
The Impact of Root-Invasive Fungi on Dominant and Invasive Plant Species in Degraded Grassland at Nanshan Pasture

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Preparation of Soil and Tested Fungi
2.3. Seed and Inoculum Preparation
2.4. Greenhouse Experiment
2.5. Harvesting and Measurements
2.6. Data Analysis and Statistical Analysis
3. Results
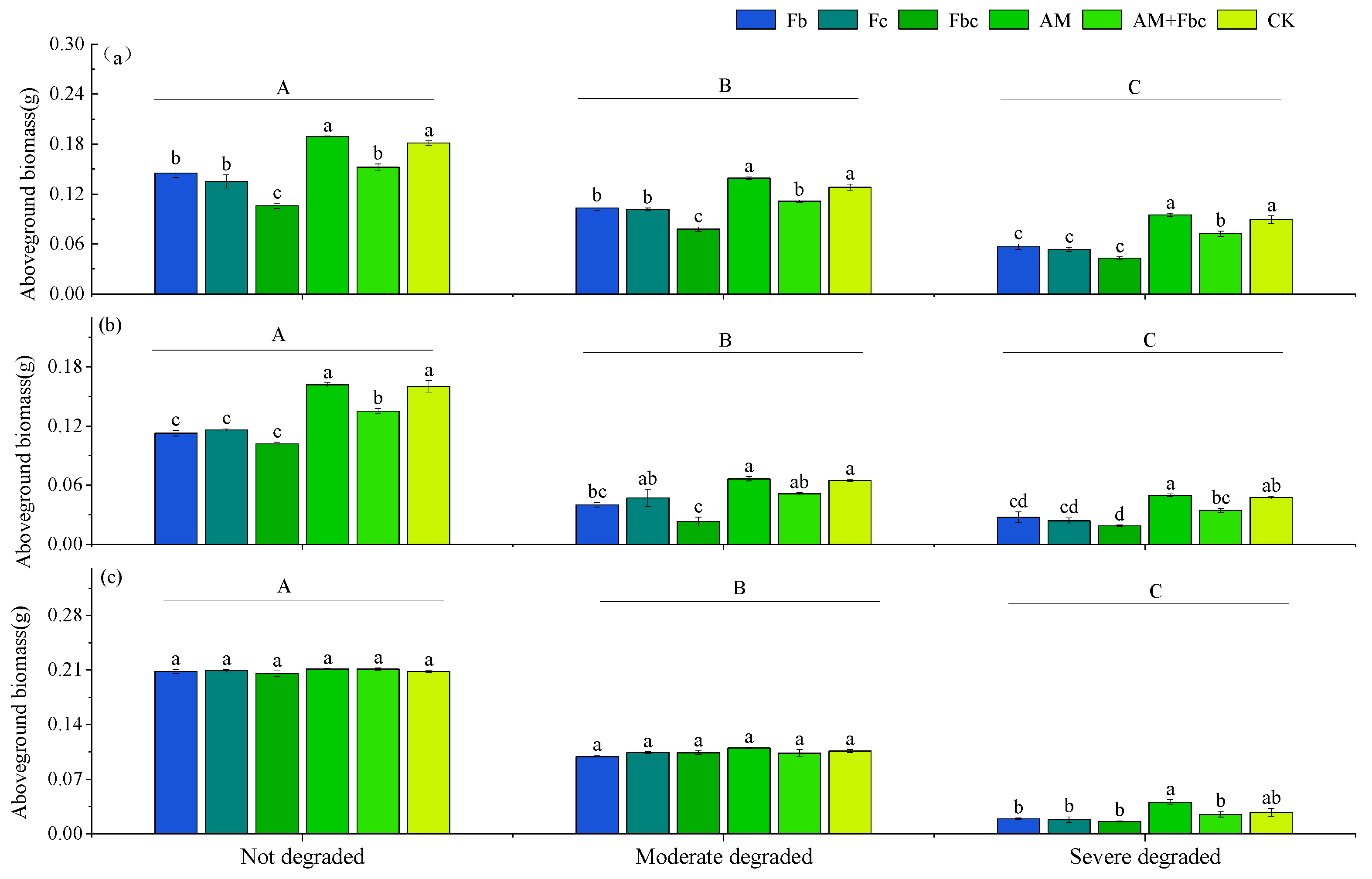
3.1. Effects of Pathogenic Fungi on Plant Biomass
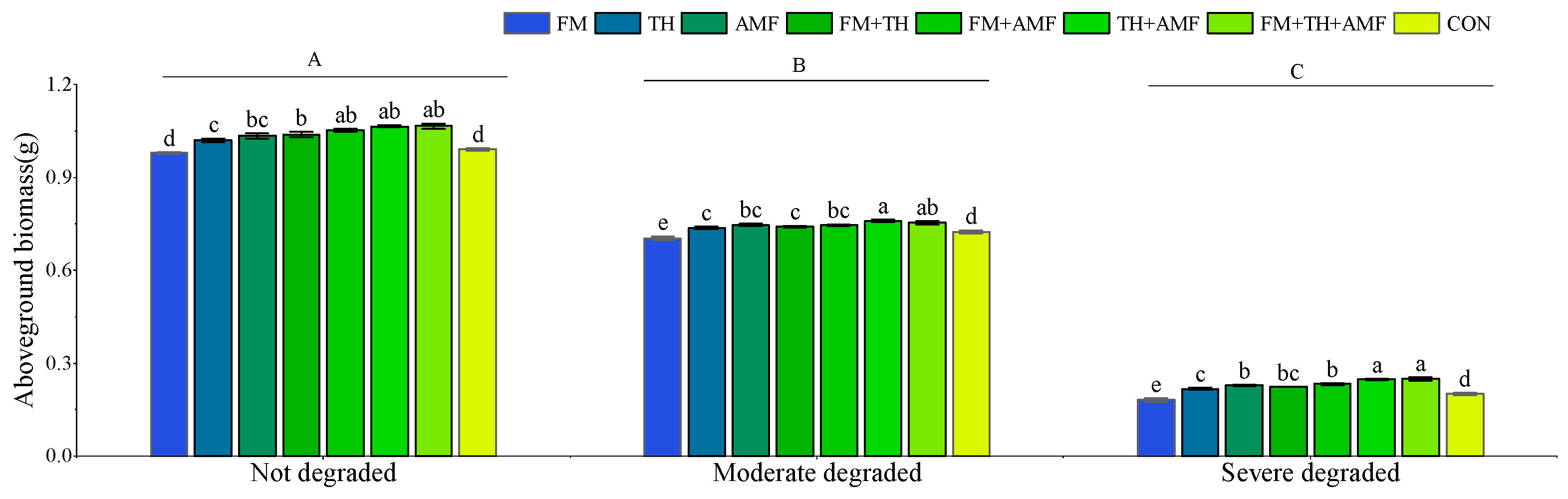
3.2. Community Total Biomass
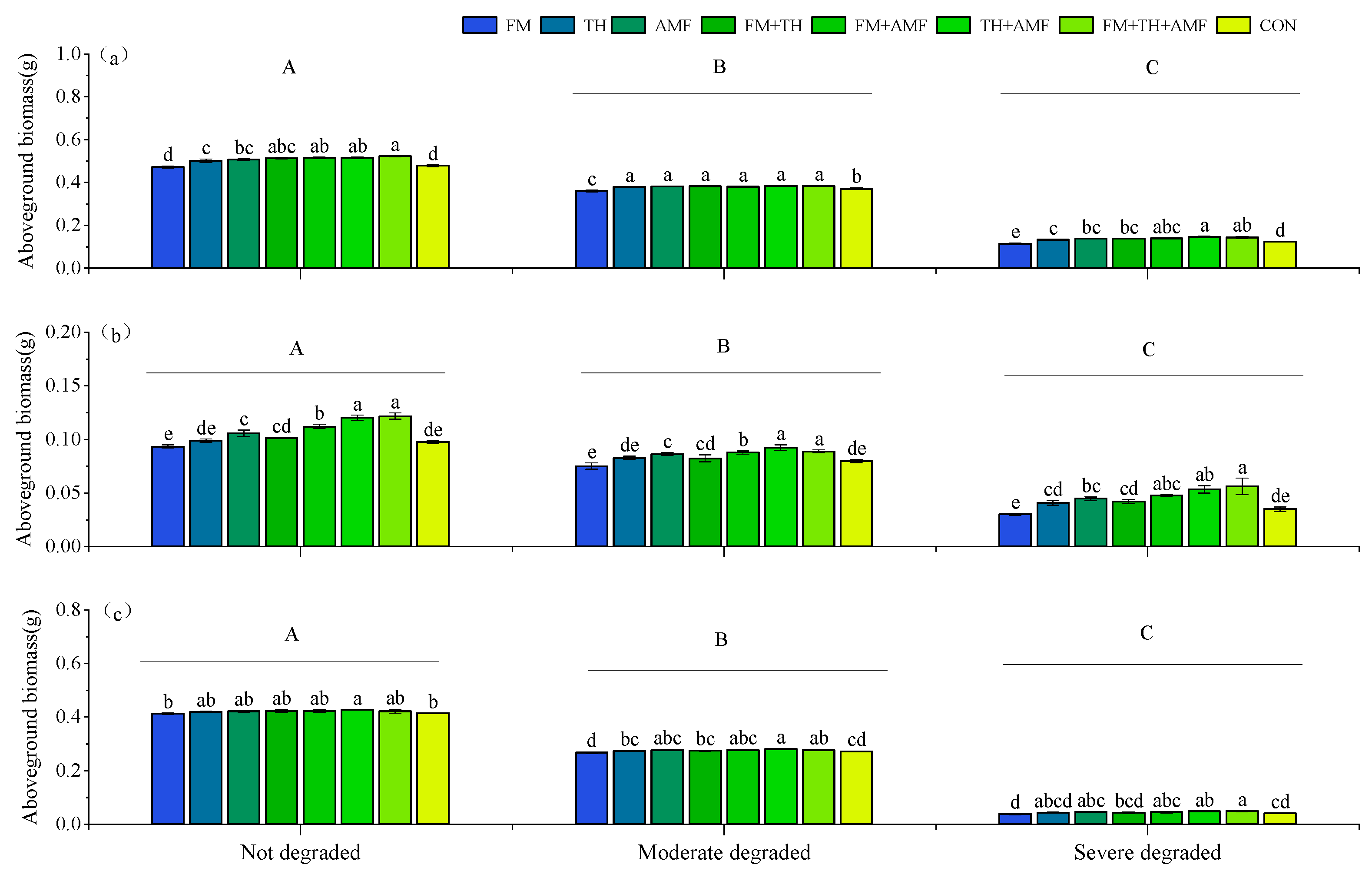
3.3. The Biomass of Each Plant Species in the Community
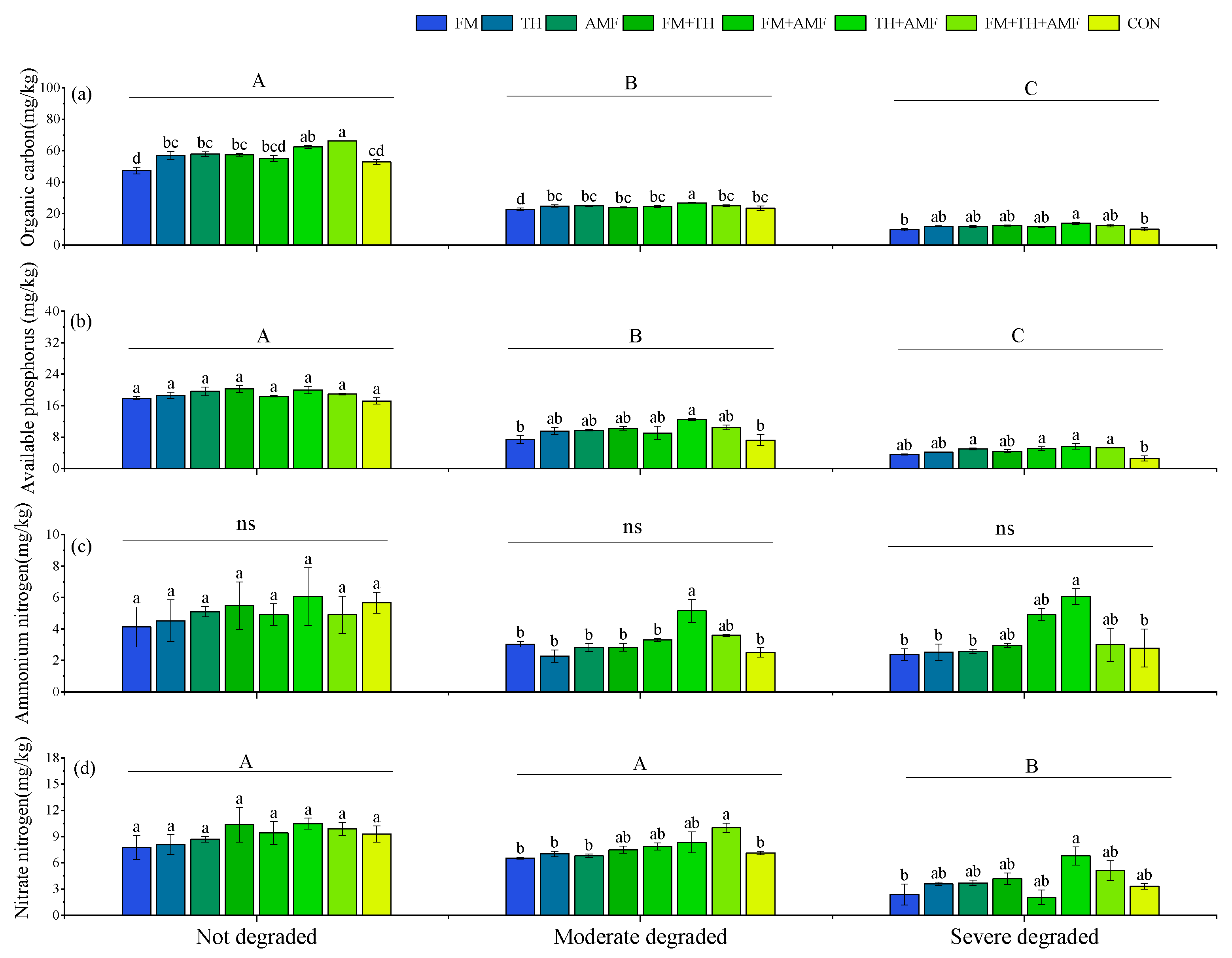
3.4. Soil Nutrients
4. Discussion
4.1. Effects of Pathogenic Fungi on Plants
4.2. The Role of Pathogenic Fungi in Plant Communities
4.3. The Role of Symbiotic Fungi in the Community
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sun, H.; Zhou, H.; Wang, P. Weed invasion and grassland quality during degraded succession of mixed artificial grassland. Grassl. Lawn 2001, 1, 33–37. [Google Scholar]
- Duan, D. Research on the establishment of artificial grassland in mountainous areas of southern China. Sci. Bull. 1992, 8, 180–183. [Google Scholar]
- Hong, F.; Wang, Y. Review and thinking on animal husbandry of artificial grassland in South China. J. Grassl. Sci. China 2006, 28, 71–75. [Google Scholar]
- Maron, J.L.; Klironomos, J.; Waller, L.; Callaway, R.M.; Austin, A. Invasive Plants Escape from Suppressive Soil Biota at Regional Scales. J. Ecol. 2014, 102, 19–27. [Google Scholar] [CrossRef] [Green Version]
- Mangla, S.; Callaway, R.M. Exotic invasive plant accumulates native soil pathogens which inhibit native plants. J. Ecol. 2007, 96, 58–67. [Google Scholar] [CrossRef]
- Callaway, R.M.; Thelen, G.C.; Barth, S.; Ramsey, P.W.; Gannon, J.E. Soil Fungi Alter Interactions between the Invader Centaurea Maculosa and North American Natives. Ecology 2004, 85, 1062–1071. [Google Scholar] [CrossRef] [Green Version]
- Lambers, H.; Mougel, C.; Jaillard, B.; Hinsinger, P. Plant-Microbe-Soil Interactions in the Rhizosphere: An Evolutionary Perspective. Plant Soil 2009, 321, 83–115. [Google Scholar]
- Lagueux, D.; Jumpponen, A.; Porras-Alfaro, A.; Herrera, J.; Chung, Y.A.; Baur, L.E.; Smith, M.D.; Knapp, A.K.; Collins, S.L.; Rudgers, J.A. Experimental Drought Re-Ordered Assemblages of Root-Associated Fungi across North American Grasslands. J. Ecol. 2021, 109, 776–792. [Google Scholar] [CrossRef]
- Shiragane, H.; Usami, T.; Shishido, M. Weed Roots Facilitate the Spread of Rosellinia Necatrix, the Causal Agent of White Root Rot. Microbes Environ. 2019, 34, 340–343. [Google Scholar] [CrossRef] [Green Version]
- Pearson, K.A.; Taylor, A.F.S.R.; Fuchs, M.E.; Woodward, S. Characterisation and Pathogenicity of Fusarium Taxa Isolated from Ragwort (Jacobaea Vulgaris) Roots. Fungal Ecol. 2016, 20, 186–192. [Google Scholar]
- Soti, P.G.; Purcell, M.; Jayachandran, K. Soil Biotic and Abiotic Conditions Negate Invasive Species Performance in Native Habitat. Ecol. Process. 2020, 9, 18. [Google Scholar] [CrossRef]
- Zhang, P.; Li, B.; Wu, J.; Hu, S. Invasive plants differentially affect soil biota through litter and rhizosphere pathways: A meta-analysis. Ecol. Lett. 2018, 22, 200–210. [Google Scholar] [CrossRef] [Green Version]
- Ravichandran, K.R.; Thangavelu, M. Role and Influence of Soil Microbial Communities on Plant Invasion. Ecol. Quest. 2017, 27, 9–23. [Google Scholar] [CrossRef] [Green Version]
- Bao, S. Soil Agricultural Chemical Elements Analysis; China Agriculture Press: Beijing, China, 2000; pp. 50–57. [Google Scholar]
- Song, C.; Han, J.; Gao, C. Research progress on soil available phosphorus and its chemical testing methods. Agric. Syst. Sci. Integr. Res. 2010, 26, 283–287. [Google Scholar]
- Nelson, D.W.; Sommers, L.E. Total Carbon, Organic Carbon, and Organic Matter. Methods Soil Anal. 1982, 9, 961–1010. [Google Scholar]
- Bever, J.D.; Mangan, S.A.; Alexander, H.M. Maintenance of Plant Species Diversity by Pathogens. Annu. Rev. Ecol. Evol. Syst. 2015, 46, 305–325. [Google Scholar] [CrossRef]
- Ampt, E.A.; van Ruijven, J.; Raaijmakers, J.M.; Termorshuizen, A.J.; Mommer, L. Linking ecology and plant pathology to unravel the importance of soil-borne fungal pathogens in species-rich grasslands. Eur. J. Plant Pathol. 2018, 154, 141–156. [Google Scholar] [CrossRef] [Green Version]
- Jarosz, A.M.; Davelos, A.L. Effects of disease in wild plant populations and the evolution of pathogen aggressiveness. New Phytol. 1995, 129, 371–387. [Google Scholar] [CrossRef]
- Tao, Z.W.; Bu, H.; Li, J.; Jia, P.; Qi, W.; Liu, K.; Du, G.Z. Effects of different artificial planting schemes on invasive weeds. Glob. Ecol. Conserv. 2021, 28, e01651. [Google Scholar] [CrossRef]
- Ke, W.; Pan, Y.; Chen, L.; Huang, J.; Zhang, J.; Long, X.; Cai, M.; Peng, C. Adaptive photosynthetic strategies of the invasive plant Sphagneticola trilobata and its hybrid to a low-light environment. Photosynthetica 2022, 60, 549–561. [Google Scholar] [CrossRef]
- Klironomos, J.N. Feedback with soil biota contributes to plant rarity and invasiveness in communities. Nature 2002, 417, 67–70. [Google Scholar] [CrossRef] [PubMed]
- Kardol, P.; De Deyn, G.; Laliberté, E.; Mariotte, P.; Hawkes, C.V. Biotic plant-soil feedbacks across temporal scales. J. Ecol. 2013, 101, 309–315. [Google Scholar] [CrossRef] [Green Version]
- Grman, E.; Robinson, T.M.P. Resource Availability and Imbalance Affect Plant-Mycorrhizal Interactions: A Field Test of Three Hypotheses. Ecology 2013, 94, 62–71. [Google Scholar] [CrossRef] [PubMed]
- Ampt, E.A.; Francioli, D.; van Ruijven, J.; Gomes, S.I.F.; Maciá-Vicente, J.G.; Termorshuizen, A.J.; Bakker, L.M.; Mommer, L. Deciphering the interactions between plant species and their main fungal root pathogens in mixed grassland communities. J. Ecol. 2022, 110, 3039–3052. [Google Scholar] [CrossRef]
- Maron, J.L.; Smith, A.L.; Ortega, Y.K.; Pearson, D.E.; Callaway, R.M. Negative Plant-Soil Feedbacks Increase with Plant Abundance, and Are Unchanged by Competition. Ecology 2016, 97, 2055–2063. [Google Scholar] [CrossRef]
- Delgado-Baquerizo, M.; Reich, P.B.; Trivedi, C.; Eldridge, D.J.; Abades, S.; Alfaro, F.D.; Bastida, F.; Berhe, A.A.; Cutler, N.A.; Gallardo, A.; et al. Multiple elements of soil biodiversity drive ecosystem functions across biomes. Nat. Ecol. Evol. 2020, 4, 210–220. [Google Scholar] [CrossRef] [PubMed]
- Fanin, N.; Gundale, M.J.; Farrell, M.; Ciobanu, M.; Baldock, J.A.; Nilsson, M.-C.; Kardol, P.; Wardle, D.A. Consistent effects of biodiversity loss on multifunctionality across contrasting ecosystems. Nat. Ecol. Evol. 2017, 2, 269–278. [Google Scholar] [CrossRef]
- Guerra, C.A.; Bardgett, R.D.; Caon, L.; Crowther, T.W.; Delgado-Baquerizo, M.; Montanarella, L.; Navarro, L.M.; Orgiazzi, A.; Singh, B.K.; Tedersoo, L.; et al. Tracking, targeting, and conserving soil biodiversity. Science 2021, 371, 239–241. [Google Scholar] [CrossRef]
- Reynolds, H.L.; Packer, A.; Bever, J.D.; Clay, K. Grassroots Ecology: Plant-Microbe-Soil Interactions as Drivers of Plant Community Structure and Dynamics. Ecology 2003, 84, 2281–2291. [Google Scholar] [CrossRef] [Green Version]
- Shinichi, N.; Schielzeth, H. A General and Simple Method for Obtaining R2 from Generalized Linear Mixed-Effects Models. Methods Ecol. Evol. 2013, 4, 133–142. [Google Scholar]
- Harman, G.E.; Uphoff, N. Symbiotic Root-Endophytic Soil Microbes Improve Crop Productivity and Provide Environmental Benefits. Scientifica 2019, 2019, 9106395. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | DF | F | p |
|---|---|---|---|
| Plant species (P) | 2 | 208.0 | <0.001 |
| Degree of degradation (D) | 2 | 50,746.9 | <0.001 |
| Fungal treatment (F) | 5 | 6696.7 | <0.001 |
| P × D | 4 | 32.3 | <0.001 |
| P × F | 10 | 501.2 | <0.001 |
| D × F | 10 | 4.0 | <0.001 |
| P × D × F | 20 | 3.6 | 0.08 |
| Treatment | DF | AB | SOC | NO3−-N | NH4+-N | AP | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| F | p | F | p | F | p | F | p | F | p | ||
| Degree of degradation (D) | 2 | 170,582.298 | <0.001 | 3492.557 | <0.001 | 73.577 | <0.01 | 11.169 | <0.001 | 770.432 | <0.001 |
| Fungal treatment (F) | 7 | 221.834 | <0.001 | 16.955 | <0.001 | 4.595 | <0.001 | 3.252 | <0.01 | 7.571 | <0.001 |
| D × F | 14 | 8.840 | <0.001 | 6.121 | <0.001 | 0.469 | 0.938 | 0.056 | 0.885 | 0.735 | 0.730 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Chang, J.; Xie, J.; Yang, L.; Sheteiwy, M.S.; Moustafa, A.-R.A.; Zaghloul, M.S.; Ren, H. The Impact of Root-Invasive Fungi on Dominant and Invasive Plant Species in Degraded Grassland at Nanshan Pasture. Agronomy 2023, 13, 1666. https://doi.org/10.3390/agronomy13071666
Zhang Y, Chang J, Xie J, Yang L, Sheteiwy MS, Moustafa A-RA, Zaghloul MS, Ren H. The Impact of Root-Invasive Fungi on Dominant and Invasive Plant Species in Degraded Grassland at Nanshan Pasture. Agronomy. 2023; 13(7):1666. https://doi.org/10.3390/agronomy13071666
Chicago/Turabian StyleZhang, Yanxia, Jiechao Chang, Jiayao Xie, Liquan Yang, Mohamed S. Sheteiwy, Abdel-Raouf A. Moustafa, Mohamed S. Zaghloul, and Haiyan Ren. 2023. "The Impact of Root-Invasive Fungi on Dominant and Invasive Plant Species in Degraded Grassland at Nanshan Pasture" Agronomy 13, no. 7: 1666. https://doi.org/10.3390/agronomy13071666