
Evidence of Bi-Directional Volatile-Mediated Communication between Drought-Stressed and Well-Watered Grapevines (Vitis vinifera L.)

 ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Treatments
2.2. Growth and Stress Conditions
2.3. Physiological Measurements
2.4. VOC Metabolite Analysis
2.4.1. Chemicals and VOC Sampling Materials
2.4.2. GC-MS Instrumentation
2.5. Nucleic Acid Analysis
2.5.1. RNA Extraction
2.5.2. cDNA Synthesis and PCR
2.5.3. Quantitative Real-Time PCR

{kind=link}
{kind=link}
{kind=link}
| Gene Name/ID | Forward Primer Sequence | Reverse Primer Sequence | References |
|---|---|---|---|
| Allene synthase (AOS) | CTTCGGAGGGATGAAAATCA | GGCTAATTGGGTGTGCAGT | [36] |
| Chorismate synthase (CHORS) | GCCTTCACATGCAGATGCTA | CTGCAACTCTCCCAATGGTT | [37] |
| Lipoxygenase (VvLOXD) | ACCCACCAAATCGTCCCACACTATG | ACCTCTTCGTTGTCTGTCCACTCTG | [38] |
| α-Pinene (VvPNaPin1) | TTGGAGAAGCTTAAGGGAGATG | GGTAGCCATGCTGTCTTAGGAG | [39] |
2.6. Statistical Analysis
3. Results
3.1. Physiological Analysis
3.2. Investigating VOC Signalling Using Volatile Metabolite and Transcript Analyses
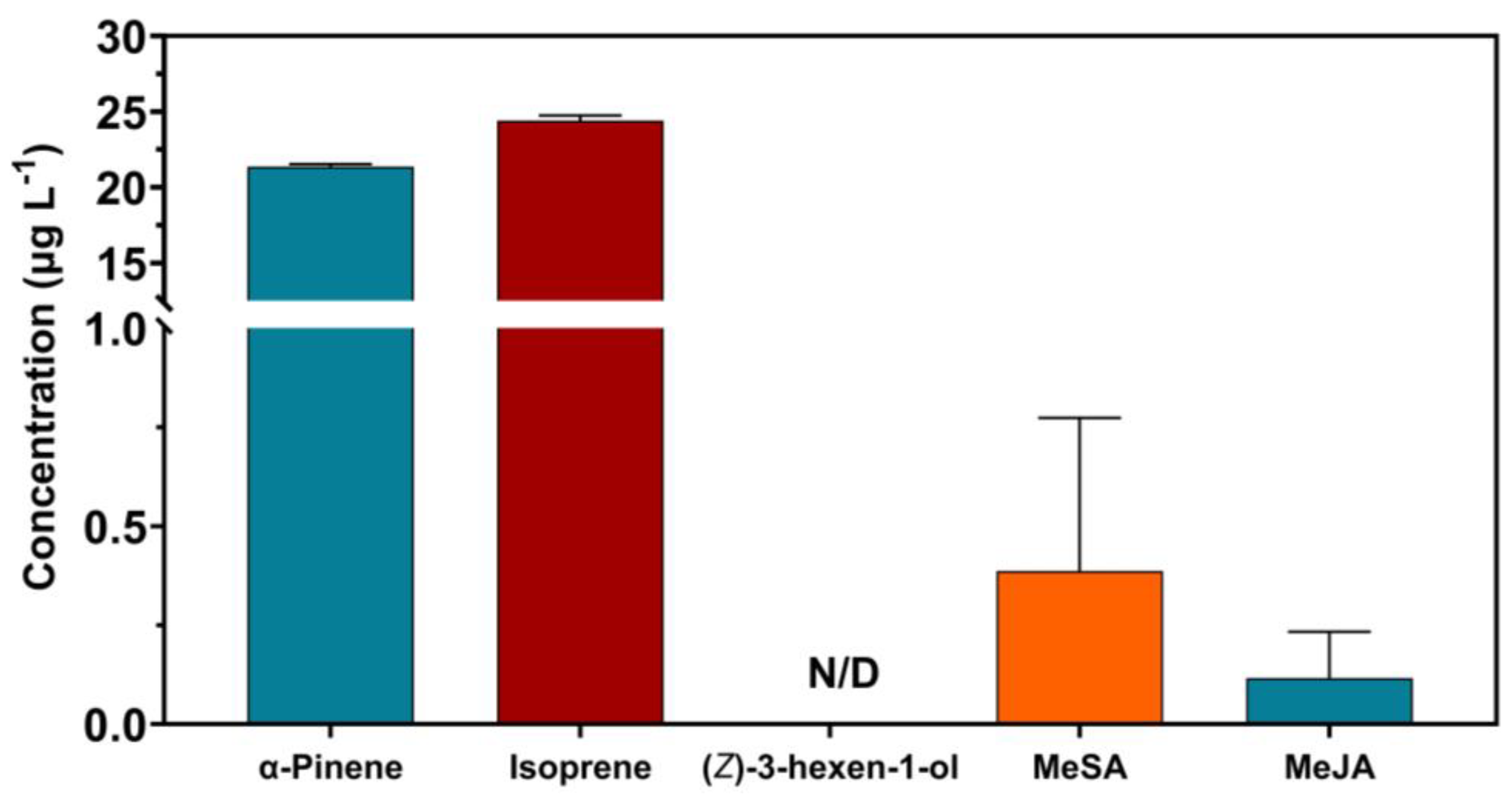
3.2.1. Constitutive VOC Emissions
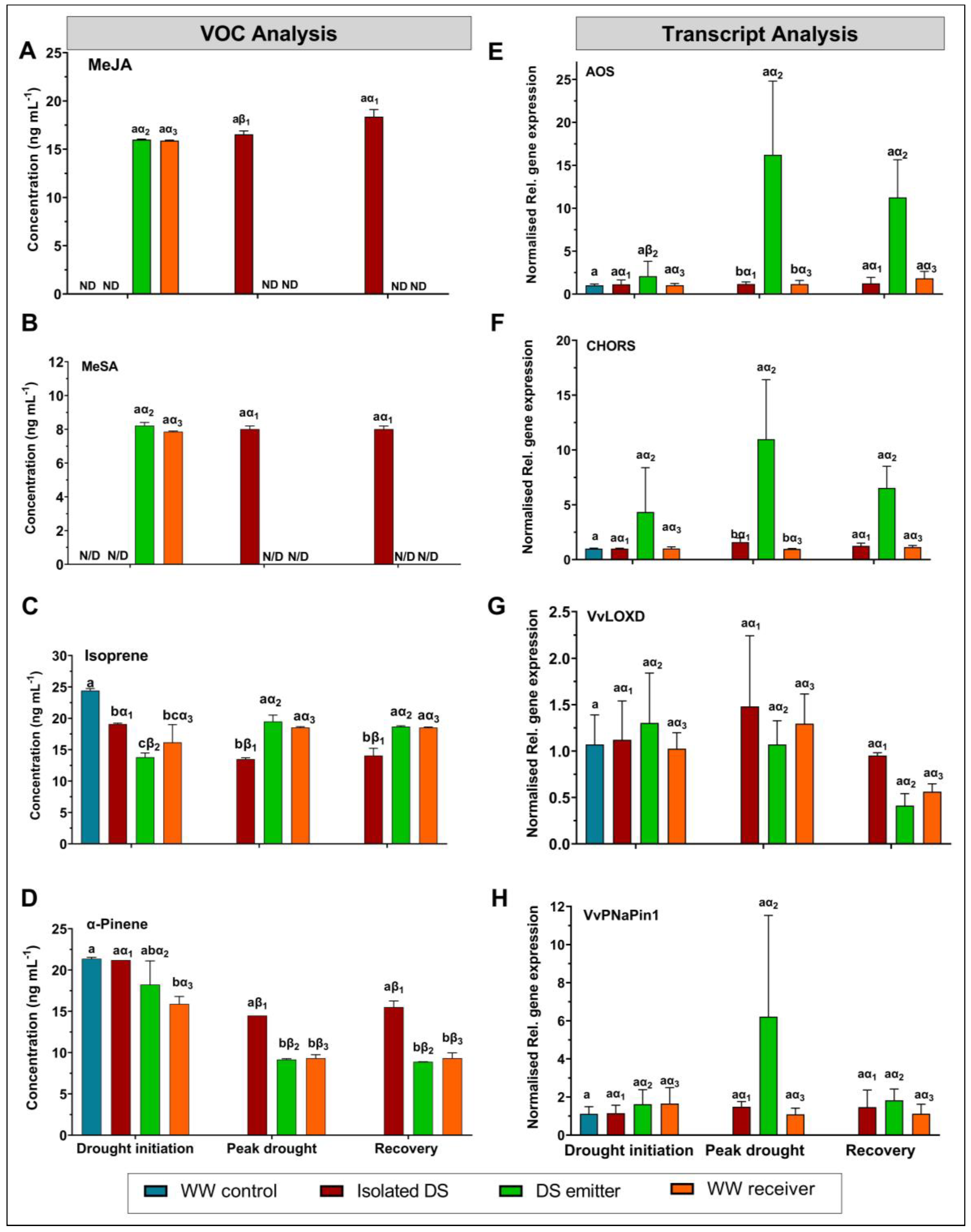
3.2.2. Synchronised Emission of MeJA, MeSA, α-Pinene, and Isoprene in Co-Located Treatment Vines
4. Discussion
4.1. Effect of Drought-Induced VOCs on the gs of Co-Located WWr
4.2. Volatile Metabolite and Gene Transcript Analyses
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dudareva, N.; Negre, F.; Nagegowda, D.A.; Orlova, I. Plant volatiles: Recent advances and future perspectives. Crit. Rev. Plant Sci. 2006, 25, 417–440. [Google Scholar] [CrossRef]
- Heil, M.; Silva Bueno, J.C. Within-plant signalling by volatiles leads to induction and priming of an indirect plant defence in nature. Proc. Natl. Acad. Sci. USA 2007, 104, 5467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niinemets, Ü.; Arneth, A.; Kuhn, U.; Monson, R.K.; Peñuelas, J.; Staudt, M. The emission factor of volatile isoprenoids: Stress, acclimation, and developmental responses. Biogeosciences 2010, 7, 2203–2223. [Google Scholar] [CrossRef] [Green Version]
- Loreto, F.; Schnitzler, J.P. Abiotic stresses and induced BVOCs. Trends Plant Sci. 2010, 15, 154–166. [Google Scholar] [CrossRef]
- Weingart, G.; Kluger, B.; Forneck, A.; Krska, R.; Schuhmacher, R. Establishment and application of a metabolomics workflow for identification and profiling of volatiles from leaves of Vitis vinifera by HS-SPME-GC-MS. Phytochem. Anal. 2012, 23, 345–358. [Google Scholar] [CrossRef]
- Niinemets, Ü.; Kännaste, A.; Copolovici, L. Quantitative patterns between plant volatile emissions induced by biotic stresses and the degree of damage. Front. Plant Sci. 2013, 4, 262. [Google Scholar] [CrossRef] [Green Version]
- Zhou, S.; Jander, G. Molecular ecology of plant volatiles in interactions with insect herbivores. J. Exp. Bot. 2021, 73, 449–462. [Google Scholar] [CrossRef]
- Brosset, A.; Blande, J.D. Volatile-mediated plant–plant interactions: Volatile organic compounds as modulators of receiver plant defence, growth, and reproduction. J. Exp. Bot. 2022, 73, 511–528. [Google Scholar] [CrossRef]
- Pierik, R.; BallarÉ, C.L.; Dicke, M. Ecology of plant volatiles: Taking a plant community perspective. Plant Cell Environ. 2014, 37, 1845–1853. [Google Scholar] [CrossRef]
- Midzi, J.; Jeffery, D.W.; Baumann, U.; Rogiers, S.; Tyerman, S.D.; Pagay, V. Stress-induced volatile emissions and signalling in inter-plant communication. Plants 2022, 11, 2566. [Google Scholar] [CrossRef]
- Arimura, G.-I.; Ozawa, R.; Nishioka, T.; Boland, W.; Koch, T.; Kühnemann, F.; Takabayashi, J. Herbivore-induced volatiles induce the emission of ethylene in neighbouring lima bean plants. Plant J. 2002, 29, 87–98. [Google Scholar] [CrossRef]
- Engelberth, J.; Alborn, H.T.; Schmelz, E.A.; Tumlinson, J.H. Airborne signals prime plants against insect herbivore attack. Proc. Natl. Acad. Sci. USA 2004, 101, 1781. [Google Scholar] [CrossRef] [Green Version]
- Karban, R.; Baldwin, I.T.; Baxter, K.J.; Laue, G.; Felton, G.W. Communication between plants: Induced resistance in wild tobacco plants following clipping of neighbouring sagebrush. Oecologia 2000, 125, 66–71. [Google Scholar] [CrossRef]
- Kessler, A.; Halitschke, R.; Diezel, C.; Baldwin, I. Priming of plant defence responses in nature by airborne signalling between Artemisia tridentata and Nicotiana attenuata. Oecologia 2006, 148, 280–292. [Google Scholar] [CrossRef]
- Sugimoto, K.; Matsui, K.; Iijima, Y.; Akakabe, Y.; Muramoto, S.; Ozawa, R.; Uefune, M.; Sasaki, R.; Alamgir, K.M.; Akitake, S.; et al. Intake and transformation to a glycoside of (Z)-3-hexenol from infested neighbours reveals a mode of plant odour reception and defence. Proc. Natl. Acad. Sci. USA 2014, 111, 7144–7149. [Google Scholar] [CrossRef] [Green Version]
- Ton, J.; D’Alessandro, M.; Jourdie, V.; Jakab, G.; Karlen, D.; Held, M.; Mauch-Mani, B.; Turlings, T.C.J. Priming by airborne signals boosts direct and indirect resistance in maize. Plant J. 2007, 49, 16–26. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.; Seo, P.J. Airborne signals from salt-stressed Arabidopsis plants trigger salinity tolerance in neighbouring plants. Plant Signal. Behav. 2014, 9, e28392. [Google Scholar] [CrossRef] [Green Version]
- Caparrotta, S.; Boni, S.; Taiti, C.; Palm, E.; Mancuso, S.; Pandolfi, C. Induction of priming by salt stress in neighbouring plants. Environ. Exp. Bot. 2018, 147, 261–270. [Google Scholar] [CrossRef]
- Zhao, M.; Wang, L.; Wang, J.; Jin, J.; Zhang, N.; Lei, L.; Gao, T.; Jing, T.; Zhang, S.; Wu, Y. Induction of priming by cold stress via inducible volatile cues in neighbouring tea plants. J. Integr. Plant Biol. 2020, 62, 1461. [Google Scholar] [CrossRef]
- Landi, M.; Araniti, F.; Flamini, G.; Lo Piccolo, E.; Trivellini, A.; Abenavoli, M.; Guidi, L. “Help is in the air”: Volatiles from salt-stressed plants increase the reproductive success of receivers under salinity. Planta 2020, 251, 48. [Google Scholar] [CrossRef]
- Jin, J.; Zhao, M.; Gao, T.; Jing, T.; Zhang, N.; Wang, J.; Zhang, X.; Huang, J.; Schwab, W.; Song, C. Amplification of early drought responses caused by volatile cues emitted from neighbouring tea plants. Hort. Res. 2021, 8, 243. [Google Scholar] [CrossRef] [PubMed]
- Tscharntke, T.; Thiessen, S.; Dolch, R.; Boland, W. Herbivory, induced resistance, and interplant signal transfer in Alnus glutinosa. Biochem. Syst. Ecol. 2001, 29, 1025–1047. [Google Scholar] [CrossRef]
- Farmer, E.E.; Ryan, C.A. Interplant communication: Airborne methyl jasmonate induces synthesis of proteinase inhibitors in plant leaves. Proc. Natl. Acad. Sci. USA 1990, 87, 7713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shulaev, V.; Silverman, P.; Raskin, I. Airborne signalling by methyl salicylate in plant pathogen resistance. Nature 1997, 385, 718–721. [Google Scholar] [CrossRef]
- Rai, V.K.; Gupta, S.C.; Singh, B. Volatile monoterpenes from Prinsepia utilis L. leaves inhibit stomatal opening in Vicia faba L. Biol. Plant. 2003, 46, 121–124. [Google Scholar] [CrossRef]
- Jiang, Y.; Ye, J.; Rasulov, B.; Niinemets, Ü. Role of stomatal conductance in modifying the dose response of stress-volatile emissions in methyl jasmonate treated leaves of cucumber (Cucumis sativa). Int. J. Mol. Sci. 2020, 21, 1018. [Google Scholar] [CrossRef] [Green Version]
- Arévalo-Marín, D.F.; Briceño-Robles, D.M.; Mosquera, T.; Melgarejo, L.M.; Sarmiento, F. Jasmonic acid priming of potato uses hypersensitive response-dependent defense and delays necrotrophic phase change against Phytophthora infestans. Physiol. Mol. Plant Pathol. 2021, 115, 101680. [Google Scholar] [CrossRef]
- Zamora, O.; Schulze, S.; Azoulay-Shemer, T.; Parik, H.; Unt, J.; Brosché, M.; Schroeder, J.I.; Yarmolinsky, D.; Kollist, H. Jasmonic acid and salicylic acid play minor roles in stomatal regulation by CO2, abscisic acid, darkness, vapor pressure deficit and ozone. Plant J. 2021, 108, 134–150. [Google Scholar] [CrossRef]
- López-Gresa, M.P.; Payá, C.; Ozáez, M.; Rodrigo, I.; Conejero, V.; Klee, H.; Bellés, J.M.; Lisón, P. A new role for green leaf volatile esters in tomato stomatal defense against Pseudomonas syringe pv. tomato. Front. Plant Sci. 2018, 9, 01855. [Google Scholar] [CrossRef]
- Dayer, S.; Tyerman, S.; Garnett, T.; Pagay, V. Relationship between hydraulic and stomatal conductance and its regulation by root and leaf aquaporins under progressive water stress and recovery and exogenous application of ABA in Vitis vinifera L. ‘Syrah’. Acta Hortic. 2017, 1188, 227–234. [Google Scholar] [CrossRef]
- Himanen, S.J.; Blande, J.D.; Klemola, T.; Pulkkinen, J.; Heijari, J.; Holopainen, J.K. Birch (Betula spp.) leaves adsorb and re-release volatiles specific to neighbouring plants: A mechanism for associational herbivore resistance? New Phytol. 2010, 186, 722–732. [Google Scholar] [CrossRef]
- Ormeño, E.; Viros, J.; Mévy, J.-P.; Tonetto, A.; Saunier, A.; Bousquet-Mélou, A.; Fernandez, C. Exogenous isoprene confers physiological benefits in a negligible isoprene emitter (Acer monspessulanum L.) under water deficit. Plants 2020, 9, 159. [Google Scholar] [CrossRef] [Green Version]
- Murray, F.W. On the computation of saturation vapor pressure. J. Appl. Meteorol. Climatol. 1967, 6, 203–204. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, research0034.1. [Google Scholar] [CrossRef] [Green Version]
- Dumin, W.; Rostas, M.; Winefield, C. Identification and functional characterisation of an allene oxide synthase from grapevine (Vitis vinifera L. Sauvignon blanc). Mol. Biol. Rep. 2018, 45, 263–277. [Google Scholar] [CrossRef]
- Dufour, M.C.; Lambert, C.; Bouscaut, J.; Mérillon, J.M.; Corio-Costet, M.F. Benzothiadiazole-primed defence responses and enhanced differential expression of defence genes in Vitis vinifera infected with biotrophic pathogens Erysiphe necator and Plasmopara viticola. Plant Pathol. 2013, 62, 370–382. [Google Scholar] [CrossRef]
- Podolyan, A.; White, J.; Jordan, B.; Winefield, C. Identification of the lipoxygenase gene family from Vitis vinifera and biochemical characterisation of two 13-lipoxygenases expressed in grape berries of Sauvignon Blanc. Funct. Plant Biol. 2010, 37, 767–784. [Google Scholar] [CrossRef]
- Matarese, F.; Scalabrelli, G.; D’Onofrio, C. Analysis of the expression of terpene synthase genes in relation to aroma content in two aromatic Vitis vinifera varieties. Funct. Plant Biol. 2013, 40, 552–565. [Google Scholar] [CrossRef]
- Balacey, S.L. Investigating the Role of Volatile Signalling in Plant Responses to Drought. Ph.D Thesis, The University of Adelaide, Adelaide, Australia, 2021. [Google Scholar]
- Zhang, J.; Davies, W.J. Changes in the concentration of ABA in xylem sap as a function of changing soil water status can account for changes in leaf conductance and growth. Plant Cell Environ. 1990, 13, 277–285. [Google Scholar] [CrossRef]
- Cai, Y.; Wang, J.; Li, S.; Zhang, L.; Peng, L.; Xie, W.; Liu, F. Photosynthetic response of an alpine plant, Rhododendron delavayi Franch, to water stress and recovery: The role of mesophyll conductance. Front. Plant Sci. 2015, 6, 1089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bourtsoukidis, E.; Schneider, H.; Radacki, D.; Schütz, S.; Hakola, H.; Hellén, H.; Noe, S.; Mölder, I.; Ammer, C.; Bonn, B. Impact of flooding and drought conditions on the emission of volatile organic compounds of Quercus robur and Prunus serotina. Trees 2013, 28, 193–204. [Google Scholar] [CrossRef]
- Jubany-Marí, T.; Munné-Bosch, S.; Alegre, L. Redox regulation of water stress responses in field-grown plants. Role of hydrogen peroxide and ascorbate. Plant Physiol. Biochem. 2010, 48, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Jardine, K.J.; Chambers, J.Q.; Holm, J.; Jardine, A.B.; Fontes, C.G.; Zorzanelli, R.F.; Meyers, K.T.; De Souza, V.F.; Garcia, S.; Gimenez, B.O.; et al. Green leaf volatile emissions during high temperature and drought stress in a Central Amazon rainforest. Plants 2015, 4, 678–690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takayama, K.; Jansen, R.M.C.; Verstappen, F.W.A.; Bouwmeester, H.J. Monitoring of lipoxygenase-related plant emission for early detection of drought stress in greenhouse. In Proceedings of the XVI Congress of the Federation of European Societies of Plant Biolology, Tampere, Finland, 22–28 August 2008. [Google Scholar]
- Rering, C.C.; Franco, J.G.; Yeater, K.M.; Mallinger, R.E. Drought stress alters floral volatiles and reduces floral rewards, pollinator activity, and seed set in a global plant. Ecosphere 2020, 11, e03254. [Google Scholar] [CrossRef]
- Pag, A.; Bodescu, A.; Astrid, K.; Tomescu, D.; Niinemets, Ã.; Copolovici, L. Volatile organic compounds emission from Betula verrucosa under drought stress. Sci. Bull. (Escorina) 2013, 8, 45–53. [Google Scholar]
- Niinemets, Ü.; Reichstein, M. Controls on the emission of plant volatiles through stomata: Differential sensitivity of emission rates to stomatal closure explained. J. Geophys. Res. 2003, 108, D7. [Google Scholar] [CrossRef]
- Fall, R.; Monson, R.K. Isoprene emission rate and intercellular isoprene concentration as influenced by stomatal distribution and conductance. Plant Physiol. 1992, 100, 987–992. [Google Scholar] [CrossRef] [Green Version]
- Loreto, F.; Ciccioli, P.; Cecinato, A.; Brancaleoni, E.; Frattoni, M.; Tricoli, D. Influence of environmental factors and air composition on the emission of [alpha]-pinene from Quercus ilex Leaves. Plant Physiol. 1996, 110, 267–275. [Google Scholar] [CrossRef] [Green Version]
- Sharkey, T.D.; Loreto, F. Water stress, temperature, and light effects on the capacity for isoprene emission and photosynthesis of kudzu leaves. Oecologia 1993, 95, 328–333. [Google Scholar] [CrossRef]
- Velikova, V. On the relationship between isoprene emission and thermotolerance in Phragmites australis leaves exposed to high temperatures and during the recovery from a heat stress. Plant Cell Environ. 2005, 28, 318–327. [Google Scholar] [CrossRef]
- Peñuelas, J.; Filella, I.; Seco, R.; Llusià, J. Increase in isoprene and monoterpene emissions after re-watering of droughted Quercus ilex seedlings. Biol. Plant. 2009, 53, 351–354. [Google Scholar] [CrossRef]
- Niinemets, Ü.; Fares, S.; Harley, P.; Jardine, K.J. Bidirectional exchange of biogenic volatiles with vegetation: Emission sources, reactions, breakdown and deposition. Plant Cell Environ. 2014, 37, 1790–1809. [Google Scholar] [CrossRef] [Green Version]
- Noe, S.M.; Copolovici, L.; Niinemets, U.; Vaino, E. Foliar limonene uptake scales positively with leaf lipid content: “Non-emitting” species absorb and release monoterpenes. Plant Biol. 2008, 10, 129–137. [Google Scholar] [CrossRef]
- Root, R.B. Organization of a plant-arthropod association in simple and diverse habitats: The fauna of collards (Brassica oleracea). Ecol. Monogr. 1973, 43, 95–124. [Google Scholar] [CrossRef]
- Tahvanainen, J.O.; Root, R.B. The influence of vegetational diversity on the population ecology of a specialized herbivore, Phyllotreta cruciferae (Coleoptera: Chrysomelidae). Oecologia 1972, 10, 321–346. [Google Scholar] [CrossRef]
- Vickers, C.E.; Gershenzon, J.; Lerdau, M.T.; Loreto, F. A unified mechanism of action for volatile isoprenoids in plant abiotic stress. Nat. Chem. Biol. 2009, 5, 283–291. [Google Scholar] [CrossRef]
- Zuo, Z.; Weraduwage, S.M.; Lantz, A.T.; Sanchez, L.M.; Weise, S.E.; Wang, J.; Childs, K.L.; Sharkey, T.D. Isoprene acts as a signalling molecule in gene networks important for stress responses and plant growth. Plant Physiol. 2019, 180, 124–152. [Google Scholar] [CrossRef] [Green Version]
- Martin, D.M.; Aubourg, S.; Schouwey, M.B.; Daviet, L.; Schalk, M.; Toub, O.; Lund, S.T.; Bohlmann, J. Functional annotation, genome organization and phylogeny of the grapevine (Vitis vinifera) terpene synthase gene family based on genome assembly, FLcDNA cloning, and enzyme assays. BMC Plant Biol. 2010, 10, 226. [Google Scholar] [CrossRef] [Green Version]



| Treatments | gs (mmol H2O m−2 s−1) | Soil Moisture (% VWC) | ||||
|---|---|---|---|---|---|---|
| Day 1 | Day 8 | Day 9 | Day 1 | Day 8 | Day 9 | |
| WWc | 121 ± 25.5 a | - | - | 21 ± 0.07 a | - | - |
| DSc | 91 ± 8.3 aα | 33 ± 3.9 bβ | 70 ± 8.3 aα | 21 ± 0.1 aα | 4 ± 0.1 aβ | 23 ± 0.6 aα |
| DSe | 146 ± 31.7 aα | 33 ± 2.4 bβ | 63 ± 10.9 aα | 24 ± 3.1 aα | 4 ± 0.3 aβ | 24 ± 2.9 aα |
| WWr | 116 ± 19.9 aα | 100 ± 24.79 aα | 115 ± 24.0 aα | 23 ± 0.9 aα | 25 ± 1.5 aα | 25 ± 1.5 aα |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Midzi, J.; Jeffery, D.W.; Baumann, U.; Capone, D.L.; Rogiers, S.Y.; Pagay, V. Evidence of Bi-Directional Volatile-Mediated Communication between Drought-Stressed and Well-Watered Grapevines (Vitis vinifera L.). Agronomy 2023, 13, 1747. https://doi.org/10.3390/agronomy13071747
Midzi J, Jeffery DW, Baumann U, Capone DL, Rogiers SY, Pagay V. Evidence of Bi-Directional Volatile-Mediated Communication between Drought-Stressed and Well-Watered Grapevines (Vitis vinifera L.). Agronomy. 2023; 13(7):1747. https://doi.org/10.3390/agronomy13071747
Chicago/Turabian StyleMidzi, Joanah, David W. Jeffery, Ute Baumann, Dimitra L. Capone, Suzy Y. Rogiers, and Vinay Pagay. 2023. "Evidence of Bi-Directional Volatile-Mediated Communication between Drought-Stressed and Well-Watered Grapevines (Vitis vinifera L.)" Agronomy 13, no. 7: 1747. https://doi.org/10.3390/agronomy13071747