Scholar]
- Steiner, J.L.; Starks, P.J.; Neel, J.P.S.; Northup, B.K.; Gowda, P.H.; Brown, M.A.; Coleman, S. Managing Tallgrass Prairies for Productivity and Ecological Function: A Long Term Grazing Experiment in the Southern Great Plains, USA. Agronomy 2019, in press. [Google Scholar]
- Huete, A.; Didan, K.; Miura, T.; Rodriguez, E.P.; Gao, X.; Ferreira, L.G. Overview of the radiometric and biophysical performance of the MODIS vegetation indices. Remote Sens. Environ. 2002, 83, 195–213. [Google Scholar] [CrossRef]
- Boegh, E.; Søgaard, H.; Broge, N.; Hasager, C.B.; Jensen, N.O.; Schelde, K.; Thomsen, A. Airborne multispectral data for quantifying leaf area index, nitrogen concentration, and photosynthetic efficiency in agriculture. Remote Sens. Environ. 2002, 81, 179–193. [Google Scholar] [CrossRef]
- Xiao, X.; Hollinger, D.; Aber, J.; Goltz, M.; Davidson, E.A.; Zhang, Q.; Moore III, B. Satellite-based modeling of gross primary production in an evergreen needleleaf forest. Remote Sens. Environ. 2004, 89, 519–534. [Google Scholar] [CrossRef]
- Xiao, X.; Zhang, Q.; Braswell, B.; Urbanski, S.; Boles, S.; Wofsy, S.; Moore III, B.; Ojima, D. Modeling gross primary production of temperate deciduous broadleaf forest using satellite images and climate data. Remote Sens. Environ. 2004, 91, 256–270. [Google Scholar] [CrossRef]
- Wagle, P.; Xiao, X.; Torn, M.S.; Cook, D.R.; Matamala, R.; Fischer, M.L.; Jin, C.; Dong, J.; Biradar, C. Sensitivity of vegetation indices and gross primary production of tallgrass prairie to severe drought. Remote Sens. Environ. 2014, 152, 1–14. [Google Scholar] [CrossRef]
- Zhou, Y.; Xiao, X.; Wagle, P.; Bajgain, R.; Mahan, H.; Basara, J.B.; Dong, J.; Qin, Y.; Zhang, G.; Luo, Y. Examining the short-term impacts of diverse management practices on plant phenology and carbon fluxes of Old World bluestems pasture. Agric. For. Meteorol. 2017, 237–238, 60–70. [Google Scholar] [CrossRef]
- Bajgain, R.; Xiao, X.; Basara, J.; Wagle, P.; Zhou, Y.; Mahan, H.; Gowda, P.; McCarthy, H.R.; Northup, B.; Neel, L.; et al. Carbon dioxide and water vapor fluxes in winter wheat and tallgrass prairie in central Oklahoma. Sci. Total Environ. 2018, 644, 1511–1524. [Google Scholar] [CrossRef]
- Doughty, R.; Xiao, X.; Wu, X.; Zhang, Y.; Bajgain, R.; Zhou, Y.; Qin, Y.; Zou, Z.; McCarthy, H.R.; Friedman, J.; et al. Responses of gross primary production of grasslands and croplands under drought, pluvial, and irrigation conditions during 2010–2016, Oklahoma, USA. Agric. Water Manag. 2018, 204, 47–59. [Google Scholar] [CrossRef]
- Zhang, Y.; Xiao, X.; Wu, X.; Zhou, S.; Zhang, G.; Qin, Y.; Dong, J. A global moderate resolution dataset of gross primary production of vegetation for 2000–2016. Sci. Data 2017, 4, 170165. [Google Scholar] [CrossRef]
- Teague, W.; Dowhower, S.; Waggoner, J. Drought and grazing patch dynamics under different grazing management. J. Arid Environ. 2004, 58, 97–117. [Google Scholar] [CrossRef]
- Díaz-Solís, H.; Grant, W.E.; Kothmann, M.M.; Teague, W.R.; Díaz-García, J.A. Adaptive management of stocking rates to reduce effects of drought on cow–calf production systems in semi-arid rangelands. Agric. Syst. 2009, 100, 43–50. [Google Scholar] [CrossRef]
- Teague, W.R.; Provenza, F.; Norton, B.; Steffens, T.; Barnes, M.; Kothmann, M.; Roath, R. Benefits of multi-paddock grazing management on rangelands: Limitations of experimental grazing research and knowledge gaps. In Grasslands: Ecology, Management and Restoration; Schröder, H., Ed.; Nova Science Publishers: Hauppauge, NY, USA, 2009; pp. 41–80. [Google Scholar]
- Torell, L.A.; Murugan, S.; Ramirez, O.A. Economics of Flexible Versus Conservative Stocking Strategies to Manage Climate Variability Risk. Rangel. Ecol. Manag. 2010, 63, 415–425. [Google Scholar] [CrossRef]
- Roche, L.M.; Cutts, B.B.; Derner, J.D.; Lubell, M.N.; Tate, K.W. On-Ranch Grazing Strategies: Context for the Rotational Grazing Dilemma. Rangel. Ecol. Manag. 2015, 68, 248–256. [Google Scholar] [CrossRef]
- Otkin, J.A.; Anderson, M.C.; Hain, C.; Svoboda, M.; Johnson, D.; Mueller, R.; Tadesse, T.; Wardlow, B.; Brown, J. Assessing the evolution of soil moisture and vegetation conditions during the 2012 United States flash drought. Agric. For. Meteorol. 2016, 218, 230–242. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Xiao, X.; Zhang, G.; Wagle, P.; Bajgain, R.; Dong, J.; Basara, J.; Jin, C.; Anderson, M.C.; Hain, C.; et al. Quantifying agricultural drought in tallgrass prairie region in the U.S. Southern Great Plains through analysis of a water-related vegetation index from MODIS images. Agric. For. Meteorol. 2017, 246 (Suppl. C), 111–122. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plot Name | Area (ha) | Plot Name | Area (ha) |
|---|---|---|---|
| Ca | 58.6 | Cb | 62.7 |
| Ra-1 | 9.3 | Rb-1 | 5.1 |
| Ra-2 | 9.2 | Rb-2 | 7.5 |
| Ra-3 | 9.5 | Rb-3 | 12.9 |
| Ra-4 | 9.2 | Rb-4 | 12.2 |
| Ra-5 | 4.4 | Rb-5 | 9.0 |
| Ra-6 | 6.7 | Rb-6 | 6.7 |
| Ra-7 | 7.6 | Rb-7 | 9.1 |
| Ra-8 | 5.9 | Rb-8 | 6.6 |
| Ra-9 | 8.7 | Rb-9 | 7.0 |
| Ra-10 | 7.6 | Rb-10 | 6.7 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, Y.; Gowda, P.H.; Wagle, P.; Ma, S.; Neel, J.P.S.; Kakani, V.G.; Steiner, J.L. Climate Effects on Tallgrass Prairie Responses to Continuous and Rotational Grazing. Agronomy 2019, 9, 219. https://doi.org/10.3390/agronomy9050219
Zhou Y, Gowda PH, Wagle P, Ma S, Neel JPS, Kakani VG, Steiner JL. Climate Effects on Tallgrass Prairie Responses to Continuous and Rotational Grazing. Agronomy. 2019; 9(5):219. https://doi.org/10.3390/agronomy9050219
Chicago/Turabian StyleZhou, Yuting, Prasanna H. Gowda, Pradeep Wagle, Shengfang Ma, James P. S. Neel, Vijaya G. Kakani, and Jean L. Steiner. 2019. "Climate Effects on Tallgrass Prairie Responses to Continuous and Rotational Grazing" Agronomy 9, no. 5: 219. https://doi.org/10.3390/agronomy9050219