
Using Ecological Stoichimetric Characteristies to Inform Grassland Management in the Karst Desertification Area

Abstract
:1. Introduction
2. Materials and Methods
2.1. Research Area
2.2. Sample Plot Setting
2.3. Sample Collection
2.4. Determination of Samples
2.5. Data Processing
3. Results
3.1. C, N and P Contents and Stoichiometric Ratios of Grassland Plants under Different Utilization Methods
3.2. C, N and P Contents and Stoichiometric Ratio of Grassland Soil under Different Utilization Methods
3.3. MBC, MBN and MBP Contents and Stoichiometric Ratio of Grassland Soil under Different Utilization Methods
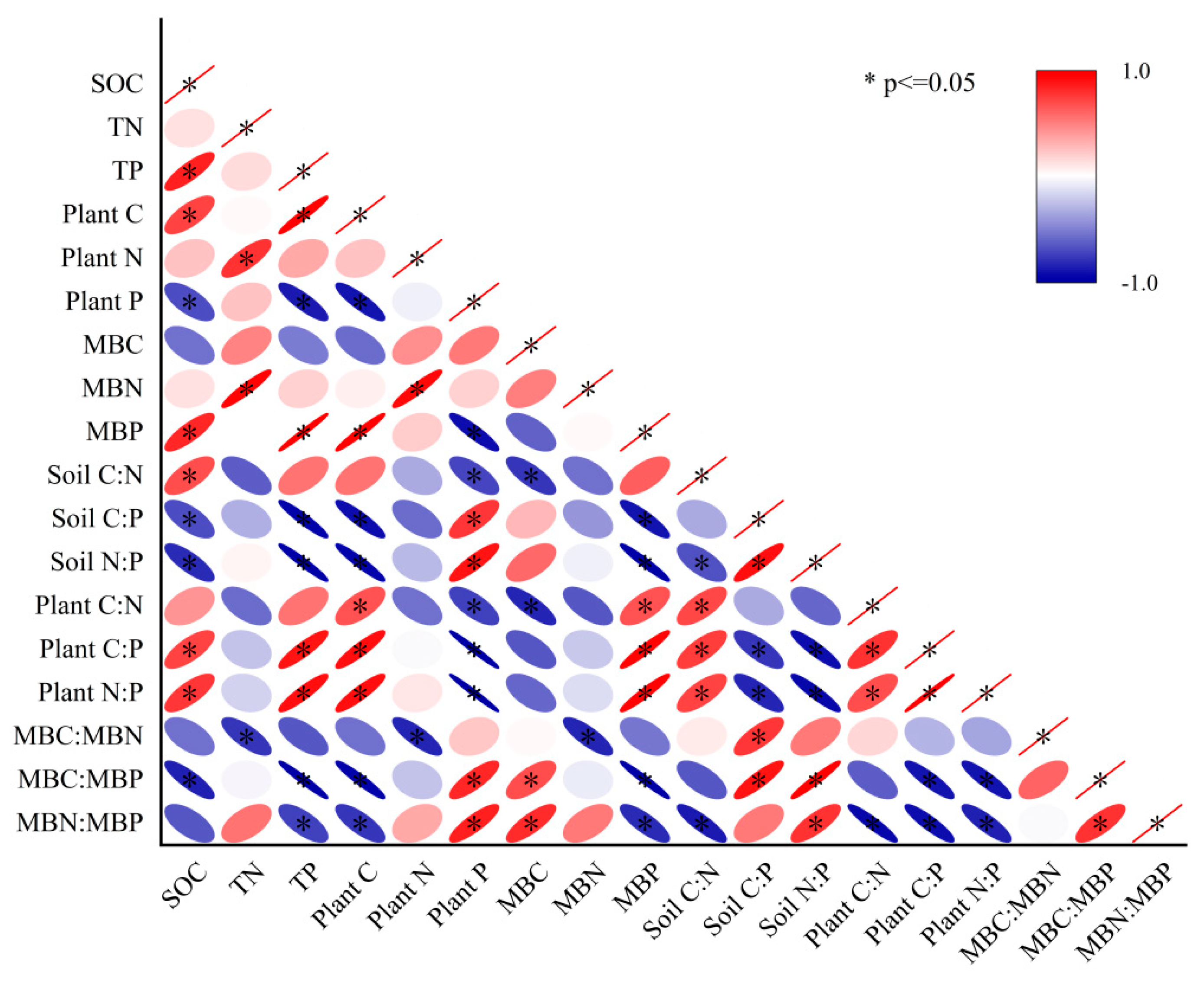
3.4. Correlation Analysis of C, N, P Contents and Stoichiometric Ratios in Plant–Soil–Microorganism
4. Discussion
4.1. Plant C, N and P Content and Stoichiometric Ratio
4.2. Soil C, N and P Nutrient Content and Stoichiometric Ratio
4.3. Microorganisms C, N and P Content and Stoichiometric Ratio
4.4. Plant–Soil–Microbial Stoichiometric Correlation Analysis
5. Conclusions
- (1)
- There were significant differences in C, N, P and the ecological stoichiometric characteristics of plants under different grassland utilization methods. Among the three treatments, the EG treatment had the highest plant C content, while the MG treatment had the lowest. The plant P of GG and MG was significantly higher than that of EG. The plant N content in GG was significantly higher than that in MG and EG. The ratio of plant N:P in the three treatments was higher than 20, and P deficiency and P limitation existed in the three treatments. The P limitation of EG was the most serious, and that of MG was the least.
- (2)
- There were some differences in soil nutrients and ecological stoichiometry among the three treatments. EG may be more beneficial to accumulate SOC and increase TP content. The C:N in EG was higher than that in GG and MG, and the decomposition rate of SOC in EG was slower, which was beneficial to the long-term maintenance of soil fertility. The C:P ratio of MG was the highest, and the net mineralization of P in MG was stronger than that of GG and EG. Soil N:P ratios under three different grassland-use patterns were all lower than 14, indicating that there was obvious N limitation.
- (3)
- There were some differences in soil microbial biomass under different grassland-utilization methods. The MBC of GG and MG was higher than that of EG, but the MBN of EG was between that of GG and MG, and the MBP of EG was the highest. The MBC:MBN under the three different grassland-utilization methods ranged from 2.48 to 3.88, which was lower than the global average range. The MBC:MBP also showed significant differences, with MG being the highest and EG the lowest. The MBN:MBP ratios of GG and MG were both higher than 9.6, indicating that they were P-limited, while the MBN:MBP of EG was lower than 8.9, indicating that it was N-limited.
- (4)
- Plant C and N were significantly correlated with soil C and N (p < 0.05). Plant P was significantly negatively correlated with soil P (p < 0.05), but MBP was significantly positively correlated with soil TP (p < 0.05). The results of MBC:MBP showed that soil microorganisms in GG and MG had a tendency to assimilate soil available phosphorus, and there was a tendency for microorganisms to compete with crops to absorb soil available phosphorus, which had a strong phosphorus fixation potential. Meanwhile, soil microorganisms in EG had a greater potential to release P from mineralized soil organic matter, and MBP could supplement soil available phosphorus pool.
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reich, P.B.; Walters, M.B.; Ellsworth, D.S. From tropics to tundra: Global convergence in plant functioning. Proc. Natl. Acad. Sci. USA 1997, 94, 13730–13734. [Google Scholar] [CrossRef] [PubMed]
- Reich, P.B.; Tjoelker, M.G.; Machado, J.L.; Oleksyn, J. Universal scaling of respiratory metabolism, size and nitrogen in plants. Nature 2006, 439, 457–461. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Hou, Y.; Zheng, R.L.; Hu, Y.X.; Li, X.N. Plant diversity and nitrogen addition affect the architecture of plant–soil–microbe stoichiometric networks. Plant Soil 2023. [Google Scholar] [CrossRef]
- Han, W.X.; Fang, J.Y.; Reich, P.B.; Ian Woodward, F.; Wang, Z.H. Biogeography and variability of eleven mineral elements in plant leaves across gradients of climate, soil and plant functional type in China. Ecol. Lett. 2011, 14, 788–796. [Google Scholar] [CrossRef]
- Song, S.Z.; Wang, X.L.; He, C.; Chi, Y.K. Effects of utilization methods on C, N, P rate and enzyme activity of artificial grassland in karst desertification area. Agronomy 2023, 13, 1368. [Google Scholar] [CrossRef]
- Sun, B.; Barnes, A.D. Editorial: Soil-root-microbe interactions promote soil and plant health. Front. Microbiol. 2023, 14, 1155234. [Google Scholar] [CrossRef]
- Chen, Z.J.; Li, Y.; Hu, M.; Xiong, Y.W.; Huang, Q.Z.; Jin, S.; Huang, G.H. Lignite bioorganic fertilizer enhanced microbial co-occurrence network stability and plant–microbe interactions in saline-sodic soil. Sci. Total Environ. 2023, 879, 163113. [Google Scholar] [CrossRef]
- Elser, J.J.; Fagan, W.F.; Kerkhof, A.J.; Swenson, N.G.; Enquist, B.J. Biological stoichiometry of plant production: Metabolism, scaling and ecological response to global change. New Phytol. 2010, 186, 593–608. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.P.; Li, Q.; Feng, R.Q.; Zhang, Z.H.; Yang, Y.F.; Liu, J. C:N:P stoichiometry of plant-soil-microbe in the secondary succession of zokor-made mounds on Qinghai-Tibet Plateau. Environ. Res. 2023, 222, 115333. [Google Scholar] [CrossRef]
- Isles, P.D.F. The misuse of ratios in ecological stoichiometry. Ecology 2020, 101, e03153. [Google Scholar] [CrossRef]
- Oldroyd, G.E.D.; Leyser, O.A. Plant’s diet, surviving in a variable nutrient environment. Science 2020, 368, eaba0196. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Ren, Z.; Ma, P.; Wang, Z.; Elser, J.J. Effects of grassland degradation on ecological stoichiometry of soil ecosystems on the Qinghai-Tibet Plateau. Sci. Total Environ. 2020, 722, 137910. [Google Scholar] [CrossRef] [PubMed]
- Cleveland, C.C.; Liptzin, D. C:N:P stoichiometry in soil: Is there a “Redfield ratio” for the microbial biomass? Biogeochemistry 2007, 85, 235–252. [Google Scholar] [CrossRef]
- Hu, P.L.; Wang, K.L.; Zeng, Z.X.; Zhang, H.; Li, S.S.; Song, X.J. Ecological stoichiometric characteristics of plants, soil, and microbes of Pennisetum purpureum cv. Guimu-1 pastures at different rehabilitation ages in a karst rocky desertification region. Acta Ecol. Sin. 2017, 37, 896–905. [Google Scholar]
- Garousi, F.; Shan, Z.J.; Ni, K.; Yang, H.; Shan, J.; Cao, J.H.; Jiang, Z.C.; Yang, J.L.; Zhu, T.B.; Müller, C. Decreased inorganic N supply capacity and turnover in calcareous soil under degraded rubber plantation in the tropical karst region. Geoderma 2021, 381, 114754. [Google Scholar] [CrossRef]
- Xiong, K.N.; Chi, Y.K.; Shen, X.Y. Research on photosynthetic leguminous forage in the karst rocky desertification regions of Southwestern China. Pol. J. Environ. Stud. 2017, 26, 2319–2329. [Google Scholar] [CrossRef]
- Wang, K.L.; Yue, Y.M.; Chen, H.S.; Zeng, F.P. Mechanisms and realization pathways for integration of scientifific poverty alleviation and ecosystem services enhancement. Bull. Chin. Acad. Sci. 2020, 35, 1264–1272. [Google Scholar]
- Pei, G.T.; Sun, J.F.; He, T.X.; Hu, B.Q. Effects of long-term human disturbances on soil microbial diversity and community structure in a karst grassland ecosystem of northwestern Guangxi, China. Chin. J. Plant Ecol. 2021, 45, 74–84. [Google Scholar] [CrossRef]
- Liu, M.; Bai, X.Y.; Tan, Q.; Luo, G.J.; Zhao, C.W.; Wu, L.H.; Hu, Z.Y.; Ran, C.; Deng, Y.H. Monitoring impacts of ecological engineering on ecosystem services with geospatial techniques in karst areas of SW China. Geocarto. Int. 2022, 7, 5091–5115. [Google Scholar] [CrossRef]
- Lu, M.; Liu, K.; Zhang, L.; Zeng, F.; Song, T.; Peng, W.; Du, H. Stoichiometric variation in soil carbon, nitrogen, and phosphorus following cropland conversion to forest in Southwest China. Forests 2022, 13, 1155. [Google Scholar] [CrossRef]
- Wen, L.; Li, D.; Xiao, X.; Tang, H.M. Alterations in soil microbial phospholipid fatty acid profile with soil depth following cropland conversion in karst region, southwest China. Environ. Sci. Pollut. Res. 2023, 30, 1502–1519. [Google Scholar] [CrossRef] [PubMed]
- Song, M.; Peng, W.X.; Du, H.; Xu, Q.G. Responses of soil and microbial C:N:P stoichiometry to vegetation succession in a karst region of Southwest China. Forests 2019, 10, 755. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.J.; Peng, W.X.; Du, H.; Song, T.Q.; Zeng, F.P.; Wang, F. Effect of different grain for green approaches on soil bacterial community in a karst region. Front. Microbiol. 2020, 11, 577242. [Google Scholar] [PubMed]
- Xiao, S.Z.; He, J.H.; Zeng, C.; Wang, J.L. Soil chemical properties under various land-use types in the karst area with a case study in Shibing County of China. J. Chem. 2021, 2021, 5523060. [Google Scholar] [CrossRef]
- Lin, X.G. Principles and Methods of Soil Microbiology Research; Higher Education Press: Beijing, China, 2010. [Google Scholar]
- Bradshaw, C.; Kautsky, U.; Kumblad, L. Ecological stoichiometry and multi-element transfer in a coastal ecosystem. Ecosystems 2012, 15, 591–603. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Chen, H.Y.H.; Chang, S.X. Meta-analysis shows that plant mixtures increase soil phosphorus availability and plant productivity in diverse ecosystems. Nat. Ecol. Evol. 2022, 6, 1112–1121. [Google Scholar] [CrossRef]
- Kohmann, M.M.; Silveira, M.L.; da Silva Cardoso, A.; Bracho, R. Short-term impacts of prescribed fire on C, N, and P dynamics in a subtropical rangeland. Plant Soil 2023. [Google Scholar] [CrossRef]
- Dai, M.; Wang, T.; Wang, Y.; Xu, J. Effects of warming and phosphorus enrichment on the C:N:P stoichiometry of Potamogeton crispus organs. Front. Plant Sci. 2022, 13, 814255. [Google Scholar] [CrossRef]
- Taggart, C.B.; Muir, J.P.; Brady, J.A.; Kan, E.; Mitchell, A.B.; Obayomi, O. Impacts of biochar on Trifolium incarnatum and Lolium multiflorum: Soil nutrient retention and loss in sandy loam amended with dairy manure. Agronomy 2023, 13, 26. [Google Scholar] [CrossRef]
- Xu, H.; Qu, Q.; Wang, Z.; Xue, S.; Xu, Z. Plant-soil-enzyme C-N-P stoichiometry and microbial nutrient limitation responses to plant-soil feedbacks during community succession: A 3-year pot experiment in China. Front. Plant Sci. 2022, 13, 1009886. [Google Scholar] [CrossRef]
- Wang, J.; He, K.; Bao, G.; He, L.; Xu, H.; Song, M.; Jin, L.; Wei, X. Effects of increased phosphorus fertilizer on C, N, and P stoichiometry in different organs of bluegrass (Poa L.) at different growth stages. Front. Ecol. Evol. 2023, 11, 1092840. [Google Scholar] [CrossRef]
- Xie, L.L.; Wang, X.L.; Ma, Y.; MA, Y.S.; Wang, Y.L.; Zhou, X.B. Effect of the no-grazing practice in regreening period on the quality and stoichiometric ratio of C, N, P of plant community in alpine meadow. Acta Agrestia Sin. 2023, 31, 1454–1460. [Google Scholar]
- Wang, M.; Zhang, Y.; Li, R.Q.; Xin, X.P.; Zhu, X.Y.; Cao, J.; Zhou, Z.Y.; Yan, R.R. Effects of Grazing Disturbance on the Stoichiometry of Nitrogen and Phosphorus in Plant Organs of Leymus chinensis Meadow Steppe. Sci. Agric. Sin. 2022, 55, 1371–1384. [Google Scholar]
- Tessier, J.T.; Raynal, D.J. Use of nitrogen to phosphorus ratios in plant tissue as an indicator of nutrient limitation and nitrogen saturation. J. Appl. Ecol. 2003, 40, 523–534. [Google Scholar] [CrossRef]
- Güsewell, S. N:P ratios in terrestrial plants: Variation and functional significance. New Phytol 2004, 164, 243–266. [Google Scholar] [CrossRef]
- He, J.S.; Wang, L.; Flynn, D.F.B.; Wang, X.P.; Ma, W.H.; Fang, J.Y. Leaf nitrogen:phosphorus stoichiometry across Chinese grassland biomes. Oecologia 2008, 155, 301–310. [Google Scholar] [CrossRef] [PubMed]
- Song, S.Z. Study on Coupling of Degraded Grassland Improvement and Semi-House Feeding Cattle and Sheep in the Karst Rocky Desertification Area. Master’s Thesis, Guizhou Normal University, Guiyang, China, 2019. [Google Scholar]
- Peco, B.; Navarro, E.; Carmona, C.P.; Medina, N.G.; Marques, M.J. Effects of grazing abandonment on soil multifunctionality: The role of plant functional traits. Agr. Ecosyst. Environ. 2017, 249, 215–225. [Google Scholar] [CrossRef]
- Li, S.Y.; Cui, Y.X.; Sun, Z.J.; Liu, H.X.; Ye, H.W. Effect of grazing exclusion on soil organic carbon and stoichiometry characteristics of soil microbial biomass in sagebrush desert. Acta Pratacul. Sin. 2023, 32, 58–70. [Google Scholar]
- Hali, A.; Sun, Z.J.; He, P.X.; Liu, H.X. Effects of grazing exclusion on soil nitrogen content and its component characteristics in sagebrush desert grassland. J. Soil Wat. Conserv. 2022, 36, 222–230. [Google Scholar]
- Li, W.; Su, T.; Shen, Y.; Ma, H.; Zhou, Y.; Lu, Q.; Wang, G.; Liu, Z.; Li, J. Effects of warming seasonal rotational grazing on plant communities’ structure and diversity in desert steppe. Ecol. Evol. 2023, 13, e9748. [Google Scholar] [CrossRef]
- Chi, Y.K.; Song, S.Z.; Zhao, D.G.; Wu, J.H. Effects of fertilization on plant-soil ecological stoichiometric characteristics of degraded artificial grassland in karst area. Fresen. Environ. Bull. 2022, 31, 10190–10198. [Google Scholar]
- Fan, D.D.; Liu, Y.J.; Cao, H.L.; Chen, H.; Kong, W.D.; Li, X.Z. On the effect of fencing on physicochemical property and microbial community of grassland soils. Sci. Technol. Rev. 2022, 40, 41–51. [Google Scholar]
- Lu, X.; Yan, Y.; Sun, J.; Zhang, X.; Chen, Y.; Wang, X.; Cheng, G. Short-term grazing exclusion has no impact on soil properties and nutrients of degraded alpine grassland in Tibet, China. Solid Earth 2015, 6, 1195–1205. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.L.; Du, G.Z.; Liu, Z.H.; Thirgood, S. Effect of fencing and grazing on a Kobresia-dominated meadow in the QinghaiTibetan Plateau. Plant Soil 2009, 319, 115–126. [Google Scholar] [CrossRef]
- Ahmad, E.H.; Demisie, W.; Zhang, M. Effects of land use on concentrations and chemical forms of phosphorus in different-size aggregates. Eurasian Soil Sci. 2017, 50, 1435–1443. [Google Scholar] [CrossRef]
- Wang, S.Q.; Yu, G.R. Ecological stoichiometry characteristics of ecosystem carbon, nitrogen and phosphorus elements. Acta Ecol. Sin. 2008, 28, 3937–3947. [Google Scholar]
- Shu, M.; Jiang, T.; Wang, D.L.; Lian, Z.; Tang, J.X.; Kong, T.; Xu, Y.Y.; Han, X.M. Soil ecological stoichiometry under the planted of Pinus sylvestris var. mongolica forests with different stand ages in the Horqin Sandy Land. Arid Zone Res. 2018, 35, 789–795. [Google Scholar]
- Tian, H.Q.; Chen, G.S.; Zhang, C.; Melillo, J.M.; Hall, C.A.S. Pattern and variation of C:N:P rations in China’s soils: Synthesis of observational data. Biogeochemistry 2010, 98, 139–151. [Google Scholar] [CrossRef]
- Jia, Y.; Xu, B.C.; Li, F.M.; Wang, X.L. Availability and contributions of soil phosphorus to forage production of seeded alfalfa in semiarid Loess Plateau. Acta Ecol. Sin. 2007, 27, 42–47. [Google Scholar]
- Zhu, Q.L.; Xing, X.Y.; Zhang, H.; An, S.S. Soil ecological stoichiometry under different vegetation area on loess hilly gully region. Acta Ecol. Sin. 2013, 33, 4674–4682. [Google Scholar]
- He, J.; Wei, Q.Q.; Zhong, Y.X.; Luo, L.L. Eco-stoichiometry characteristics of grape fields with different planting years in the east foothills of Helan Mountain. Agric. Res. Arid Areas 2020, 38, 23–30. [Google Scholar]
- Zhao, D.; Li, F.; Yang, Q.; Wang, R.; Song, Y.; Tao, Y. The influence of different types of urban land use on soil microbial biomass and functional diversity in Beijing, China. Soil Use Manag. 2013, 29, 230–239. [Google Scholar] [CrossRef]
- Wu, L.F.; Wang, Z.Q.; Wang, Y.; Liu, Y.G.; Yang, B.; Zhang, Y.F. Relationship between soil C, N, P stoichiometric characteristics and enzyme activity in karst plateau soils with different degree of rocky desertification. Ecol. Environ. Sci. 2019, 28, 2332–2340. [Google Scholar]
- Yang, X.W.; Liu, J.; Hou, M.Y.; Cheng, X.M.; Huang, X.X. Enzyme activities and stoichiometric characteristics of rhizometric and non-rhizosphere soil in different ancient tea gardens in Fengqing County. Chin. J. Appl. Eco. 2023, 29, 1368. [Google Scholar]
- Tian, J.; Sheng, M.Y.; Wang, P.; Wen, P.C. Influence of land use change on litter and soil C, N, P stoichiometric characteristics and soil enzyme activity in karst ecosystem, Southwest China. Environ. Sci. 2019, 40, 4278–4286. [Google Scholar]
- Liu, L.B.; Zhong, Q.L.; Ni, J. Ecosystem C:N:P stoichiometry and storages of a secondary plateau-surface karst forest in Guizhou Province, southwestern China. Acta Ecol. Sin. 2019, 39, 8606–8614. [Google Scholar]
- Wan, F.; Meng, Z.J.; Dang, X.H.; Wang, R.D.; Zhang, H.M. C, N and P ecological stoichiometry characteristics of a Stipa species plant–soil system subject to grazing exclusion in a desert steppe. Acta Pratacul. Sin. 2020, 29, 49–55. [Google Scholar]
- Zhang, J.H.; Li, M.X.; Xu, L.; Zhu, J.X.; Dai, G.H.; He, N.P. C:N:P stoichiometry in terrestrial ecosystems in China. Sci. Total Environ. 2021, 795, 148849. [Google Scholar] [CrossRef]
- Arunachalam, A.; Pandey, H.N. Ecosystem restoration of jhum fallows in northeast india: Microbial C and N along altitudinal and successional gradients. Restor. Ecol. 2003, 11, 168–173. [Google Scholar] [CrossRef]
- Manzoni, S.; Trofymow, J.A.; Jackson, R.B.; Porporato, A. Stoichiometric controls on carbon, nitrogen, and phosphorus dynamics in decomposing litter. Ecol. Monogr. 2010, 80, 89–106. [Google Scholar] [CrossRef] [Green Version]
- Mooshammer, M.; Wanek, W.; Zechmeister-Boltenstern, S.; Richter, A. Stoichiometric imbalances between terrestrial decomposer communities and their resources: Mechanisms and implications of microbial adaptations to their resources. Front. Microbiol. 2014, 5, 22. [Google Scholar] [CrossRef] [Green Version]
- Heuch, C.; Weig, A.; Spohn, M. Soil microbial biomass C:N:P stoichiometry and microbial use of organic phosphorus. Soil Biol. Biochem. 2015, 85, 119–129. [Google Scholar] [CrossRef]
- Li, J.W.; Liu, Y.L.; Hai, X.Y.; Shangguan, Z.P.; Deng, L. Dynamics of soil microbial C:N:P stoichiometry and its driving mechanisms following natural vegetation restoration after farmland abandonment. Sci. Total. Environ. 2019, 693, 133613. [Google Scholar] [CrossRef] [PubMed]
- Tischer, A.; Potthast, K.; Hamer, U. Land-use and soil depth affect resource and microbial stoichiometry in a tropical mountain rainforest region of southern Ecuador. Oecologia 2014, 175, 375–393. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Fang, F.; Tang, H. Patterns and internal stability of carbon, nitrogen, and phosphorus in soils and soil microbial biomass in terrestrial ecosystems in China: A Data Synthesis. Forests 2021, 12, 1544. [Google Scholar] [CrossRef]
- Hartman, W.H.; Richardson, C.J. Differential nutrient limitation of soil microbial biomass and metabolic quotients(qCO2): Is there a biological stoichiometry of soil microbes. PLoS ONE 2013, 8, 57127. [Google Scholar] [CrossRef]
- González-Chávez, M.C.A.; Aitkenhead-Peterson, J.A.; Gentry, T.J.; Zuberer, D.; Hons, F.; Loeppert, R. Soil microbial community, C, N, and P responses to long-term tillage and crop rotation. Soil Till. Res. 2016, 106, 285–293. [Google Scholar] [CrossRef]
- Wang, B.R.; Yang, J.J.; An, S.S.; Zhang, H.X.; Bai, X.J. Effects of vegetation and topography features on ecological stoichiometry of soil and soil microbial biomass in the hilly-gully region of the Loess Plateau, China. Chin. J. Appl. Ecol. 2018, 29, 247–259. [Google Scholar]
- Yu, Y.F.; Peng, W.X.; Song, T.Q.; Zeng, F.P.; Wang, K.L.; Wen, L.; Fan, F.J. Stoichiometric characteristics of plant and soil C, N and P in different forest types in depressions between karst hills, southwest China. Chin. J. Appl. Ecol. 2014, 25, 947–954. [Google Scholar]
- Peng, P.Q.; Zhang, W.J.; Tong, C.L.; Qiu, S.J.; Zhang, W.C. Soil C, N and P contents and their relationships with soil physical properties in wetlands of Dongting Lake floodplain. Chin. J. Appl. Ecol. 2005, 16, 1872–1878. [Google Scholar]


{kind=link}
| Utilization Methods | C/(g/kg) | N/(g/kg) | P/(g/kg) | C:N | C:P | N:P |
|---|---|---|---|---|---|---|
| Grazing grassland | 429.56 ± 9.92 b | 21.47 ± 0.16 a | 0.81 ± 0.08 a | 20.01 ± 0.61 c | 531.74 ± 42.68 b | 26.61 ± 2.62 b |
| Mowing grassland | 406.70 ± 5.25 c | 18.54 ± 0.13 bc | 0.87 ± 0.10 a | 21.93 ± 0.22 b | 474.63 ± 61.02 bc | 21.63 ± 2.67 bc |
| Enclosed grassland | 472.45 ± 12.90 a | 19.79 ± 0.03 b | 0.42 ± 0.02 b | 23.88 ± 0.68 a | 1133.09 ± 40.40 a | 47.49 ± 2.38 a |
| Utilization Methods | C/(g/kg) | N/(g/kg) | P/(g/kg) | C:N | C:P | N:P |
|---|---|---|---|---|---|---|
| Grazing grassland | 19.48 ± 1.12 ab | 2.19 ± 0.09 a | 0.85 ± 0.05 b | 8.91 ± 0.97 c | 22.78 ± 0.83 b | 2.58 ± 0.36 b |
| Mowing grassland | 17.37 ± 0.42 b | 1.74 ± 0.10 bc | 0.53 ± 0.01 c | 10.04 ± 0.67 b | 32.88 ± 0.79 a | 3.29 ± 0.27 a |
| Enclosed grassland | 22.45 ± 1.39 a | 1.83 ± 0.09 b | 1.26 ± 0.020 a | 12.31 ± 1.91 a | 17.91 ± 2.07 c | 1.46 ± 0.08 c |
| Utilization Methods | MBC (mg/kg) | MBN (mg/kg) | MBP (mg/kg) | MBC:MBN | MBC:MBP | MBN:MBP |
|---|---|---|---|---|---|---|
| Grazing grassland | 350.24 ± 37.56 a | 142.46 ± 10.45 a | 10.95 ± 37.56 b | 2.48 ± 0.41 bc | 32.34 ± 5.93 b | 13.11 ± 1.81 a |
| Mowing grassland | 315.61 ± 31.44 ab | 81.53 ± 7.23 c | 7.50 ± 0.20 c | 3.88 ± 0.32 a | 42.09 ± 3.98 a | 10.86 ± 0.71 a |
| Enclosed grassland | 270.89 ± 11.44 b | 96.93 ± 10.14 b | 18.82 ± 0.65 a | 2.81 ± 0.19 b | 14.39 ± 0.20 c | 5.14 ± 0.37 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pan, G.; Song, S.; Wang, X.; Chi, Y. Using Ecological Stoichimetric Characteristies to Inform Grassland Management in the Karst Desertification Area. Agronomy 2023, 13, 1841. https://doi.org/10.3390/agronomy13071841
Pan G, Song S, Wang X, Chi Y. Using Ecological Stoichimetric Characteristies to Inform Grassland Management in the Karst Desertification Area. Agronomy. 2023; 13(7):1841. https://doi.org/10.3390/agronomy13071841
Chicago/Turabian StylePan, Guochang, Shuzhen Song, Xueling Wang, and Yongkuan Chi. 2023. "Using Ecological Stoichimetric Characteristies to Inform Grassland Management in the Karst Desertification Area" Agronomy 13, no. 7: 1841. https://doi.org/10.3390/agronomy13071841