
How Are Warm-Season Pastures’ Nutritive Value and Fermentation Characteristics Affected by Open Pasture, Silvopasture, and Sward Herbage Maturity?

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
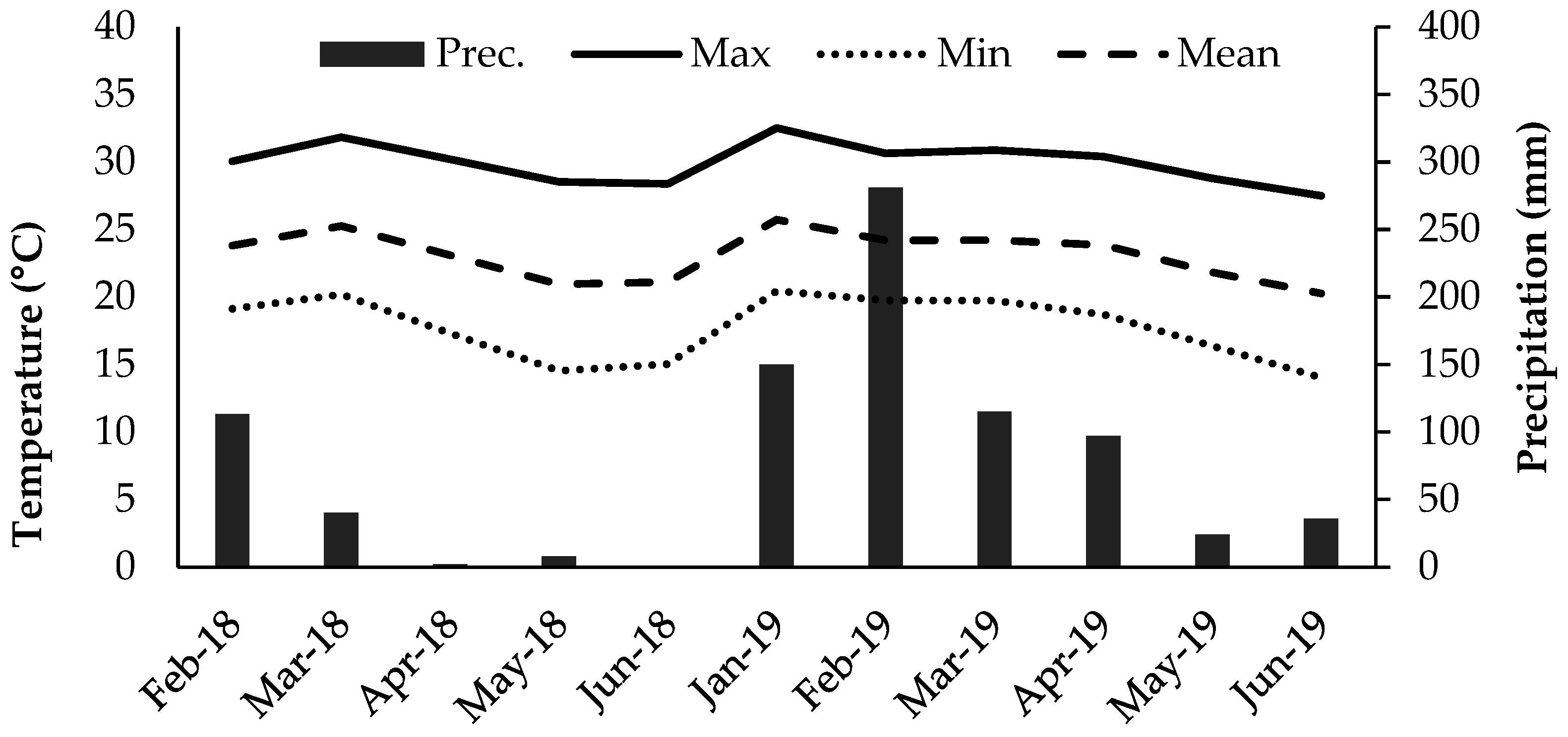
2.1. Experimental Area
2.2. Implementation of the Pasture Systems
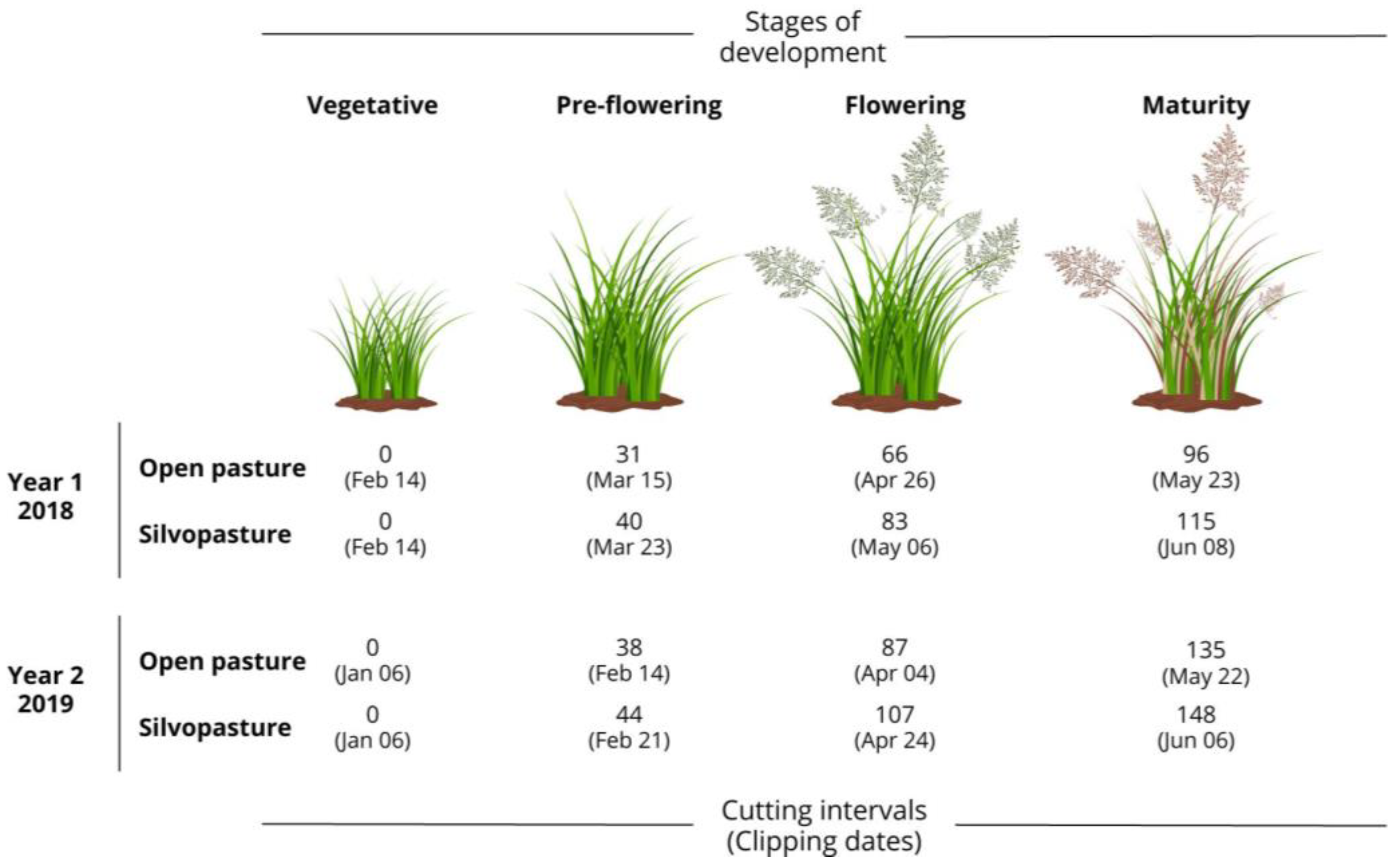
2.3. Treatments and Experimental Design
2.4. Evaluation of Forage Mass and Chemical Composition
2.5. Gas Production and In Vitro Degradability
2.6. Statistical Analyses
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Oliveira, C.C.; Alves, F.V.; de Almeida, R.G.; Gamarra, É.L.; Villela, S.D.J.; Martins, P.G.M.D.A. Thermal comfort indices assessed in integrated production systems in the Brazilian savannah. Agrofor. Syst. 2018, 92, 1659–1672. [Google Scholar] [CrossRef]
- Cardoso, A.S.; Alari, F.O.; de Andrade, N.; Nascimento, T.S.; van Cleef, F.D.O.S.; Alves, B.J.R.; Ruggieri, A.C.; Malheiros, E.B. Nitrous oxide emissions in silvopastoral systems: Key driving variables and seasonality. Agric. For. Meteorol. 2022, 316, 108851. [Google Scholar] [CrossRef]
- de Oliveira, A.F.; Menezes, G.L.; Gonçalves, L.C.; de Araújo, V.E.; Ramirez, M.A.; Júnior, R.G.; Jayme, D.G.; Lana, Â.M.Q. Pasture traits and cattle performance in silvopastoral systems with Eucalyptus and Urochloa: Systematic review and meta-analysis. Livest. Sci. 2022, 262, 104973. [Google Scholar] [CrossRef]
- Carvalho, C.A.B.D.; Silva, P.H.F.D.; Zanella, P.G.; Diavão, J.; Paiva, A.J. Morphogenetic and structural traits of tillers and herbage accumulation of Tanganyika grass under shading levels. Rev. Bras. Saúde E Produção Anim. 2022, 23, 1–6. [Google Scholar] [CrossRef]
- Oliveira, E.P.D.; Silveira, L.P.D.O.; Teodoro, P.E.; Ascoli, F.G.; Torres, F.E. Effect of shading and seed encrusting on the development of Panicum maximum Jacq cultivars. Biosci. J. 2014, 54, 1682–1691. [Google Scholar]
- Paciullo, D.S.C.; Gomide, C.D.M.; Castro, C.D.; Maurício, R.M.; Fernandes, P.B.; Morenz, M.J.F. Morphogenesis, biomass and nutritive value of Panicum maximum under different shade levels and fertilizer nitrogen rates. Grass Forage Sci. 2017, 72, 590–600. [Google Scholar] [CrossRef] [Green Version]
- Almeida, E.M.; de Almeida, R.G.; Es, M.; de Freitas, P.V.D.X.; Ribeiro, F.M.; Fernandes, P.B.; Garcia, E.D.C. Silvopastoral systems: An approach on the integration of biotic and abiotic components. Rev. Científica Rural 2019, 21, 438–454. [Google Scholar]
- Gobbi, K.F.; García, R.; Garcez Neto, A.F.; Pereira, O.G.; Rocha, G.C. Nutritional value of signalgrass and forage peanut submitted to shading. Arch. Zootec. 2010, 59, 379–390. [Google Scholar]
- Rezaeifard, M.; Jafari, A.A.; Assareh, M.H. Effects of phenological stages on forage yield quality traits in cocksfoot (Dactylis glomerata). J. Food Agric. Environ. 2010, 8, 365–369. [Google Scholar]
- Tesk, C.R.M.; Pedreira, B.C.; Pereira, D.H.; Pina, D.D.S.; Ramos, T.A.; Mombach, M.A. Impact of grazing management on forage qualitative characteristics: A review. Sci. Electron. Arch. 2018, 11, 188–197. [Google Scholar] [CrossRef] [Green Version]
- Sousa, L.F.; Maurício, R.M.; Moreira, G.R.; Gonçalves, L.C.; Borges, I.; Pereira, L.G.R. Nutritional evaluation of “Braquiarão” grass in association with “Aroeira” trees in a silvopastoral system. Agrofor. Syst. 2010, 79, 189–199. [Google Scholar] [CrossRef]
- Barros, J.S.; Meirelles, P.R.L.; Gomes, V.C.; Pariz, C.M.; Fachiolli, D.F.; Santana, E.A.R.; Costa, C.; Castilhos, C.A.; Souza, D.M. Nutritive value of xaraés palisade grass in three light intensities. Arq. Bras. Med. Veterinária E Zootec. 2019, 71, 1703–1711. [Google Scholar] [CrossRef] [Green Version]
- Lima, M.A.; Paciullo, D.S.; Silva, F.F.; Morenz, M.J.; Gomide, C.A.; Rodrigues, R.A.; Bretas, I.L.; Chizzotti, F.H. Evaluation of a long-established silvopastoral Brachiaria decumbens system: Plant characteristics and feeding value for cattle. Crop Pasture Sci. 2019, 70, 814–825. [Google Scholar] [CrossRef]
- Ruggieri, A.C.; Cardoso, A.S.; Ongaratto, F.; Casagrande, D.R.; Barbero, R.P.; Brito, L.D.F.; Oliveira, A.A.; Koscheck, J.F.W.; Reis, R.A. Grazing Intensity impacts on herbage mass, sward structure, greenhouse gas emissions, and animal performance: Analysis of brachiaria Pastureland. Agronomy 2020, 10, 1750. [Google Scholar] [CrossRef]
- Raij, B.V.; Cantarella, H.; Quaggio, J.A.; Furlani, A.M.C. Recomendações de Adubação e Calagem Para o Estado de São Paulo; Instituto Agronômico/Fundação IAC: Campinas, Brazil, 1997; pp. 237–239. [Google Scholar]
- Neel, J.P.S.; Felton, E.E.D.; Singh, S.; Sexstone, A.J.; Belesky, D.P. Open pasture, silvopasture and sward herbage maturity effects on nutritive value and fermentation characteristics of cool-season pasture. Grass Forage Sci. 2016, 71, 259–269. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis of the Association of Official Analytical Chemists, 17th ed.; Association of Official Analytical Chemists: Gaithersburg, MD, USA, 2000. [Google Scholar]
- ANKOM. Acid Detergent Fiber in Feeds. Filter Bag Technique (for A200, A200I); Ankom Technology Method 8; Ankom Technology Corp.: Macedon, NY, USA, 2006. [Google Scholar]
- Theodorou, M.K.; Williams, B.A.; Dhanoa, M.S.; McAllan, A.B.; France, J.A. Simple gas production method using a pressure transducer to determine the fermentation kinetics of ruminant feeds. Anim. Feed Sci. Technol. 1994, 48, 185–197. [Google Scholar] [CrossRef]
- Mauricio, R.M.; Mould, F.L.; Dhanoa, M.S.; Owen, E.; Channa, K.S.; Theodorou, M.K. A semi-automated in vitro gas production technique for ruminant feedstuff evaluation. Anim. Feed Sci. Technol. 1999, 79, 321–330. [Google Scholar] [CrossRef]
- Goering, H.K.; Van Soest, P.J. Forage Fiber Analysis: Apparatus Reagents, Procedures, and Some Applications; Agricultural Handbook 379; U.S. Agricultural Research Service: Washington, DC, USA, 1970; 20p.
- van Cleef, F.O.S.; van Cleef, E.H.C.B.; Longhini, V.Z.; Nascimento, T.S.; Ezequiel, J.M.B.; Ruggieri, A.C. Feedlot performance, carcass characteristics, and meat characteristics of lambs grown under silvopastoral systems. Can. J. Anim. Sci. 2019, 100, 385–388. [Google Scholar] [CrossRef]
- Cardoso, A.S.; Longhini, V.Z.; Berça, A.S.; Ongaratto, F.; Siniscalchi, D.; Reis, R.A.; Ruggieri, A.C. Pasture management and greenhouse gases emissions. Biosci. J. 2022, 38, e38099. [Google Scholar] [CrossRef]
- Pezzopane, J.R.M.; Bosi, C.; Nicodemo, M.L.F.; Santos, P.M.; Cruz, P.G.D.; Parmejiani, R.S. Microclimate and soil moisture in a silvopastoral system in southeastern Brazil. Bragantia 2015, 74, 110–119. [Google Scholar] [CrossRef] [Green Version]
- Kephart, K.D.; Buxton, D.R. Forage quality responses of C3 and C4 perennial grasses to shade. Crop Sci. 1993, 33, 831–837. [Google Scholar] [CrossRef]
- Wilson, J.R. Cell wall characteristics in relation to forage digestion by ruminants. J. Agric. Sci. 1994, 122, 173–182. [Google Scholar] [CrossRef]
- Caldeira, R.R. Avaliação da Produção e da Qualidade do Capim Panicum maximum cv. Massai. Master Thesis, Universidade de Brasília, Brasília, Brazil, 2016. [Google Scholar]
- Krahl, G.; Baldissera, T.C.; Pinto, C.E.; Garagorry, F.C.; Werner, S.S.; Lopes, C.F.; Ribeiro Filho, H.M.N. Can pasture defoliation management targets be similar under full sun and shaded conditions? Crop Pasture Sci. 2022, 74, 259–269. [Google Scholar] [CrossRef]
- Peralta, M.D.L.A.; Garcia, A.R.R.; Olmedo, F.M.; Mariela, P.M.; Aniano Aguirre, H.; Gasga Perez, L. Physical-chemical quality of Urochloa grasses in different phenological stages. Int. J. Agric. Environ. Biores. 2020, 5, 162–171. [Google Scholar] [CrossRef]
- Hodgson, J. Grazing Management—Science into Practice; Longman Scientific & Technical: Essex, New Zealand, 1990; 203p. [Google Scholar]
- Garcez, B.S.; Alves, A.A.; Macedo, E.D.O.; Santos, C.M.D.; Araújo, D.L.D.C.; Lacerda, M.D.S.B. Ruminal degradation of Panicum grasses in three post-regrowth ages. Ciênc. Anim. Bras. 2020, 21, 1–8. [Google Scholar] [CrossRef]
- Asaadi, A.M.; Yazdi, A.K. Phenological stage effects on forage quality of four forbs species. J. Food Agric. Environ. 2011, 9, 380–384. [Google Scholar]
- Garcez, B.S.; Alves, A.A.; Araújo, D.L.C.; Lacerda, M.D.S.B.; Souza, L.G.C.; Carvalho, L.F. Ruminal degradability of colonião grass (Panicum maximum Jacq. cv. Colonião) at three post-regrowth ages. Acta Vet. Bras. 2016, 10, 130–134. [Google Scholar] [CrossRef]
- Monção, F.P.; Costa, M.A.M.S.; Sampaio, J.P.; Gomes, M.; Leal, D.B.; Maranhão, C.M.A.; Chamone, J.M.A. Yield and nutritional value of BRS Capiaçu grass at different regrowth ages. Semin. Ciências Agrárias 2019, 40, 2045–2056. [Google Scholar] [CrossRef] [Green Version]
- Van Soest, P.J. Nutritional Ecology of the Ruminant; Cornell University Press: New York, NY, USA, 1994; 476p. [Google Scholar]
- Santo, A.X.; Silva, L.D.F.; Lançanova, J.A.C.; Ribeiro, E.L.A.; Mizubuti, I.Y.; Fortaleza, A.P.S.; Henz, E.L.; Júnior, F.L.M. Fractionation of carbohydrates, protein and "in vitro" ruminal degradation kinetics by gas production technique of supplementary rations containing sunflower cake. Arq. Bras. Med. Veterinária E Zootec. 2017, 69, 234–242. [Google Scholar] [CrossRef] [Green Version]
- Velásquez, P.A.T.; Berchielli, T.T.; Reis, R.A.; Rivera, A.R.; Dian, P.H.M.; Teixeira, I.A.M.D.A. Chemical composition, fractionation of carbohydrates and crude protein and in vitro digestibility on tropical forages in the different cutting ages. Rev. Bras. Zootec. 2010, 39, 1206–1213. [Google Scholar] [CrossRef] [Green Version]
- Xue, Z.; Liu, N.; Wang, Y.; Yang, H.; Wei, Y.; Moriel, P.; Palmer, E.; Zhang, Y. Combining orchardgrass and alfalfa: Effects of forage ratios on in vitro rumen degradation and fermentation characteristics of silage compared with hay. Animals 2019, 10, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopes, C.M.; Paciullo, D.S.C.; Araújo, S.A.C.; Gomide, C.A.M.; Morenz, M.J.F.; Villela, S.D.J. Herbage mass, morphological composition, and nutritive value of signalgrass, submitted to shading and fertilization levels. Arq. Bras. Med. Veterinária E Zootec. 2017, 69, 225–233. [Google Scholar] [CrossRef] [Green Version]
- Collins, M.; Newman, Y.C. Forage quality. In Forages, 7th ed.; Collins, M., Nelson, C.J., Moore, K.J., Barnes, R.F., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2018; Volume 1, pp. 269–285. [Google Scholar]



{kind=link}
{kind=link}
| Properties | SPS | OP |
|---|---|---|
| pH (CaCl2) | 5.3 | 5.4 |
| Organic matter (g dm−3) | 26 | 25 |
| P (g dm−3) | 38 | 36 |
| S (g dm−3) | 5 | 5 |
| Ca (mmolc dm−3) | 24 | 26 |
| Mg (mmolc dm−3) | 13 | 13 |
| K (mmolc dm−3) | 2.8 | 3.0 |
| Al (mmolc dm−3) | 0 | 0 |
| H + Al (mmolc dm−3) | 31 | 28 |
| Sum of bases (mmolc dm−3) | 38.5 | 42.4 |
| Cation exchange capability | 69 | 70 |
| Base saturation (V%) | 56 | 60 |
| Variable | Systems | SEM | SD | Effect | p-Value | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| OP | SPS | 1 | 2 | 3 | 4 | S | SD | S × SD | |||
| Forage mass a | 5740 | 5704 | 2811 | 2931 | 4322 | 7006 | 8475 | LIN | 0.959 | <0.001 | 0.736 |
| Leaf (%) | 58.74 | 58.14 | 11.33 | 83.41 | 65.96 | 42.80 | 42.99 | LIN | 0.826 | <0.001 | 0.523 |
| Stem (%) | 20.56 | 21.32 | 10.23 | 8.25 | 15.65 | 29.31 | 29.71 | LIN | 0.762 | <0.001 | 0.769 |
| Dead (%) b | 20.68 | 20.53 | 12.23 | 8.34 | 18.39 | 27.36 | 27.57 | LIN | 0.959 | 0.004 | 0.953 |
| LSR c | 5.10 | 5.19 | 3.09 | 11.98 | 5.23 | 1.76 | 1.99 | LIN | 0.909 | <0.001 | 0.417 |
| Variable | System (S) | SEM | Stage of Development (SD) | Effect | p-Value | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| OP | SPS | 1 | 2 | 3 | 4 | S | DS | S × DS | |||
| CP a | 76.5 | 84.4 | 14.5 | 118.7 | 81.3 | 65.8 | 48.0 | LIN | 0.023 | <0.001 | 0.994 |
| NDF b | 742.4 | 736.6 | 22.3 | 736.1 | 753.8 | 736.1 | 732.0 | QUA | 0.243 | 0.020 | 0.250 |
| ADF c | 391.9 | 390.7 | 22.2 | 376.6 | 385.5 | 397.8 | 404.4 | LIN | 0.818 | 0.002 | 0.879 |
| HEM d | 353.9 | 345.7 | 31.9 | 353.8 | 357.5 | 342.0 | 345.5 | NS | 0.292 | 0.488 | 0.787 |
| CEL e | 347.4 | 345.2 | 20.0 | 347.9 | 344.6 | 350.4 | 342.5 | NS | 0.665 | 0.673 | 0.817 |
| LIG f | 29.4 | 34.0 | 10.8 | 28.9 | 25.3 | 34.8 | 37.4 | LIN | 0.074 | 0.004 | 0.406 |
| Ash | 101.2 | 102.6 | 11.9 | 10.58 | 110.1 | 94.6 | 97.3 | LIN | 0.649 | <0.001 | 0.859 |
| Variable | System (S) | SEM | Stage of Development (DS) | SEM | p-Value | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Open Pasture | SPS | 1 | 2 | 3 | 4 | S | DS | S × DS | |||
| Vf (mL g−1 OM) a | 158 | 168 | 3.89 | 196a | 164b | 140c | 150bc | 14 | 0.0389 | <0.0001 | |
| L (h) b | 2.90 | 5.36 | 0.75 | 6.37a | 6.79a | 1.51b | 1.85b | 2.39 | 0.0213 | 0.0098 | |
| Variable | System (S) | SEM | Stage of Development (SD) | Effect | p-Value | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Open Pasture | SPS | 1 | 2 | 3 | 4 | S | DS | S vs. DS | |||
| DIVDM 24 h | 284.5 | 297.6 | 7.8 | 344.0 | 319.8 | 274.9 | 225.4 | LIN | 0.235 | <0.001 | 0.127 |
| DIVDM 48 h | 447.9 | 453.8 | 7.1 | 552.4 | 502.0 | 394.0 | 354.8 | LIN | 0.562 | <0.001 | 0.147 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferreira, M.R.; Cardoso, A.d.S.; Andrade, M.E.B.; Brito, T.R.; Ruggieri, A.C. How Are Warm-Season Pastures’ Nutritive Value and Fermentation Characteristics Affected by Open Pasture, Silvopasture, and Sward Herbage Maturity? Agronomy 2023, 13, 1756. https://doi.org/10.3390/agronomy13071756
Ferreira MR, Cardoso AdS, Andrade MEB, Brito TR, Ruggieri AC. How Are Warm-Season Pastures’ Nutritive Value and Fermentation Characteristics Affected by Open Pasture, Silvopasture, and Sward Herbage Maturity? Agronomy. 2023; 13(7):1756. https://doi.org/10.3390/agronomy13071756
Chicago/Turabian StyleFerreira, Mariane Rodrigues, Abmael da Silva Cardoso, Marina Elizabeth Barbosa Andrade, Thais Ribeiro Brito, and Ana Cláudia Ruggieri. 2023. "How Are Warm-Season Pastures’ Nutritive Value and Fermentation Characteristics Affected by Open Pasture, Silvopasture, and Sward Herbage Maturity?" Agronomy 13, no. 7: 1756. https://doi.org/10.3390/agronomy13071756