
The Effect of Increasing Irrigation Rates on the Carbon Isotope Discrimination of Apple Leaves
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
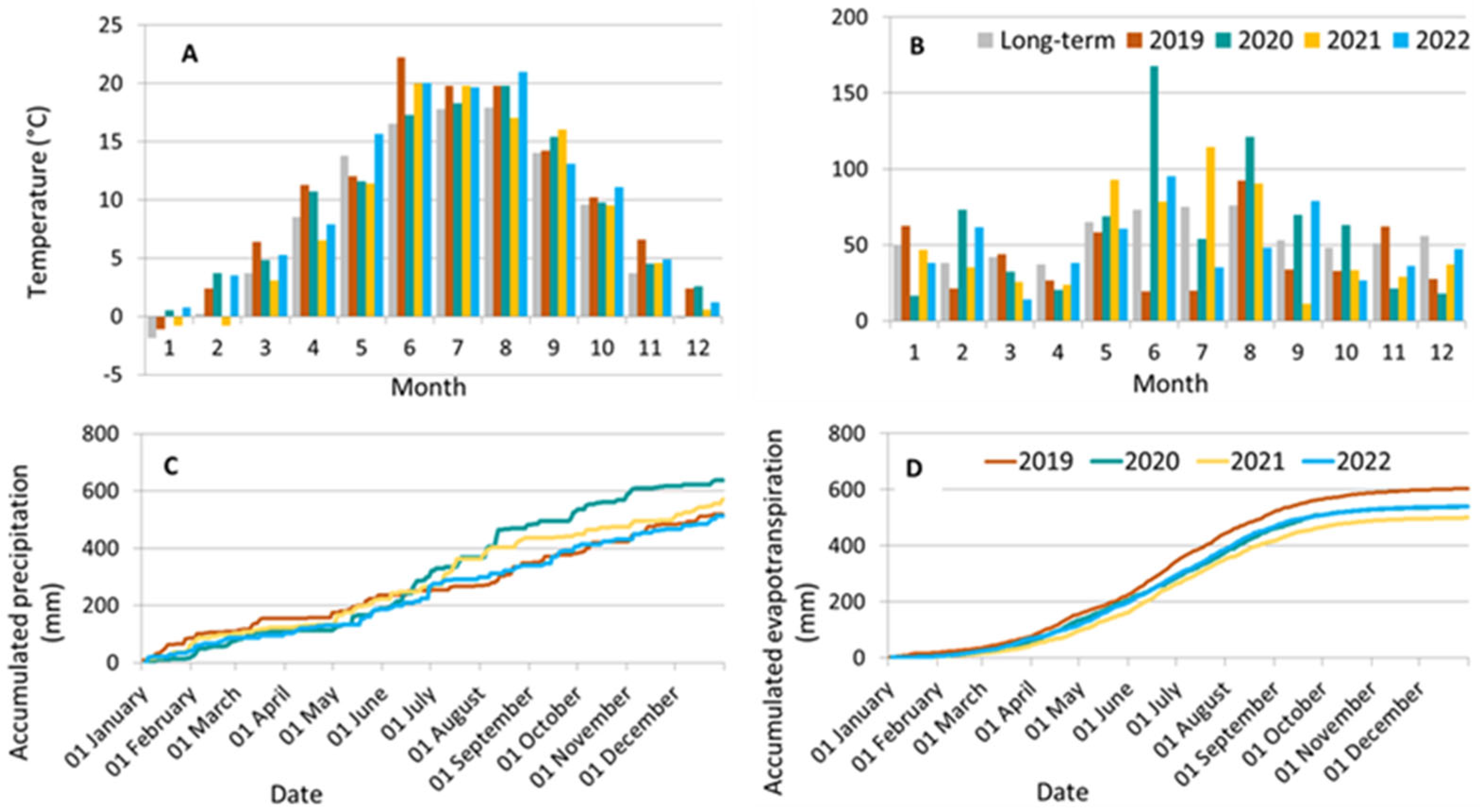
2.1. Site Characteristics
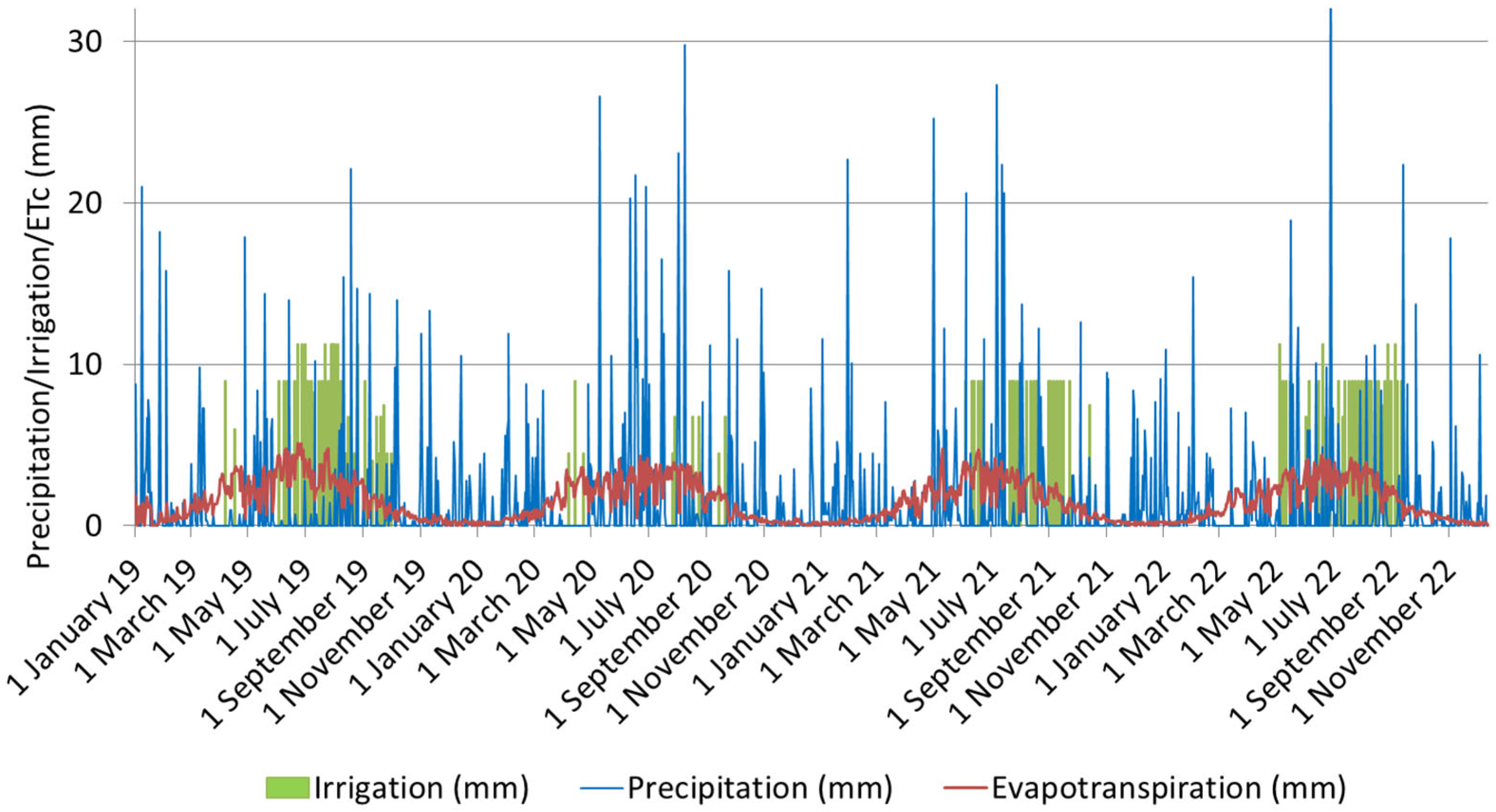
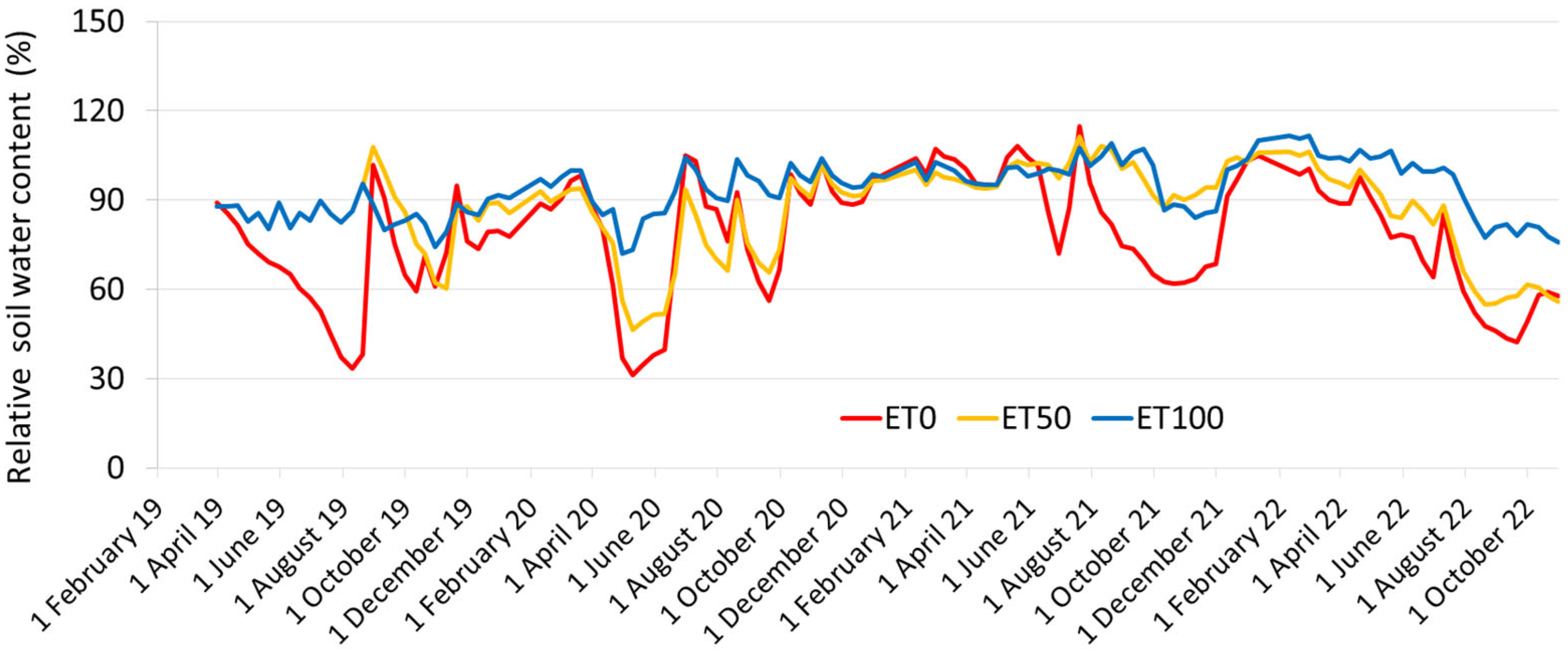
2.2. Layout of the Orchard and Experimental Irrigation Treatments
2.3. Terms of Sampling of Apple Leaves
2.3.1. Experiment A
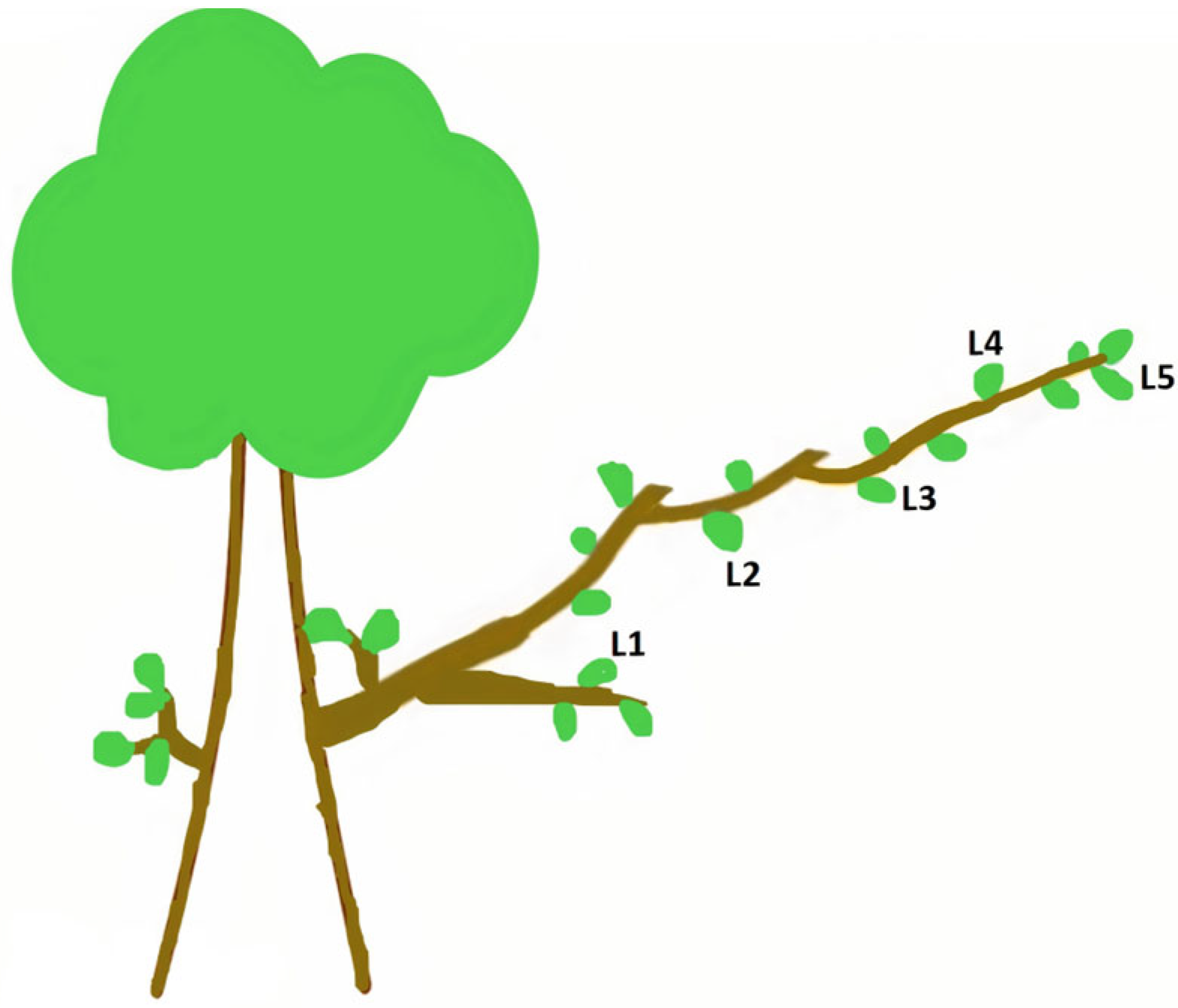
2.3.2. Experiment B
2.4. Chemical Analysis of the Leaves
2.5. Statistical Analysis
3. Results
3.1. The Effect of Irrigation on Δ13C—Experiment A
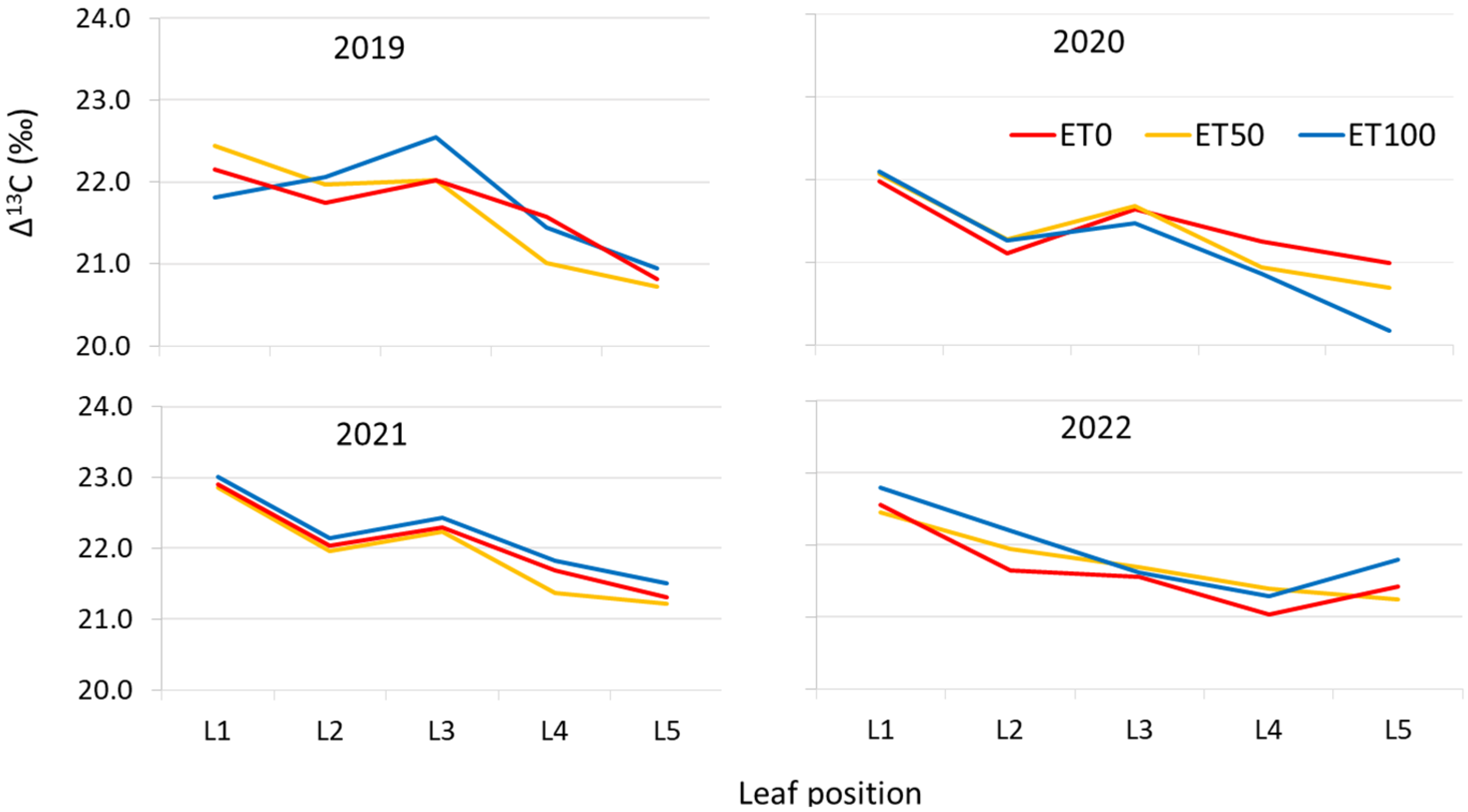
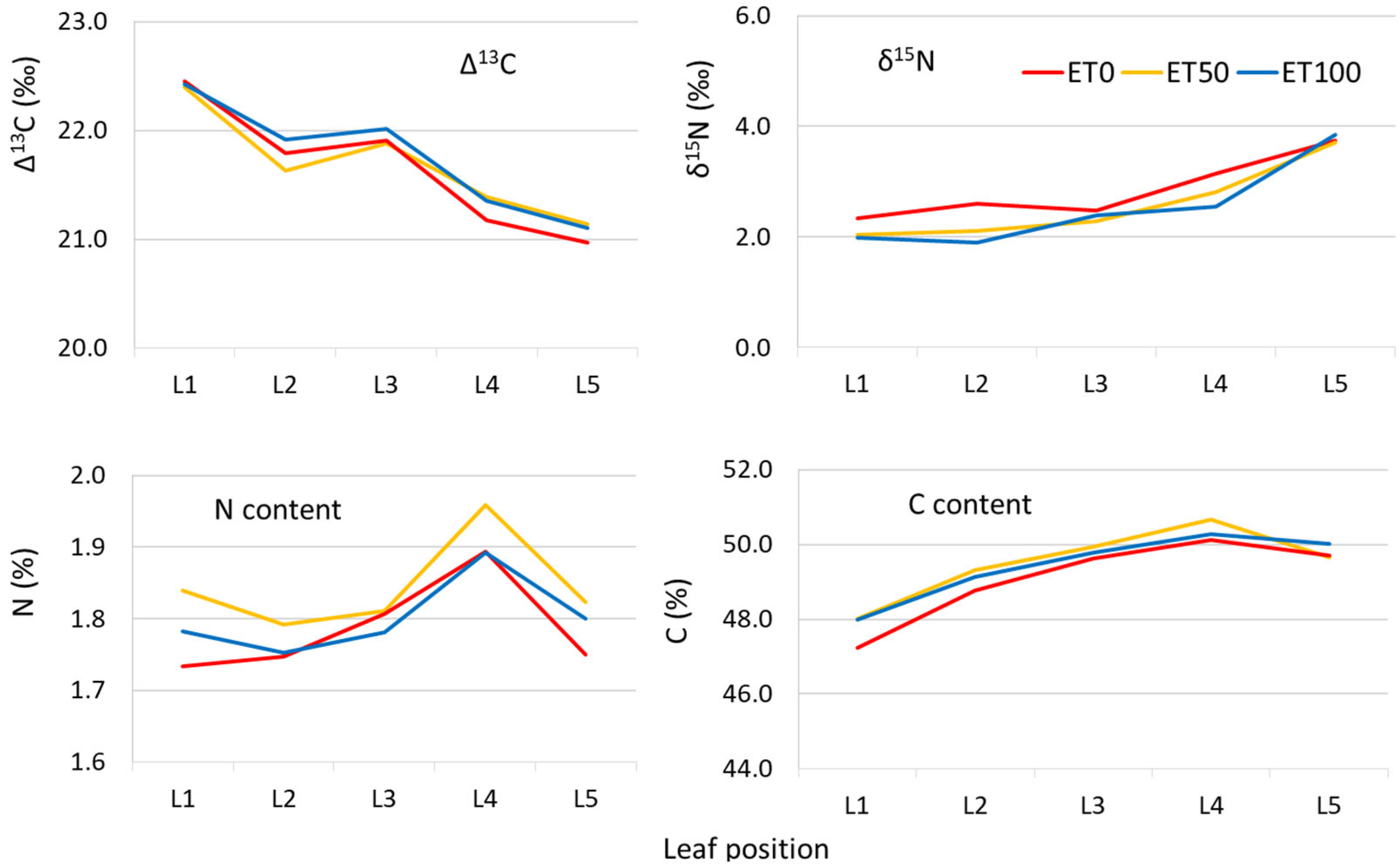
3.2. The Effect of Irrigation on Δ13C—Experiment B
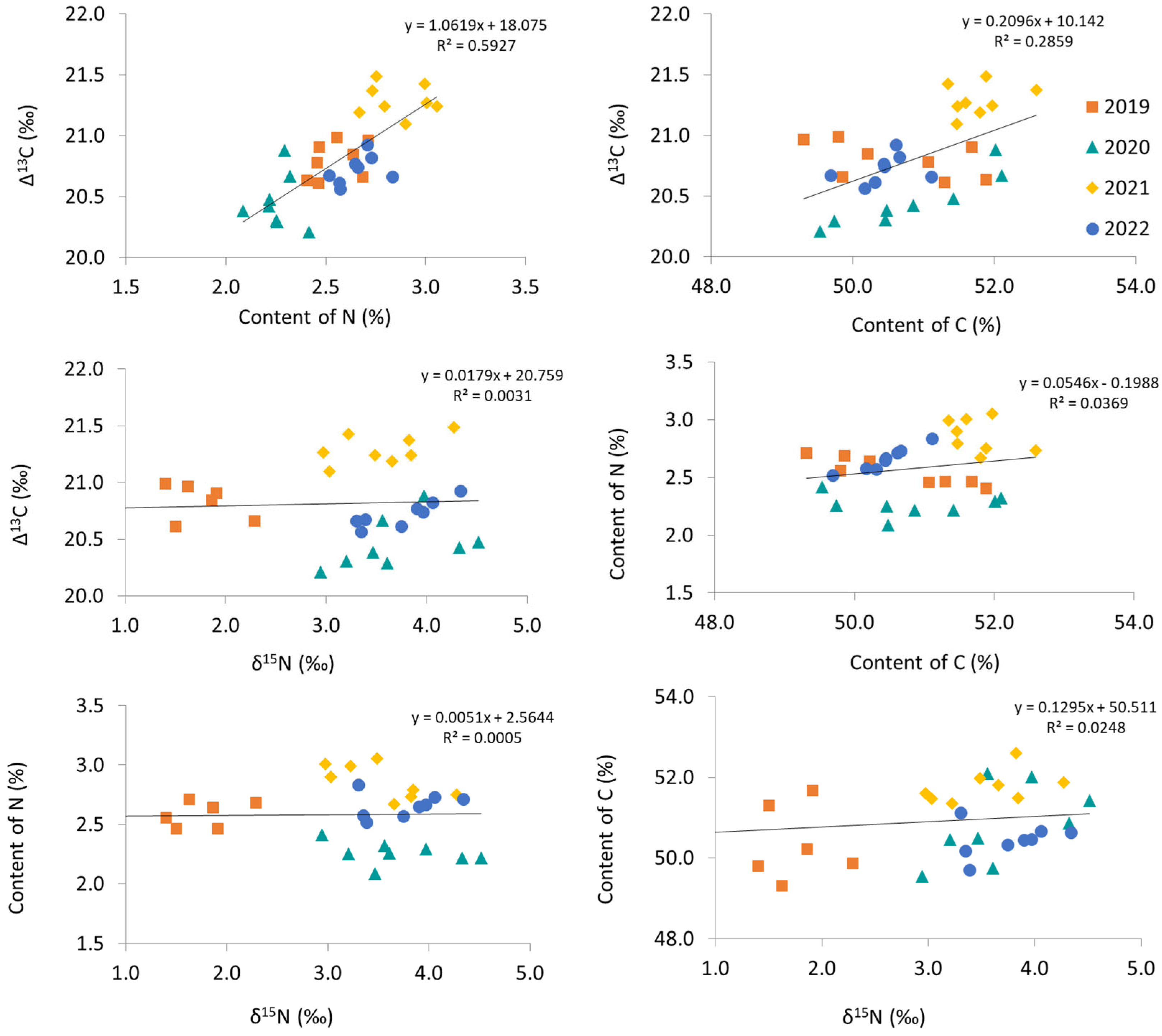
3.3. The Relationships between Δ13C and the Contents of N and C and δ15N of Apple Leaves
4. Discussion
4.1. The Effect of Irrigation Rates on Δ13C
4.2. The Year Variability of Δ13C
4.3. The Indices for the Use of Carbon from the Previous Year
4.4. The Effect of Leaf Position (Experiment B)
4.5. The Relationships among Δ13C, δ15N, and N and C Contents of Apple Leaves
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Potopová, V.; Trnka, M.; Vizina, A.; Semerádová, D.; Balek, J.; Chawdhery, M.R.A.; Musiolková, M.; Pavlík, P.; Možný, M.; Štěpánek, P.; et al. Projection of 21st century irrigation water requirements for sensitive agricultural crop commodities across the Czech Republic. Agric. Water Manag. 2022, 262, 107337. [Google Scholar] [CrossRef]
- Trnka, M.; Balek, J.; Brázdil, R.; Dubrovský, M.; Eitzinger, J.; Hlavinka, P.; Chuchma, F.; Možný, M.; Prášil, I.; Růžek, P.; et al. Observed changes in the agroclimatic zones in the Czech Republic between 1961 and 2019. Plant Soil Environ. 2021, 67, 154–163. [Google Scholar] [CrossRef]
- Trnka, M.; Rötter, R.P.; Ruiz-Ramos, M.; Kersebaum, K.C.; Olesen, J.E.; Žalud, Z.; Semenov, M.A. Adverse weather conditions for European wheat production will become more frequent with climate change. Nat. Clim. Chang. 2014, 4, 637–643. [Google Scholar] [CrossRef]
- Spitz, P.; Hemerka, I. The development of the irrigation requirement in the dry regions of the Czech Republic with reference to possible climatic change. Acta Univ. Agric. Silvic. Mendel. Brun. 2006, 54, 171–180. [Google Scholar] [CrossRef] [Green Version]
- Gu, Z.; Qi, Z.; Burghate, R.; Yuan, S.; Jiao, X.; Xu, J. Irrigation scheduling approaches and applications: A review. J. Irrig. Drain. Eng. 2020, 146, 04020007. [Google Scholar] [CrossRef]
- Glenn, D.M. An analysis of ash and isotopic carbon discrimination (δ13C) methods to evaluate water use efficiency in apple. Sci. Hortic. 2014, 171, 32–36. [Google Scholar] [CrossRef]
- CSN 750434; Amelioration-Irrigation Requirement. Czech State Norm: Hradec Králové, Czech Republic, 2017; (In Czech with English Abstract). Available online: https://www.technicke-normy-csn.cz/csn-75-0434-750434-225463.html (accessed on 15 June 2020).
- Litschmann, T. Virrib: A soil moisture sensor and its application in agriculture. Commun. Soil Sci. Plant Anal. 1991, 22, 409–418. [Google Scholar] [CrossRef]
- Park, R.; Epstein, S. Carbon isotope fractionation during photosynthesis. Geochim. Cosmochim. Acta 1960, 21, 110–126. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Ehleringer, J.R.; Hubick, K.T. Carbon isotope discrimination and photosynthesis. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1989, 40, 503–537. [Google Scholar] [CrossRef]
- Condon, A.G.; Richards, R.A.; Farquhar, G.D. Carbon isotope discrimination is positively correlated with grain yield and dry matter production in field-grown wheat. Crop Sci. 1987, 27, 996–1001. [Google Scholar] [CrossRef]
- Kunz, K.; Hu, Y.; Schmidhalter, U. Carbon isotope discrimination as a key physiological trait to phenotype drought/heat resistance of future climate-resilient German winter wheat compared with relative leaf water content and canopy temperature. Front. Plant Sci. 2022, 13, 1043458. [Google Scholar] [CrossRef]
- Mininni, A.N.; Tuzio, A.C.; Brugnoli, E.; Dichio, B.; Sofo, A. Carbon isotope discrimination and water use efficiency in interspecific Prunus hybrids subjected to drought stress. Plant Physiol. Biochem. 2022, 175, 33–43. [Google Scholar] [CrossRef]
- Casagrande Biasuz, E.; Kalcsits, L.A. Apple rootstocks affect functional leaf traits with consequential effects on carbon isotope composition and vegetative vigour. AoB Plants 2022, 14, plac020. [Google Scholar] [CrossRef]
- Glenn, D.M. Canopy gas exchange and water use efficiency of ‘Empire’ apple in response to particle film, irrigation, and microclimatic factors. J. Am. Soc. Hortic. Sci. 2010, 135, 25–32. [Google Scholar] [CrossRef] [Green Version]
- Zhu, L.; Liang, Z.S.; Xu, X.; Li, S.H.; Monneveux, P. Evidences for the association between carbon isotope discrimination and grain yield-Ash content and stem carbohydrate in spring wheat grown in Ningxia (Northwest China). Plant Sci. 2009, 176, 758–767. [Google Scholar] [CrossRef]
- Bchir, A.; Escalona, J.M.; Gallé, A.; Hernández-Montes, E.; Tortosa, I.; Braham, M.; Medrano, H. Carbon isotope discrimination (δ13C) as an indicator of vine water status and water use efficiency (WUE): Looking for the most representative sample and sampling time. Agric. Water Manag. 2016, 167, 11–20. [Google Scholar] [CrossRef]
- Brillante, L.; Martínez-Lüscher, J.; Yu, R.; Kurtural, S.K. Carbon isotope discrimination (δ13 C) of grape musts is a reliable tool for zoning and the physiological ground-truthing of sensor maps in precision viticulture. Front. Environ. Sci. 2020, 8, 561477. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, X.; Liu, X.; Zhang, X.; Shao, L.; Sun, H.; Chen, S. The effects of nitrogen supply and water regime on instantaneous WUE, time-integrated WUE and carbon isotope discrimination in winter wheat. Field Crops Res. 2013, 144, 236–244. [Google Scholar] [CrossRef]
- Sidhu, R.S.; Bound, S.A.; Hunt, I. Crop load and thinning methods impact yield, nutrient content, fruit quality, and physiological disorders in ‘Scilate’ apples. Agronomy 2022, 12, 1989. [Google Scholar] [CrossRef]
- Baïram, E.; LeMorvan, C.; Delaire, M.; Buck-Sorlin, G. Fruit and leaf response to different source–sink ratios in apple, at the scale of the fruit-bearing branch. Front. Plant Sci. 2019, 10, 1039. [Google Scholar] [CrossRef]
- Racskó, J. Crop load, fruit thinning and their effects on fruit quality of apple (Malus domestica Borkh.). Acta Agrar. Debr. 2006, 24, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Štěpánek, P.; Trnka, M.; Chuchma, F.; Zahradníček, P.; Skalák, P.; Farda, A.; Fiala, R.; Hlavinka, P.; Balek, J.; Semerádová, D.; et al. Drought prediction system for central europe and its validation. Geosciences 2018, 8, 104. [Google Scholar] [CrossRef] [Green Version]
- Kohut, M.; Fiala, R.; Chuchma, F.; Rožnovský, J.; Hora, P. Drought monitoring on the CHMI website. In Proceedings of the Mendel a Bioklimatologie, Brno, Czech Republic, 3–5 September 2014; pp. 1–19, ISBN 978-80-210-6983-1. [Google Scholar]
- Allen, R.G.; Pereira, L.S.; Raes, D.; Smith, M. Crop Evapotranspiration-Guidelines for Computing Crop Water Requirements-FAO Irrigation and Drainage Paper 56; FAO: Rome, Italy, 1998; ISBN 9251042195. [Google Scholar]
- Robinson, D. Δ15N as an integrator of the nitrogen cycle. Trends Ecol. Evol. 2001, 16, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Plavcová, L.; Mészáros, M.; Jupa, R.; Scháňková, K.; Kovalíková, Z.; Náměstek, J.; Mahrová, A. Yield and water relations of two apple cultivars under irrigation. Irrig. Sci. 2023, in press. [CrossRef]
- Raimanová, I.; Svoboda, P.; Kurešová, G.; Haberle, J. The effect of different post-anthesis water supply on the carbon isotope discrimination of winter wheat grain. Plant Soil Environ. 2016, 62, 329–334. [Google Scholar] [CrossRef] [Green Version]
- Cui, N.; Du, T.; Kang, S.; Li, F.; Hu, X.; Wang, M.; Li, Z. Relationship between stable carbon isotope discrimination and water use efficiency under regulated deficit irrigation of pear-jujube tree. Agric. Water Manag. 2009, 96, 1615–1622. [Google Scholar] [CrossRef]
- Torbenson, M.C.A.; Büntgen, U.; Esper, J.; Urban, O.; Balek, J.; Reinig, F.; Krusic, P.J.; del Castillo, E.M.; Brázdil, R.; Semerádová, D.; et al. Central European agroclimate over the past 2000 years. J. Clim. 2023, 36, 4429–4441. [Google Scholar] [CrossRef]
- Flohr, P.; Jenkins, E.; Williams, H.R.S.; Jamjoum, K.; Nuimat, S.; Müldner, G. What can crop stable isotopes ever do for us? An experimental perspective on using cereal carbon stable isotope values for reconstructing water availability in semi-arid and arid environments. Veg. Hist. Archaeobot. 2019, 28, 497–512. [Google Scholar] [CrossRef] [Green Version]
- Wahbi, A.; Shaaban, A.S.A. Relationship between carbon isotope discrimination (Δ), yield and water use efficiency of durum wheat in Northern Syria. Agric. Water Manag. 2011, 98, 1856–1866. [Google Scholar] [CrossRef]
- Dalal, R.C.; Strong, W.M.; Cooper, J.E.; King, A.J. Relationship between water use and nitrogen use efficiency discerned by 13C discrimination and 15N isotope ratio in bread wheat grown under no-till. Soil Tillage Res. 2013, 128, 110–118. [Google Scholar] [CrossRef]
- Spitz, P.; Zavadil, J.; Duffková, R.; Korsuň, S.; Nechvátal, M.; Hemerka, I. Methods of Management of Crops Irrigation Regimes of Crops with Calculation Programme IRRIPROG. Certified Methods; Nechvátal, I.M., Ed.; Výzkumný Ústav Meliorací a Ochrany Půdy, v.v.i.: Prague, Czech Republic, 2011; ISBN 978-80-87361-11-5. [Google Scholar]
- Jiang, X.; He, L. Investigation of effective irrigation strategies for high-density apple orchards in Pennsylvania. Agronomy 2021, 11, 732. [Google Scholar] [CrossRef]
- Lo, T.H.; Rudnick, D.R.; Singh, J.; Nakabuye, H.N.; Katimbo, A.; Heeren, D.M.; Ge, Y. Field assessment of interreplicate variability from eight electromagnetic soil moisture sensors. Agric. Water Manag. 2020, 231, 105984. [Google Scholar] [CrossRef] [Green Version]
- Lind, K.R.; Siemianowski, O.; Yuan, B.; Sizmur, T.; VanEvery, H.; Banerjee, S.; Cademartiri, L. Evidence for root adaptation to a spatially discontinuous water availability in the absence of external water potential gradients. Proc. Natl. Acad. Sci. USA 2021, 118, e2012892118. [Google Scholar] [CrossRef]
- Svoboda, P.; Haberle, J.; Moulik, M.; Raimanová, I.; Kurešová, G.; Mészáros, M. The effect of drip irrigation on the length and distribution of apple tree roots. Horticulturae 2023, 9, 405. [Google Scholar] [CrossRef]
- McQueen, J.C. Carbohydrate storage and remobilisation: The mechanisms involved in woody stem tissue of apple trees. Doctoral Thesis, University of Waikato, Hamilton, New Zealand, 2003. [Google Scholar]
- Breen, K.; Tustin, S.; Palmer, J.; Boldingh, H.; Close, D. Revisiting the role of carbohydrate reserves in fruit set and early-season growth of apple. Sci. Hortic. 2020, 261, 109034. [Google Scholar] [CrossRef]
- Loescher, W.H.; McCamant, T.; Keller, J.D. Carbohydrate reserves, translocation, and storage in woody plant roots. HortScience 1990, 25, 274–281. [Google Scholar] [CrossRef] [Green Version]
- Imada, S.; Tako, Y. Seasonal accumulation of photoassimilated carbon relates to growth rate and use for new aboveground organs of young apple trees in following spring. Tree Physiol. 2022, 42, 2294–2305. [Google Scholar] [CrossRef]
- Davies, W.J.; Wilkinson, S.; Loveys, B. Stomatal control by chemical signalling and the exploitation of this mechanism to increase water use efficiency in agriculture. New Phytol. 2002, 153, 449–460. [Google Scholar] [CrossRef] [Green Version]
- Quinlan, J.D. Mobilization of 14C in the spring following autumn assimilation of 14CO2 by an apple rootstock. J. Hortic. Sci. 1969, 44, 107–110. [Google Scholar] [CrossRef]
- Vogado, N.O.; Winter, K.; Ubierna, N.; Farquhar, G.D.; Cernusak, L.A. Directional change in leaf dry matter δ13C during leaf development is widespread in C3 plants. Ann. Bot. 2020, 126, 981–990. [Google Scholar] [CrossRef]
- Dercon, G.; Clymans, E.; Diels, J.; Merckx, R.; Deckers, J. Differential 13C isotopic discrimination in maize at varying water stress and at low to high nitrogen availability. Plant Soil 2006, 282, 313–326. [Google Scholar] [CrossRef]
- Clay, D.E.; Engel, R.E.; Long, D.S.; Liu, Z. Nitrogen and water stress interact to influence carbon-13 discrimination in wheat. Soil Sci. Soc. Am. J. 2001, 65, 1823–1828. [Google Scholar] [CrossRef]
- Ngao, J.; Martinez, S.; Marquier, A.; Bluy, S.; Saint-Joanis, B.; Costes, E.; Pallas, B. Spatial variability in carbon- and nitrogen-related traits in apple trees: The effects of the light environment and crop load. J. Exp. Bot. 2021, 72, 1933–1945. [Google Scholar] [CrossRef] [PubMed]
- Evans, R.D. Physiological mechanisms influencing plant nitrogen isotope composition. Trends Plant Sci. 2001, 6, 121–126. [Google Scholar] [CrossRef]
- Peri, P.L.; Ladd, B.; Pepper, D.A.; Bonser, S.P.; Laffan, S.W.; Amelung, W. Carbon (δ 13C) and nitrogen (δ 15N) stable isotope composition in plant and soil in Southern Patagonia’s native forests. Glob. Chang. Biol. 2012, 18, 311–321. [Google Scholar] [CrossRef]
- Ma, J.Y.; Sun, W.; Liu, X.N.; Chen, F.H. Variation in the stable carbon and nitrogen isotope composition of plants and soil along a precipitation gradient in northern China. PLoS ONE 2012, 7, e51894. [Google Scholar] [CrossRef]
- Massonnet, C.; Costes, E.; Rambal, S.; Dreyer, E.; Regnard, J.L. Stomatal regulation of photosynthesis in apple leaves: Evidence for different water-use strategies between two cultivars. Ann. Bot. 2007, 100, 1347–1356. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.H.; Cheng, L.; Liang, D.; Zou, Y.J.; Ma, F.W. Growth, gas exchange, water-use efficiency, and carbon isotope composition of ’Gale Gala´ apple trees grafted onto 9 wild Chinese rootstocks in response to drought stress. Photosynthetica 2012, 50, 401–410. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Layer | Texture | Volume Weight | FWC (1) WP (2) | AWC (3) | Corg Total N | pH (KCl) | Available Nutrients (4) P, K, and Mg |
|---|---|---|---|---|---|---|---|
| cm | - | g m−3 | % vol. | % vol. | % | - | Mg kg−1 |
| 0–30 | Silt loam | 1.45 | 31.9 | 18.6 | 1.53 | 6.32 | 124.20 |
| 13.3 | 0.16 | 260.80 | |||||
| 214.70 | |||||||
| 30–60 | Silt loam | 1.42 | 34.1 | 20.0 | 0.90 | 6.42 | 11.60 |
| 14.1 | 0.10 | 147.50 | |||||
| 191.20 | |||||||
| 60–90 | Silt loam | 1.45 | 33.0 | 18.1 | 0.33 | 6.37 | 1.60 |
| 14.9 | 0.05 | 121.80 | |||||
| 162.50 |
| Factor | p | Factor Level | Δ13C ‰ | Std. Deviation ‰ | ||
|---|---|---|---|---|---|---|
| Treatment | 0.002 | ET0 | 20.77 | 0.38 | b | |
| (Treat) | ET50 | 20.73 | 0.37 | b | ||
| ET75 | 20.80 | 0.36 | ab | |||
| ET100 | 20.95 | 0.38 | a | |||
| Term | 0.867 | Spring | 20.82 | 0.40 | ||
| Summer | 20.81 | 0.35 | ||||
| Year | <0.001 | 2019 | 20.80 | 0.21 | c | |
| 2020 | 20.45 | 0.27 | b | |||
| 2021 | 21.29 | 0.19 | a | |||
| 2022 | 20.72 | 0.22 | c | |||
| Average | 20.81 | 0.38 | ||||
| Year × Term | <0.001 | 2019 | Spring | 20.86 | 0.20 | b |
| 2019 | Summer | 20.73 | 0.21 | b | ||
| 2020 | Spring | 20.30 | 0.17 | c | ||
| 2020 | Summer | 20.61 | 0.27 | b | ||
| 2021 | Spring | 21.32 | 0.18 | a | ||
| 2021 | Summer | 21.26 | 0.21 | a | ||
| 2022 | Spring | 20.79 | 0.13 | b | ||
| 2022 | Summer | 20.64 | 0.27 | b | ||
| 2019 | ET0 | 20.80 | 0.24 | |||
| Year × Treat | 0.801 | 2019 | ET50 | 20.64 | 0.10 | |
| 2019 | ET75 | 20.81 | 0.23 | |||
| 2019 | ET100 | 20.95 | 0.15 | |||
| 2020 | ET0 | 20.36 | 0.28 | |||
| 2020 | ET50 | 20.44 | 0.27 | |||
| 2020 | ET75 | 20.39 | 0.16 | |||
| 2020 | ET100 | 20.63 | 0.33 | |||
| 2021 | ET0 | 21.23 | 0.15 | |||
| 2021 | ET50 | 21.23 | 0.27 | |||
| 2021 | ET75 | 21.24 | 0.14 | |||
| 2021 | ET100 | 21.46 | 0.12 | |||
| 2022 | ET0 | 20.72 | 0.22 | |||
| 2022 | ET50 | 20.61 | 0.19 | |||
| 2022 | ET75 | 20.76 | 0.27 | |||
| 2022 | ET100 | 20.78 | 0.20 | |||
| Treat × Term | 0.675 | |||||
| Treat × Term × Year | 0.495 | |||||
| Spring | Summer | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Factor | p | Factor Level | Δ13C ‰ | Std. Deviation ‰ | Factor | p | Factor Level | Δ13C ‰ | Std. Deviation ‰ | ||||
| Treatment | 0.039 | ET0 | 20.80 | 0.39 | ab | Treatment | 0.043 | ET0 | 20.75 | 0.39 | a | ||
| (Treat) | ET50 | 20.72 | 0.45 | b | (Treat) | ET50 | 20.73 | 0.29 | a | ||||
| ET75 | 20.83 | 0.39 | ab | ET75 | 20.78 | 0.36 | a | ||||||
| ET100 | 20.92 | 0.42 | a | ET100 | 20.99 | 0.34 | a | ||||||
| Year | <0.001 | 2019 | 20.86 | 0.20 | b | Year | <0.001 | 2019 | 20.86 | 0.20 | b | ||
| 2020 | 20.30 | 0.17 | c | 2020 | 20.30 | 0.17 | b | ||||||
| 2021 | 21.32 | 0.18 | a | 2021 | 21.32 | 0.18 | a | ||||||
| 2022 | 20.79 | 0.13 | b | 2022 | 20.79 | 0.13 | b | ||||||
| Average | 20.82 | 0.40 | Average | 20.81 | 0.35 | ||||||||
| Year × | 0.289 | 2019 | ET0 | 20.96 | 0.03 | Year × | 0.835 | 2019 | ET0 | 20.63 | 0.26 | ||
| Treat | 2019 | ET50 | 20.66 | 0.15 | Treat | 2019 | ET50 | 20.61 | 0.06 | ||||
| 2019 | ET75 | 20.85 | 0.27 | 2019 | ET75 | 20.78 | 0.23 | ||||||
| 2019 | ET100 | 20.99 | 0.16 | 2019 | ET100 | 20.91 | 0.17 | ||||||
| 2020 | ET0 | 20.29 | 0.36 | 2020 | ET0 | 20.42 | 0.23 | ||||||
| 2020 | ET50 | 20.21 | 0.08 | 2020 | ET50 | 20.67 | 0.14 | ||||||
| 2020 | ET75 | 20.31 | 0.08 | 2020 | ET75 | 20.48 | 0.19 | ||||||
| 2020 | ET100 | 20.38 | 0.02 | 2020 | ET100 | 20.88 | 0.30 | ||||||
| 2021 | ET0 | 21.19 | 0.18 | 2021 | ET0 | 21.27 | 0.15 | ||||||
| 2021 | ET50 | 21.37 | 0.06 | 2021 | ET50 | 21.10 | 0.35 | ||||||
| 2021 | ET75 | 21.24 | 0.21 | 2021 | ET75 | 21.24 | 0.06 | ||||||
| 2021 | ET100 | 21.49 | 0.14 | 2021 | ET100 | 21.42 | 0.11 | ||||||
| 2022 | ET0 | 20.76 | 0.03 | 2022 | ET0 | 20.67 | 0.34 | ||||||
| 2022 | ET50 | 20.66 | 0.15 | 2022 | ET50 | 20.56 | 0.26 | ||||||
| 2022 | ET75 | 20.92 | 0.10 | 2022 | ET75 | 20.61 | 0.32 | ||||||
| 2022 | ET100 | 20.82 | 0.01 | 2022 | ET100 | 20.74 | 0.31 | ||||||
| Factor | p | Factor Level | Δ13C ‰ | Std. Deviation ‰ | ||
|---|---|---|---|---|---|---|
| Year | <0.001 | 2019 | 21.74 | 0.66 | b | |
| 2020 | 21.30 | 0.57 | c | |||
| 2021 | 22.05 | 0.59 | a | |||
| 2022 | 21.78 | 0.54 | b | |||
| Treatment | 0.022 | ET0 | 21.69 | 0.56 | b | |
| (Treat) | ET50 | 21.66 | 0.63 | b | ||
| ET100 | 21.81 | 0.73 | a | |||
| Leaf position | <0.001 | L1 | 22.50 | 0.40 | a | |
| L2 | 21.78 | 0.41 | b | |||
| L3 | 21.94 | 0.39 | b | |||
| L4 | 21.31 | 0.36 | c | |||
| L5 | 21.07 | 0.47 | d | |||
| Average | 21.72 | 0.64 | ||||
| Year × Leaf | 0.001 | 2019 | L1 | 22.44 | 0.31 | ab |
| 2019 | L2 | 21.92 | 0.24 | bcd | ||
| 2019 | L3 | 22.20 | 0.31 | b | ||
| 2019 | L4 | 21.34 | 0.26 | ef | ||
| 2019 | L5 | 20.83 | 0.22 | fg | ||
| 2020 | L1 | 22.05 | 0.20 | bc | ||
| 2020 | L2 | 21.22 | 0.27 | ef | ||
| 2020 | L3 | 21.60 | 0.14 | de | ||
| 2020 | L4 | 21.02 | 0.33 | f | ||
| 2020 | L5 | 20.62 | 0.38 | g | ||
| 2021 | L1 | 22.93 | 0.23 | a | ||
| 2021 | L2 | 22.05 | 0.17 | bc | ||
| 2021 | L3 | 22.32 | 0.09 | b | ||
| 2021 | L4 | 21.63 | 0.21 | de | ||
| 2021 | L5 | 21.34 | 0.14 | ef | ||
| 2022 | L1 | 22.60 | 0.15 | ab | ||
| 2022 | L2 | 21.93 | 0.23 | bcd | ||
| 2022 | L3 | 21.62 | 0.17 | de | ||
| 2022 | L4 | 21.23 | 0.27 | ef | ||
| 2022 | L5 | 21.49 | 0.33 | de | ||
| Treat × Leaf | 0.409 | ET0 | L1 | 22.40 | 0.44 | |
| ET0 | L2 | 21.63 | 0.40 | |||
| ET0 | L3 | 21.88 | 0.33 | |||
| ET0 | L4 | 21.39 | 0.34 | |||
| ET0 | L5 | 21.14 | 0.35 | |||
| ET50 | L1 | 22.46 | 0.37 | |||
| ET50 | L2 | 21.79 | 0.38 | |||
| ET50 | L3 | 21.91 | 0.31 | |||
| ET50 | L4 | 21.18 | 0.31 | |||
| ET50 | L5 | 20.97 | 0.31 | |||
| ET100 | L1 | 22.66 | 0.39 | |||
| ET100 | L2 | 21.91 | 0.45 | |||
| ET100 | L3 | 22.02 | 0.53 | |||
| ET100 | L4 | 21.35 | 0.42 | |||
| ET100 | L5 | 21.10 | 0.69 | |||
| Year × Treat | 0.013 | |||||
| 3-way interaction | 0.617 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Haberle, J.; Raimanová, I.; Svoboda, P.; Moulik, M.; Mészáros, M.; Kurešová, G. The Effect of Increasing Irrigation Rates on the Carbon Isotope Discrimination of Apple Leaves. Agronomy 2023, 13, 1623. https://doi.org/10.3390/agronomy13061623
Haberle J, Raimanová I, Svoboda P, Moulik M, Mészáros M, Kurešová G. The Effect of Increasing Irrigation Rates on the Carbon Isotope Discrimination of Apple Leaves. Agronomy. 2023; 13(6):1623. https://doi.org/10.3390/agronomy13061623
Chicago/Turabian StyleHaberle, Jan, Ivana Raimanová, Pavel Svoboda, Michal Moulik, Martin Mészáros, and Gabriela Kurešová. 2023. "The Effect of Increasing Irrigation Rates on the Carbon Isotope Discrimination of Apple Leaves" Agronomy 13, no. 6: 1623. https://doi.org/10.3390/agronomy13061623