
Nitrogen Application Alleviates Impairments for Jatropha curcas L. Seedling Growth under Salinity Stress by Regulating Photosynthesis and Antioxidant Enzyme Activity
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Site Descriptions
2.2. Experimental Design
2.3. Data Collection
2.3.1. Plant Biomass and Leaf Area Measurements
2.3.2. Antioxidant Enzyme Activity Measurements
2.3.3. Photosynthetic Pigment Contents Measurements
2.3.4. Photosynthetic Characteristics and Chlorophyll Fluorescence Measurement
2.4. Statistical Analysis
3. Results
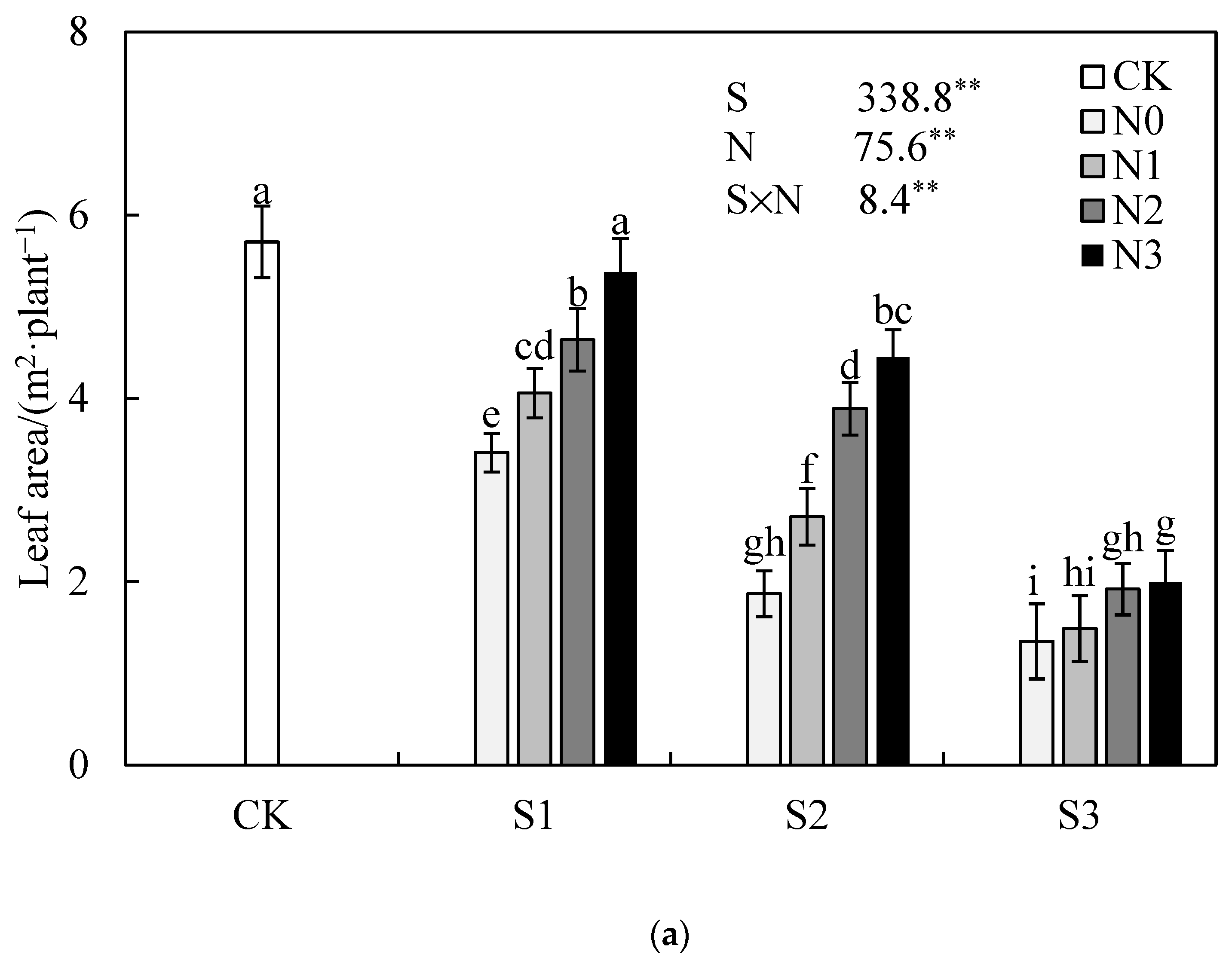
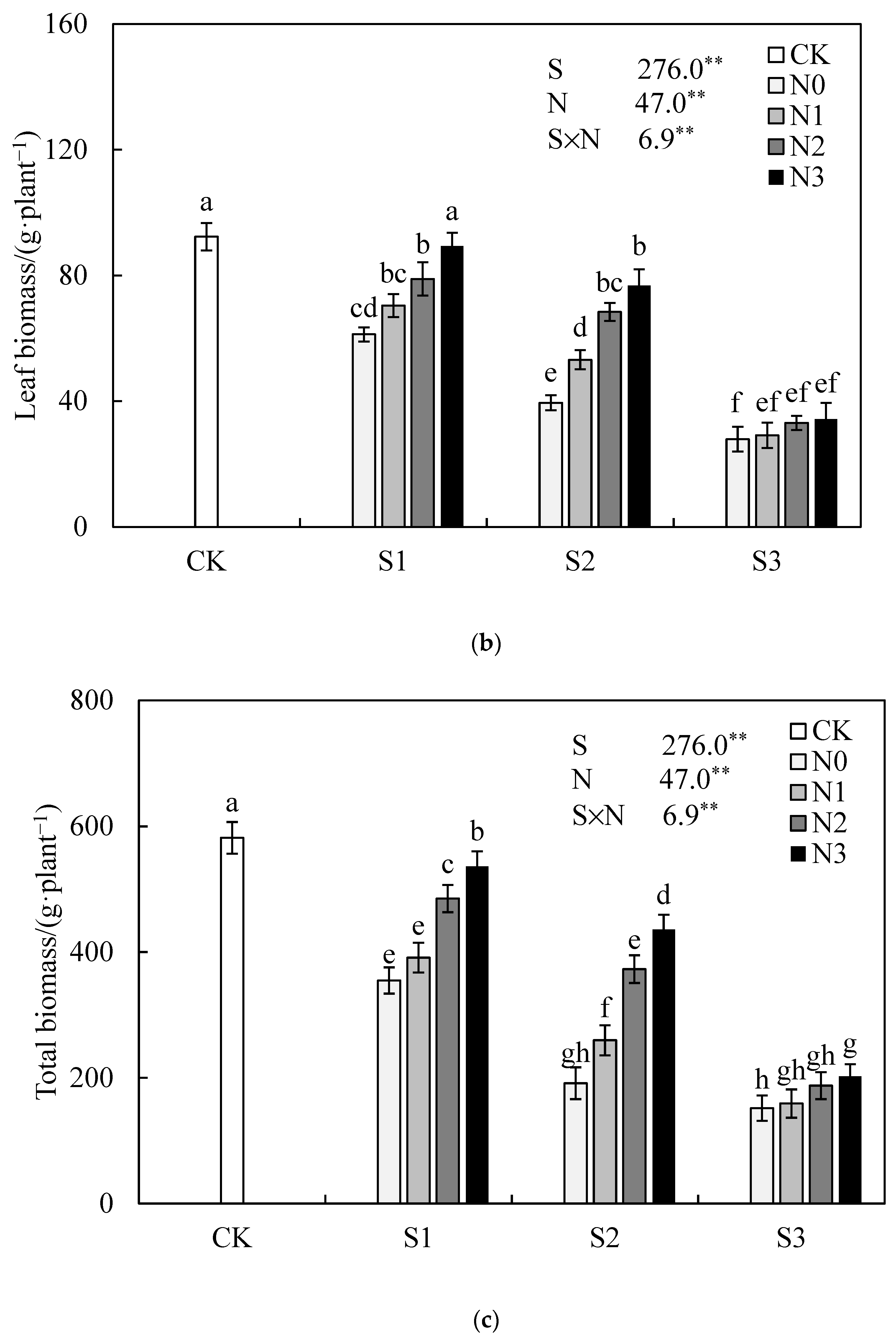
3.1. Effect of Nitrogen Application Rate on Jatropha curcas L. Growth under Salinity Stress
3.2. Effect of Nitrogen Application Rate on Antioxidant Enzyme Activity of Jatropha curcas L. Seedlings under Salinity Stress
3.3. Effect of Nitrogen Application Rate on the Photosynthetic Pigment of Jatropha curcas L. Seedlings under Salinity Stress
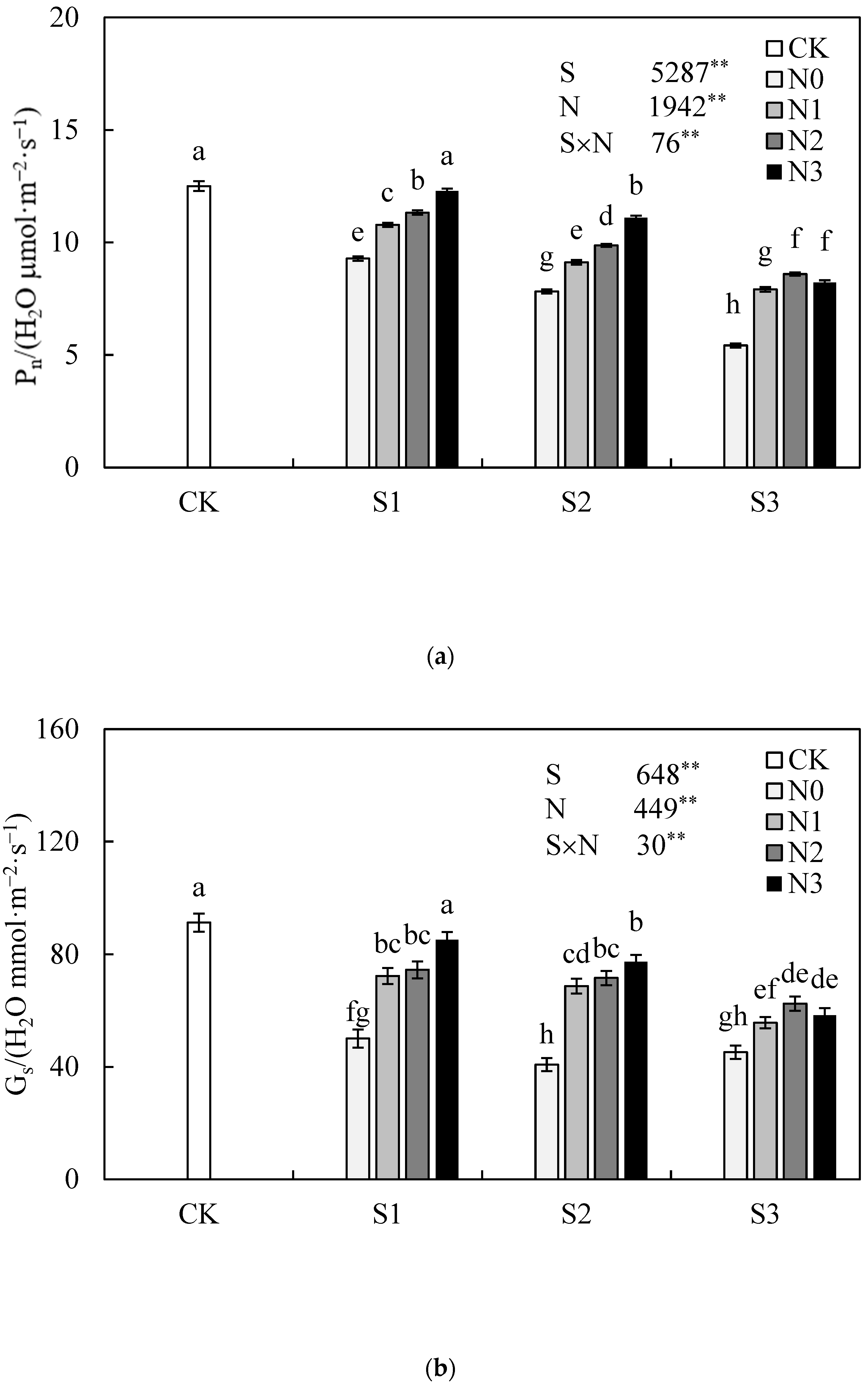
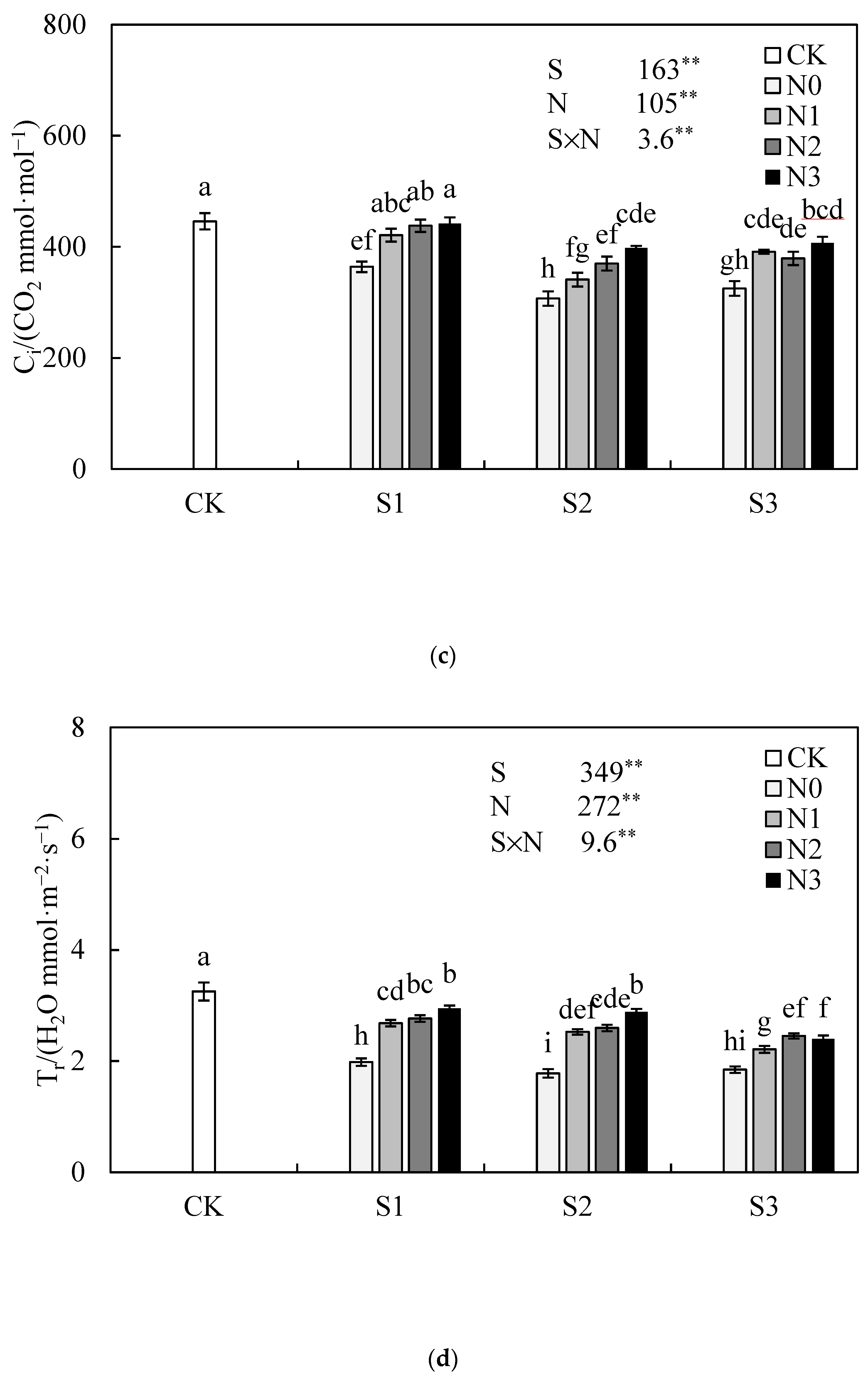
3.4. Effect of Nitrogen Application Rate on Leaf Photosynthetic Characteristic of Jatropha curcas L. Seedlings under Salinity Stress
3.5. Effect of Nitrogen Application Rate on Chlorophyll Fluorescence of Jatropha curcas L. Seedlings under Salinity Stress
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Lozano-Isla, F.; Campos, M.L.; Endres, L.; Bezerra-Neto, E.; Pompelli, M.F. Effects of seed storage time and salt stress on the germination of Jatropha curcas L. Ind. Crops Prod. 2018, 118, 214–224. [Google Scholar] [CrossRef]
- Corte-Real, N.; Endres, L.; Santos, K.; Figueirêdo, R.; Arruda, E.; Ulisses, C.; Pompelli, M.; Figueirêdo, R.; Pompelli, M. Morphoanatomy and ontogeny of the fruit and seeds of Jatropha curcas L.: A promising biofuel plant. Promis. Future Jatropha Curcas Propr. Potential Appl. Hauppauge NY Nova Sci. Publ. Inc 2016, 8, 141–158. [Google Scholar]
- Silitonga, A.S.; Hassan, M.H.; Ong, H.C.; Kusumo, F. Analysis of the performance, emission and combustion characteristics of a turbocharged diesel engine fuelled with Jatropha curcas biodiesel-diesel blends using kernel-based extreme learning machine. Environ. Sci. Pollut. Res. 2017, 24, 25383–25405. [Google Scholar] [CrossRef]
- Abrar, M.M.; Saqib, M.; Abbas, G.; Atiq-ur-Rahman, M.; Mustafa, A.; Shah, S.A.A.; Mehmood, K.; Maitlo, A.A.; Sun, N.; Xu, M. Evaluating the contribution of growth, physiological, and ionic components towards salinity and drought stress tolerance in Jatropha curcas. Plants 2020, 9, 1574. [Google Scholar] [CrossRef] [PubMed]
- Reubens, B.; Achten, W.M.; Maes, W.; Danjon, F.; Aerts, R.; Poesen, J.; Muys, B. More than biofuel? Jatropha curcas root system symmetry and potential for soil erosion control. J. Arid Environ. 2011, 75, 201–205. [Google Scholar] [CrossRef] [Green Version]
- Pompelli, M.F.; Barata-Luís, R.; Vitorino, H.S.; Gonçalves, E.R.; Rolim, E.V.; Santos, M.G.; Almeida-Cortez, J.S.; Ferreira, V.M.; Lemos, E.E.; Endres, L. Photosynthesis, photoprotection and antioxidant activity of purging nut under drought deficit and recovery. Biomass Bioenergy 2010, 34, 1207–1215. [Google Scholar] [CrossRef]
- de Lima Cabral, G.A.; Binneck, E.; de Souza, M.C.P.; da Silva, M.D.; Costa Ferreira Neto, J.R.; Pompelli, M.F.; Endres, L.; Kido, É.A. First expressed TFome of physic nut (Jatropha curcas L.) after salt stimulus. Plant Mol. Biol. Report. 2020, 38, 189–208. [Google Scholar] [CrossRef]
- Gao, S.; Ouyang, C.; Wang, S.; Xu, Y.; Tang, L.; Chen, F. Effects of salt stress on growth, antioxidant enzyme and phenylalanine ammonia-lyase activities in Jatropha curcas L. seedlings. Plant Soil Env. 2008, 54, 374–381. [Google Scholar] [CrossRef] [Green Version]
- de Oliveira, D.F.B.; Endres, L.; Silva, J.V.; Clemente, P.R.A. Pre-colonized seedlings with arbuscular mycorrhizal fungi: An alternative for the cultivation of Jatropha curcas L. in salinized soils. Theor. Exp. Plant Physiol. 2017, 29, 129–142. [Google Scholar] [CrossRef]
- Pompelli, M.F.; Jarma-Orozco, A.; Rodríguez-Páez, L.A. Salinity in Jatropha curcas: A review of physiological, biochemical, and molecular factors involved. Agriculture 2022, 12, 594. [Google Scholar] [CrossRef]
- Patel, A.D.; Panchal, N.S.; Pandey, I.B.; Pandey, A.N. Growth, water status and nutrient accumulation of seedlings of Jatropha curcas L. (Euphorbiaceae) in response to soil salinity. In Anales de Biología; Facultad de Biología, Universidad de Murcia: Murcia, Spain, 2010; pp. 59–71. [Google Scholar]
- Liu, B.; Yang, Q.; Xu, M. Effects of water-nitrogen addition on Jatropha curcas L. physiological characteristics under salt stress. J. Drain. Irrig. Mach. Eng. 2018, 36, 679–684. [Google Scholar]
- da Silva Cavalcante, P.G.; dos Santos, C.M.; de Lins Wanderley Filho, H.C.; Avelino, J.R.L.; Endres, L. Morpho-physiological adaptation of Jatropha curcas L. to salinity stress. Aust. J. Crop Sci. 2018, 12, 563–571. [Google Scholar] [CrossRef]
- Silva, E.d.; Ribeiro, R.; Ferreira-Silva, S.; Viégas, R.; Silveira, J. Comparative effects of salinity and water stress on photosynthesis, water relations and growth of Jatropha curcas plants. J. Arid Environ. 2010, 74, 1130–1137. [Google Scholar] [CrossRef]
- Iqbal, N.; Umar, S.; Khan, N.A. Nitrogen availability regulates proline and ethylene production and alleviates salinity stress in mustard (Brassica juncea). J. Plant Physiol. 2015, 178, 84–91. [Google Scholar] [CrossRef]
- Reda, M.; Migocka, M.; Kłobus, G. Effect of short-term salinity on the nitrate reductase activity in cucumber roots. Plant Sci. 2011, 180, 783–788. [Google Scholar] [CrossRef]
- Syeed, S.; Anjum, N.A.; Nazar, R.; Iqbal, N.; Masood, A.; Khan, N.A. Salicylic acid-mediated changes in photosynthesis, nutrients content and antioxidant metabolism in two mustard (Brassica juncea L.) cultivars differing in salt tolerance. Acta Physiol. Plant 2011, 33, 877–886. [Google Scholar] [CrossRef]
- Hessini, K.; Issaoui, K.; Ferchichi, S.; Saif, T.; Abdelly, C.; Siddique, K.H.; Cruz, C. Interactive effects of salinity and nitrogen forms on plant growth, photosynthesis and osmotic adjustment in maize. Plant Physiol. Biochem. 2019, 139, 171–178. [Google Scholar] [CrossRef]
- Singh, M.; Singh, V.P.; Prasad, S.M. Nitrogen alleviates salinity toxicity in Solanum lycopersicum seedlings by regulating ROS homeostasis. Plant Physiol. Biochem. 2019, 141, 466–476. [Google Scholar] [CrossRef]
- Novoa, R.; Loomis, R. Nitrogen and plant production. Plant Soil 1981, 58, 177–204. [Google Scholar] [CrossRef]
- Peng, J.; Feng, Y.; Wang, X.; Li, J.; Xu, G.; Phonenasay, S.; Luo, Q.; Han, Z.; Lu, W. Effects of nitrogen application rate on the photosynthetic pigment, leaf fluorescence characteristics, and yield of indica hybrid rice and their interrelations. Sci. Rep. 2021, 11, 7485. [Google Scholar] [CrossRef]
- Huang, G.; Shu, Y.; Peng, S.; Li, Y. Leaf photosynthesis is positively correlated with xylem and phloem areas in leaf veins in rice (Oryza sativa) plants. Ann. Bot. 2022, 129, 619–631. [Google Scholar] [CrossRef]
- Ru, C.; Hu, X.; Chen, D.; Song, T.; Wang, W.; Lv, M.; Hansen, N.C. Nitrogen modulates the effects of short-term heat, drought and combined stresses after anthesis on photosynthesis, nitrogen metabolism, yield, and water and nitrogen use efficiency of wheat. Water 2022, 14, 1407. [Google Scholar] [CrossRef]
- Muhammad, I.; Yang, L.; Ahmad, S.; Zeeshan, M.; Farooq, S.; Ali, I.; Khan, A.; Zhou, X.B. Irrigation and nitrogen fertilization alter soil bacterial communities, soil enzyme activities, and nutrient availability in maize crop. Front. Microbiol. 2022, 13, 105. [Google Scholar] [CrossRef]
- Muhammad, I.; Yang, L.; Ahmad, S.; Farooq, S.; Al-Ghamdi, A.A.; Khan, A.; Zeeshan, M.; Elshikh, M.S.; Abbasi, A.M.; Zhou, X.-B. Nitrogen fertilizer modulates plant growth, chlorophyll pigments and enzymatic activities under different irrigation regimes. Agronomy 2022, 12, 845. [Google Scholar] [CrossRef]
- Zhang, Y.; Luo, J.; Peng, F.; Xiao, Y.; Du, A. Application of bag-controlled release fertilizer facilitated new root formation, delayed leaf, and root senescence in peach trees and improved nitrogen utilization efficiency. Front. Plant Sci. 2021, 12, 627313. [Google Scholar] [CrossRef]
- Shen, Z.; Pu, X.; Wang, S.; Dong, X.; Cheng, X.; Cheng, M. Silicon improves ion homeostasis and growth of liquorice under salt stress by reducing plant Na+ uptake. Sci. Rep. 2022, 12, 5089. [Google Scholar] [CrossRef]
- Ahanger, M.A.; Qin, C.; Begum, N.; Maodong, Q.; Dong, X.X.; El-Esawi, M.; El-Sheikh, M.A.; Alatar, A.A.; Zhang, L. Nitrogen availability prevents oxidative effects of salinity on wheat growth and photosynthesis by up-regulating the antioxidants and osmolytes metabolism, and secondary metabolite accumulation. BMC Plant Biol. 2019, 19, 479. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.; Singh, V.P.; Prasad, S.M. Responses of photosynthesis, nitrogen and proline metabolism to salinity stress in Solanum lycopersicum under different levels of nitrogen supplementation. Plant Physiol. Biochem. 2016, 109, 72–83. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhang, D.; Cai, Z.; Wang, L.; Wang, J.; Sun, L.; Fan, X.; Shen, S.; Zhao, J. Spectral technology and multispectral imaging for estimating the photosynthetic pigments and SPAD of the Chinese cabbage based on machine learning. Comput. Electron. Agric. 2022, 195, 106814. [Google Scholar] [CrossRef]
- Gong, B.; Wen, D.; VandenLangenberg, K.; Wei, M.; Yang, F.; Shi, Q.; Wang, X. Comparative effects of NaCl and NaHCO3 stress on photosynthetic parameters, nutrient metabolism, and the antioxidant system in tomato leaves. Sci. Hortic. 2013, 157, 1–12. [Google Scholar] [CrossRef]
- Zhang, M.S.; Tan, F. Relationship between ratio of Chlorophyll a and b under water stress and drought resistance of different sweet potato varieties. Seed 2001, 4, 23–25. [Google Scholar]
- Liu, C.Y.; Yan, M.; Huang, X.B. Effects of salt stress on growth and physiological characteristics of pomegranate (Punica granatum L.) cuttings. Pak. J. Bot. 2018, 50, 457–464. [Google Scholar]
- Suzuki, N.; Koussevitzky, S.; Mittler, R.; Miller, G. ROS and redox signalling in the response of plants to abiotic stress. Plant Cell Environ. 2012, 35, 259–270. [Google Scholar] [CrossRef]
- Feghhenabi, F.; Hadi, H.; Khodaverdiloo, H.; Van Genuchten, M.T.; Pessarakli, M. Improving wheat (Triticum aestivum L.) antioxidative defense mechanisms against salinity stress by exogenous application of potassium silicate. J. Plant Nutr. 2022, 45, 2887–2905. [Google Scholar] [CrossRef]
- Turan, M.; Ekinci, M.; Kul, R.; Boynueyri, F.G.; Yildirim, E. Mitigation of salinity stress in cucumber seedlings by exogenous hydrogen sulfide. J. Plant Res. 2022, 135, 517–529. [Google Scholar] [CrossRef]
- Liang, Y.; Ding, R. Influence of silicon on microdistribution of mineral ions in roots of salt-stressed barley as associated with salt tolerance in plants. Sci. China Ser. C Life Sci. 2002, 45, 298–308. [Google Scholar] [CrossRef] [PubMed]
- Harrison, E.L.; Arce Cubas, L.; Gray, J.E.; Hepworth, C. The influence of stomatal morphology and distribution on photosynthetic gas exchange. Plant J. 2020, 101, 768–779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, Q.; Gu, S.; Kang, S.; Du, T.; Tong, L.; Wood, J.D.; Ding, R. Mild water and salt stress improve water use efficiency by decreasing stomatal conductance via osmotic adjustment in field maize. Sci. Total Environ. 2022, 805, 150364. [Google Scholar] [CrossRef]
- Pan, T.; Liu, M.; Kreslavski, V.D.; Zharmukhamedov, S.K.; Nie, C.; Yu, M.; Kuznetsov, V.V.; Allakhverdiev, S.I.; Shabala, S. Non-stomatal limitation of photosynthesis by soil salinity. Crit. Rev. Environ. Sci. Technol. 2021, 51, 791–825. [Google Scholar] [CrossRef]
- Jamil, M.; Lee, K.J.; Kim, J.M.; Kim, H.-S.; Rha, E.S. Salinity reduced growth PS2 photochemistry and chlorophyll content in radish. Sci. Agric. 2007, 64, 111–118. [Google Scholar] [CrossRef] [Green Version]
- Peng, W.; Wang, C.-D.; Wang, X.-L.; Wu, Y.-L.; Zhang, Y.; Sun, Y.-G.; Yi, S.; Mi, G.-H. Increasing nitrogen absorption and assimilation ability under mixed NO3− and NH4+ supply is a driver to promote growth of maize seedlings. J. Integr. Agric. 2023, 22, 1896–1908. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Soil Salinity (g/kg) | Nitrogen Application Rate (gN/Plant) | ||
|---|---|---|---|---|
| 1 DAE | 60 DAE | Total | ||
| CK | 0 | 0 | 0 | 0 |
| S1N0 | 2 | 0 | 0 | 0 |
| S1N1 | 10 | 10 | 20 | |
| S1N2 | 30 | 30 | 60 | |
| S1N3 | 50 | 50 | 100 | |
| S2N0 | 4 | 0 | 0 | 0 |
| S2N1 | 10 | 10 | 20 | |
| S2N2 | 30 | 30 | 60 | |
| S2N3 | 50 | 50 | 100 | |
| S3N0 | 6 | 0 | 0 | 0 |
| S3N1 | 10 | 10 | 20 | |
| S3N2 | 30 | 30 | 60 | |
| S3N3 | 50 | 50 | 100 | |
| Treatments | POD/(U·g−1·min−1) | SOD/(U·g−1·min−1) | CAT/(U·g−1·min−1) |
|---|---|---|---|
| CK | 83.9 ± 1.0 k | 20.4 ± 0.8 k | 60.3 ± 1.2 l |
| S1N0 | 85.4 ± 1.0 k | 23.2 ± 0.3 j | 68.0 ± 0.6 k |
| S1N1 | 115.5 ± 0.5 h | 29.8 ± 0.3 i | 78.0 ± 0.8 h |
| S1N2 | 144.9 ± 0.6 f | 48.2 ± 0.2 e | 92.1 ± 0.6 e |
| S1N3 | 176.4 ± 0.6 c | 67.3 ± 0.3 b | 108.9 ± 0.4 c |
| S2N0 | 107.3 ± 0.6 i | 31.2 ± 0.2 h | 71.1 ± 0.7 j |
| S2N1 | 148.7 ± 0.5 e | 40.1 ± 0.3 g | 87.1 ± 0.9 f |
| S2N2 | 198.1 ± 0.8 b | 59.4 ± 0.3 c | 116.1 ± 0.7 b |
| S2N3 | 231.1 ± 0.7 a | 74.2 ± 0.6 a | 128.1 ± 0.6 a |
| S3N0 | 89.3 ± 0.4 j | 20.1 ± 0.3 k | 61.0 ± 1.1 l |
| S3N1 | 116.4 ± 0.6 h | 30.8 ± 0.3 h | 74.0 ± 0.6 i |
| S3N2 | 139.9 ± 0.6 g | 43.5 ± 0.2 f | 84.0 ± 0.5 g |
| S3N3 | 172.3 ± 0.7 d | 52.6 ± 0.7 d | 97.9 ± 0.6 d |
| Significant of factors (F-value) | |||
| Salinity stress (S) | 5452 ** | 2252 ** | 943 ** |
| Nitrogen application (N) | 13,188 ** | 9947 ** | 2349 ** |
| S × N | 270 ** | 138 ** | 76 ** |
| Treatments | Chl a/(mg/g FW) | Chl b/(mg/g FW) | Chl a+b/(mg/g FW) | Chl a/b ratio | Car/(mg/g FW) |
|---|---|---|---|---|---|
| CK | 2.15 ± 0.21 b | 1.03 ± 0.02 b | 3.18 ± 0.11 a | 2.09 ± 0.13 j | 0.63 ± 0.01 c |
| S1N0 | 1.74 ± 0.00 e | 0.72 ± 0.01 f | 2.46 ± 0.01 f | 2.43 ± 0.01 c | 0.51 ± 0.00 e |
| S1N1 | 1.81 ± 0.02 c | 0.83 ± 0.00 d | 2.65 ± 0.02 e | 2.18 ± 0.01 i | 0.56 ± 0.01 e |
| S1N2 | 2.06 ± 0.01 c | 1.07 ± 0.01 b | 3.14 ± 0.02 c | 1.93 ± 0.01 l | 0.67 ± 0.01 b |
| S1N3 | 2.27 ± 0.00 a | 1.20 ± 0.01 a | 3.47 ± 0.01 b | 1.90 ± 0.01 l | 0.79 ± 0.01 a |
| S2N0 | 1.41 ± 0.01 j | 0.59 ± 0.01 j | 2.00 ± 0.01 k | 2.38 ± 0.02 d | 0.41 ± 0.00 j |
| S2N1 | 1.49 ± 0.01 h | 0.66 ± 0.01 h | 2.15 ± 0.01 i | 2.26 ± 0.02 f | 0.44 ± 0.01 i |
| S2N2 | 1.69 ± 0.01 f | 0.77 ± 0.01 e | 2.46 ± 0.01 g | 2.20 ± 0.01 h | 0.53 ± 0.00 f |
| S2N3 | 1.83 ± 0.01 d | 0.89 ± 0.00 c | 2.72 ± 0.02 d | 2.05 ± 0.01 k | 0.61 ± 0.01 d |
| S3N0 | 1.03 ± 0.01 l | 0.40 ± 0.00 l | 1.43 ± 0.02 m | 2.61 ± 0.01 a | 0.27 ± 0.00 k |
| S3N1 | 1.36 ± 0.01 k | 0.55 ± 0.00 k | 1.91 ± 0.01 l | 2.48 ± 0.02 b | 0.40 ± 0.00 j |
| S3N2 | 1.45 ± 0.01 i | 0.64 ± 0.01 i | 2.09 ± 0.01 j | 2.26 ± 0.02 e | 0.47 ± 0.01 h |
| S3N3 | 1.54 ± 0.01 g | 0.69 ± 0.01 g | 2.23 ± 0.01 h | 2.23 ± 0.02 g | 0.50 ± 0.01 g |
| Significant of factors (F-value) | |||||
| Salinity stress (S) | 4219 ** | 3306 ** | 1734 ** | 815 ** | 1262 ** |
| Nitrogen application (N) | 1419 ** | 1640 ** | 359 ** | 426 ** | 773 ** |
| N × S | 54 ** | 76 ** | 22 ** | 37 ** | 24 ** |
| Treatments | Fv/Fm | ΦPSⅡ | qP | NPQ | ETR |
|---|---|---|---|---|---|
| CK | 0.781 ± 0.011 ab | 0.521 ± 0.014 a | 0.751 ± 0.012 a | 1.473 ± 0.016 c | 199.3 ± 5.4 a |
| S1N0 | 0.777 ± 0.009 abc | 0.449 ± 0.008 ef | 0.651 ± 0.004 fg | 1.522 ± 0.011 c | 142.3 ± 4.4 g |
| S1N1 | 0.802 ± 0.005 a | 0.485 ± 0.007 abc | 0.714 ± 0.005 ab | 1.034 ± 0.013 i | 177.8 ± 3.3 bc |
| S1N2 | 0.809 ± 0.007 a | 0.489 ± 0.007 abc | 0.720 ± 0.005 ab | 0.913 ± 0.006 j | 185.0 ± 3.5 ab |
| S1N3 | 0.818 ± 0.008 a | 0.506 ± 0.010 a | 0.734 ± 0.009 a | 0.807 ± 0.012 k | 196.5 ± 1.8 a |
| S2N0 | 0.774 ± 0.008 abc | 0.441 ± 0.010 f | 0.638 ± 0.006 gh | 1.603 ± 0.013 b | 131.4 ± 3.78 h |
| S2N1 | 0.793 ± 0.005 abc | 0.475 ± 0.006 bcd | 0.691 ± 0.006 cd | 1.314 ± 0.011 f | 164.4 ± 2.9 de |
| S2N2 | 0.799 ± 0.004 ab | 0.481 ± 0.007 abcd | 0.703 ± 0.007 bc | 1.281 ± 0.010 g | 170.2 ± 5.0 cd |
| S2N3 | 0.813 ± 0.005 a | 0.495 ± 0.011 ab | 0.727 ± 0.009 a | 1.197 ± 0.006 h | 191.9 ± 5.4 a |
| S3N0 | 0.752 ± 0.005 bc | 0.437 ± 0.004 f | 0.621 ± 0.004 h | 1.719 ± 0.011 a | 117.2 ± 3.7 i |
| S3N1 | 0.782 ± 0.004 abc | 0.455 ± 0.006 def | 0.663 ± 0.007 ef | 1.492 ± 0.009 c | 149.6 ± 3.7 fg |
| S3N2 | 0.749 ± 0.046 c | 0.467 ± 0.009 cde | 0.683 ± 0.008 d | 1.397 ± 0.009 d | 159.2 ± 2.6 def |
| S3N3 | 0.784 ± 0.005 abc | 0.463 ± 0.011 cdef | 0.672 ± 0.006 de | 1.352 ± 0.008 e | 153.4 ± 3.4 efg |
| Significant of factors (F-value) | |||||
| Salinity stress (S) | 6.5 ** | 10.9 ** | 52.0 ** | 1722 ** | 68.4 ** |
| Nitrogen application (N) | 3.6 * | 17.8 ** | 80.4 ** | 1342 ** | 103.24 ** |
| S × N | 0.48 NS | 0.80 NS | 2.31 ** | 69.4 * | 2.99 ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Z.; Tan, S.; Yang, Q.; Chen, S.; Qi, C.; Liu, X.; Liang, J.; Wang, H. Nitrogen Application Alleviates Impairments for Jatropha curcas L. Seedling Growth under Salinity Stress by Regulating Photosynthesis and Antioxidant Enzyme Activity. Agronomy 2023, 13, 1749. https://doi.org/10.3390/agronomy13071749
Yang Z, Tan S, Yang Q, Chen S, Qi C, Liu X, Liang J, Wang H. Nitrogen Application Alleviates Impairments for Jatropha curcas L. Seedling Growth under Salinity Stress by Regulating Photosynthesis and Antioxidant Enzyme Activity. Agronomy. 2023; 13(7):1749. https://doi.org/10.3390/agronomy13071749
Chicago/Turabian StyleYang, Zhao, Shuai Tan, Qiliang Yang, Shaomin Chen, Changmin Qi, Xiaogang Liu, Jiaping Liang, and Haidong Wang. 2023. "Nitrogen Application Alleviates Impairments for Jatropha curcas L. Seedling Growth under Salinity Stress by Regulating Photosynthesis and Antioxidant Enzyme Activity" Agronomy 13, no. 7: 1749. https://doi.org/10.3390/agronomy13071749