
Effects of Combined Application of Biological Agent and Fertilizer on Fungal Community Structure in Rhizosphere Soil of Panax notoginseng
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Site and Experimental Setup
2.2. Determination of Soil Physical and Chemical Properties
2.3. DNA Extraction and Amplification
2.4. Bioinformatic Analysis
3. Results
3.1. Physical and Chemical Properties of Rhizosphere Soil of P. notoginseng
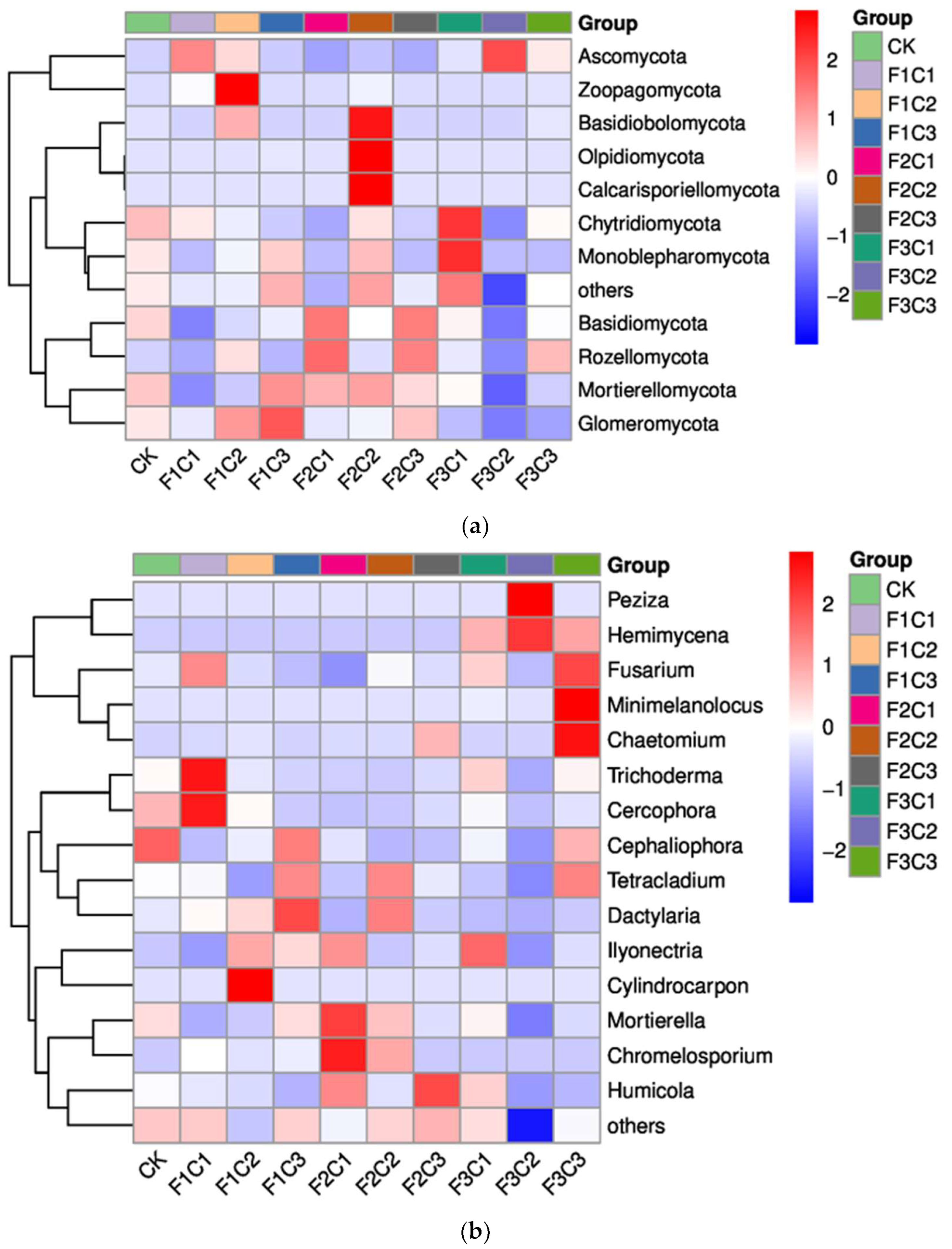
3.2. Diversity and Structural Characteristics of Fungi in Rhizosphere Soil of P. notoginseng
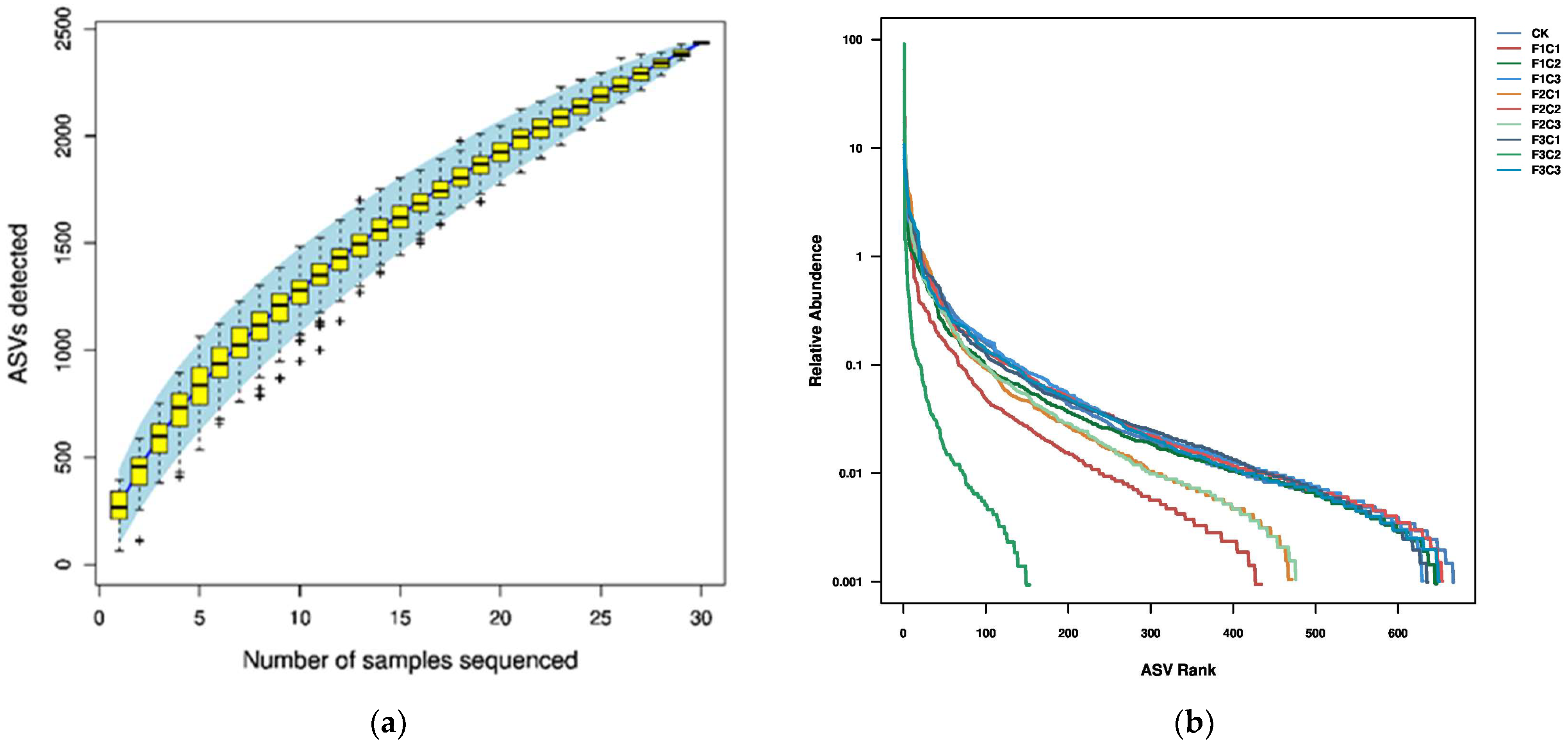
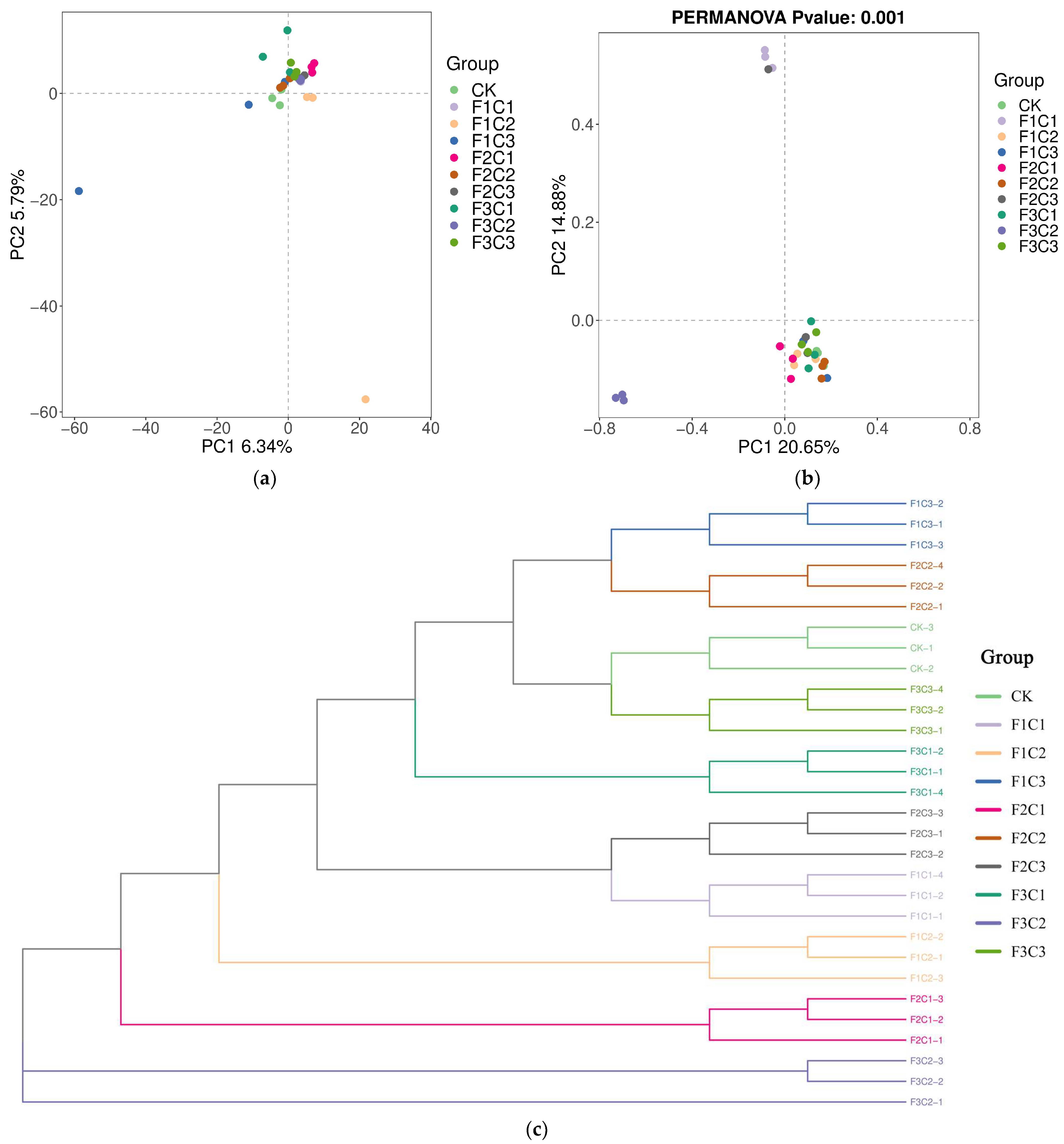
3.2.1. Diversity and Abundance
Analysis of the Alpha and Beta Diversity of Fungi Community
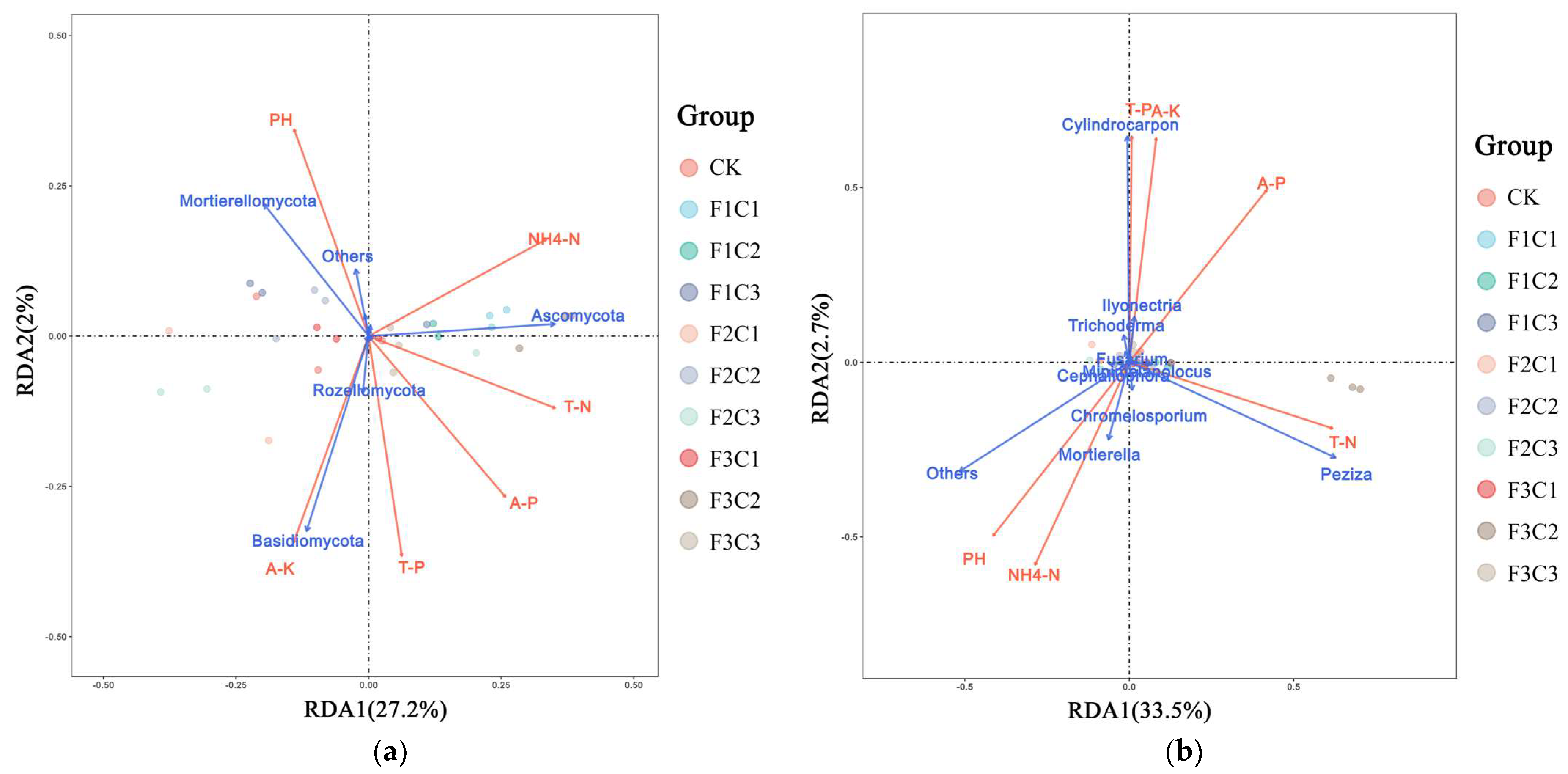
3.3. Relationship between Rhizosphere Soil Microorganisms and Environmental Physicochemical Parameters
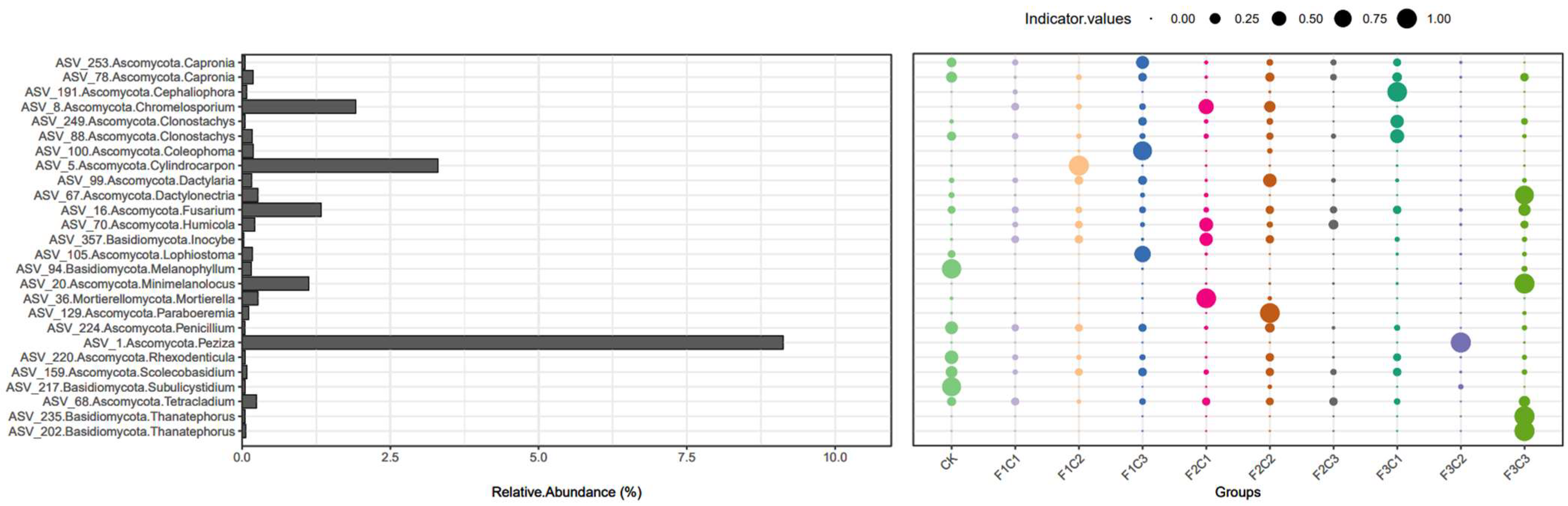
3.4. Indicator Analysis
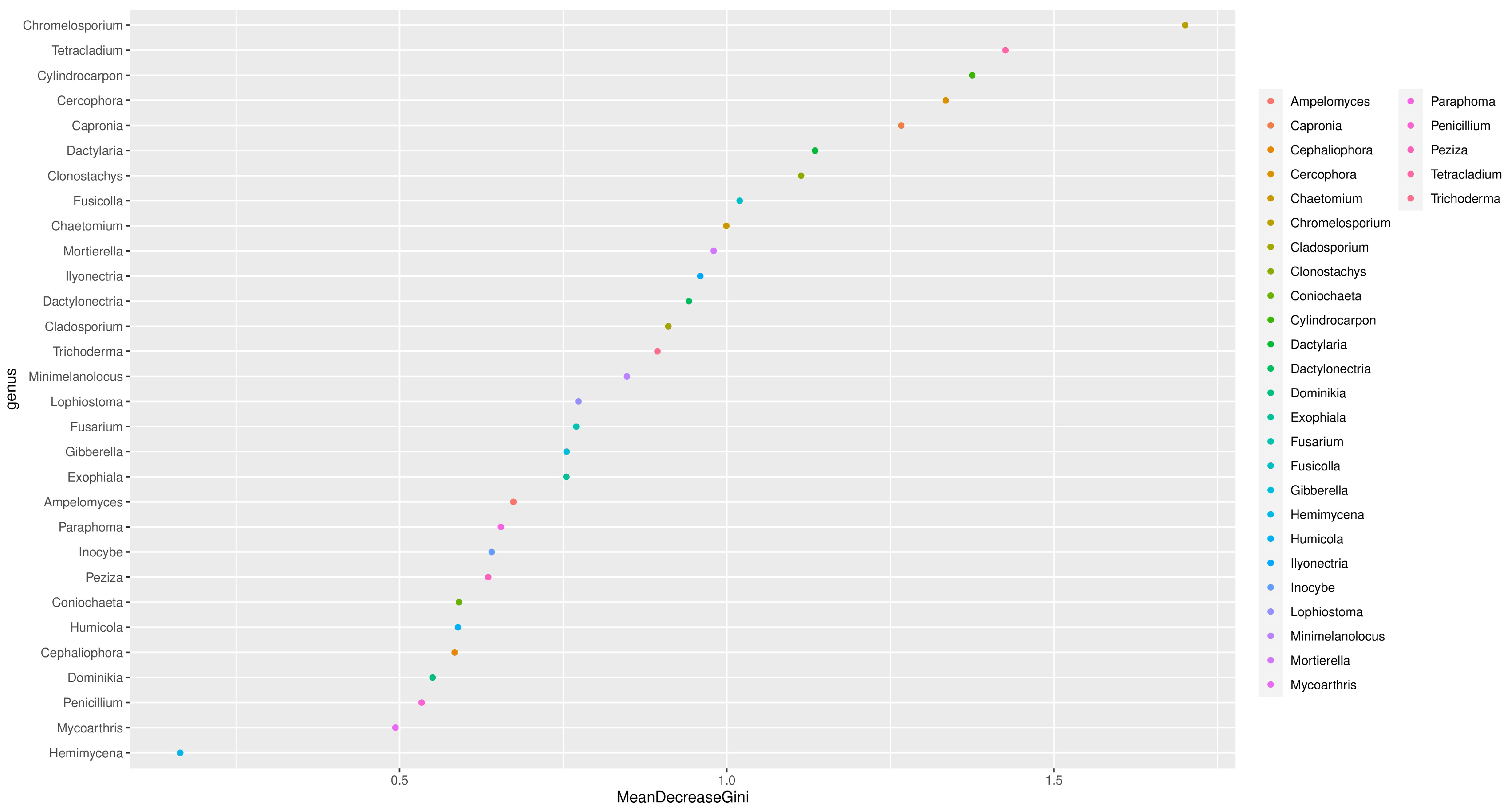
3.5. Random Forest Analysis
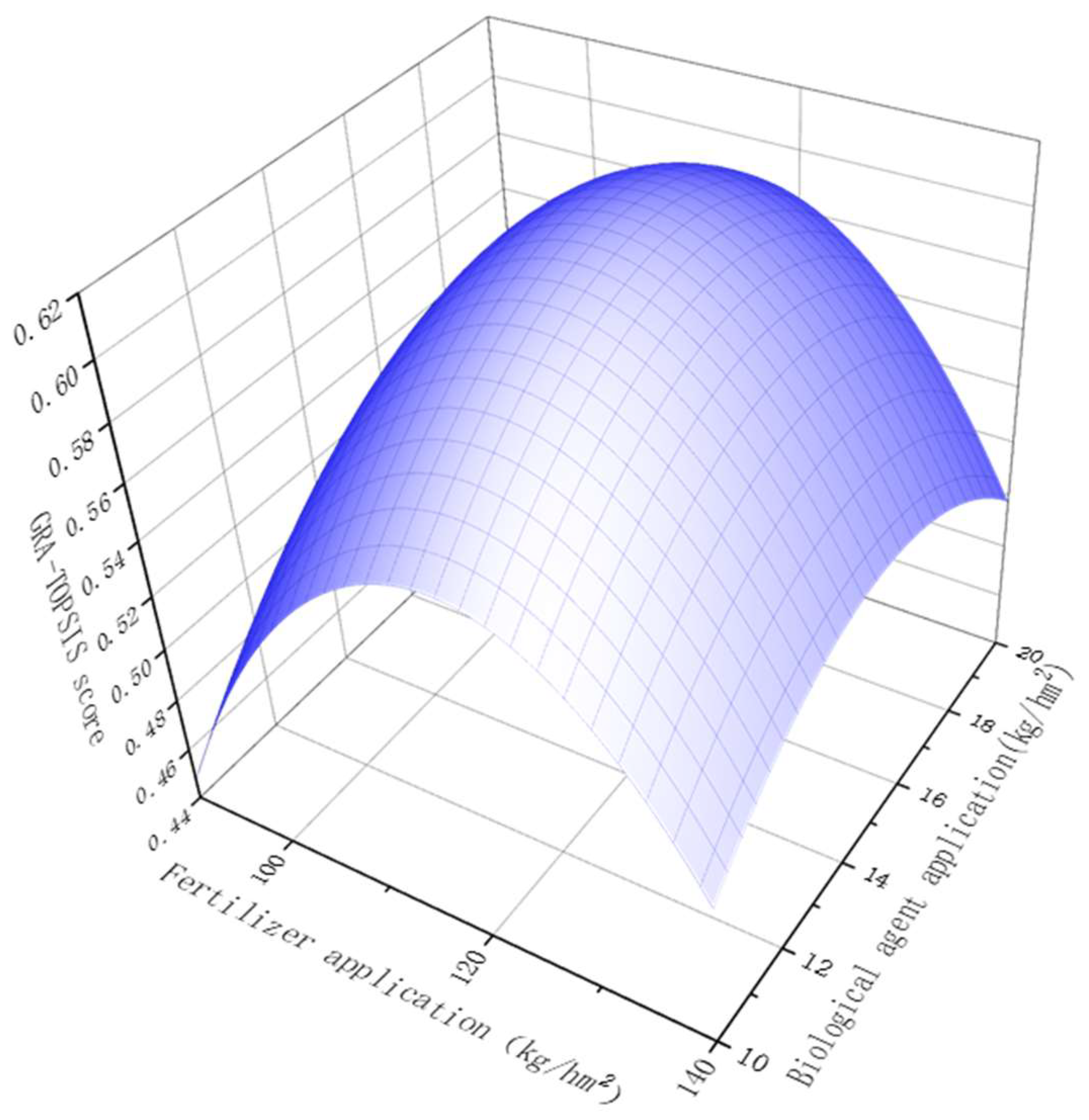
3.6. GRA-TOPSIS Analysis
3.7. Establish the Best Model in Theory
4. Discussion
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sun, H.X.; Qin, F.; Ye, Y.P. Relationship between haemolytic and adjuvant activity and structure of protopanaxadiol-type saponins from the roots of Panax notoginseng. Vaccine 2005, 23, 5533–5542. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Han, Z.; Li, G.; Zhang, S.; Luo, Y. Therapeutic Potential and Cellular Mechanisms of Panax notoginseng on Prevention of Aging and Cell Senescence-Associated Diseases. Aging Dis. 2017, 8, 721–739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, M.; Yuan, Y.; Huang, H.; Ye, C.; Guo, C.; Xu, Y.; Wang, W.; He, X.; Liu, Y.; Zhu, S. Steaming combined with biochar application eliminates negative plant-soil feedback for sanqi cultivation. Soil Tillage Res. 2019, 189, 189–198. [Google Scholar] [CrossRef]
- Guo, W.Q.; Chen, Y.G.; Shi, R.Z.; He, K.; Wang, J.F.; Shao, J.H.; Wan, J.B.; Gao, J.L. 20(S)-Protopanaxdiol Suppresses the Abnormal Granule-Monocyte Differentiation of Hematopoietic Stem Cells in 4T1 Breast Cancer-Bearing Mouse. Evid. Based Complement Altern. Med. 2020, 2020, 8747023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hawthorne, B.; Lund, K.; Freggiaro, S.; Kaga, R.; Meng, J. The mechanism of the cytotoxic effect of Panax notoginseng extracts on prostate cancer cells. Biomed. Pharmacother. 2022, 149, 10. [Google Scholar] [CrossRef]
- Santhakumar, A.B.; Battino, M.; Alvarez-Suarez, J.M. Dietary polyphenols: Structures, bioavailability and protective effects against atherosclerosis. Food Chem. Toxicol. 2018, 113, 49–65. [Google Scholar] [CrossRef]
- Xia, P.; Guo, H.; Zhao, H.; Jiao, J.; Deyholos, M.K.; Yan, X.; Liu, Y.; Liang, Z. Optimal fertilizer application for Panax notoginseng and effect of soil water on root rot disease and saponin contents. J. Ginseng. Res. 2016, 40, 38–46. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Ye, C.; Su, Y.; Peng, W.; Lu, R.; Liu, Y.; Huang, H.; He, X.; Yang, M.; Zhu, S. Soil Acidification caused by excessive application of nitrogen fertilizer aggravates soil-borne diseases: Evidence from literature review and field trials. Agric. Ecosyst. Environ. 2022, 340, 108176. [Google Scholar] [CrossRef]
- Fu, Y.; Dou, X.; Lu, Q.; Qin, J.; Luo, J.; Yang, M. Comprehensive assessment for the residual characteristics and degradation kinetics of pesticides in Panax notoginseng and planting soil. Sci. Total Environ. 2020, 714, 136718. [Google Scholar] [CrossRef]
- Jiang, N.; Qin, L.; Ye, Y. Research advances in diseases of Panax notoginseng. J. South. Argic. 2011, 42, 1070–1074. [Google Scholar]
- Li, J.B.; Bao, Y.L.; Wang, Z.R.; Yang, Q.; Cui, X.M. Research progress in diseases of Panax notoginseng. Physiol. Mol. Plant Pathol. 2022, 121, 8. [Google Scholar] [CrossRef]
- Tan, Y.; Cui, Y.; Li, H.; Kuang, A.; Li, X.; Wei, Y.; Ji, X. Rhizospheric soil and root endogenous fungal diversity and composition in response to continuous Panax notoginseng cropping practices. Microbiol. Res. 2017, 194, 10–19. [Google Scholar] [CrossRef]
- Ma, Y.N.; Chen, C.J.; Li, Q.Q.; Xu, F.R.; Cheng, Y.X.; Dong, X. Monitoring Antifungal Agents of Artemisia annua against Fusarium oxysporum and Fusarium solani, Associated with Panax notoginseng Root-Rot Disease. Molecules 2019, 24, 213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuoqing, M.; Shidong, L.I.; Xingzhong, L.I.U.; Yujun, C.; Yunhua, L.I.; Yong, W.; Rongjun, G.U.O.; Zhenyuan, X.I.A.; Keqin, Z. The Causal Microorganisms of Panax notoginseng Root Rot Disease. Sci. Agric. Sin. 2006, 39, 1371–1378. [Google Scholar]
- Wei, S.; Sun, Y.; Xi, G.; Zhang, H.; Xiao, M.; Yin, R. Development of a single-tube nested PCR-lateral flow biosensor assay for rapid and accurate detection of Alternaria panax Whetz. PLoS ONE 2018, 13, e0206462. [Google Scholar] [CrossRef]
- Sunera; Amna; Saqib, S.; Uddin, S.; Zaman, W.; Ullah, F.; Ayaz, A.; Asghar, M.; Rehman, S.U.; Munis, M.F.H.; et al. Characterization and phytostimulatory activity of bacteria isolated from tomato (Lycopersicon esculentum Mill.) rhizosphere. Microb Pathog 2020, 140, 103966. [Google Scholar] [CrossRef]
- Ou, X.; Cui, X.; Zhu, D.; Guo, L.; Liu, D.; Yang, Y. Lowering Nitrogen and Increasing Potassium Application Level Can Improve the Yield and Quality of Panax notoginseng. Front. Plant Sci. 2020, 11, 595095. [Google Scholar] [CrossRef]
- Tuo, Y.; Wang, Z.; Zheng, Y.; Shi, X.; Liu, X.; Ding, M.; Yang, Q. Effect of water and fertilizer regulation on the soil microbial biomass carbon and nitrogen, enzyme activity, and saponin content of Panax notoginseng. Agric. Water Manag. 2023, 278, 108145. [Google Scholar] [CrossRef]
- Li, J.; Yang, Q.; Shi, Z.; Zang, Z.; Liu, X. Effects of deficit irrigation and organic fertilizer on yield, saponin and disease incidence in Panax notoginseng under shaded conditions. Agric. Water Manag. 2021, 256, 107056. [Google Scholar] [CrossRef]
- Zhang, Y.; Liang, J.; Tang, Z.; Yang, Q. Rain Shelter Cultivation Reduces Root Rot Incidence of Panax notoginseng by Altering Root Exudates and Bacterial Communities under Micro-Irrigation and Fertilization. Agronomy 2023, 13, 1257. [Google Scholar] [CrossRef]
- Singh, B.K.; Millard, P.; Whiteley, A.S.; Murrell, J.C. Unravelling rhizosphere-microbial interactions: Opportunities and limitations. Trends Microbiol. 2004, 12, 386–393. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, W.; Wang, W. Effects of rhizosphere microbes and status of rihzosphere soil nutrients under different vegetations in south subtropical region. Soil Environ. Sci. 2002, 11, 279–282. [Google Scholar]
- Wu, Z.; Hao, Z.; Zeng, Y.; Guo, L.; Huang, L.; Chen, B. Molecular characterization of microbial communities in the rhizosphere soils and roots of diseased and healthy Panax notoginseng. Antonie Van Leeuwenhoek 2015, 108, 1059–1074. [Google Scholar] [CrossRef]
- Vacheron, J.; Desbrosses, G.; Bouffaud, M.L.; Touraine, B.; Moenne-Loccoz, Y.; Muller, D.; Legendre, L.; Wisniewski-Dye, F.; Prigent-Combaret, C. Plant growth-promoting rhizobacteria and root system functioning. Front. Plant Sci. 2013, 4, 356. [Google Scholar] [CrossRef] [Green Version]
- Chiappero, J.; Cappellari, L.d.R.; Sosa Alderete, L.G.; Palermo, T.B.; Banchio, E. Plant growth promoting rhizobacteria improve the antioxidant status in Mentha piperita grown under drought stress leading to an enhancement of plant growth and total phenolic content. Ind. Crops Prod. 2019, 139, 111553. [Google Scholar] [CrossRef]
- Cavagnaro, T.R.; Bender, S.F.; Asghari, H.R.; Heijden, M. The role of arbuscular mycorrhizas in reducing soil nutrient loss. Trends Plant Sci. 2015, 20, 283–290. [Google Scholar] [CrossRef] [Green Version]
- Bailey, K.L.; Lazarovits, G. Suppressing soil-borne diseases with residue management and organic amendments. Soil Tillage Res. 2003, 72, 169–180. [Google Scholar] [CrossRef]
- De Corato, U. Disease-suppressive compost enhances natural soil suppressiveness against soil-borne plant pathogens: A critical review. Rhizosphere 2020, 13, 100192. [Google Scholar] [CrossRef]
- Hashem, A.; Tabassum, B.; Fathi Abd Allah, E. Bacillus subtilis: A plant-growth promoting rhizobacterium that also impacts biotic stress. Saudi J. Biol. Sci. 2019, 26, 1291–1297. [Google Scholar] [CrossRef]
- Zhang, G.; Zhao, Z.; Yin, X.A.; Zhu, Y. Impacts of biochars on bacterial community shifts and biodegradation of antibiotics in an agricultural soil during short-term incubation. Sci. Total Environ. 2021, 771, 144751. [Google Scholar] [CrossRef]
- Qin, X.Y.; Zhang, K.D.; Fan, Y.Z.; Fang, H.; Nie, Y.; Wu, X.L. The Bacterial MtrAB Two-Component System Regulates the Cell Wall Homeostasis Responding to Environmental Alkaline Stress. Microbiol. Spectr. 2022, 15, e0231122. [Google Scholar] [CrossRef] [PubMed]
- Goswami, D.; Thakker, J.N.; Dhandhukia, P.C.; Tejada Moral, M. Portraying mechanics of plant growth promoting rhizobacteria (PGPR): A review. Cogent Food Agric. 2016, 2, 1127500. [Google Scholar] [CrossRef]
- Tripathi, D.K.; Singh, V.P.; Kumar, D.; Chauhan, D.K. Impact of exogenous silicon addition on chromium uptake, growth, mineral elements, oxidative stress, antioxidant capacity, and leaf and root structures in rice seedlings exposed to hexavalent chromium. Acta Physiol. Plant. 2011, 34, 279–289. [Google Scholar] [CrossRef]
- Emmert, E.A.B.; Handelsman, J. Biocontrol of plant disease: A (Gram-) positive perspective. FEMS Microbiol. Lett. 1999, 171, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Stein, T. Bacillus subtilis antibiotics: Structures, syntheses and specific functions. Mol. Microbiol. 2005, 56, 845–857. [Google Scholar] [CrossRef]
- Caulier, S.; Nannan, C.; Gillis, A.; Licciardi, F.; Bragard, C.; Mahillon, J. Overview of the Antimicrobial Compounds Produced by Members of the Bacillus subtilis Group. Front. Microbiol. 2019, 10, 19. [Google Scholar] [CrossRef] [Green Version]
- Radhakrishnan, R.; Hashem, A.; Abd Allah, E.F. Bacillus: A Biological Tool for Crop Improvement through Bio-Molecular Changes in Adverse Environments. Front. Physiol. 2017, 8, 667. [Google Scholar] [CrossRef] [Green Version]
- Sun, B.; Bai, Z.; Bao, L.; Xue, L.; Zhang, S.; Wei, Y.; Zhang, Z.; Zhuang, G.; Zhuang, X. Bacillus subtilis biofertilizer mitigating agricultural ammonia emission and shifting soil nitrogen cycling microbiomes. Environ. Int. 2020, 144, 105989. [Google Scholar] [CrossRef]
- Gadhave, K.R.; Devlin, P.F.; Ebertz, A.; Ross, A.; Gange, A.C. Soil Inoculation with Bacillus spp. Modifies Root Endophytic Bacterial Diversity, Evenness, and Community Composition in a Context-Specific Manner. Microb. Ecol. 2018, 76, 741–750. [Google Scholar] [CrossRef] [Green Version]
- Chowdappa, P.; Mohan Kumar, S.P.; Jyothi Lakshmi, M.; Upreti, K.K. Growth stimulation and induction of systemic resistance in tomato against early and late blight by Bacillus subtilis OTPB1 or Trichoderma harzianum OTPB3. Biol. Control 2013, 65, 109–117. [Google Scholar] [CrossRef]
- Xueling, Z.; Chunhua, Z.; Ruyue, G.; Yichao, D.; Yangang, W.; Zhongxin, G.; Zhenzhong, Z.; Xueling, R. Effects of Biological Fertilizer on Rhizosphere and Endophytic Bacterial Community of Panax notoginseng with Continuous Cropping Disorder. J. Henan Agric. Sci. 2021, 50, 78–91. [Google Scholar] [CrossRef]
- Tang, J.; Han, H.; Liu, B.; Yang, Q.; Liu, X.; Liu, Y. Effects of irrigation frequency and fertilization amount on active ingredient accumulation and morbidity of Panax notoginseng. Trans. Chin. Soc. Agric. Eng. 2020, 36, 55–63. [Google Scholar]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Rodrigues, J.; Piedade, S.M.D.S.; Lara, I.A.R.d.; Henrique, F.H. Type I error in multiple comparison tests in analysis of variance. Acta Scientiarum. Agron. 2022, 45, e57742. [Google Scholar] [CrossRef]
- Jia, M.; Sun, X.; Chen, M.; Liu, S.; Zhou, J.; Peng, X. Deciphering the microbial diversity associated with healthy and wilted Paeonia suffruticosa rhizosphere soil. Front. Microbiol. 2022, 13, 967601. [Google Scholar] [CrossRef]
- Xiong, W.; Zhao, Q.; Zhao, J.; Xun, W.; Li, R.; Zhang, R.; Wu, H.; Shen, Q. Different continuous cropping spans significantly affect microbial community membership and structure in a vanilla-grown soil as revealed by deep pyrosequencing. Microb. Ecol. 2015, 70, 209–218. [Google Scholar] [CrossRef]
- Wang, C.; Lu, X.; Mori, T.; Mao, Q.; Zhou, K.; Zhou, G.; Nie, Y.; Mo, J. Responses of soil microbial community to continuous experimental nitrogen additions for 13 years in a nitrogen-rich tropical forest. Soil Biol. Biochem. 2018, 121, 103–112. [Google Scholar] [CrossRef]
- Chen, M.; Liu, H.; Yu, S.; Wang, M.; Pan, L.; Chen, N.; Wang, T.; Chi, X.; Du, B. Long-term continuously monocropped peanut significantly changed the abundance and composition of soil bacterial communities. PeerJ 2020, 8, e9024. [Google Scholar] [CrossRef]
- Chen, M.; Zhang, J.; Liu, H.; Wang, M.; Pan, L.; Chen, N.; Wang, T.; Jing, Y.; Chi, X.; Du, B. Long-term continuously monocropped peanut significantly disturbed the balance of soil fungal communities. J. Microbiol. 2020, 58, 563–573. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, Y.; Li, Z.-Z.; Zhao, Q.; Huang, X.-Y.; Huang, K.-F. Effect of continuous cropping on the rhizosphere soil and growth of common buckwheat. Plant Prod. Sci. 2019, 23, 81–90. [Google Scholar] [CrossRef] [Green Version]
- Mazzola, M.; Manici, L.M. Apple replant disease: Role of microbial ecology in cause and control. Annu. Rev. Phytopathol. 2012, 50, 45–65. [Google Scholar] [CrossRef] [PubMed]
- Yim, B.; Smalla, K.; Winkelmann, T. Evaluation of apple replant problems based on different soil disinfection treatments—Links to soil microbial community structure? Plant Soil 2012, 366, 617–631. [Google Scholar] [CrossRef]
- Weiss, S.; Bartsch, M.; Winkelmann, T. Transcriptomic analysis of molecular responses in Malus domestica ‘M26′ roots affected by apple replant disease. Plant Mol. Biol. 2017, 94, 303–318. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Wang, G.; Kang, X.; Hu, H.; Wang, Y.; Zhang, X.; Sun, X.; Zhang, H.; Hu, Z.; Xi, B. Long-term fertilization alters microbial community but fails to reclaim soil organic carbon stocks in a land-use changed soil of the Tibetan Plateau. Land Degrad. Dev. 2019, 31, 531–542. [Google Scholar] [CrossRef]
- Ren, N.; Wang, Y.; Ye, Y.; Zhao, Y.; Huang, Y.; Fu, W.; Chu, X. Effects of Continuous Nitrogen Fertilizer Application on the Diversity and Composition of Rhizosphere Soil Bacteria. Front. Microbiol. 2020, 11, 1948. [Google Scholar] [CrossRef]
- Francioli, D.; Schulz, E.; Lentendu, G.; Wubet, T.; Buscot, F.; Reitz, T. Mineral vs. Organic Amendments: Microbial Community Structure, Activity and Abundance of Agriculturally Relevant Microbes Are Driven by Long-Term Fertilization Strategies. Front. Microbiol. 2016, 7, 1446. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.; Ye, G.; Kuzyakov, Y.; Liu, D.; Fan, J.; Ding, W. Long-term manure application increases soil organic matter and aggregation, and alters microbial community structure and keystone taxa. Soil Biol. Biochem. 2019, 134, 187–196. [Google Scholar] [CrossRef]
- Dou, Y.; Lu, K.; Deng, P.; Zhang, F.; Zhang, X.; Luo, W. Study on the nitrogenous transformation in weak alkaline soil amended with different rates of nitrite. Appl. Chem. Ind. 2019, 48, 1830–1832, 1836. [Google Scholar]
- Chen, B.; Liu, E.; Tian, Q.; Yan, C.; Zhang, Y. Soil nitrogen dynamics and crop residues. A review. Agron. Sustain. Dev. 2014, 34, 429–442. [Google Scholar] [CrossRef] [Green Version]
- Oladele, S.O.; Adeyemo, A.J.; Awodun, M.A. Influence of rice husk biochar and inorganic fertilizer on soil nutrients availability and rain-fed rice yield in two contrasting soils. Geoderma 2019, 336, 1–11. [Google Scholar] [CrossRef]
- Vejan, P.; Abdullah, R.; Khadiran, T.; Ismail, S.; Nasrulhaq Boyce, A. Role of Plant Growth Promoting Rhizobacteria in Agricultural Sustainability—A Review. Molecules 2016, 21, 573. [Google Scholar] [CrossRef] [Green Version]
- Ali, M.A.; Ren, H.; Ahmed, T.; Luo, J.; An, Q.; Qi, X.; Li, B. Antifungal Effects of Rhizospheric Bacillus Species against Bayberry Twig Blight Pathogen Pestalotiopsis versicolor. Agronomy 2020, 10, 1811. [Google Scholar] [CrossRef]
- You, C.; Zhang, C.; Kong, F.; Feng, C.; Wang, J. Comparison of the effects of biocontrol agent Bacillus subtilis and fungicide metalaxyl–mancozeb on bacterial communities in tobacco rhizospheric soil. Ecol. Eng. 2016, 91, 119–125. [Google Scholar] [CrossRef]
- Gajbhiye, A.; Rai, A.R.; Meshram, S.U.; Dongre, A.B. Isolation, evaluation and characterization of Bacillus subtilis from cotton rhizospheric soil with biocontrol activity against Fusarium oxysporum. World J. Microbiol. Biotechnol. 2010, 26, 1187–1194. [Google Scholar] [CrossRef]
- Rahman, M.M.E.; Hossain, D.M.; Suzuki, K.; Shiiya, A.; Suzuki, K.; Dey, T.K.; Nonaka, M.; Harada, N. Suppressive effects of Bacillus spp. on mycelia, apothecia and sclerotia formation of Sclerotinia sclerotiorum and potential as biological control of white mold on mustard. Australas. Plant Pathol. 2016, 45, 103–117. [Google Scholar] [CrossRef]
- Sui, J.; Yu, Q.; Yang, K.; Yang, J.; Li, C.; Liu, X. Effects of Bacillus subtilis T6-1 on the Rhizosphere Microbial Community Structure of Continuous Cropping Poplar. Biology 2022, 11, 791. [Google Scholar] [CrossRef]
- Zhao, W.; Guo, Q.; Li, S.; Wang, P.; Dong, L.; Su, Z.; Zhang, X.; Lu, X.; Ma, P. Effects of Bacillus subtilis NCD-2 and broccoli residues return on potato Verticillium wilt and soil fungal community structure. Biol. Control 2021, 159, 104628. [Google Scholar] [CrossRef]
- Miao, C.P.; Mi, Q.L.; Qiao, X.G.; Zheng, Y.K.; Chen, Y.W.; Xu, L.H.; Guan, H.L.; Zhao, L.X. Rhizospheric fungi of Panax notoginseng: Diversity and antagonism to host phytopathogens. J. Ginseng. Res. 2016, 40, 127–134. [Google Scholar] [CrossRef] [Green Version]
- Cabral, A.; Groenewald, J.Z.; Rego, C.; Oliveira, H.; Crous, P.W. Cylindrocarpon root rot: Multi-gene analysis reveals novel species within the Ilyonectria radicicola species complex. Mycol. Prog. 2011, 11, 655–688. [Google Scholar] [CrossRef] [Green Version]
- Lekberg, Y.; Arnillas, C.A.; Borer, E.T.; Bullington, L.S.; Fierer, N.; Kennedy, P.G.; Leff, J.W.; Luis, A.D.; Seabloom, E.W.; Henning, J.A. Nitrogen and phosphorus fertilization consistently favor pathogenic over mutualistic fungi in grassland soils. Nat. Commun. 2021, 12, 3484. [Google Scholar] [CrossRef]
- Wu, F.; Cui, X.; Yang, Y.; Guan, H. Effects of soil pH values regulated by different fertilization on the disease incidence and growth of Panax notoginseng. J. Yunnan Univ. Nat. Sci. 2017, 39, 908–914. [Google Scholar]
- Kamble, P.N.; Gaikwad, V.B.; Kuchekar, S.R.; Bååth, E. Microbial growth, biomass, community structure and nutrient limitation in high pH and salinity soils from Pravaranagar (India). Eur. J. Soil Biol. 2014, 65, 87–95. [Google Scholar] [CrossRef]
- Rousk, J.; Brookes, P.C.; Bååth, E. Investigating the mechanisms for the opposing pH relationships of fungal and bacterial growth in soil. Soil Biol. Biochem. 2010, 42, 926–934. [Google Scholar] [CrossRef]
- Rousk, J.; Baath, E.; Brookes, P.C.; Lauber, C.L.; Lozupone, C.; Caporaso, J.G.; Knight, R.; Fierer, N. Soil bacterial and fungal communities across a pH gradient in an arable soil. ISME J. 2010, 4, 1340–1351. [Google Scholar] [CrossRef] [PubMed]
- Compant, S.; Brader, G.; Muzammil, S.; Sessitsch, A.; Lebrihi, A.; Mathieu, F. Use of beneficial bacteria and their secondary metabolites to control grapevine pathogen diseases. BioControl 2012, 58, 435–455. [Google Scholar] [CrossRef] [Green Version]
- Mousavi Khaneghah, A.; Abhari, K.; Eş, I.; Soares, M.B.; Oliveira, R.B.A.; Hosseini, H.; Rezaei, M.; Balthazar, C.F.; Silva, R.; Cruz, A.G.; et al. Interactions between probiotics and pathogenic microorganisms in hosts and foods: A review. Trends Food Sci. Technol. 2020, 95, 205–218. [Google Scholar] [CrossRef]
- Chen, Y.; Du, J.; Li, Y.; Tang, H.; Yin, Z.; Yang, L.; Ding, X. Evolutions and Managements of Soil Microbial Community Structure Drove by Continuous Cropping. Front. Microbiol. 2022, 13, 839494. [Google Scholar] [CrossRef]
- Wolińska, A.; Kuźniar, A.; Zielenkiewicz, U.; Banach, A.; Błaszczyk, M. Indicators of arable soils fatigue — Bacterial families and genera: A metagenomic approach. Ecol. Indic. 2018, 93, 490–500. [Google Scholar] [CrossRef]
- Pervaiz, Z.H.; Iqbal, J.; Zhang, Q.; Chen, D.; Wei, H.; Saleem, M. Continuous Cropping Alters Multiple Biotic and Abiotic Indicators of Soil Health. Soil Syst. 2020, 4, 59. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Fertilizer (kg·hm−2) | Biological Agents (kg·hm−2) |
|---|---|---|
| CK | 0 | 0 |
| F1C1 | 80 | 10 |
| F1C2 | 80 | 10 |
| F1C3 | 80 | 10 |
| F2C1 | 110 | 15 |
| F2C2 | 110 | 15 |
| F2C3 | 110 | 15 |
| F3C1 | 140 | 20 |
| F3C2 | 140 | 20 |
| F3C3 | 140 | 20 |
| Treatments | PH | NH4-N(mg·kg−1) | AP (mg·kg−1) | AK (mg·kg−1) | TN (mg·kg−1) | TP (mg·kg−1) | Yield (g) |
|---|---|---|---|---|---|---|---|
| CK | 7.55 ± 0.06 a | 38.44 ± 0.94 a | 65.66 ± 4.25 d | 24.76 ± 1.25 a | 368.90 ± 10.27 a | 335.15 ± 7.56 c | 10.91 ± 0.72 a |
| F1C1 | 7.38 ± 0.03 a | 34.19 ± 0.50 b | 71.34 ± 4.36 cd | 26.10 ± 2.28 a | 369.27 ± 6.97 a | 448.72 ± 32.69 ab | 11.09 ± 0.48 a |
| F1C2 | 7.41 ± 0.02 a | 34.19 ± 1.20 b | 73.64 ± 8.82 cd | 26.4 ± 2.4 a | 373.94 ± 7.32 a | 396.32 ± 11.07 bc | 11.49 ± 0.29 a |
| F1C3 | 7.51 ± 0.01 a | 25.22 ± 0.52 c | 69.62 ± 10.23 cd | 25.50 ± 1.96 a | 369.83 ± 8.75 a | 449.36 ± 20.03 ab | 11.90 ± 0.51 a |
| F2C1 | 7.33 ± 0.01 a | 25.41 ± 0.45 c | 97.98 ± 8.04 abcd | 30.48 ± 2.49 a | 377.58 ± 11.52 a | 445.51 ± 14.69 ab | 12.55 ± 1.04 a |
| F2C2 | 7.45 ± 0.02 a | 25.55 ± 0.29 c | 99.42 ± 6.16 abc | 30.44 ± 2.09 a | 389.94 ± 11.25 a | 430.32 ± 14.77 ab | 11.99 ± 1.07 a |
| F2C3 | 7.36 ± 0.06 a | 24.44 ± 1.69 c | 91.50 ± 12.82 bcd | 30.42 ± 1.18 a | 379.77 ± 14.9 a | 455.99 ± 20.19 ab | 11.66 ± 1.51 a |
| F3C1 | 7.40 ± 0.02 a | 26.04 ± 0.12 c | 120.6 ± 8.99 ab | 30.60 ± 0.76 a | 399.46 ± 12.07 a | 495.13 ± 27.46 a | 11.71 ± 0.42 a |
| F3C2 | 7.32 ± 0.03 b | 27.08 ± 0.46 c | 126.37 ± 7.54 a | 31.05 ± 1.39 a | 403.62 ± 1.96 a | 410.43 ± 3.34 b | 11.44 ± 0.74 a |
| F3C3 | 7.36 ± 0.04 a | 25.83 ± 0.48 b | 125.46 ± 7.11 a | 30.65 ± 3.22 a | 400.68 ± 2.48 a | 453.64 ± 22.35 ab | 13.05 ± 20 a |
| Treatments | Ascomycota | Mortierellomycota | Basidiomycota |
|---|---|---|---|
| CK | 50.54 | 21.48 | 18.17 |
| F1C1 | 83.41 | 5.15 | 4.22 |
| F1C2 | 67.17 | 11.08 | 11.29 |
| F1C3 | 49.06 | 26.71 | 13.09 |
| F2C1 | 40.61 | 23.36 | 26.13 |
| F2C2 | 47.53 | 25.12 | 14.49 |
| F2C3 | 42.86 | 19.77 | 25.63 |
| F3C1 | 53.85 | 16.59 | 15.75 |
| F3C2 | 95.32 | 1.05 | 3.10 |
| F3C3 | 63.47 | 11.53 | 14.39 |
| Treatments | Peziza | Mortierella | Ilyonectria | Cylindrocarpon | Fusarium | Trichoderma | Cephaliophora | Minimelanolocus |
|---|---|---|---|---|---|---|---|---|
| CK | 0.02 | 8.85 | 1.89 | 0.12 | 2.34 | 1.75 | 3.28 | 0.02 |
| F1C1 | 0 | 2.77 | 0.64 | 0.07 | 5.09 | 5.52 | 0.59 | 0.02 |
| F1C2 | 0.01 | 4.25 | 6.04 | 33.02 | 2.09 | 1.25 | 1.16 | 0.05 |
| F1C3 | 0 | 8.80 | 4.61 | 0.30 | 1.56 | 0.93 | 2.95 | 0.02 |
| F2C1 | 0.01 | 17.03 | 6.63 | 0.08 | 0.69 | 0.86 | 1.05 | 0.05 |
| F2C2 | 0 | 10.17 | 1.89 | 0.19 | 2.71 | 0.81 | 0.50 | 0.05 |
| F2C3 | 0.01 | 5.28 | 2.55 | 0.15 | 2.14 | 1.06 | 0.61 | 0.04 |
| F3C1 | 0 | 7.59 | 7.85 | 0.26 | 3.73 | 2.41 | 1.23 | 0.44 |
| F3C2 | 91.16 | 0.19 | 0.44 | 0.01 | 1.52 | 0.26 | 0.10 | 0.01 |
| F3C3 | 0.06 | 5.05 | 2.56 | 0.42 | 6.38 | 1.84 | 2.29 | 11.72 |
| Treatments | ASV | Chao1 | Shannon | Simpson | ACE |
|---|---|---|---|---|---|
| CK | 342 ± 15.00 a | 342.31 ± 14.62 a | 5.99 ± 0.13 a | 0.95 ± 0.011 a | 342.63 ± 14.58 a |
| F1C1 | 224 ± 7.00 c | 224.14 ± 7.41 c | 3.49 ± 0.21 c | 0.69 ± 0.052 b | 224.07 ± 7.93 c |
| F1C2 | 311 ± 36.00 ab | 310.65 ± 36.21 ab | 4.96 ± 0.44 b | 0.86 ± 0.045 a | 310.40 ± 36.05 ab |
| F1C3 | 307 ± 46.00 ab | 306.68 ± 46.25 ab | 6.01 ± 0.04 a | 0.95 ± 0.007 a | 306.83 ± 46.29 |
| F2C1 | 232 ± 9.00 bc | 232.01 ± 9.01 bc | 5.43 ± 0.10 ab | 0.95 ± 0.004 a | 231.82 ± 9.18 bc |
| F2C2 | 341 ± 6.00 a | 340.98 ± 6.59 a | 5.97 ± 0.09 a | 0.96 ± 0.005 a | 341.16 ± 6.68 a |
| F2C3 | 233 ± 7.00 bc | 233.33 ± 7.22 bc | 4.77 ± 0.36 b | 0.87 ± 0.044 a | 233.26 ± 7.50 bc |
| F3C1 | 320 ± 9.00 a | 320.09 ± 9.20 a | 5.91 ± 0.20 a | 0.95 ± 0.009 a | 319.95 ± 9.42 a |
| F3C2 | 75 ± 3.00 d | 74.54 ± 3.90 d | 0.77 ± 0.25 d | 0.16 ± 0.065 c | 74.56 ± 4.49 d |
| F3C3 | 340 ± 15.00 a | 340.34 ± 15.50 a | 5.87 ± 0.08 a | 0.95 ± 0.012 a | 340.33 ± 15.48 a |
| Treatments | Indicative Species | Indicator.Values |
|---|---|---|
| CK | ASV_94.Basidiomycota.Melanophyllum | 0.93 |
| ASV_217.Basidiomycota.Subulicystidium | 0.90 | |
| F1C1 | \ | \ |
| F1C2 | ASV_5.Ascomycota.Cylindrocarpon | 0.99 |
| F1C3 | ASV_100.Ascomycota.Coleophoma | 0.88 |
| F2C1 | ASV_36.Mortierellomycota.Mortierella | 0.96 |
| F2C2 | ASV_129.Ascomycota.Paraboeremia | 0.96 |
| F2C3 | \ | \ |
| F3C1 | ASV_191.Ascomycota.Cephaliophora | 0.95 |
| F3C2 | ASV_1.Ascomycota.Peziza | 1.00 |
| F3C3 | ASV_235.Basidiomycota.Thanatephorus | 1.00 |
| ASV_202.Basidiomycota.Thanatephorus | 1.00 | |
| ASV_20.Ascomycota.Minimelanolocus | 0.99 | |
| ASV_67.Ascomycota.Dactylonectria | 0.87 |
| Treatments | Score | Sort |
|---|---|---|
| CK | 0.51 | 6 |
| F1C1 | 0.48 | 7 |
| F1C1 | 0.45 | 8 |
| F1C2 | 0.53 | 5 |
| F2C3 | 0.57 | 3 |
| F2C2 | 0.57 | 4 |
| F2C3 | 0.60 | 2 |
| F3C1 | 0.42 | 10 |
| F3C2 | 0.63 | 1 |
| F3C3 | 0.45 | 9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Zhou, Y.; Zhang, X.; Cao, N.; Li, B.; Liang, J.; Yang, Q. Effects of Combined Application of Biological Agent and Fertilizer on Fungal Community Structure in Rhizosphere Soil of Panax notoginseng. Agronomy 2023, 13, 2093. https://doi.org/10.3390/agronomy13082093
Liu Y, Zhou Y, Zhang X, Cao N, Li B, Liang J, Yang Q. Effects of Combined Application of Biological Agent and Fertilizer on Fungal Community Structure in Rhizosphere Soil of Panax notoginseng. Agronomy. 2023; 13(8):2093. https://doi.org/10.3390/agronomy13082093
Chicago/Turabian StyleLiu, Yanwei, Yingjie Zhou, Xiaofan Zhang, Ni Cao, Bin Li, Jiaping Liang, and Qiliang Yang. 2023. "Effects of Combined Application of Biological Agent and Fertilizer on Fungal Community Structure in Rhizosphere Soil of Panax notoginseng" Agronomy 13, no. 8: 2093. https://doi.org/10.3390/agronomy13082093