
Organic or Inorganic Amendments Influence Microbial Community in Rhizosphere and Decreases the Incidence of Tomato Bacterial Wilt
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Experimental Design
2.2.1. Tomato Seedlings and Transplanting
2.2.2. Field Trial
2.2.3. Tomato Plant and Soil Sample Collection
2.2.4. Determination of Physical and Chemical Properties of Soil and Plants
2.2.5. DNA Extraction and High-Throughput Sequencing
2.2.6. Ralstonia Solanacearum Quantitative PCR
2.3. Statistical Analysis
3. Results
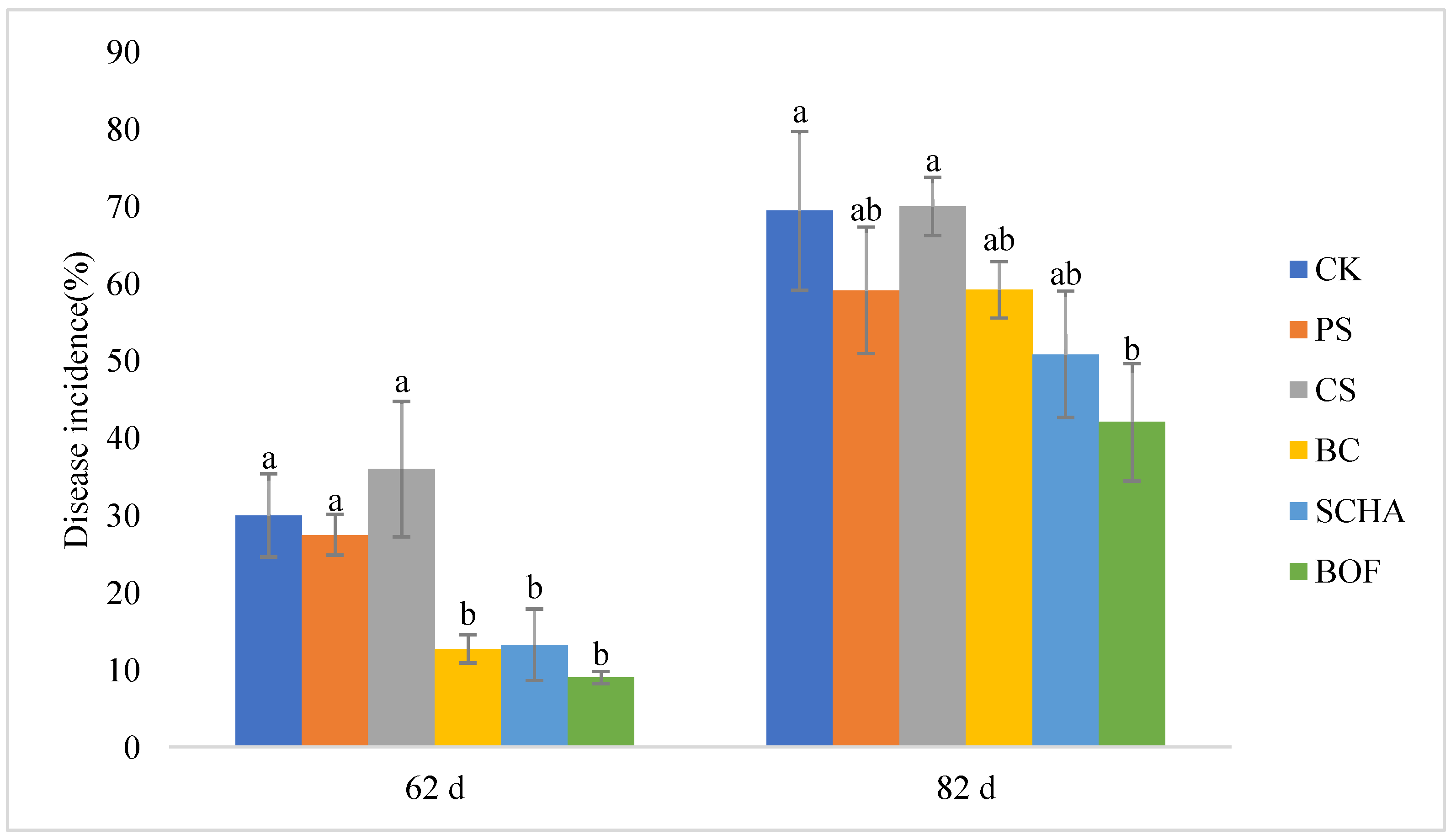
3.1. Effects of Different Soil Control Measures on the Incidence of Tomato Bacterial Wilt
3.2. Quantity of R. solanacearum in Soil and Roots
3.3. Variance Analysis of Disease Incidence and Soil Physicochemical Properties among Different Treatments, Spearman Correlation Analysis of Physicochemical Properties, Morbidity, and Number of Pathogenic Bacteria
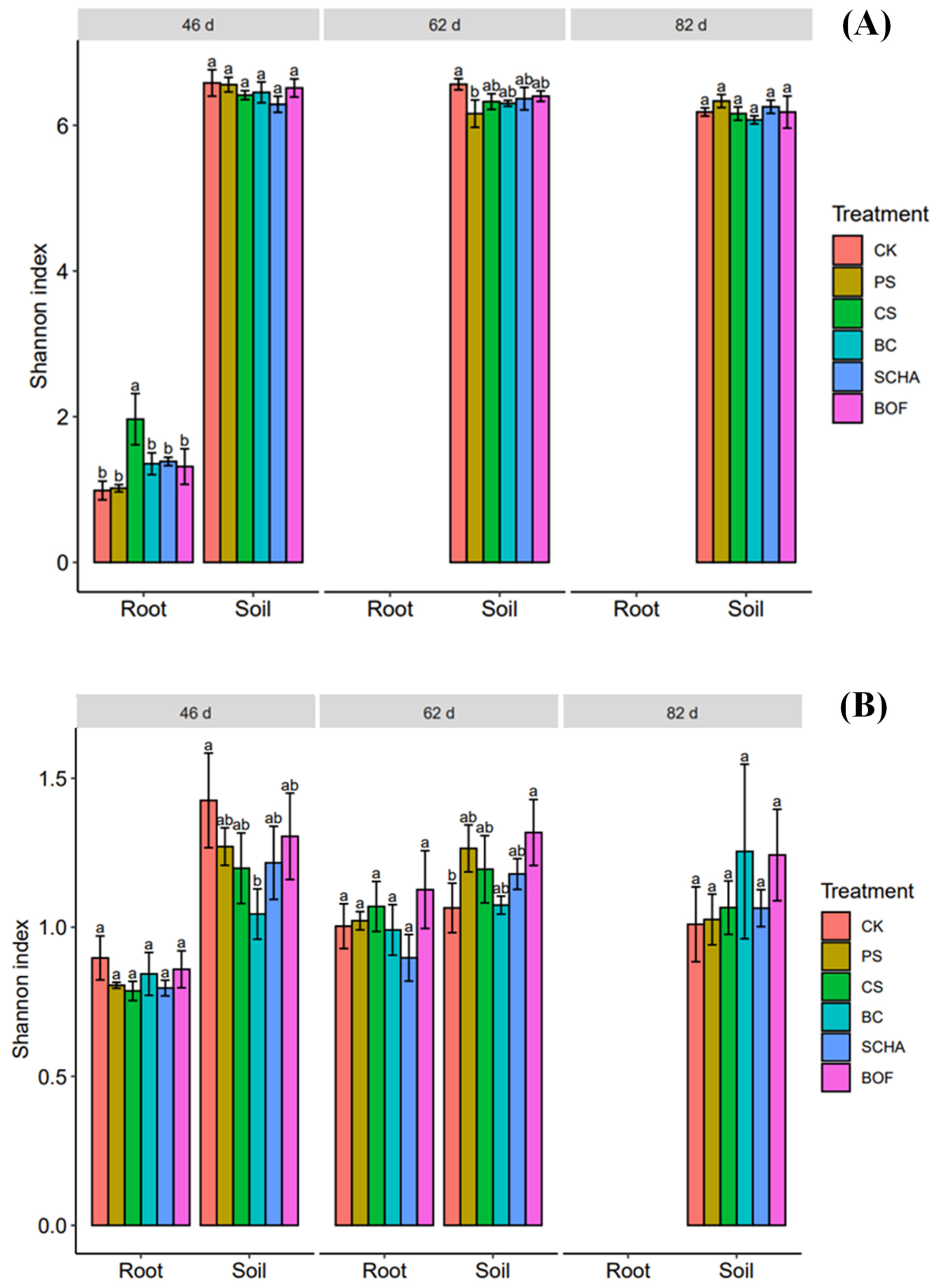
3.4. Soil and Root Microbial Community Structure
3.5. Correlation Analysis
4. Discussion
4.1. Effect of Amendments on Incidence of Tomato Bacterial Wilt, the Number of R. solanacearum and Chemical Factors in Soil and Plant Parts
4.2. Effect of Amendments on Microbial Diversity in Rhizosphere Soil and Root
4.3. Effect of Amendments on the Interaction between Soil Microbes and Tomato Bacterial Wilt
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Khan, S.; Hanjra, M.A.; Mu, J.X. Water management and crop production for food security in China: A review. Agric. Water Manag. 2009, 96, 349–360. [Google Scholar] [CrossRef]
- Irikiin, Y.; Nishiyama, M.; Otsuka, S.; Senoo, K. Rhizobacterial community-level, sole carbon source utilization pattern affects the delay in the bacterial wilt of tomato grown in rhizobacterial community model system. Appl. Soil Ecol. 2006, 34, 27–32. [Google Scholar] [CrossRef]
- Trigalet, A.; Trigalet, D. Invasiveness in tomato plant of Tn5-induced avirulent mutants of Pseudomonas solanacearun. Physiol. Mol. Plant Pathol. 1986, 28, 423–430. [Google Scholar] [CrossRef]
- Trigalet, A.; Trigalet-Demery, D. Use of avirulent mutants of Pseudomonas solanacearum for the biological control of bacterial wilt of tomato plants. Physiol. Mol. Plant Pathol. 1990, 36, 27–38. [Google Scholar] [CrossRef]
- Yuliar; Nion, Y.A.; Toyota, K.K. Recent trends in control methods for bacterial wilt diseases caused by Ralstonia solanacearum. Microbes Environ. 2015, 30, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Dong, Y.H.; Li, J.G. Effects of Soil Amendments on Suppression of Bacterial Wilt of Tomato. J. Agro-Environ. Sci. 2013, 32, 1368–1374. [Google Scholar]
- Bi, J.J.; Song, A.L.; Li, S.D.; Chen, M.S.; Wang, Y.N.; Wang, S.; Si, Z.Y.; Wang, E.Z.; Zhang, J.Y.; Asante-Badu, B.; et al. Plant physiology, microbial community, and risks of multiple fungal diseases along a soil nitrogen gradien. Appl. Soil Ecol. 2022, 175, 104445. [Google Scholar] [CrossRef]
- Amtmann, A.; Troufflard, S.; Armengaud, P. The effect of potassium nutrition on pest and disease resistance in plants. Physiol. Plant 2008, 133, 682–691. [Google Scholar] [CrossRef]
- He, X.; Jiang, J.; Dong, Y. Effect of potassium on resistance and mechanism of tomato blight. J. Anhui Agric. Sci. 2017, 45, 154–156. [Google Scholar]
- Jiang, J.; Li, J.; Dong, Y. Effect of calcium nutrition on resistance of tomato against bacterial wilt induced by Ralstonia solanacearum. Eur. J. Plant Pathol. 2013, 136, 547–555. [Google Scholar] [CrossRef]
- Wang, L.; Cai, K.Z.; Chen, Y.T.; Wang, G.P. Silicon-mediated tomato resistance against Ralstonia solanacearum is associated with modification of soil microbial community structure and activity. Biol. Trace Elem. Res. 2013, 152, 275–283. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Hou, Y.G.; Huang, D.P.; Hao, Z.X.; Wang, X.F.; Wei, Z.; Jousset, A.; Tan, S.Y.; Xu, D.B.; Shen, Q.R. Application of biochar reduces Ralstonia solanacearum infection via effects on pathogen chemotaxis, swarming motility, and root exudate adsorption. Plant Soil 2017, 415, 269–281. [Google Scholar] [CrossRef]
- Gao, Y.; Lu, Y.; Lin, W.P.; Tian, J.H.; Cai, K.Z. Biochar suppresses Bacterial Wilt of tomato by improving soil chemical properties and shifting soil microbial community. Microorganisms 2019, 7, 676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, Y.; Liao, Z.; Zhang, J.; Kong, W.; He, C. Effects of ecological organic fertilizer on tomato blight and soil microbial diversity. Appl. Ecol. 2003, 03, 349–353. [Google Scholar]
- Wu, X.J.; Liu, Y.; Shang, Y.W.; Liu, D.; Liesack, W.; Cui, Z.L.; Peng, J.J.; Zhang, F.S. Peat-vermiculite alters microbiota composition towards increased soil fertility and crop productivity. Plant Soil 2002, 470, 21–34. [Google Scholar] [CrossRef]
- Wu, X.J.; Rensing, C.; Han, D.F.; Ke-Qing, X.; Dai, Y.X.; Tang, Z.X.; Liesack, W.; Peng, J.J.; Cui, Z.L.; Zhang, F.S. Genome-Resolved metagenomics reveals distinct phosphorus acquisition strategies between soil microbiomes. Msystems 2022, 7, e0110721. [Google Scholar] [CrossRef]
- Cui, J.; Wang, W.Q.; Peng, Y.; Zhou, F.W.; He, D.Y.; Wang, J.J.; Chang, Y.J.; Yang, J.; Wang, W.; Yao, D.R.; et al. Effects of simulated Cd deposition on soil Cd availability, microbial response, and crop Cd uptake in the passivation-remediation process of Cd-contaminated purple soil. Sci. Total Environ. 2019, 683, 782–792. [Google Scholar] [CrossRef]
- Egamberdieva, D.; Wirth, S.; Li, L.; Abd-Allah, E.F.; Lindstrom, K. Microbial cooperation in the rhizosphere improves liquorice growth under salt stress. Bioengineered 2017, 8, 433–438. [Google Scholar] [CrossRef] [Green Version]
- Ge, Y.Y.; Wang, Q.L.; Wang, L.; Liu, W.X.; Liu, X.Y.; Huang, Y.J.; Christie, P. Response of soil enzymes and microbial communities to root extracts of the alien Alternanthera philoxeroides. Arch. Agron. Soil Sci. 2017, 64, 708–717. [Google Scholar] [CrossRef]
- Jaiswal, A.K.; Elad, Y.; Paudel, I.; Graber, E.R.; Cytryn, E.; Frenkel, O. Linking the belowground microbial composition, diversity and activity to soilborne disease suppression and growth promotion of tomato amended with biochar. Sci. Rep. 2017, 117, 204. [Google Scholar] [CrossRef] [Green Version]
- Rogovska, N.; Laird, D.; Leandro, L.; Aller, D. Biochar ect on severity of soybean root disease caused by Fusarium virguliforme. Plant Soil 2017, 413, 111–126. [Google Scholar] [CrossRef]
- Hayward, A.C. Biology and epidemiology of bacterial wilt caused by Pseudomonas solanacearum. Annu. Rev. Phytopathol. 1991, 29, 65–87. [Google Scholar] [CrossRef] [PubMed]
- Solke, H.B. Book Review: Bacterial Wilt Disease and the Ralstonia solanacearum Species Complex. Eur. J. Plant Pathol. 2006, 114, 227–228. [Google Scholar]
- Wei, Z. Effect and Mechanism of Bio-Organic Fertilizer in the Prevention and Control of Soil-Borne Tomato Blight; D. Nanjing Agricultural University: Nanjing, China, 2012. [Google Scholar]
- Shen, Z.; Huang, Y.; Cao, Y.; Wang, D.; Liu, H.; Li, R.; Shen, Q. Comparison of soil bacterial community characteristics of healthy and verticilage-affected tomato. Soils 2021, 53, 5–12. [Google Scholar]
- Bo, S. Soil Agrochemical Analysis, 3rd ed.; M. China Agriculture Press: Beijing, China, 2000. [Google Scholar]
- Song, A.L.; Li, Z.M.; Liao, Y.L.; Liang, Y.C.; Wang, E.Z.; Wang, S.; Bi, J.J.; Si, Z.Y.; Lu, Y.H.; Nie, J.; et al. Soil bacterial communities interact with silicon fraction transformation and promote rice yield after long-term straw return. Soil Ecol. Lett. 2021, 3, 395–408. [Google Scholar] [CrossRef]
- Fan, F.; Yu, B.; Wang, B.; George, T.S.; Yin, H.; Xu, D.; Li, D.; Song, A. Microbial mechanisms of the contrast residue decomposition and priming effect in soils with different organic and chemical fertilization histories. Soil Biol. Biochem. 2019, 135, 213–221. [Google Scholar] [CrossRef]
- Lin, W.; Jiang, N.; Li, P.; Fan, X.; Gao, Y.; Wang, G.; Cai, K. Silicon impacts on soil microflora under Ralstonia Solanacearum inoculation. J. Agric. Sci. 2020, 19, 251–264. [Google Scholar] [CrossRef]
- Wang, F.; Li, Z.; Chen, Y.; Yang, S.; Xu, Y. Research progress on the effect and mechanism of calcium inhibition of plant diseases. J. Biotech. Bulletin. 2017, 33, 1–7. [Google Scholar]
- Teixeira, P.J.P.; Colaianni, N.R.; Fitzpatrick, C.R.; Dang, J.L. Beyond pathogens: Microbiota interactions with the plant immune system. J. Curr. Opin. Microbiol. 2019, 49, 7–17. [Google Scholar] [CrossRef]
- Yamazaki, H. Relation between Resistance to Bacterial Wilt and Calcium Nutrition in Tomato Seedlings. Jpn. Agric. Res. Q. 2001, 35, 163–169. [Google Scholar] [CrossRef] [Green Version]
- Graber, E.R.; Frenkel, O.; Jaiswal, A.K.; Elad, Y. How may biochar influence severity of diseases caused by soilborne pathogens? Carbon Manag. 2014, 5, 169–183. [Google Scholar] [CrossRef]
- De, M.; Erika, V.L.; Neyla, T.; De, S.L.; José, R.; Pinto, K.M.S.; Da, C.; Diogo, P.; Da, F.; Rafaela, F.; et al. Biochar from different sources against tomato bacterial wilt disease caused by Ralstonia solanacearum. J. Soil Sci. Plant Nut. 2021, 22, 1–9. [Google Scholar]
- Claudio, M.; Flavio, F.; Tania, S.; Paolo, S.; Federica, G.; Davide, M. Organic amendment effectively recovers soil functionality in degraded vineyards. Eur. J. Agron. 2018, 101, 210–221. [Google Scholar]
- Jiang, L.H.; Bonkowski, M.; Luo, L.; Kardol, P.; Zhang, Y.; Chen, X.Y.; Li, D.M.; Xiao, Z.G.; Hu, F.; Liu, M.Q. Combined addition of chemical and organic amendments enhances plant resistance to aboveground herbivores through increasing microbial abundance and diversity. Biol. Fert. Soils. 2020, 56, 1007–1022. [Google Scholar] [CrossRef]
- Pastrana, A.M.; Shea, E.A.; Fernandez-Bayo, J.D.; Allison, B.; Watson, D.C.; Toniato, J.L.; Gordon, T.R.; Simmons, C.W. Impact of biosolarization with almond hull and shell amendments for the control of Fusarium oxysporum f. sp. lactucae in a lettuce/tomato cropping system. Crop Prot. 2022, 152, 105856. [Google Scholar] [CrossRef]
- Zhang, Y.; Hu, A.; Zhou, J.; Zhang, W.; Li, P. Comparison of bacterial communities in soil samples with and without tomato bacterial wilt caused by Ralstonia solanacearum species complex. BMC Microbiol. 2020, 20, 89. [Google Scholar] [CrossRef]
- Wang, Z. Research on the Occurrence Conditions and Control of Bacterial Wilt in Hydroponic Tomatoes; Huazhong Agricultural University: Wuhan, China, 2008. [Google Scholar]
- Yu, W.; Yi, Y.; Yang, L. Effects of calcium and nitrogen content in soil on resistance to tomato fusarium wilt. Soil Fertil. Sci. China 2016, 134–140. [Google Scholar]
- Huber, D.M. Fertilizers and soil-borne diseases. Soil Use Manag. 1990, 6, 168–173. [Google Scholar] [CrossRef]
- Yang, T.; Wang, Y.; Wang, J.; Wei, Z.; Dong, C.; Shen, Q.; Xu, Y. Study on the prevention and control of tomato soil blight and its pro-growth effect of different matrix bio-organic fertilizers. Soil 2021, 53, 961–968. [Google Scholar]
- Gong, Z.; Gao, X.; Huang, Y.; Li, R.; Wang, D.; Shen, Q. Study on continuous application of bio-organic fertilizer to improve cucumber yield and quality in facilitie. J. Nanjing Agric. Univ. 2016, 39, 777–783. [Google Scholar]
- Zhuang, Z.; Gao, J.; Chui, J.; Wang, L.; Xiao, Y. Effects of seaweed bio-organic fertilizer on tomato growth, soil organic matter and pH. Anhui Agric. Sci. 2017, 45, 104–106. [Google Scholar]
- Shen, Z.; Wang, B.; Zhu, J.; Hu, H.; Tao, C.; Ou, Y.; Deng, X.; Ling, N.; Li, R.; Shen, Q. Lime and ammonium carbonate fumigation coupled with bio-organic fertilizer application steered banana rhizosphere to assemble a unique microbiome against Panama disease. Microb. Biotechnol. 2019, 12, 515–527. [Google Scholar] [CrossRef] [PubMed]
- Tao, C.Y.; Li, R.; Xiong, W.; Shen, Z.Z.; Liu, S.S.; Wang, B.B.; Ruan, Y.Z.; Geisen, S.; Shen, Q.R.; Kowalchuk, G.A. Bio-organic fertilizers stimulate indigenous soil Pseudomonas populations to enhance plant disease suppression. Microbiome 2020, 8, 137. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Li, J.; Xiao, Y.; Gu, Y.; Liu, H.; Liang, Y.; Liu, X.; Hu, J.; Meng, D.; Yin, H. An Integrated Insight into the Relationship between Soil Microbial Community and Tobacco Bacterial Wilt Disease. Front. Microbiol. 2017, 8, 2179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wen, T.; Zhao, M.; Liu, T.; Huang, Q.; Yuan, J.; Shen, Q. High abundance of Ralstonia solanacearum changed tomato rhizosphere microbiome and metabolome. BMC Plant Biol. 2020, 20, 166. [Google Scholar] [CrossRef] [Green Version]
- Bonilla, N.; Gutierrez-Barranquero, J.A.; Vicente, A.D.; Cazorla, F.M. Enhancing Soil quality and plant health through suppressive organic amendments. Diversity 2012, 4, 475–491. [Google Scholar] [CrossRef]
- He, J.; Li, J.; Zheng, Y. Reflections on the relationship between microbial diversity and stability in soil ecosystems. Biodiversity 2013, 21, 412–421. [Google Scholar]
- Garbeva, P.; Veen, J.A.; Elsas, J.D. Microbial diversity in soil: Selection of microbial populations by plant and soil type and implications for disease suppressiveness [Review]. Ann. Rev. Phytopathol. 2004, 42, 243–270. [Google Scholar] [CrossRef]
- Fan, J.; Tan, J.; Wang, R.; Deng, J.; Zhang, J.; Xu, D.; Shi, H.; Xiang, B. Analysis of Soil Physical and Chemical Properties and Bacterial Community Structure for Tobacco R. wilt disease. China Tob. Sci. 2021, 42, 15–21. [Google Scholar]
- Li, J.G.; Ren, G.D.; Jia, Z.J.; Dong, Y.H. Composition and activity of rhizosphere microbial communities associated with healthy and diseased greenhouse tomatoes. Plant Soil 2014, 380, 337–347. [Google Scholar] [CrossRef]
- Xiang, L.G.; Guo, H.; Zhou, H.; Xie, H.L.; Cai, L.T.; Wang, H.C.; Ding, W.; Yu, Z.H. Fungal community structure and diversity of the rhizosphere soil and stems of healthy and diseased tobacco plants infected with bacterial wilt. Plant Prot. 2020, 46, 189–196. [Google Scholar]
- Ren, H.; Xu, Q.; Qi, X.; Yu, Z.; Zheng, X.; Zhang, S.; Wang, Z. Differences in the body and rhizosphere flora between healthy and wilted myrica. Chin. J. Appl. Ecol. 2021, 32, 3107–3118. [Google Scholar]
- Li, F.; Chen, L.; Redmile-Gordon, M.; Zhang, J.B.; Zhang, C.Z.; Ning, Q.; Li, W. Mortierella elongata’s roles in organic agriculture and crop growth promotion in a mineral soil. Land Degrad. Dev. 2018, 29, 1642–1651. [Google Scholar] [CrossRef]
- Xiong, W.; Li, R.; Ren, Y.; Liu, C.; Zhao, Q.Y.; Wu, H.S.; Jousset, A.; Shen, Q.R. Distinct roles for soil fungal and bacterial communities associated with the suppression of vanilla Fusarium wilt disease. Soil Biol. Biochem. 2017, 107, 198–207. [Google Scholar] [CrossRef]
- Muhammad, A.; Islam, W.; Shabbir, A.; Khan, K.A.; Ghramh, H.A.; Huang, Z.; Chen, H.Y.H.; Lu, G.L. Plant defense against fungal pathogens by antagonistic fungi with Trichoderma in focus. Microb. Pathog. 2019, 129, 7–18. [Google Scholar]
- Li, T.; Deng, X.; Li, R.; Liu, H.; Shen, Z.; Li, R.; Shen, Q. Effects of tomato bacterial wilt on soil fungal community diversity. J/OL Biotechnol. Bull. 2022, 1–9. [Google Scholar]
- Yang, X.; Zhang, L.; Huang, X.; Wu, W.; Zhou, X.; Du, L.; Li, H.; Liu, Y. Analysis of microbial community structure differences between soybean and rape rhizosphere based on high-throughput sequencing. J. Appl. Ecol. 2019, 30, 2345–2351. [Google Scholar]
- Padhi, E.M.T.; Maharaj, N.; Lin, S.Y.; Mishchuk, D.O.; Chin, E.; Godfrey, K.; Foster, E.; Polek, M.; Leveau, H.J.; Slupsky, C.M. Metabolome and microbiome signatures in the roots of citrus affected by Huanglongbing. J. Phytopathol. 2019, 109, 2022–2032. [Google Scholar] [CrossRef]
- Xiong, W.; Guo, S.; Jousset, A.; Zhao, Q.Y.; Wu, H.S.; Li, R.; Kowalchuk, G.A.; Shen, Q.R. Bio-fertilizer application induces soil suppressiveness against Fusarium wilt disease by reshaping the soil microbiome. Soil Biol. Biochem. 2017, 114, 238–247. [Google Scholar] [CrossRef]
- Deng, X.; Zhang, N.; Li, Y.; Zhu, C.; Qu, B.; Liu, H.; Li, R.; Bai, Y.; Shen, Q.; Falcao, S.J. Bio-organic soil amendment promotes the suppression of Ralstonia solanacearum by inducing changes in the functionality and composition of rhizosphere bacterial communities. New Phytol. 2022, 235, 1558–1574. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | TN g·kg−1 | TP g·kg−1 | TK g·kg−1 | pH | SOM g·kg−1 | ASi g·kg−1 |
|---|---|---|---|---|---|---|
| PS | - | - | 631.67 ± 0.33 | 12.07 ± 0.02 | 185.86 ± 1.41 | |
| CS | 2.89 ± 0.09 | 1.04 ± 0.03 | 2.94 ± 0.69 | 10.72 ± 0.12 | 108.24 ± 0.85 | 8.57 ± 0.41 |
| BC | 9.33 ± 0.93 | 2.97 ± 0.01 | 17.47 ± 0.12 | 9.96 ± 0.05 | 138.78 ± 5.08 | 0.46 ± 0.01 |
| SCHA | 3.27 ± 0.47 | 0.27 ± 0.01 | 62.80 ± 0.65 | 9.52 ± 0.06 | 97.84 ± 0.43 | 0.58 ± 0.02 |
| BOF | 29.87 ± 3.37 | 14.24 ± 0.03 | 7.57 ± 0.52 | 4.68 ± 0.15 | 380.13 ± 8.39 | 0.20 ± 0.02 |
| Treatment | Incidence (%) | pH | NH4+-N (mg/kg) | NO3−-N (mg/kg) | AP (mg/kg) | AK (mg/kg) | ASi (mg/kg) | ECa (cmol/kg) |
|---|---|---|---|---|---|---|---|---|
| CK | 30.00 ± 5.31a | 4.77 ± 0.11a | 21.00 ± 7.81b | 64.09 ± 23.78b | 400.20 ± 14.50a | 477.36 ± 40.56c | 30.45 ± 3.07b | 3.95 ± 0.26ab |
| PS | 27.50 ± 2.63a | 4.45 ± 0.11abc | 45.10 ± 14.82ab | 110.76 ± 30.30ab | 577.80 ± 20.94a | 656.88 ± 43.98ab | 52.08 ± 10.06b | 3.03 ± 0.43b |
| CS | 36.00 ± 8.45a | 4.59 ± 0.06ab | 33.29 ± 11.88ab | 84.54 ± 34.53ab | 594.98 ± 61.56a | 502.10 ± 40.78c | 187.12 ± 85.11a | 4.04 ± 0.58ab |
| BC | 12.75 ± 1.65b | 4.22 ± 0.13c | 47.50 ± 14.61ab | 93.85 ± 28.54ab | 539.29 ± 64.82a | 538.56 ± 24.88bc | 39.53 ± 11.31b | 2.47 ± 0.32b |
| SCHA | 13.25 ± 4.66b | 4.31 ± 0.12bc | 72.55 ± 12.35a | 180.15 ± 44.36a | 454.41 ± 122.32a | 781.07 ± 74.84a | 32.38 ± 1.92b | 4.81 ± 1.05a |
| BOF | 9.00 ± 0.07b | 4.46 ± 0.12bc | 55.21 ± 18.31ab | 176.78 ± 34.56a | 404.37 ± 50.21a | 503.37 ± 26.68c | 26.97 ± 1.92b | 3.60 ± 0.27ab |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, S.; Bai, Z.; Zhang, Z.; Bi, J.; Wang, E.; Sun, M.; Asante-Badu, B.; Zhang, J.; Njyenawe, M.C.; Song, A.; et al. Organic or Inorganic Amendments Influence Microbial Community in Rhizosphere and Decreases the Incidence of Tomato Bacterial Wilt. Agronomy 2022, 12, 3029. https://doi.org/10.3390/agronomy12123029
Wang S, Bai Z, Zhang Z, Bi J, Wang E, Sun M, Asante-Badu B, Zhang J, Njyenawe MC, Song A, et al. Organic or Inorganic Amendments Influence Microbial Community in Rhizosphere and Decreases the Incidence of Tomato Bacterial Wilt. Agronomy. 2022; 12(12):3029. https://doi.org/10.3390/agronomy12123029
Chicago/Turabian StyleWang, Sai, Zhanbing Bai, Zhuo Zhang, Jingjing Bi, Enzhao Wang, Miaomiao Sun, Bismark Asante-Badu, Jiayin Zhang, Marie Claire Njyenawe, Alin Song, and et al. 2022. "Organic or Inorganic Amendments Influence Microbial Community in Rhizosphere and Decreases the Incidence of Tomato Bacterial Wilt" Agronomy 12, no. 12: 3029. https://doi.org/10.3390/agronomy12123029