
Over-Expression of Soybean GmSAUL1 Enhances Disease Resistance in Nicotiana tabacum

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Transient Over-Expression of GmSAUL1a Induces HR Cell Death on the Leaves of Nicotiana benthamiana Plants
2.2. Stable Transgenic N. tabacum Plants Over-Expressing GmSAUL1a Displayed an Auto-Immune Phenotype
2.3. GmSAUL1a-Overexpressing N. tabacum Plants Exhibit Enhanced Resistance against Different Biotrophic Pathogens
2.4. Enhanced Disease Resistance of GmSAUL1a-Overexpressing Tobacco Plants Is Correlated with Elevated Callose Deposition
2.5. Over-Expressing GmSAUL1a in Tobacco Has Opposite Effects on the Activation of NtMPK4 and NtMPK6 Responding to flg22 Elicitation
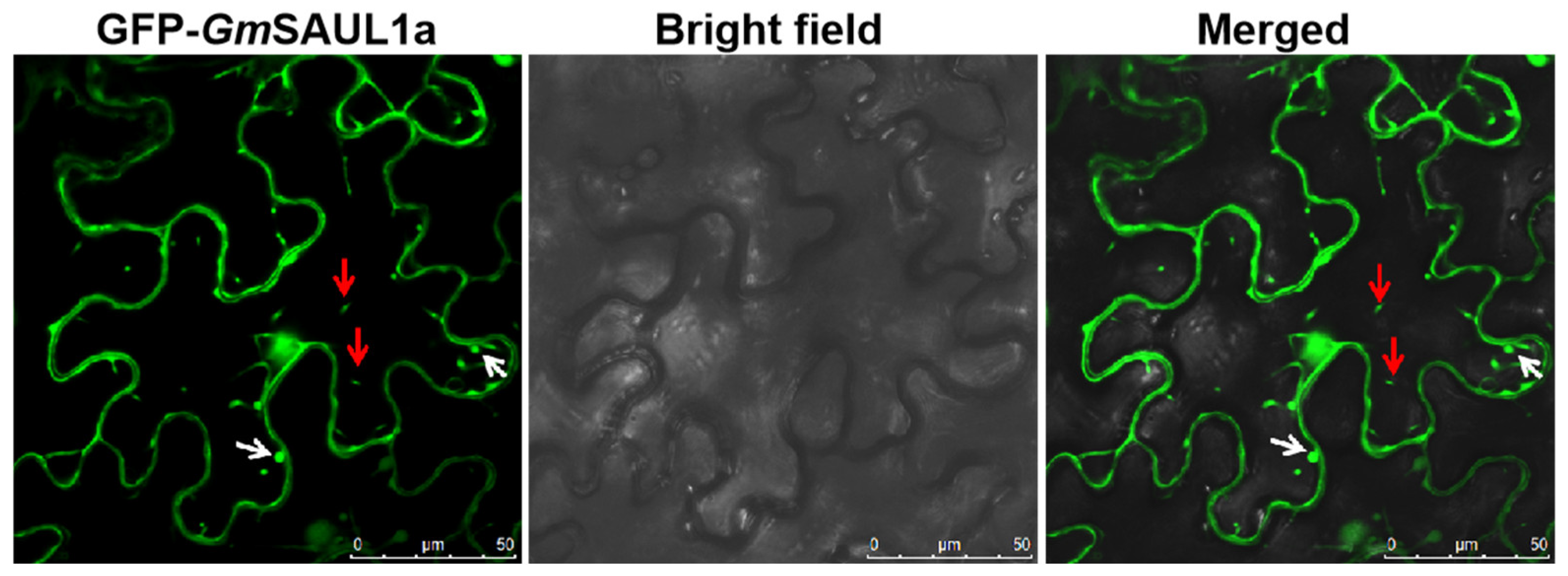
2.6. Subcellular Localization of GmSAUL1a
3. Discussion
3.1. The Function of SAUL1 Homologs Is Conserved across Plant Species
3.2. The Roles of GmSAUL1s in PTI and ETI
3.3. Silencing GmSAUL1a/1b in Soybean and Over-Expressing GmSAUL1a in N. tabacum Activate Immunity through Different Downstream MPKs
4. Materials and Methods
4.1. Plant Materials
4.2. MAPK Activity Assay
4.3. Histochemical Assays
4.4. SA Quantification
4.5. Generation of Transgenic N. tabacum Samsun (NN) Plants Over-Expressing GmSAUL1a
- GmSAUL1a-pE1776-F: aagGGTACCATGATGGCTGCGAGCTG
- GmSAUL1a-pE1776-R: ttgCCCGGGTTCATCCCATGTTTGGAAAGATT
4.6. The Pst DC3000 Infection
4.7. RNA Isolation and RT-qPCR
- NtPR1-F: GCTGAGGGAAGTGGCGATTTC
- NtPR1-R: CCTAGCACATCCAACACGAACC
- NtPR3-F: GACCCATCCAATTGACAAACCAA
- NtPR3-R: CCTGTGGTGTCATCCAGAACC
- NtPR5-F: GGCGATTGTGGCTCAAACC
- NtPR5-R: GAAATCTTGCTTCGTACCTGAGA
- NtUBQ5-F: CCTAACGGGTAAAACAATCAC
- NtUBQ5-R: AGCCATAAAAGTTCCAGCAC
4.8. Callose Deposition
4.9. GUS Staining
4.10. Sub-Cellular Localization of GFP-GmSAUL1a
- pGDG-F-Sal1: aaaGTC GAC ATGATGGCTGCGAGCTGG
- pGDG-R-BamH1: ttgGGA TCC TCATCCCATGTTTGGAAAGATTC
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Jones, J.D.G.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Chinchilla, D.; Zipfel, C.; Robatzek, S.; Kemmerling, B.; Nürnberger, T.; Jones, J.D.G.; Felix, G.; Boller, T. A flagellin-induced complex of the receptor FLS2 and BAK1 initiates plant defence. Nature 2007, 448, 497–500. [Google Scholar] [CrossRef] [PubMed]
- Boller, T.; Felix, G. A renaissance of elicitors: Perception of microbe-associated molecular patterns and danger signals by pattern-recognition receptors. Annu. Rev. Plant Biol. 2009, 60, 379–406. [Google Scholar] [CrossRef]
- Tena, G.; Boudsocq, M.; Sheen, J. Protein kinase signaling networks in plant innate immunity. Curr. Opin. Plant Biol. 2011, 14, 519–529. [Google Scholar] [CrossRef]
- Zhou, J.; Zhang, Y. Plant Immunity: Danger perception and signaling. Cell 2020, 181, 978–989. [Google Scholar] [CrossRef]
- Yuan, M.; Jiang, Z.; Bi, G. Pattern-recognition receptors are required for NLR-mediated plant immunity. Nature 2021, 592, 105–109. [Google Scholar] [CrossRef] [PubMed]
- Ngou, B.P.M.; Ahn, H.K.; Ding, P.; Jones, J.D. Mutual potentiation of plant immunity by cell-surface and intracellular receptors. Nature 2021, 592, 110–115. [Google Scholar] [CrossRef]
- Vierstra, R.D. The ubiquitin-26S proteasome system at the nexus of plant biology. Nat. Rev. Mol. Cell Biol. 2009, 10, 385–397. [Google Scholar] [CrossRef]
- Smalle, J.; Vierstra, R.D. The Ubiquitin 26S proteasome proteolytic pathway. Annu. Rev. Plant Biol. 2004, 55, 555–590. [Google Scholar] [CrossRef] [PubMed]
- Hershko, A.; Clechanover, A. The ubiquitin system. Annu. Rev. Biochem. 1998, 67, 425–479. [Google Scholar] [CrossRef]
- Cheng, Y.T.; Li, X. Ubiquitination in NB-LRR-mediated immunity. Curr. Opin. Plant Biol. 2012, 15, 392–399. [Google Scholar] [CrossRef] [PubMed]
- Zeng, L.; Vega-Sanchez, M.E.; Zhu, T.; Wang, G.L. Ubiquitination-mediated protein degradation and modification: An emerging theme in plant-microbe interactions. Cell Res. 2006, 16, 413–426. [Google Scholar] [CrossRef]
- Zhou, B.; Zeng, L. Conventional and unconventional ubiquitinnation in plant immunity. Mol. Plant Pathol. 2017, 18, 1313–1330. [Google Scholar] [CrossRef]
- Zeng, L.; Qu, S.; Bordeos, A.; Yang, C.; Baraoidan, M.; Yan, H.; Xie, Q.; Nahm, B.H.; Leung, H.; Wang, G.L. Soptted leaf 11, a negative regulator of plant cell death and defense, encodes a U-box/armadillo repeat protein endowed with E3 ubiquitin ligase activity. Plant Cell 2004, 16, 2795–2808. [Google Scholar] [CrossRef]
- Raab, S.; Drechsel, G.; Zarepour, M.; Hartung, W.; Koshiba, T.; Bittner, F.; Hoth, S. Identification of a novel E3 ubiquitin ligase that is required for suppression of premature senescence in Arabidopsis. Plant J. 2009, 59, 39–51. [Google Scholar] [CrossRef]
- Vogelmann, K.; Drechsel, G.; Bergler, J.; Subert, C.; Philippar, K.; Soll, J.; Engelmann, J.C.; Engelsdorf, T.; Voll, L.M.; Hoth, S. Early senescence and cell death in Arabidopsis saul1 mutants involves the PAD4-dependent salicylic acid pathway. Plant Physiol. 2012, 159, 1477–1487. [Google Scholar] [CrossRef] [PubMed]
- Disch, E.M.; Tong, M.; Kotur, T.; Koch, G.; Wolf, C.A.; Li, X.; Hoth, S. Membrane-associated ubiquitin ligase SAUL1 suppresses temperature-and humidity-dependent autoimmunity in Arabidopsis. Mol. Plant-Microbe Interact. 2016, 29, 69–80. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.H.; Lee, I.C.; Kim, J.; Kim, J.H.; Chung, E.H.; Kim, H.J.; Park, S.J.; Kim, Y.M.; Kang, S.K.; Nam, H.G.; et al. NORE1/SAUL1 integrates temperature-dependent defense programs involving SGT1b and PAD4 pathways and leaf senescence in Arabidopsis. Physiol. Plant. 2016, 158, 180–199. [Google Scholar] [CrossRef]
- Tong, M.; Kotur, T.; Liang, W.; Vogelmann, K.; Kleine, T.; Leister, D.; Brieske, C.; Yang, S.; Ludke, D.; Wiermer, M.; et al. E3 ligase SAUL1 serves as a positive regulator of PAMP-triggered immunity and its homeostasis is monitored by immune receptor SOC3. New Phytol. 2017, 215, 1516–1532. [Google Scholar] [CrossRef]
- Liang, W.; van Wersch, S.; Tong, M.; Li, X. TIR-NB-LRR immune receptor SOC3 pairs with truncated TIR-NB protein CHS1 or TN2 to monitor the homeostasis of E3 ligase SAUL1. New Phytol. 2019, 221, 2054–2066. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, Y.; Liu, J.; Ding, Y.; Wang, S.; Zhang, X.; Liu, Y.; Yang, S. Temperature-dependent autoimmunity mediated by chs1 requires its neighboring TNL gene SOC3. New Phytol. 2016, 213, 1330–1345. [Google Scholar] [CrossRef]
- Colcombet, J.; Hirt, H. Arabidopsis MAPKs: A complex signal-ling network involved in multiple biological processes. Biochem. J. 2008, 413, 217–226. [Google Scholar] [CrossRef]
- Pitzschke, A.; Schikora, A.; Hirt, H. MAPK cascade signalling networks in plant defence. Curr. Opin. Plant. Biol. 2009, 12, 421–426. [Google Scholar] [CrossRef]
- Pitzschke, A.; Djamei, A.; Bitton, F.; Hirt, H. A major role of the MEKK1-MKK1/2-MPK4 pathway in ROS signalling. Mol. Plant 2009, 2, 120–137. [Google Scholar] [CrossRef]
- Meng, X.; Zhang, S. MAPK cascades in plant disease resistance signaling. Annu. Rev. Phytopathol. 2013, 51, 245–266. [Google Scholar] [CrossRef]
- Zhang, S.; Klessig, D.F. Resistance gene N-mediated de novo synthesis and activation of a tobacco mitogen-activated protein kinase by Tobacco mosaic virus infection. Proc. Natl. Acad. Sci. USA 1998, 95, 7433–7438. [Google Scholar] [CrossRef] [PubMed]
- Ren, D.; Liu, Y.; Yang, K.Y.; Han, L.; Mao, G.; Glazebrook, J.; Zhang, S. A fungal-responsive MAPK cascade regulates phytoalexin biosynthesis in Arabidopsis. Proc. Natl. Acad. Sci. USA 2008, 105, 5638–5643. [Google Scholar] [CrossRef] [PubMed]
- Mao, G.; Meng, X.; Liu, Y.; Zheng, Z.; Chen, Z.; Zhang, S. Phosphorylation of a WRKY transcription factor by two pathogen-responsive MAPKs drives phytoalexin biosynthesis in Arabidopsis. Plant Cell 2011, 23, 1639–1653. [Google Scholar] [CrossRef]
- Petersen, M.; Brodersen, P.; Naested, H.; Andreasson, E.; Lindhart, U.; Johansen, B.; Nielsen, H.B.; Lacy, M.; Austin, M.J.; Parker, J.E.; et al. Arabidopsis MAP kinase 4 negatively regulates systemic acquired resistance. Cell 2000, 103, 1111–1120. [Google Scholar] [CrossRef] [PubMed]
- Qiu, J.L.; Zhou, L.; Yun, B.W.; Nielsen, H.B.; Fiil, B.K.; Petersen, K.; Mackinlay, J.; Loake, G.J.; Mundy, J.; Morris, P.C. Arabidopsis mitogen-activated protein kinase kinases MKK1 and MKK2 have overlapping functions in defense signaling mediated by MEKK1, MPK4, and MKS1. Plant Physiol. 2008, 148, 212–222. [Google Scholar] [CrossRef]
- Gao, M.; Liu, J.; Bi, D.; Zhang, Z.; Cheng, F.; Chen, S.; Zhang, Y. MEKK1, MKK1/MKK2 and MPK4 function together in a mitogen-activated protein kinase cascade to regulate innate immunity in plants. Cell Res. 2008, 18, 1190–1198. [Google Scholar] [CrossRef] [PubMed]
- Kong, Q.; Qu, N.; Gao, M.; Zhang, Z.; Ding, X.; Yang, F.; Li, Y.; Dong, O.X.; Chen, S.; Li, X.; et al. The MEKK1-MKK1/MKK2-MPK4 kinase cascade negatively regulates immunity mediated by a mitogen-activated protein kinase kinase kinase in Arabidopsis. Plant Cell 2012, 24, 2225–2236. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.Z.; Horstman, H.D.; Braun, E.; Graham, M.A.; Zhang, C.; Navarre, D.; Qiu, W.L.; Lee, Y.; Nettleton, D.; Hill, J.H. Soybean homologs of MPK4 negatively regulate defense responses and positively regulate growth and development. Plant Physiol. 2011, 157, 1363–1378. [Google Scholar] [CrossRef]
- Liu, J.Z.; Braun, E.; Qiu, W.L.; Shi, Y.F.; Marcelino-Guimars, F.C.; Navarre, D.; Hill, J.H.; Whitham, S.A. Positive and negative roles for soybean MPK6 in regulating defense responses. Mol. Plant-Microbe Interact. 2014, 27, 824–834. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.Z.; Graham, M.A.; Pedley, K.F.; Whitham, S.A. Gaining insight into soybean defense responses using functional genomics approaches. Brief. Funct. Genom. 2015, 14, 283–290. [Google Scholar] [CrossRef]
- Liu, J.Z.; Fang, Y.; Pang, H. The current status of the soybean-soybean mosaic virus (SMV) pathosystem. Front. Microbiol. 2016, 7, 1906. [Google Scholar] [CrossRef]
- Liu, J.Z.; Whitham, S.A. Over-expression of a nuclear-localized DnaJ domain-containing HSP40 from soybean reveals its roles in cell death and disease resistance. Plant J. 2013, 74, 110–121. [Google Scholar] [CrossRef]
- Xu, H.Y.; Zhang, C.; Li, Z.C.; Wang, Z.R.; Jiang, X.X.; Shi, Y.F.; Tian, S.N.; Braun, E.; Mei, Y.; Qiu, W.L.; et al. The MAPK kinase kinase GmMEKK1 regulates cell death and defense responses. Plant Physiol. 2018, 178, 907–922. [Google Scholar] [CrossRef]
- Tian, S.N.; Liu, D.D.; Zhong, C.L.; Xu, H.Y.; Yang, S.; Fang, Y.; Ran, J.; Liu, J.Z. Silencing GmFLS2 enhances the susceptibility of soybean to bacterial pathogen through attenuating the activation of GmMAPK signaling pathway. Plant Sci. 2020, 292, 110386. [Google Scholar] [CrossRef]
- Hashimi, S.M.; Wu, N.N.; Ran, J.; Liu, J.Z. Silencing Autophagy-Related Gene 2 (ATG2) Results in Accelerated Senescence and Enhanced Immunity in Soybean. Int. J. Mol. Sci. 2021, 22, 11749. [Google Scholar]
- Liu, D.D.; Lan, H.J.; Masoud, H.S.; Ye, M.Y.; Dai, X.Y.; Zhong, C.L.; Tian, S.N.; Liu, J.Z. Silencing GmBIR1 in Soybean Results in Activated Defense Responses. Int. J. Mol. Sci. 2022, 23, 7450. [Google Scholar]
- Li, J.M.; Ye, M.Y.; Wang, C.; Ma, X.H.; Wu, N.N.; Zhong, C.L.; Zhang, Y.; Cheng, N.; Nakata, P.A.; Zeng, L.; et al. Soybean GmSAUL1, a Bona Fide U-Box E3 Ligase, Negatively Regulates Immunity Likely through Repressing the Activation of GmMPK3. Int. J. Mol. Sci. 2023, 24, 6240. [Google Scholar]
- Whitham, S.; McCormick, S.; Baker, B. The N gene of tobacco confers resistance to tobacco mosaic virus in transgenic tomato. Proc. Natl. Acad. Sci. USA 1996, 93, 8776–8781. [Google Scholar]
- Zhu, Y.; Qian, W.; Hua, J. Temperature Modulates Plant Defense Responses through NB-LRR Proteins. PLoS Pathog. 2010, 6, e1000844. [Google Scholar] [CrossRef]
- Whitham, S.; Dinesh-Kumar, S.P.; Choi, D.; Hehl, R.; Corr, C.; Baker, B. The product of the tobacco mosaic virus resistance gene N: Similarity to toll and the interleukin-1 receptor. Cell 1994, 78, 1101–1115. [Google Scholar] [CrossRef]
- Meshi, T.; Takamatsu, N.; Ohno, T.; Okada, Y. Molecular cloning of the complementary DNA copies of the common and cowpea strains of tobacco mosaic virus RNA. Virology 1982, 118, 4–75. [Google Scholar]
- Vogelmann, K.; Subert, C.; Danzberger, N.; Drechsel, G.; Bergler, J.; Kotur, T.; Burmester, T.; Hoth, S. Plasma membrane-association of SAUL1-type plant U-box armadillo repeat proteins is conserved in land plants. Front. Plant Sci. 2014, 5, 37. [Google Scholar] [CrossRef]
- Tao, K.; Waletich, J.R.; Wise, H.; Arredondo, F.; Tyler, B.M. Tethering of multi-vesicular bodies and the tonoplast to the plasma membrane in plants. Front. Plant Sci. 2019, 10, 636. [Google Scholar] [CrossRef]
- Knop, J.; Lienemann, T.; El-Kilani, H.; Falke, S.; Krings, C.; Sindalovskaya, M.; Bergler, J.; Betzel, C.; Hoth, S. Structural Features of a Full-Length Ubiquitin Ligase Responsible for the Formation of Patches at the Plasma Membrane. Int. J. Mol. Sci. 2021, 22, 9455. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Liu, Y.; Bruffett, K.; Lee, J.; Hause, G.; Walker, J.; Zhang, S. Haplo-insufficiency of MPK3 in MPK6 mutant background uncovers a novel function of these two MAPKs in Arabidopsis ovule development. Plant Cell 2008, 20, 602–613. [Google Scholar] [CrossRef] [PubMed]
- Menke, F.L.; van Pelt, J.A.; Pieterse, C.M.; Klessig, D.F. Silencing of the mitogen-activated protein kinase MPK6 compromises disease resistance in Arabidopsis. Plant Cell 2004, 16, 897–907. [Google Scholar] [CrossRef]
- Zhang, S.; Liu, Y. Activation of salicylic acid-induced protein kinase, a mitogen-activated protein kinase, induces multiple defense responses in tobacco. Plant Cell 2001, 13, 1877–1889. [Google Scholar]
- Ren, D.; Yang, H.; Zhang, S. Cell death mediated by MAPK is associated with hydrogen peroxide production in Arabidopsis. J. Biol. Chem. 2002, 277, 559–565. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhao, L.; Zhao, J.; Li, Y.; Wang, J.; Guo, R.; Gan, S.; Liu, C.J.; Zhang, K. S5H/DMR6 Encodes a Salicylic Acid 5-Hydroxylase That Fine-Tunes Salicylic Acid Homeostasis. Plant Physiol. 2017, 175, 1082–1093. [Google Scholar] [CrossRef]
- Leister, R.T.; Dahlbeck, D.; Day, B.; Li, Y.; Chesnokova, O.; Staskawicz, B.J. Molecular genetic evidence for the role of SGT1 in the intramolecular complementation of Bs2 protein activity in Nicotiana benthamiana. Plant Cell 2005, 17, 1268–1278. [Google Scholar] [CrossRef]
- Horsch, R.B.; Fry, J.E.; Hofmann, N.L.; Eichholtz, D.; Rogers, S.G.; Fraley, R.T. A simple and general method for transferring genes into plants. Science 1985, 227, 1229–1231. [Google Scholar] [CrossRef]
- Li, Z.C.; Ren, Q.W.; Guo, Y.; Ran, J.; Ren, X.T.; Wu, N.N.; Liu, X.; Liu, J.Z. Dual roles of GSNOR1 in cell death and immunity in tetraploid Nicotiana tabacum. Front. Plant Sci. 2021, 12, 596234. [Google Scholar] [CrossRef] [PubMed]
- Hinge, V.R.; Chavhan, R.L.; Kale, S.P.; Suprasanna, P.; Kadam, U.S. Engineering Resistance against Viruses in Field Crops Using CRISPR-Cas9. Curr. Genom. 2021, 22, 214–231. [Google Scholar] [CrossRef] [PubMed]
- Heese, A.; Hann, D.R.; Gimenez-Ibanez, S.; Jones, A.M.; He, K.; Li, J.; Schroeder, J.I.; Peck, S.C.; Rathjen, J.P. The receptor-like kinase SERK3/BAK1 is a central regulator of innate immunity in plants. Proc. Natl. Acad. Sci. USA 2007, 104, 12217–12222. [Google Scholar] [CrossRef]
- Jefferson, R.A.; Kavanagh, T.A.; Bevan, M.W. GUS fusions: Beta-glucuronidase as a sensitive and versatile gene fusion marker in higher plants. EMBO J. 1987, 6, 3901–3907. [Google Scholar] [CrossRef]
- Goodin, M.; Dietzgen, R.G.; Schichnes, D.; Ruzin, S.; Jackson, A.O. pGD vectors: Versatile tools for the expression of green and red fluorescent protein fusions in Agroinfiltrated plant leaves. Plant J. 2022, 31, 375–383. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.Z.; Blancaflor, E.B.; Nelson, R.S. The tobacco mosaic virus 126-kilodalton protein, a constituent of the virus replication complex, alone or within the complex aligns with and traffics along microfilaments. Plant Physiol. 2005, 138, 1853–1865. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.-M.; Ye, M.-Y.; Ma, X.-H.; Wu, N.-N.; Liu, J.-Z. Over-Expression of Soybean GmSAUL1 Enhances Disease Resistance in Nicotiana tabacum. Agronomy 2023, 13, 1533. https://doi.org/10.3390/agronomy13061533
Li J-M, Ye M-Y, Ma X-H, Wu N-N, Liu J-Z. Over-Expression of Soybean GmSAUL1 Enhances Disease Resistance in Nicotiana tabacum. Agronomy. 2023; 13(6):1533. https://doi.org/10.3390/agronomy13061533
Chicago/Turabian StyleLi, Jun-Mei, Mei-Yan Ye, Xiao-Han Ma, Ni-Ni Wu, and Jian-Zhong Liu. 2023. "Over-Expression of Soybean GmSAUL1 Enhances Disease Resistance in Nicotiana tabacum" Agronomy 13, no. 6: 1533. https://doi.org/10.3390/agronomy13061533