


Ammonium Phytotoxicity and Tolerance: An Insight into Ammonium Nutrition to Improve Crop Productivity


1
Institute of Agriculture and Life Science, Gyeongsang National University, Jinju 52828, Republic of Korea
2
Department of Horticulture, Division of Applied Life Science (BK21 Four), Graduate School, Gyeongsang National University, Jinju 52828, Republic of Korea
3
Research Institute of Life Science, Gyeongsang National University, Jinju 52828, Republic of Korea
*
Author to whom correspondence should be addressed.
Agronomy 2023, 13(6), 1487; https://doi.org/10.3390/agronomy13061487
Submission received: 12 April 2023
/
Revised: 25 May 2023
/
Accepted: 25 May 2023
/
Published: 28 May 2023
(This article belongs to the Topic Plants Nutrients)
Abstract
:Ammonium sensitivity is considered a globally stressful condition that affects overall crop productivity. The major toxic symptom associated with ammonium nutrition is growth retardation, which has been associated with a high energy cost for maintaining ion, pH, and hormone homeostasis and, eventually, the NH3/NH4+ level in plant tissues. While certain species/genotypes exhibit extreme sensitivity to ammonium, other species/genotypes prefer ammonium to nitrate as a form of nitrogen. Some of the key tolerance mechanisms used by the plant to deal with NH4+ toxicity include an enhanced activity of an alternative oxidase pathway in mitochondria, greater NH4+ assimilation plus the retention of the minimum level of NH4+ in leaves, and/or poor response to extrinsic acidification or pH drop. Except for toxicity, ammonium can be considered as an energy-efficient nutrition in comparison to nitrate since it is already in a reduced form for use in amino acid metabolism. Through effective manipulation of the NH4+/NO3 − ratio, ammonium nutrition can be used to increase productivity, quality, and resistance to various biotic and abiotic stresses of crops. This review highlights recent advancements in ammonium toxicity and tolerance mechanisms, possible strategies to improve ammonium tolerance, and omics-based understanding of nitrogen use efficiency (NUE) in plants.
1. Introduction
Nitrogen (N) holds a prominent position in the metabolic system of plants, as it is part of amino acids, which are the building blocks of proteins and enzymes. Among all the mineral elements, N is regarded as one of the vital macronutrients for all living tissues in plants, including metabolism, resource distribution, growth, and development [1]. Nitrogen uptake is necessary for plant growth as it is crucial for the production of coenzymes, proteins, and photosynthetic processes. Although dinitrogen gas makes up about 78% of the atmosphere, plants cannot use it precisely. About 65% of the available N for the biosphere is produced via the biological N fixation process, which converts atmospheric N2 to ammonium. Ammonium (NH4+) and nitrate (NO3−) are the two forms of accessible N that plants can take up from the soil and are assimilated into amino acids, either in the roots or shoots, depending on the plant species. In many plant species, the majority of the nitrate taken up by the roots is transferred to the shoot, where it is further assimilated into amino acids [2]. However, in a few species such as peas, amino acid synthesis primarily occurs in the roots, before being transported by the xylem to mature source leaves, where the amino-N is used in a variety of metabolic activities or loaded into the phloem to provide N to developing sinks, such as fruit and seeds [3]. Therefore, the main factors affecting soil nitrogen input for plant nutrition and agricultural yield are the decomposition of organic matter, the use of synthetic fertilizers, and biological nitrogen fixation through nitrogenase enzyme activity. Due to low bioavailability and greater dependence of crop growth on nitrogen, there is a massive global market for N-based fertilizers, such as ammonium (NH4+), nitrate (NO3−), and urea (CH4N2O) [4]. Estimates from the FAO show that 107.7 Mt of elemental N from synthetic fertilizers were consumed globally for agriculture in 2018, with China, India, the United States, the EU28, and Brazil contributing 68% of the overall N use [5]. However, about 50% of nutrient losses from N-fertilizers, significantly contribute to low fertilizer use, which prevents crops from reaching their yield potential, increases the cost of farmers’ investments in agricultural inputs, and worsens food and nutrition insecurity [6]. Moreover, N-fertilizer losses contribute to the eutrophication of surface waters, contamination of subsurface water, and atmospheric greenhouse gas emissions, all of which worsen the impact of agriculture on the environment [7]. Therefore, in order to achieve sustainable agriculture, it is imperative to enhance yield and nitrogen use efficiency (NUE), as well as reduce nitrogen application and losses to the environment.
Although inorganic nitrogen in the form of nitrate is primarily accessible to plants, ammonium can be the dominant N ion in some soils and hydroponic cultures, while amino acids can also be taken up by natural and agricultural crops [8]. The availability of ammonium (NH4+) in soils and subsequently the primary productivity of terrestrial ecosystems is controlled by two important soil microbial processes, known as gross ammonification (GA) and gross nitrification (GN) rates. Numerous factors, such as the pH, chemical makeup of the soil, temperature, the buildup of organic molecules, oxygenation, CO2, and light affect the accumulation of NH4+ in ecosystems. Typically, low pH, low temperature, the buildup of compounds with phenolic bases, and insufficient oxygen supply prevent many nitrifying bacteria, leading to higher rates of net ammonification than net nitrification [9]. Although ammonium toxicity may be pervasive, the point at which symptoms start to occur varies greatly among plant species. Generally, many domesticated plants such as potato, tomato, bean, pea, sugar beet, strawberry, and mustard are sensitive to NH4+, while few crops such as rice, onion, cranberry, and blueberry are highly adapted to NH4+ [9,10]. Toxic quantities of ammonium build up in plant tissues when the net rate of converting ammonium into amino acids and amides is less than the rate of its intake and cellular synthesis, via amino acid catabolism, phenylpropanoid metabolism, nitrate reduction, and photorespiration. Therefore, plants must maintain a delicate balance between NH4+ absorption, production, and consumption in order to prevent toxicity. This review presents a brief overview of ammonium toxicity and its underlying tolerance mechanisms with a special emphasis on how ammonium nutrition can be potentially used to improve plant productivity, quality, and resistance to biotic and abiotic stresses. It also provides an update on recent advancements in nitrogen metabolism supported by omics research and the future perspectives to improve ammonium tolerance.
2. Nitrogen Metabolism and Ammonium Assimilation
N is a nutrient that plants can obtain from a variety of compounds, viz., nitrate, urea, ammonium, and amino acids and use for a variety of metabolic processes, including the synthesis of nucleic acids and proteins, while also as signaling and storage molecules. An essential physiological mechanism for the growth and development of plants is the assimilation of nitrogen into carbon skeletons. Under aerobic conditions, NO3− acts as the main source of N with rapid mobility and storability in vacuoles; however, its reduction to NH4+ is required for the synthesis of many organic compounds and proteins. All inorganic N is first converted to ammonium since it is the only reduced N form that plants can employ to assimilate into N-carrying amino acids [8]. A two-stage process is involved to convert nitrate into NH4+; in the first step, nitrate reductase converts NO3− into nitrite in the cytoplasm, and in the second step, nitrite is turned into NH4+ by nitrite reductase in the plastids [11]. Then, ammonia is assimilated into glutamine (Gln) and glutamate (Glu) through the glutamate synthase/glutamine-2-oxoglutarate amino transferase (GS/GOGAT) cycle [12,13] (Figure 1). The enzyme, glutamine synthetase (GS) has a vital role in the assimilation of NH4+ into glutamine, while it is also a rate-limiting step in the NH4+ assimilation. Additionally, GOGAT and GS have two isoforms and their location has been identified as being tissue specific. For instance, NADH-GOGAT (NADH-dependent glutamate synthase) and GS1 are implicated in nitrogen assimilation in the roots, while Fd-GOGAT (reduced ferredoxin-dependent glutamate synthase) and GS2 are primarily involved in nitrogen assimilation in leaves [13]. Among five GS1 genes in Arabidopsis (GLN1;1–GLN1;5), GLN1;2 is the main isoenzyme responsible for GS1 activity in the roots. The gln1;2 mutant exhibits poor GS activity, excessive ammonium buildup, and decreased plant growth when supplied with ammonia, indicating that GLN1;2 is crucial for both assimilation of ammonia and detoxification in roots [14]. NADH-GOGAT serves as the primary isoform in the roots of Arabidopsis and exogenous ammonium supply greatly induces the expression of this isoform. Under the conditions of ammonium availability, the nadh-gogat mutant exhibits lower glutamate formation and biomass buildup [15]. However, as endogenous NH4+ concentration rises, a different route regulated by the mitochondrial NAD(H)-dependent glutamate dehydrogenase (GDH) helps to lower this internal NH4+ concentration (Figure 1). The GDH enzyme catalyzes the reversible amination of 2-oxoglutarate (2-OG) to form glutamate, using ammonium as a substrate. Therefore, the GDH enzyme is theoretically able to either assimilate or release ammonium. Evidence suggests that under normal conditions, GDH operates in the direction of Glu deamination to form ammonium [16]. However, when ammonium concentration increases above a certain threshold, particularly under stress conditions [17], or when plants are grown under sole NH4+ N source, the GDH activity is greatly induced to lower the internal concentration of NH4+ [18]. The ability of GDH to release N from amino acids results in the creation of a keto-acid, 2-OG, and NH3, which can, then, be independently recycled for use in respiration and the synthesis of amides, respectively. The C-skeletons generated by other metabolic pathways, including respiration and photosynthesis, can also be used to integrate ammonia into the amino acids, glutamine, and glutamate. However, ammonium is thought to be toxic to plants at higher concentrations, as it can result in proton extrusion, which is linked to ammonium uptake, cytosolic pH alterations, and the dissociation of photophosphorylation in plants.
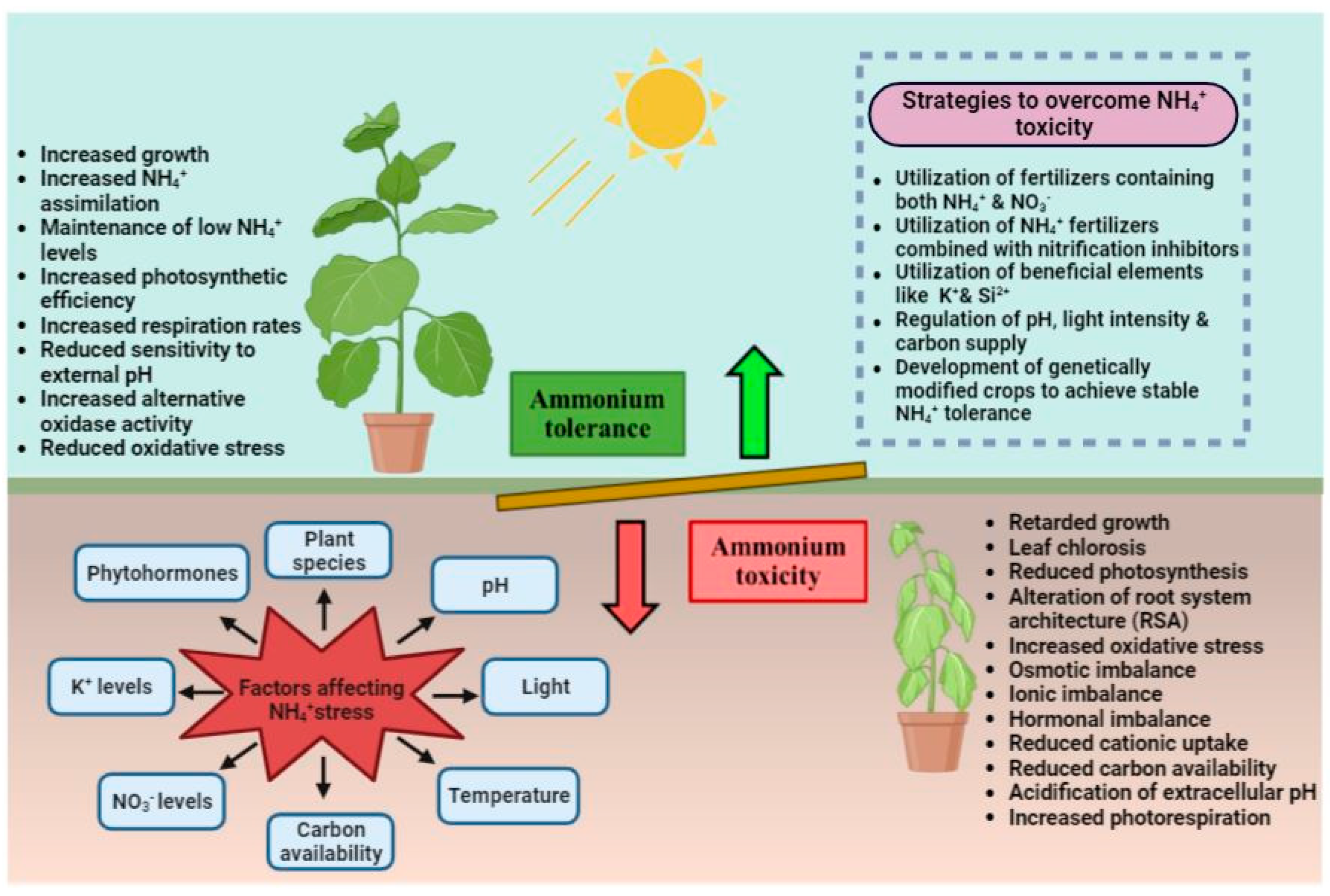
3. Signs of Ammonium-Induced Toxicity
Ammonium toxicity often develops, when a plant is exposed to high levels of ammonium in the environment, or alternatively, when plant cells spontaneously overproduce ammonium due to enhanced proteolytic activity as a consequence of environmental stress. Widely varying documented NH4+ toxicity symptoms typically manifest at external NH4+ values over 0.1 to 0.5 mmol L−1 [20]. The primary phenotypic indicators of ammonium toxicity in plants are restricted root growth and chlorosis of the leaves [10]. Ammonium toxicity modifies the biochemical and physiological makeup of the plant, affecting its intracellular pH, osmotic balance, metabolism of phytohormones and polyamines, and ability to absorb nutrients (Figure 2). Ammonium toxicity can also cause a reduction in the amount of carbon availability, damage to the chloroplast’s ultrastructure, an increase in proton efflux, ineffective transmembrane ammonium cycling, suppression of the enzyme GDP-mannose pyrophosphorylase, as well as oxidative stress [17,19]. Ammonium toxicity is also accompanied by lower levels of chlorophyll a and b and carotenoids, a decline in photosynthetic rates, and a rise in the production of ethylene [19,21]. In cucumber, higher levels of ammonium were found to be toxic, causing toxicity symptoms such as suppression of growth, chlorosis, curling of leaves, and low tissue accumulation of calcium and magnesium, while the addition of lesser amounts of NH4+ to NO3− has drastically improved the growth rate [22]. Increased ammonium levels have negatively impacted the uptake of cations, such as Ca, K, Mg, and Si, in papaya, while also reducing growth [23]. Excess ammonium has resulted in calcium deficit, restricted N uptake, altered N/Ca ratio as well as physiological changes in tomato plants [24]. Recently, the interactive role of ammonium nutrition with iron (Fe) homeostasis has been demonstrated in the model plant Brachypodium distachyon [25]. It was shown that plants that had been fed with ammonium showed heightened susceptibility to iron deficiency, indicating that ammonium nutrition resulted in decreased Fe uptake and root-to-shoot transfer. Thus, in order to better comprehend and enhance ammonium use efficiency and tolerance in plants, future studies are required to explain the mechanisms driving the interaction between Fe and NH4+, particularly regarding the mechanism leading to Fe deficiency under ammonium stress.
Root development is more strongly impacted by excess NH4+ than any other plant organ. Higher NO3− concentrations promote lateral root growth, while higher NH4+ concentrations result in stunted roots with slow growth rates. Overabundance of NH4+ primarily shortens primary roots by preventing cell expansion and division in the meristems. Under these circumstances, GDP-mannose pyrophosphorylase (GMPase)-mediated NH4+ efflux increases in the meristematic zone, leading to a higher NH4+ influx in mature root cells [26]. Investigations in Lotus japonicus, employing split-root systems have revealed that root-specific NH4+-induced reactions are regulated by NH4+ transporters that are more closely related to NH4+ sensing than its assimilation. In both L. japonicus and Arabidopsis, AMT1;3 mediates the short-root phenotype induced by NH4+ nutrition [27,28]. Ammonium suppresses root gravitropism in a similar way as it inhibits primary root growth. This phenomenon is linked to a considerable delay in the asymmetrical auxin distribution in the elongation zone of the root and a significant reduction in the expression of PIN2, the auxin exporter. ARG1 (Altered Response to Gravity 1) acts as an antagonist to PIN2 to re-establish root gravitropism through PIN3-mediated lateral auxin distribution and AUX1-induced shootward auxin reflux at the root apex under ammonium input [29]. The defective root gravitropism is greatly influenced by the antagonistic interaction between PIN2 and ARG1 in maintaining the asymmetric auxin fluxes under ammonium supply. Thus, PIN2 and ARG1 are the key targets of NH4+-induced impairment of root gravitropism.
4. Postulated Theories of Ammonium Toxicity and Tolerance
The ability of a plant to tolerate ammonium largely depends on its capacity to assimilate ammonium. Shoots are typically the most affected area of the plant to NH4+ feeding, after roots. The early signs of NH4+ toxicity arise at the root level with a significant alteration of the root system architecture (RSA), as roots are the prime NH4+sensor [30] (Figure 2). The exogenous NH4+ eventually reaches the leaves once the root system’s storage capacity has been exhausted. The harmful effects of NH4+ on leaves are mainly associated with its impact on photosynthesis and its ability to cause oxidative stress. For instance, a higher concentration of NH4+ (5 mM) has affected photosynthesis by binding NH3 in the oxygen-evolving complex (OEC) of photosystem II in the cyanobacterium, Synechocystis [31]. In A. thaliana, NH4+ nutrition did not alter the photosynthetic efficiency but raised the amounts of mitochondrial reactive oxygen species (ROS) in the leaf [32]. In line with this, few other studies have confirmed that NH4+ nutrition caused enhanced lipid peroxidation and glutathione-ascorbate cycle enzyme activity, or alterations in the redox state, implying higher ROS production [33,34]. The tolerance mechanisms used by plants to deal with NH4+ toxicity have not been well described because the primary causes of NH4+ toxicity have not been determined clearly. According to the classical hypothesis of ammonium toxicity, some of the key traits contributing to NH4+ tolerance in plants include enhanced alternative oxidase pathway activity in mitochondria, greater NH4+ assimilation and retention of low levels of NH4+ in leaves, and/or poor sensitivity to extrinsic pH acidification [35]. Therefore, several factors, including root carbon metabolism and the capacity to maintain high respiration rates, the ability to limit NH4+ accumulation inside tissues, and tolerance to acidification of the root zone affect a plant’s overall ability to withstand high NH4+ concentrations. The recent hypothesis on NH4+ toxicity, proposed by Coskun et al. [19], suggests gaseous NH3 as the predominant N species transported across the cell membrane instead of NH4+, as opposed to the conventional NH4+ futile cycle hypothesis [10] that has made significant advancements in the understanding of NH4 uptake in plants. This result disproves the idea that excessive energy consumption is linked to NH4+ efflux and puts forth that NH4+ accumulation in cells and nutrient imbalance caused by lower cationic concentrations are the primary causes of NH4+ toxicity. The NH4+ is transported into cells through various transporters including (1) the MEP/AMT/Rh protein family that transports NH3 by deprotonating NH4+ with high affinity under fewer NH4+concentrations [36]; (2) the aquaporins (AQP) that aid in the non-electrogenic NH3 flow through the plasma membrane at high NH4+concentrations [37]; (3) the non-specific cation channels (NSCC) and K+ specific channels that transport NH4+ ions in competition with K+, which are similar in size and surface charge density (Figure 1) [38]. One of the main causes of NH4+ toxicity is the impact of NH4+ nutrition on reducing the concentrations of cations (K+, Ca2+, and Mg2+) in plant tissue. The harmful effects of NH4+ nutrition can, however, be reduced by raising K+ concentrations in the root media. K+ can lessen the toxicity of NH4+ by enhancing the incorporation of NH4+ into organic N-compounds via the activation of enzymes such as glutamate dehydrogenase, glutamine synthetase, etc., as well as by limiting the absorption of NH4+ by low-affinity transporters [39]. K+ concentrations in the root medium have the potential to obstruct the transport of NH3 by specifically regulating the activity of AQP and the plant’s water balance [40].
5. Signaling Responses Involved in Ammonium Sensing and Tolerance
Effective nutrient harvesting by plant roots requires a quick and sensitive assessment of nutrient availability in the environment. Nutrient transporters located at the plasma membrane are the key players in sensing nutrients in the external environment. For example, NRT1.1/NPF6.3 in Arabidopsis and Mep2 in yeast have been recognized as the major transporters of nitrate and ammonium, respectively, across the plasma membrane [41]. Numerous factors are engaged in the crosstalk signaling that controls plant growth and development under the conditions of NH4+ sensitivity and tolerance.
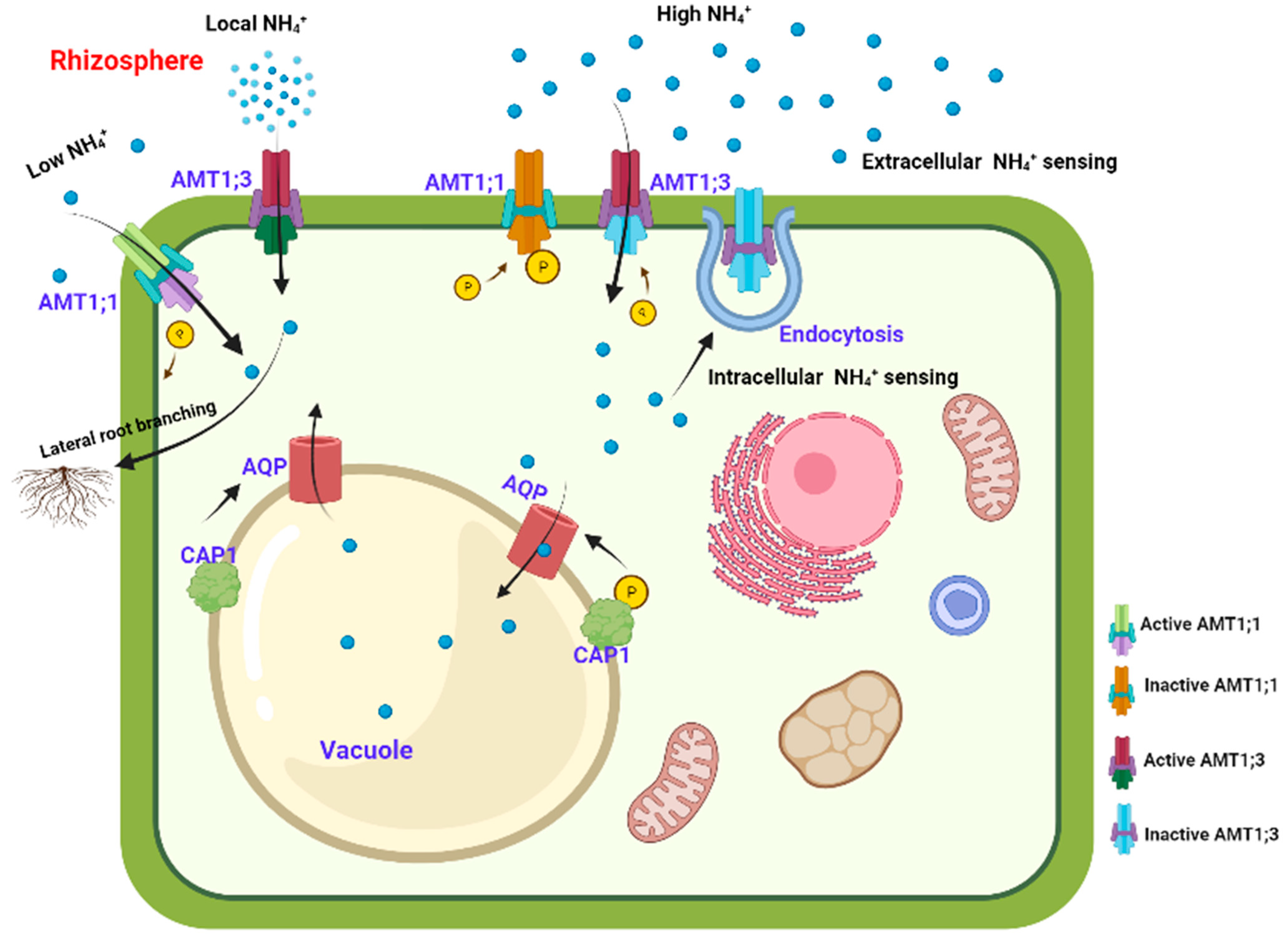
5.1. AMT-Mediated Extracellular Ammonium Sensing Responses
The extracellular sensing and transportation of ammonium are mediated through the phospho-dependent allosteric mechanism of activating the C-terminal domain of AMT1;1 in Arabidopsis [41,42]. The cytosolic C-terminus of each monomer of the trimeric AMT1 complex connects to the pore region of the adjacent subunit to transactivate the ammonium transport. The trans-inactivation of both the individual monomer as well as the entire trimeric complex is facilitated by the phosphorylation of threonine residue in the C-terminus of any monomer that leads to the relieving of contact between the C-terminus and the conducting pore. The critical threonine residue T460 in the C-terminal domain of AMT1;1 is dephosphorylated in conditions of low ammonium availability, whereas conditions of high ammonium supply result in phosphorylation and deactivation of the AMT1;1 trimer. Moreover, it has been demonstrated that glutamine, a product of ammonium assimilation, and methionine sulfoximine (MSX), the glutamine synthetase inhibitor, are unable to induce T460 phosphorylation, suggesting that extracellular ammonium alone can specifically trigger the phospho-dependent inactivation of AMT1;1. Thus, in order to prevent excessive ammonium absorption, it has been postulated that AMT1;1 itself functions as a transceptor, whereby it attracts a kinase from the cytosol following extracellular ammonium binding, and confers either direct or indirect phosphorylation of AMT1;1. Intriguingly, trans-inactivation by C-terminal phosphorylation extends to heterotrimeric AMT1 complexes in addition to homotrimeric ones, as demonstrated for AMT1;1, AMT1;2 [43], and AMT1;3 [44]. This greatly increases the potential for regulating the exchange of signals between AMT isoforms via protein–protein interactions. In furtherance, AMT1;3 plays a crucial role in ammonium-mediated lateral root branching. Therefore, it appears that AMT1;1 and AMT1;3 are significant players in various signaling pathways, and their colocalization and physical interaction enable the coordination of both ammonium sensing and transport in response to the external availability of ammonium in the rhizosphere (Figure 3). Surprisingly as AMT1;3 seems to resist the C-terminal phosphorylation and inactivation caused by ammonium, an alternate feedback mechanism is activated in response to increased ammonium supply [45]. Following ammonium addition, AMT1;3 proteins aggregate and are subsequently removed from the plasma membrane through the endocytic pathway, as demonstrated in the glutamine synthetase mutant gln1;2. Hence this alternative shutoff mechanism helps to prevent ammonium toxicity in root cells.
5.2. CAP1-Mediated Intracellular Ammonium Sensing Responses
In addition to ammonium assimilation, alternative mechanisms are also in operation to provide tolerance to ammonium toxicity. It has been shown in Arabidopsis that CAP1, a receptor-like kinase, mediates the compartmentation of ammonium in vacuoles under conditions of high intracellular ammonium [46] (Figure 3). The tonoplast localized unique regulator of the NH4+ homeostasis signal, CAP1 has been found to regulate the polar development of root hairs by maintaining tip-focused cytoplasmic Ca2+ gradients as well as ROS gradients. A CAP1 knockout mutant (cap1-1), exhibited an increased concentration of cytoplasmic NH4+. In addition, on Murashige and Skoog media, the root hair growth of cap1-1 was suppressed, while NH4+ depletion restored the Ca2+ gradient required for normal growth. The comparatively alkaline cytosolic pH of cap1-1 root hairs and the reduced net NH4+ inflow across the vacuolar membrane suggested that CAP1 mutation promoted NH4+ accumulation in the cytoplasm. Moreover, ectopic expression of CAP1 in the yeast mutant npr1 accomplished the missing function of NPR1, a crucial component of the ammonium-sensing pathway, and restored the phosphorylation as well as the ammonium transport activity of Mep2. This suggests that CAP1 may also function in plants as a regulatory component in intracellular ammonium-sensing events [46]. Autophosphorylation of CAP1 upon exposure to ammonium has been confirmed in Arabidopsis through a phosphoproteomics approach [46,47]. Moreover, the location of CAP1 in the tonoplast suggests that CAP1 may phosphorylate vacuolar ammonium transporters. The aquaporins, TIP2;1 and TIP2;3 that are located in the tonoplast and involved in ammonium compartmentation might be the potential putative targets for CAP1. However, so far only the transcriptional upregulation of the ammonia-conducting aquaporin TIP2;3 has been evidenced in ammonium treated cap1-1 mutants [48]. Thus, the transcriptional regulators responsible for amplifying the ammonium-sensing event and orchestrating the subsequent ammonium responses are still unclear. Hence, future studies that examine the local integration of external and internal ammonium-dependent signals to control AMT-dependent ammonium transport are of utmost importance.
5.3. Nitrate, Auxins, NO, and Polyamines-Mediated Signaling of NH4+ Tolerance
The alleviating role of NO3− in ammonium toxicity has been mediated through the crosstalk signaling among NO3−, NRT1.1/NPF6.3, NO3− transporter, and auxin. Since NO3− induces the expression of the NRT1.1/NPF6.3 transporter gene, plants lacking NO3− in the root medium may experience disruptions in this signaling pathway–for example, short and less branched roots in the case of NH4+ and urea-fed Medicago truncatula, respectively [49]. The auxin-responsive reporter DR5:GUS-reporter has been used in a mutant strain of Arabidopsis to study the function of auxin signaling in the growth of primary roots fed with NH4+. The study showed that the response of the reporter to auxins under NH4+ nutrition was dramatically reduced, whereas exogenous auxin-only partially restored root growth [50]. It was determined that NRT1.1/NPF6.3 had a NO3--independent function since NRT1.1/NPF6.3 defective mutants had a better tolerance for high concentrations of NH4+ and NO3−, which when administered did not increase this tolerance towards NH4+. In NO3−-mediated reduction of ammonium stress, NO3− efflux channels, such as SLAH3, also played a significant role [51]. In the absence of NO3−, SLAH3-defective mutants are hypersensitive to high NH4+ concentrations as well as low pH.
Plants lack the essential enzyme, carbamoyl phosphate synthase (CPS)-type I of animals; nevertheless, they appear to possess a functional CPS-type II that utilizes glutamine as the NH3 source for carbamoyl-P synthesis. As a result, plants are unable to produce urea directly from NH4+, which would lower the internal NH4+ concentration under stress. The plant urea cycle comprises two phases: the biosynthetic phase, which produces arginine, and the catabolic phase, where arginine can be broken down by arginase to produce urea and ornithine or act as a substrate for arginine decarboxylase to make agmatine. Either agmatine or ornithine can further produce putrescine, spermidine, and other polyamines. Arginine has been proposed as an intermediate in NO synthesis and links the urea cycle with NH4+ tolerance. It has been implied that nitric oxide (NO) derived from NO3- through nitrate reductase also plays a profound role in auxin-signaling by acting downstream of auxin. This was confirmed by arginase-negative mutants of Arabidopsis, through which increased NO builds up and effluxes, while promoting the development of lateral and adventitious roots [52]. The root tips, developing lateral roots, and hypocotyls of these mutants consistently displayed enhanced auxin signaling. However, the source of NO in plants fed with NH4+ or urea as the only N nutrition remains elusive. It was also found that the exogenous application of polyamines such as spermidine and spermine has led to the increased synthesis of NO in Arabidopsis [53]. It has been hypothesized that an excess of ammonium resulting either from NH4+ nutrition or from polyamine metabolism may operate as a feedback inhibitor of polyamine deamination, thereby reducing polyamine metabolism. Further, metabolic studies in peas and Arabidopsis have revealed that ammonium supplementation caused arginine accumulation, presumably indicating impaired polyamine production [44,54].
6. Ameliorated Crop Quality and Productivity by Ammonium Nutrition
In agricultural settings, the two most common types of inorganic nitrogen taken by the roots of higher plants are ammonium (NH4+) and nitrate (NO3−). In the rhizosphere environment, the absorption of nitrate nitrogen is associated with the inorganic cation uptake and the release of OH−, leading to an alkaline rhizosphere, while ammonium nitrogen absorption takes place by the uptake of inorganic anion and the release of H+, causing an acidic rhizosphere. An overabundance of free NH4 + in the cytosol is likely the primary source of ammonium toxicity and plants employ a number of rational ways to manage NH4+ levels, including absorption into organic compounds, storage of NH4 in the vacuole, and outflow of NH4 to the apoplast and rhizosphere. Despite ammonium stress affecting almost all plant species, there is considerable intra- and interspecific variation in response to ammonium feeding. While some species/genotypes exhibit severe sensitivity to ammonium when growing, other species/genotypes exhibit ammonium preference. The influence of ammonium and nitrate nutrition varied among selected bedding plants, such as Salvia splendens, marigold, Ageratum houstonianum, and Petunia hybrida [55]. Salvia and marigold were highly sensitive to NH4+, while ageratum, being tolerant to NH4+, was more sensitive to NO3−. The study also revealed that NH4+-fed plants contained more anions, such as more Cl- and fewer cations than NO3−-fed plants. NH4+ nutrition has significantly enhanced the chlorophyll contents of ageratum and petunia, while NO3− nutrition increased the chlorophyll contents of Salvia [56]. Upon investigating the effect of NH4+ and NO3− in the presence and absence of Cl−, the uptake of NO3- was suppressed by NH4+, while the uptake of NH4+ was unaffected by NO3− and the uptake of Cl- was counteracted by both NH4+ and NO3− [57]. In Arabidopsis, ammonium as a predominant N source negatively impacted biomass production [58,59], while in another study, ammonium nutrition was found to be beneficial in the improvement of glucosinolate metabolism [60]. Ammonium as a sole N source has improved the grain quality in wheat by increasing the grain reserve proteins, such as gliadins and glutenins [61]. The effects of N nutrition in the form of NH4+ and NO3- have been extensively studied in many plant species. However, the requirement of N sources varied among species. For example, NO3− nutrition positively regulated the growth of wheat [62], sugar beet [63], tobacco [64], beans [65], and canola [66], whereas NH4+ nutrition was found suitable for the growth of larch and pine [67], tomato [68], and rice [69]. Nevertheless, employing NO3− as the exclusive source of nitrogen causes N leaching and nitrate buildup in plants [70,71]. For the majority of species, a combination of ammonium and nitrate is recommended as an excellent nutrient recipe than using nitrate or ammonium alone. Numerous studies have achieved high productivity and quality in many plant species by combining NH4+ and NO3- in a particular ratio (Table 1). For instance, optimum concentrations of NH4+ and NO3- have greatly influenced the growth and flowering of Petunia hybrida, grown in rockwool and peat-lite media. The highest shoot dry weight was obtained when equal concentrations of NH4+ and NO3− (9 me. L−1 each) were added to the peat-lite media. Flowering was delayed at increased concentrations of both forms of nitrogen; however, increased NO3- highly affected flowering [72]. The japonica rice cultivars exhibited a high ammonium preference along with a partial requirement of nitrate (NH4+:NO3−,75:25) for efficient photosynthetic activity, higher NUE and faster biomass accumulation [73]. In the case of cabbage, an equal ratio of ammonium and nitrate has greatly improved the seedling fresh weight, root length, and contents of phosphate (H2PO4–), and mineral nutrients (K, Ca, and Mg), while also significantly reducing the nitrate content in leaves [74]. Upon investigating the effect of various NH4+:NO3− proportions in strawberries, the ideal NH4+: NO3− ratio for superior fruit development and quality in the semi-hydroponic fertigation system was found to be 29:71 [75]. In Capsicum annuum L, the ratio of ammonium to nitrate at 25:75 drastically improved root growth, and the accumulations of nutrient elements (N, P, and K) accumulation and dry matter [76]. The treatment also improved the fruit quality by raising vitamin C, contents of total phenols, flavonoids, soluble sugar and protein, dry matter, and pungency levels. The ammonium tolerance was further substantiated by greater levels of the enzymes, GS and GOGAT as well as higher relative expression of the genes encoding these enzymes. Similarly, the NH4+:NO3− ratio of 25:75 has been shown to enhance N uptake and assimilation in flowering Chinese cabbage, mostly via controlling the pH of the nutrient solution [77]. It has been well established that NH4+ uptake and assimilation are associated with proton release processes leading to the acidification or pH drop of the rhizosphere. Thus, the interaction in the decrease in pH and K+ uptake in NH4+ tolerance was investigated in hydroponically cultured ageratum, salvia, cabbage, and lettuce plants at three NH4+:NO3− ratios (0:100, 50:50, and 100:0) [78]. The plants, such as ageratum and lettuce, were found to be more tolerant to NH4+ feeding, which exhibited a rapid decrease of solution pH as well as improved K+ influx when compared to NH4+ sensitive, salvia, and cabbage. In another study, the effect of NH4+ and NO3− ratios (0:100, 50:50, and 100:0) on growth, photosynthesis, nitrogen, and carbohydrate concentrations alongside nitrogen assimilation enzymes were evaluated among NH4+ sensitive (cabbage) and tolerant (lettuce) species. The study reinforced that both plants benefitted from equal ratios of NH4+ and NO3− (50:50) with enhanced growth and quality. Moreover, in contrast to cabbage, the GS and GDH activities in lettuce were boosted in response to an increase in the level of external NH4+ leading to reduced NH4+ accumulation [79]. Thus, the co-provision of NH4+ with NO3- triggers a synergistic growth response that can outpace the maximum growth rates on either nitrogen source alone. For all of the above studies indicated, it is significant to note that the optimal ratio of NH4+:NO3- is also influenced by the total N concentration and the source of NH4+-N. The requirement of total N concentration varied among plant species, ranging between 5.0 and 15 mM L-1, and in all the trials, the total N content was maintained constant, while the ratios of NH4+ to NO3- were altered (Table 1). Additionally, the source of NH4+-N plays a vital role in determining the plant responses to nutrition, osmotic balance, stress, and overall plant development. Ammonium sulfate (AS) is one of the most widely used N fertilizers, which comprises 21% N and 60% sulfur (S). It is a great source of S for crops that demand huge supplies and offers a balanced supply of both N and S. In terms of nutrition, N and S play very close relationships with one another, particularly because both nutrients are necessary for the synthesis of chlorophyll and are components of amino acids. Sulfur is a structural element found in amino acids, protein disulfide bonds, cofactors, and vitamins. S deficiency impacts the biomass, general morphology, yield, and nutritional value of the plants [80]. Hence, the addition of a N source in the form of AS, also meets the S requirements of plants, promoting overall growth.
AS may have certain potential agronomic and environmental advantages over other N fertilizers, such as urea and ammonium nitrate (AN), including (1) minimal to no N loss via NH3 volatilization upon surface application to acid or neutral soils, (2) a better source of N for saline soils as it lessens the detrimental effects of NaCl on plant growth and for saline-sodic calcareous soils, as it enhances soil structure, (3) minimal NO3-N leaching from AS than AN, (4) improved N efficiency due to less denitrification than AN and reduced emission of greenhouse gases (NO and N2O), and (5) the formation of more acidic root rhizosphere by preferred absorption of NH4-N of AS over NO3-N of AN, which may boost soil phosphorus (P) and micronutrient availability [81]. Although AN has the highest proportion of N (33–34%), the use of AN is not recommended for flooded rice soils that are either rainfed or irrigated mainly due to the significant N losses from the denitrification of nitrate. Therefore, AS becomes the preferred choice of fertilizer for rice growers [81]. On the other hand, other N fertilizers, such as ammonium chloride, improved the yield of ratoon in sugarcane during the first cycle but decreased the yield in subsequent cycles. As sugarcane is moderately sensitive to soil salinity, the decrease in yield could be attributed to the saline effect caused by the residual Cl- from NH4Cl- [82]. Similarly, the application of NH4Cl- has provoked the accumulation of cadmium in wheat due to the increased Cl- concentration in the soil [83]. Hence, rationale utilization of the optimum concentration of NH4Cl-, especially in saline-sensitive crops is highly crucial, as Cl- appears to be the preferable counter ion to NH4+, the increased Cl- ions exacerbate the symptoms of salinity stress, in addition to ammonium toxicity.
Ammonium fertilizers are ineffective in cold climates and irrigated systems due to the significantly decreased activity of nitrifying microorganisms. Since NH4+ is a cation, it greatly adheres to most soils, especially to clays and organic matter; thus, there is minimal leaching off with water. Hence, this causes more ammonium toxicity in irrigated systems where nitrification is greatly inhibited. Therefore, management of appropriate NH4+-N source and concentration in irrigated fields is of prime requirement. Recently, several studies have examined the interaction effect between the NH4+-N forms and irrigation systems in cultivated fields. For example, AS application has drastically improved the lint yield in a drip-irrigated, moderately saline cotton field by reducing rhizosphere soil pH and salinity as well as improving P nutrition [84]. In another study, AN fertilization at 120% crop evapotranspiration (ETc), and drip irrigation conditions significantly improved the water use efficiency, chlorophyll, tuber quality, and yield of potatoes compared to AS and urea fertilization [85]. Eggplant is sensitive to NH4+ nutrition in solution culture, while it has responded positively to an appropriate ratio (50:50) of AN fertilization with 75% irrigation level through improved vegetative growth, water use efficiency, and yield compared to AS fertilization [86].
Although nitrate fertilizers are less likely to be hazardous to plants than ammonium fertilizers, they contribute to significant N loss through NO3- leaching and higher NO emissions. Inhibitors of soil nitrification, including dicyandiamide (DCD), have been suggested as an efficient way to lessen soil N loss and N2O emissions. It has been extensively demonstrated that using nitrification inhibitors in conjunction with ammonium fertilizers in agricultural fields can reduce the environmental impact of nitrogen fertilizers [87]. However, the maintenance of N in the NH4+ form in soil by such compounds also raises concerns for crops that are sensitive to ammonium [88]. Hence, future research is needed to determine how nitrification inhibitors and the increased NH4+-N concentrations in soil affect crop growth and yields.

{kind=link}
{kind=link}
{kind=link}
Table 1.
Influence of ammonium to nitrate ratio on growth, physiology, and nitrogen metabolism in plants.
Table 1.
Influence of ammonium to nitrate ratio on growth, physiology, and nitrogen metabolism in plants.
| Plant Species | NH4+: NO3- Ratios Investigated | Total N Concentration (mM L−1) | Source of the NH4+ ions | Optimum NH4+: NO3− Ratio | Effect on Growth, Photosynthesis, and Nitrogen Metabolism | Reference |
|---|---|---|---|---|---|---|
| Chrysanthemum morifolium var. circus | 1:100, 40:60, and 80:20 | 5.0 | (NH4)2SO4 | 80:20 (at pH 7) | Highly vegetative, reproductive, and increased vesicular-arbuscular mycorrhizal (VAM) fungi growth and nutrient content (N, P, K) at pH 7 | [89] |
| Triticum durum ‘Barkay’ | 0:100, 25:75, and 50:50 | 1.4, 2.8, and 5.5, respectively | (NH4)2SO4 and NH4NO3 | 25:75 | Increased grain yield and total yield | [90] |
| Fragaria × ananassa Duch) cvs. ‘Camarosa’ and ‘Selva’ | 0:100. 25:75, 50:50, and 75:25 | 3.26 | NH4H2PO4 and (NH4)2SO4 | 25:75 | Greater vegetative growth (number of leaves, leaf fresh and weights, chlorophyll index, leaf area, and net photosynthetic and transpiration rates), reproductive growth (number of flowers and fruits, percentage of fruit set, fruit length, fresh weight and dry weights of fruit, and percent total soluble sugars and Ca content of fruit), and overall yield | [91] |
| Persea Americana Mill. | 0:4, 1:3, 2:2, 3:1, and 4:0 | 3.0 | (NH4)2SO4 | 1:3 | Increased dry matter accumulation and development of secondary shoots | [92] |
| Lycopersicon esculentum Mill. ‘Xinpinbaoguan’ | 0:100, 25:75, and 50:50 | 5.0 | (NH4)2SO4 | 25:75 (both homogenous and heterogenous supply) | Higher shoot and root biomass and total N uptake | [93] |
| Lycopersicon esculentum Mill. ‘Xi Nong 2011’ | 0:100, 25:75, 50:50, 75:25, and 100:0 | 8.0 | NH4Cl | 25:75 (unstressed conditions) 50:50 (chilling stress conditions) | Increased biomass, relative growth rate (RGR), net photosynthetic rate, chlorophyll content, contents of soluble protein and free amino acids and higher activities of GS and NADH-GOGAT (under unstressed conditions) | [94] |
| Gerbera jamesonii L. | 0:100, 20:80, 40:60, and 60:40 | 11.2 | (NH4)2SO4 | 20:80 | Greater biomass, number of flowers, diameters of flower stalk and flower disk, vase life, fresh and dry weights of inflorescences, and relative water content of petals | [95] |
| Phalaenopsis ‘Golden Peoker’ and Dendrobium ‘Valentine’ | 100:0, 75:25, 50:50, 25:75, and 0:100 | 3.0 | (NH4)2SO4 | 60:40 (Phalaenopsis) 50:50 (Dendrobium) | Bigger plants, greater dry matter, diameters of stem and pseudobulb, average number of pseudo bulbs and mineral nutrients | [96] |
| Brassica chinensis L. | 0:15, 5:10, 7.5:7.5, and 10:5 | 15.0 | (NH4)2SO4 | 5:10 | Greater biomass, photosynthetic activity, and low nitrate accumulation under water deficit and low light intensity conditions | [97] |
| Saccharum officinarum L. | 0:100, 25:75, 50:50, 75:25, and 100:0 | 15.0 | NH4Cl | 0:100 and 25:75 | Greater biomass, leaf surface area, root growth, leaf gas exchange, and nutrient content | [98] |
| Brassica oleraceae L. var. acephala | 0:100, 25:75, 50:50, and 75:25 | 10.0 | (NH4)2SO4, NH4NO3, and (NH4)3PO4 | 75:25 | Higher biomass, leaf water content, nutrient content (N, P, K, Fe, Mn, Zn, and Cu), and lower nitrate content | [99] |
| Brassica oleracea var. alboglabraCoffea arabica L. | 1:4, 1:8, and 1:12 0:100, 12.5:87.5, 50:50, and 100:0 | 8.0 12.0 | (NH4)2SO4 NH4NO3 | 1:12 50:50 | Increased leaf number, leaf area, and shoot and total fresh weights Highest dry mass yield, greater nutrient absorption, and photosynthetic ability | [100,101] |
| Brassica napus L. N-efficient genotype (D4-15) and N-inefficient genotype (D2-1) | 0:100, 25:75, 50:50, 75:25, and 100:0 | 6.0 | NH4NO3 | 0:100 (N-efficient genotype) 75:25 (N-inefficient genotype) | Increased shoot and root biomass, well-developed root system, net photosynthetic rate, transpiration rate, stomatal conductance, nutrient content, better ammonium uptake, upregulation of ammonium transporter genes and greater tolerance to oxidative stress caused by NH4+ toxicity in N-inefficient genotype | [102] |
| Vaccinium spp. ‘Emerald’ | 1:1, 2:1, and 4:1 | 7.5 | (NH4)2SO4 | 2:1 | Greater plant height, crown width, chlorophyll content, root activity, and upregulation of nitrate transporter genes (NRT1.5/NPF7.3, NRT2) | [103] |
| Lactuca sativa var. longifolia | 100:0, 20:80, 40:60, and 60:40 | 5.3 | (NH4)2SO4 | 20:80 | Increased growth, head weight, and reduced nitrate accumulation in leaves | [104] |
| Brassica Pekinensis ‘Jinwa no. 2’ | 0:100, 10:90, 15:85, and 25:75 | 5.0 | (NH4)2SO4 | 15:85 (normal light intensity) 10:90 (low light intensity) | Increased activities of nitrate reductase (NR) and glutamine synthetase (GS), high nitric oxide (NO) levels in leaves, upregulation of NR and GS1 genes, and upregulation of proteins related to photosynthesis, carbon and energy metabolism, stress and defense, and protein folding and degradation responses | [105] |
| Brassica alboglabra L. H. Bailey | 0:100, 10:90, 25:75, and 50:50 | 5.0 | NH4Cl | 25:75 | Increased plant height, stem diameter, fresh and dry weights, length, surface area, volume and diameter of root, total N content, and higher NUE | [106] |
| Lycopersicon esculentum Mill. | 0:100, 8:92, 15:85, and 20:80 | 12.0 | (NH4)2SO4 | 15:85 (grafted tomato) 8:92 (non-grafted tomato) | Increased biomass production, number of trichomes, epidermal cell density, stomatal density, fruits per plant, fruit size, average fruit weight, and total soluble solids | [107] |
| Brassica lee ssp. namai Ssamchoo ‘Chunssamhwang 51’ | 0:100, 16.7:83.3, 33.3:66.7, and 50:50 NH4+:NO3-:Urea 17:83:0, 50:50:0; 25:50:25, and 0:50:50 | 13.0 | (NH4)2SO4 | 16.7:83.3 (NH4+: NO3-) and 25:50:25 (NH4+:NO3-:Urea) (combined with 10.7 mmol L−1 Si) | Increased germination rate, germination energy, mean germination rate, specific leaf weight, fresh weight, root activity, and chlorophyll and vitamin C contents | [108] |
| Dracocephalum moldavica L. | 0:100, 25:75, 50:50, 75:25, and 100:0 | 7.7 | (NH4)2SO4 and NH4NO3 | 0:100, 25:75, and 75:25 | 0:100—Highest growth parameters, biomass, and contents of total phenolics, total flavonoids, and carotenoids. 25:75—Highest nutrient (N, Ca, and Mg) and geraniol accumulation 75:25—Highest accumulation of phenolic compounds, viz., p-coumaric acid, rosmarinic acid, caffeic acid, quercetin, gallic acid and rutin and medicinal compounds, viz., geranial and geranyl acetate | [109] |
7. Ameliorated Crop Resilience to Biotic and Abiotic Stresses by Ammonium Nutrition
Ammonium stress can have beneficial effects on crop resilience, to biotic and abiotic stresses, and generate high-quality products, although it may have a modest detrimental influence on productivity in agricultural contexts. The synthesis of most defense-related secondary metabolites depends on nitrogen metabolism, which also serves as a key step in the production of nitric oxide (NO), whose function in plant-pathogen interactions has been extensively investigated [110]. Hence, nitrogen metabolism closely relates to plant immunity. Although plants that have been fed nitrate have generally shown enhanced resistance to pathogenic attacks [111,112], few species, particularly tomatoes, have shown notable resistance to pathogens when fed with ammonium. Ammonium-mediated repression of the virulence of Fusarium oxysporum has been demonstrated in tomatoes, mainly by inhibiting the penetration of virulence components through cellophane [113]. Another study demonstrated that the mild toxicity generated by ammonium stress has led to the buildup of H2O2 in tomato plants, which served as a signal to activate systemic acquired acclimation (SAA), and thereby provided resistance to the infections caused by Pseudomonas syringae [114]. Similarly, ammonium-mediated alterations in carbon and nitrogen metabolisms have been reported to increase the resistance against P. syringae in tomatoes [115]. In rice, the overexpression of the ammonium transporter AMT1;2 improved NUE and resistance to sheath blight disease caused by Rhizoctonia solani, further indicating that ammonium uptake and assimilation are necessary for this defense [116].
In furtherance, it has been recognized that an optimum nitrate-to-ammonium ratio can enhance protein synthesis, decrease the lipid peroxidation rate, and boost the plant’s ability to withstand various abiotic stresses. Ammonium feeding may cause a multitude of reactions, many of which are defense responses that are typical of various abiotic stress conditions. The significance of ammonium in alleviating heavy metal toxicity has been proven through some of the studies. The positive effect of NH4NO3 in improving manganese tolerance has been reported in Trifolium subterraneum L. genotypes [117]. Ammonium nutrition has been found to increase the aluminum (Al) tolerance in soybean [118], rice [119], and Lespedeza bicolor [120]. The primary symptom of Al toxicity is the inhibition of root elongation. Ammonium nutrition, however, dramatically increased root length and reduced Al buildup in roots under Al toxicity. The major mechanism of Al tolerance in the above-mentioned species was the restricted Al absorption, as a result of competition between positively charged Al+ and NH4+ for binding sites in the apoplast of root cell walls. Ammonium has been shown to have a strong mitigating effect on cadmium (Cd) toxicity by increasing antioxidant activity and glutathione-ascorbate cycle efficiency in rice [121]. The higher Cd detoxification effect of NH4+ compared to NO3- has been demonstrated in the hyperaccumulator, Solanum nigrum L [122]. Ammonium-mediated Cd toxicity in S. nigrum has been affected through decreased absorption and accumulation of Cd via downregulation of Cd transport-related genes, increased immobilization of Cd in the cell wall of roots by binding to pectin and cell wall components, and through the upregulation of expansin, (SnExp), a gene responsible for metal tolerance. In contrast to the report on the impact of NH4+ in worsening iron deficiency symptoms of the model plant, B. distachyon [25], the alleviating effects of NH4NO3 and NO in iron deficiency has been documented in pears [123]. The study showed that NH4NO3 significantly reduced the chlorosis of leaves caused by iron deficiency, mostly through promoting the activity of ferric chelate reductase and nitrogen assimilation enzymes. Ammonium has been found to improve the salinity tolerance of plants such as citrus [124], Sorghum bicolor [125], maize, [126], and tomato [127]. Ammonium has ameliorated the polyethylene glycol (PEG)-induced water stress tolerance in rice through improved regulation of NH4+ uptake, assimilation, and aquaporin activity [128,129]. An appropriate ratio of NH4+: NO3- (10:90) has effectively mitigated the low light intensity stress in Brassica pekinensis by modulating the expression of ammonium assimilatory enzymes, such as GS and NR [105]. In cucumber, the greater flux rate of NH4+ into vascular bundles of the lateral vein, midrib, and shoot tip, due to the increased conversion of NO3- to NH4+, has substantially aided in overcoming low-temperature stress, as an energy conservatory mechanism [130]. An equal ratio of ammonium and nitrate (50:50) was found to improve the biomass, photosynthetic efficiency, protein content, and decrease lipid peroxidation in baby lettuce under heat stress conditions [131].
8. Omics Investigations in Comprehending NUE
Numerous key regulatory genes, such as transporters, reductase, synthase, glutamate dehydrogenase, and aminotransferase are involved in the various processes of nitrogen metabolism, including the uptake, translocation, assimilation, and remobilization. Until recently several “omics” studies have focused primarily on rapidly altering gene expression and metabolites in plants under various sources of nitrogen. For instance, a comparative transcriptomic analysis between the cultivated and wild lettuce revealed the tolerance mechanisms exhibited by wild under nitrogen starvation conditions [132]. The study evidenced an enhanced expression of nitrate transporter genes (NRT2.1, NRT2.4, and NRT2.5) and glutamate synthase genes (GLN1 and GLN2) in wild lettuce compared to the cultivated one, suggesting high-efficiency N uptake under low nitrogen conditions. Hence, the study gave insights into improving the NUE of cultivated lettuce by the introgression of novel alleles from the wild species in the future. In perennial ryegrass, transcriptome profiling under various nitrogen levels has identified four crucially enriched pathways, such as the photosynthesis-antenna protein, carotenoid biosynthesis, steroid biosynthesis, and C5-branched dibasic acid metabolism, particularly at excessive nitrogen stress conditions [133]. Transcriptomic analysis was used to examine the processes of ammonium toxicity and tolerance in an aquatic macrophyte Myriophyllum aquaticum under situations of high NH4+ stress [134]. The study evidenced the transcriptional upregulation of chlorophyll a/b binding proteins genes contributing to high NH4+ resistance in the leaves. Moreover, the upregulation of nitrogen metabolism genes, along with the upregulation of catalase genes aided in combating NH4+ stress, by means of counteracting the reactive oxygen species.
A comparative transcriptome analysis between high NUE and low NUE cultivars of Brassica juncea showed that under low nitrate treatments, the high NUE cultivar had an upregulation of genes involved in N absorption, remobilization, and assimilation [135]. Furthermore, the co-expression network analysis has identified a number of nitrate regulatory modules that may be involved in the control of nitrate reductase activity, phenylpropanoid biosynthesis, and carbon:nitrogen interaction in B. juncea. The activation of phenylpropanoid and ROS scavenging pathways to counteract NH4+ toxicity has also been confirmed by the transcriptional responses of the common duckweed Lemna minor under ammonium toxicity [136]. In Camelia sinensis, NRT, AMT, AQP, GS, and GOGAT have been identified as key differentially expressed genes (DEGs) regulating ammonium uptake and assimilation [137]. Analysis of the transcriptome of Salicornia neei, a halophyte grown in saline water containing 3 mM ammonium has led to the precise identification of the ammonium detoxification mechanism, mainly through the upregulation of glutamine synthetase and glutamate synthase genes involved in ammonium homeostasis [138].
As N is necessary for the synthesis of a wide range of metabolites, such as amino acids, proteins, lipids, nucleic acids, and chlorophyll, then, measuring metabolite levels using metabolomic techniques can reveal fundamental details about how biological processes react to physiological or environmental changes brought on by N status [139]. A clear discrimination of metabolic profiles of tea leaves and roots in response to N deficiency was observed by untargeted metabolomics using a GC-TOF/MS method [140]. Surprisingly, N deprivation elevated the concentrations of major phenylpropanoids, organic acids, and amino acids in roots, while in leaves there was a significant downregulation of amino acids. Thus, the differential metabolite accumulation in tea leaves and roots corresponds to their quality and stress response, respectively, under N deficiency conditions. An integrative metabolomic response of multiple species in response to ammonium nutrition showed that the accumulation of the essential amino acids varied depending on the species; however, the synthesis of Gln and/or asparagine (Asn) served as a more precise biomarker of ammonium nutrition [141]. The high NUE in maize fed with a mixture of urea and ammonium has been demonstrated with an integrated transcriptomics and metabolomics study [142]. In addition to a rapid elevation of the transcripts of nitrogen assimilation genes, including ZmGLN1;2, ZmGLN1;5, ZmAMT1.1a, ZmGOT1, and ZmGOT3, the combination of urea and ammonium also caused a significant buildup of amino acids, including tyrosine (Tyr), arginine (Arg), phenylalanine (Phe), and methionine (Met), in shoots. Quantitative proteomics analysis was carried out to investigate the ammonium-mediated growth inhibition of tomato roots. Even though the proteins (K4BPV5 and K4D9J3) corresponding to ammonium assimilation were upregulated, there was limited assimilation of ammonium leading to root growth inhibition. The reason for the higher accumulation of ammonium leading to toxicity could be explained by the downregulation of a protein, Q5NE21 corresponding to carbonic anhydrase, a key player in C metabolism [143].
9. Strategies to Overcome Ammonium Toxicity
The vast understanding of ammonium toxicity through extensive studies has led to the development of several strategies for overcoming ammonium toxicity, both at the laboratory and agricultural field levels (Figure 2). In laboratory conditions, ammonium toxicity can be reduced by increasing CO2 concentrations, light intensities, and exogenous organic or inorganic carbon supplies [22,144,145]. The regulation of the pH of the nutrient solution has also been suggested as an efficient way to manage the symptoms of ammonium toxicity [146]. The beneficial role of silicon (Si) in mitigating ammonium toxicity and other abiotic stresses has been extensively studied [147,148]. Silicon application has effectively reversed the adverse effects of ammonium toxicity and enhanced the growth parameters, photosynthesis, and NUE of many plant species (Table 2). The ability of potassium and nitrate to counteract the harmful effects of ammonium on plants suggests that the strategic use of mineral fertilizers, including K+ and NO3-, in the field could maximize nitrogen utilization and increase the safety of agricultural plants. It is a recommended practice to fertilize soils with ammonium and nitrate, in the form of NH4NO3 to balance the effects of NH4+ as NO3- absorption helps in the alkalinization of the rhizosphere. However, the development of genetically modified (GM) plants is the potential strategy for achieving stable ammonium tolerance. In such cases, it may be beneficial to overexpress the transporters or enzymes involved in the assimilation, detoxification, and compartmentation of ammonia, whereas silencing the genes responsible for ammonia uptake and translocation may result in nitrogen deficiency or other detrimental effects as it can block crucial metabolic pathways [149]. The role of hormones, such as ABA that are involved in AMOS1/EGY1-dependent retrograde signaling of ammonium stress and auxins at elevated concentrations have been reported to alleviate ammonium toxicity [26,50]. Table 2: alleviation of ammonium toxicity using silicon supplementation in various plant species.
10. Conclusions and Future Perspectives
Ammonium assimilation into plant metabolites requires limited energy than nitrate assimilation, as it is already in reduced form. Thus, ammonium nutrition can be considered an energy-saving option that can be readily utilized by plants. However, the toxic effect of ammonium demands the optimization of its use to minimize its detrimental effects. Utilizing high-efficiency N fertilizers that contain both NH4+ and NO3- can significantly aid in managing ammonium toxicity issues and increase plant NUE in agricultural fields. To achieve high plant productivity at the laboratory level, different concentrations, and combinations of NH4+ and NO3- must be investigated, because the ideal ratio of NH4+ to NO3- probably varies for different species. Future research is highly needed to confirm the positive effects of ammonium in increasing plant quality, productivity, and resilience to biotic and abiotic stressors and their mechanisms behind. Moreover, the combined use of physiological and multi-omics studies will allow the exploration of the molecular intricacies of ammonium toxicity and tolerance in the future.
Author Contributions
Conceptualization, J.S. (Jayabalan Shilpha) and B.R.J., writing—original draft preparation, J.S. (Jayabalan Shilpha) and J.S. (Jinnan Song), validation, J.S. (Jayabalan Shilpha) and J.S. (Jinnan Song), writing—review and editing, J.S. (Jayabalan Shilpha) and B.R.J. supervision, B.R.J. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
Not applicable.
Acknowledgments
Jayabalan Shilpha thankfully acknowledges the National Research Foundation of Korea (NRF) for the financial support through the Brain Pool Program, funded by the Ministry of Science and ICT, Republic of Korea.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Crawford, N.M. Nitrate: Nutrient and Signal for Plant Growth. Plant Cell 1995, 7, 859. [Google Scholar] [PubMed]
- Xu, G.; Fan, X.; Miller, A.J. Plant Nitrogen Assimilation and Use Efficiency. Annu. Rev. Plant Biol. 2012, 63, 153–182. [Google Scholar] [CrossRef] [PubMed]
- Atkins, C.A.; Pate, J.S.; Peoples, M.B.; Joy, K.W. Amino Acid Transport and Metabolism in Relation to the Nitrogen Economy of a Legume Leaf. Plant Physiol. 1983, 71, 841–848. [Google Scholar] [CrossRef]
- Santi, C.; Bogusz, D.; Franche, C. Biological Nitrogen Fixation in Non-Legume Plants. Ann. Bot. 2013, 111, 743–767. [Google Scholar] [CrossRef] [PubMed]
- Menegat, S.; Ledo, A.; Tirado, R. Greenhouse Gas Emissions from Global Production and Use of Nitrogen Synthetic Fertilisers in Agriculture. Sci. Rep. 2022, 12, 14490. [Google Scholar] [CrossRef]
- Bindraban, P.S.; Dimkpa, C.O.; White, J.C.; Franklin, F.A.; Melse-Boonstra, A.; Koele, N.; Pandey, R.; Rodenburg, J.; Senthilkumar, K.; Demokritou, P. Safeguarding Human and Planetary Health Demands a Fertilizer Sector Transformation. Plants People Planet 2020, 2, 302–309. [Google Scholar] [CrossRef]
- Coskun, D.; Britto, D.T.; Shi, W.; Kronzucker, H.J. Nitrogen Transformations in Modern Agriculture and the Role of Biological Nitrification Inhibition. Nat. Plants 2017, 3, 17074. [Google Scholar] [CrossRef]
- Godlewski, M.; Adamczyk, B. The Ability of Plants to Secrete Proteases by Roots. Plant Physiol. Biochem. 2007, 45, 657–664. [Google Scholar] [CrossRef]
- Britto, D.T.; Kronzucker, H.J. Ecological Significance and Complexity of N-Source Preference in Plants. Ann. Bot. 2013, 112, 957–963. [Google Scholar] [CrossRef]
- Britto, D.T.; Kronzucker, H.J. NH4+ Toxicity in Higher Plants: A Critical Review. J. Plant Physiol. 2002, 159, 567–584. [Google Scholar] [CrossRef]
- Rosales, E.P.; Iannone, M.F.; Groppa, M.D.; Benavides, M.P. Nitric Oxide Inhibits Nitrate Reductase Activity in Wheat Leaves. Plant Physiol. Biochem. 2011, 49, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Lea, P.J.; Miflin, B.J. Glutamate Synthase and the Synthesis of Glutamate in Plants. Plant Physiol. Biochem. 2003, 41, 555–564. [Google Scholar] [CrossRef]
- Esposito, S.; Guerriero, G.; Vona, V.; Di Martino Rigano, V.; Carfagna, S.; Rigano, C. Glutamate Synthase Activities and Protein Changes in Relation to Nitrogen Nutrition in Barley: The Dependence on Different Plastidic Glucose-6P Dehydrogenase Isoforms. J. Exp. Bot. 2005, 56, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Guan, M.; de Bang, T.C.; Pedersen, C.; Schjoerring, J.K. Cytosolic Glutamine Synthetase Gln1; 2 Is the Main Isozyme Contributing to GS1 Activity and Can Be Up-Regulated to Relieve Ammonium Toxicity. Plant Physiol. 2016, 171, 1921–1933. [Google Scholar] [CrossRef]
- Konishi, N.; Ishiyama, K.; Matsuoka, K.; Maru, I.; Hayakawa, T.; Yamaya, T.; Kojima, S. NADH-dependent Glutamate Synthase Plays a Crucial Role in Assimilating Ammonium in the Arabidopsis Root. Physiol. Plant. 2014, 152, 138–151. [Google Scholar] [CrossRef]
- Labboun, S.; Tercé-Laforgue, T.; Roscher, A.; Bedu, M.; Restivo, F.M.; Velanis, C.N.; Skopelitis, D.S.; Moshou, P.N.; Roubelakis-Angelakis, K.A.; Suzuki, A. Resolving the Role of Plant Glutamate Dehydrogenase. I. in Vivo Real Time Nuclear Magnetic Resonance Spectroscopy Experiments. Plant Cell Physiol. 2009, 50, 1761–1773. [Google Scholar] [CrossRef]
- Skopelitis, D.S.; Paranychianakis, N.V.; Paschalidis, K.A.; Pliakonis, E.D.; Delis, I.D.; Yakoumakis, D.I.; Kouvarakis, A.; Papadakis, A.K.; Stephanou, E.G.; Roubelakis-Angelakis, K.A. Abiotic Stress Generates ROS That Signal Expression of Anionic Glutamate Dehydrogenases to Form Glutamate for Proline Synthesis in Tobacco and Grapevine. Plant Cell 2006, 18, 2767–2781. [Google Scholar] [CrossRef]
- Tercé-Laforgue, T.; Dubois, F.; Ferrario-Méry, S.; Pou de Crecenzo, M.-A.; Sangwan, R.; Hirel, B. Glutamate Dehydrogenase of Tobacco Is Mainly Induced in the Cytosol of Phloem Companion Cells When Ammonia Is Provided Either Externally or Released during Photorespiration. Plant Physiol. 2004, 136, 4308–4317. [Google Scholar] [CrossRef]
- Coskun, D.; Britto, D.T.; Li, M.; Becker, A.; Kronzucker, H.J. Rapid Ammonia Gas Transport Accounts for Futile Transmembrane Cycling under NH3/NH4+ Toxicity in Plant Roots. Plant Physiol. 2013, 163, 1859–1867. [Google Scholar] [CrossRef]
- Van Katwijk, M.M.; Vergeer, L.H.T.; Schmitz, G.H.W.; Roelofs, J.G.M. Ammonium Toxicity in Eelgrass Zostera marina. Mar. Ecol. Prog. Ser. 1997, 157, 159–173. [Google Scholar] [CrossRef]
- Barreto, R.F.; Prado, R.M.; Leal, A.J.F.; Troleis, M.J.B.; Junior, G.B.S.; Monteiro, C.C.; Santos, L.C.N.; Carvalho, R.F. Mitigation of Ammonium Toxicity by Silicon in Tomato Depends on the Ammonium Concentration. Acta Agric. Scand. Sect. B Soil Plant Sci. 2016, 66, 483–488. [Google Scholar] [CrossRef]
- Roosta, H.R.; Schjoerring, J.K. Effects of Ammonium Toxicity on Nitrogen Metabolism and Elemental Profile of Cucumber Plants. J. Plant Nutr. 2007, 30, 1933–1951. [Google Scholar] [CrossRef]
- Silva, B.S.; Prado, R.D.M.; Hurtado, A.C.; de Andrade, R.A.; da Silva, G.P. Ammonia Toxicity Affects Cations Uptake and Growth in Papaya Plants Inclusive with Silicon Addition. Acta Biológica Colomb. 2020, 25, 345–353. [Google Scholar] [CrossRef]
- Bonomelli, C.; de Freitas, S.T.; Aguilera, C.; Palma, C.; Garay, R.; Dides, M.; Brossard, N.; O’Brien, J.A. Ammonium Excess Leads to Ca Restrictions, Morphological Changes, and Nutritional Imbalances in Tomato Plants, Which Can Be Monitored by the N/Ca Ratio. Agronomy 2021, 11, 1437. [Google Scholar] [CrossRef]
- De la Peña, M.; Marín-Peña, A.J.; Urmeneta, L.; Coleto, I.; Castillo-González, J.; van Liempd, S.M.; Falcón-Pérez, J.M.; Álvarez-Fernández, A.; González-Moro, M.B.; Marino, D. Ammonium Nutrition Interacts with Iron Homeostasis in Brachypodium distachyon. J. Exp. Bot. 2022, 73, 263–274. [Google Scholar] [CrossRef]
- Li, B.; Li, G.; Kronzucker, H.J.; Baluška, F.; Shi, W. Ammonium Stress in Arabidopsis: Signaling, Genetic Loci, and Physiological Targets. Trends Plant Sci. 2014, 19, 107–114. [Google Scholar] [CrossRef]
- Rogato, A.; D’Apuzzo, E.; Barbulova, A.; Omrane, S.; Parlati, A.; Carfagna, S.; Costa, A.; Schiavo, F.L.; Esposito, S.; Chiurazzi, M. Characterization of a Developmental Root Response Caused by External Ammonium Supply in Lotus japonicus. Plant Physiol. 2010, 154, 784–795. [Google Scholar] [CrossRef]
- Lima, J.E.; Kojima, S.; Takahashi, H.; von Wirén, N. Ammonium Triggers Lateral Root Branching in Arabidopsis in an AMMONIUM TRANSPORTER1; 3-Dependent Manner. Plant Cell 2010, 22, 3621–3633. [Google Scholar] [CrossRef]
- Zou, N.; Li, B.; Chen, H.; Su, Y.; Kronzucker, H.J.; Xiong, L.; Baluška, F.; Shi, W. GSA-1/ARG 1 Protects Root Gravitropism in Arabidopsis under Ammonium Stress. New Phytol. 2013, 200, 97–111. [Google Scholar] [CrossRef]
- Domínguez-Valdivia, M.D.; Aparicio-Tejo, P.M.; Lamsfus, C.; Cruz, C.; Martins-Loução, M.A.; Moran, J.F. Nitrogen Nutrition and Antioxidant Metabolism in Ammonium-tolerant And-sensitive Plants. Physiol. Plant. 2008, 132, 359–369. [Google Scholar] [CrossRef]
- Drath, M.; Kloft, N.; Batschauer, A.; Marin, K.; Novak, J.; Forchhammer, K. Ammonia Triggers Photodamage of Photosystem II in the Cyanobacterium Synechocystis sp. Strain PCC 6803. Plant Physiol. 2008, 147, 206–215. [Google Scholar] [CrossRef] [PubMed]
- Podgorska, A.; Gieczewska, K.; Łukawska-kuzma, K.; Rasmusson, A.G.; Gardeström, P.; Szal, B. Long-Term Ammonium Nutrition of Arabidopsis Increases the Extrachloroplastic NAD (P) H. Plant Cell Environ. 2013, 36, 2038–2049. [Google Scholar]
- Zhu, Z.; Gerendas, J.; Bendixen, R.; Schinner, K.; Tabrizi, H.; Sattelmacher, B.; Hansen, U.-P. Different Tolerance to Light Stress in NO3−-and NH4+-Grown Phaseolus vulgaris L. Plant Biol. 2000, 2, 558–570. [Google Scholar] [CrossRef]
- Patterson, K.; Cakmak, T.; Cooper, A.; Lager, I.D.A.; Rasmusson, A.G.; Escobar, M.A. Distinct Signalling Pathways and Transcriptome Response Signatures Differentiate Ammonium-and Nitrate-supplied Plants. Plant. Cell Environ. 2010, 33, 1486–1501. [Google Scholar] [CrossRef]
- Zhu, Y.; Di, T.; Xu, G.; Chen, X.I.; Zeng, H.; Yan, F.; Shen, Q. Adaptation of Plasma Membrane H+-ATPase of Rice Roots to Low PH as Related to Ammonium Nutrition. Plant. Cell Environ. 2009, 32, 1428–1440. [Google Scholar] [CrossRef]
- Khademi, S.; O’Connell, J., III; Remis, J.; Robles-Colmenares, Y.; Miercke, L.J.W.; Stroud, R.M. Mechanism of Ammonia Transport by Amt/MEP/Rh: Structure of AmtB at 1.35 A. Science 2004, 305, 1587–1594. [Google Scholar] [CrossRef]
- Pantoja, O. High Affinity Ammonium Transporters: Molecular Mechanism of Action. Front. Plant Sci. 2012, 3, 34. [Google Scholar] [CrossRef] [PubMed]
- Balkos, K.D.; Britto, D.T.; Kronzucker, H.J. Optimization of Ammonium Acquisition and Metabolism by Potassium in Rice (Oryza sativa L. Cv. IR-72). Plant. Cell Environ. 2010, 33, 23–34. [Google Scholar]
- Szczerba, M.W.; Britto, D.T.; Balkos, K.D.; Kronzucker, H.J. Alleviation of Rapid, Futile Ammonium Cycling at the Plasma Membrane by Potassium Reveals K+-Sensitive and-Insensitive Components of NH4+ Transport. J. Exp. Bot. 2008, 59, 303–313. [Google Scholar] [CrossRef]
- Coskun, D.; Britto, D.T.; Kronzucker, H.J. The Nitrogen–Potassium Intersection: Membranes, Metabolism, and Mechanism. Plant. Cell Environ. 2017, 40, 2029–2041. [Google Scholar] [CrossRef]
- Liu, Y.; von Wirén, N. Ammonium as a Signal for Physiological and Morphological Responses in Plants. J. Exp. Bot. 2017, 68, 2581–2592. [Google Scholar] [CrossRef] [PubMed]
- Lanquar, V.; Loqué, D.; Hormann, F.; Yuan, L.; Bohner, A.; Engelsberger, W.R.; Lalonde, S.; Schulze, W.X.; von Wirén, N.; Frommer, W.B. Feedback Inhibition of Ammonium Uptake by a Phospho-Dependent Allosteric Mechanism in Arabidopsis. Plant Cell 2009, 21, 3610–3622. [Google Scholar] [CrossRef] [PubMed]
- Neuhauser, B.; Dynowski, M.; Mayer, M.; Ludewig, U. Regulation of NH4+ Transport by Essential Cross Talk between AMT Monomers through the Carboxyl Tails. Plant Physiol. 2007, 143, 1651–1659. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Gu, R.; Xuan, Y.; Smith-Valle, E.; Loqué, D.; Frommer, W.B.; von Wirén, N. Allosteric Regulation of Transport Activity by Heterotrimerization of Arabidopsis Ammonium Transporter Complexes in Vivo. Plant Cell 2013, 25, 974–984. [Google Scholar] [CrossRef]
- Wang, Q.; Zhao, Y.; Luo, W.; Li, R.; He, Q.; Fang, X.; Michele, R.D.; Ast, C.; von Wirén, N.; Lin, J. Single-Particle Analysis Reveals Shutoff Control of the Arabidopsis Ammonium Transporter AMT1; 3 by Clustering and Internalization. Proc. Natl. Acad. Sci. USA 2013, 110, 13204–13209. [Google Scholar] [CrossRef] [PubMed]
- Bai, L.; Ma, X.; Zhang, G.; Song, S.; Zhou, Y.; Gao, L.; Miao, Y.; Song, C.-P. A Receptor-like Kinase Mediates Ammonium Homeostasis and Is Important for the Polar Growth of Root Hairs in Arabidopsis. Plant Cell 2014, 26, 1497–1511. [Google Scholar] [CrossRef]
- Engelsberger, W.R.; Schulze, W.X. Nitrate and Ammonium Lead to Distinct Global Dynamic Phosphorylation Patterns When Resupplied to Nitrogen-starved Arabidopsis Seedlings. Plant J. 2012, 69, 978–995. [Google Scholar] [CrossRef]
- Bai, L.; Zhou, Y.; Ma, X.; Gao, L.; Song, C.-P. Arabidopsis CAP1-Mediated Ammonium Sensing Required Reactive Oxygen Species in Plant Cell Growth. Plant Signal. Behav. 2014, 9, e29582. [Google Scholar] [CrossRef]
- Esteban, R.; Royo, B.; Urarte, E.; Zamarreño, A.M.; Garcia-Mina, J.M.; Moran, J.F. Both Free Indole-3-Acetic Acid and the Photosynthetic Efficiency Play a Relevant Role in the Response of Medicago truncatula to Urea and Ammonium Nutrition under Axenic Conditions. Front. Plant Sci 2016, 7, 140. [Google Scholar] [CrossRef]
- Yang, H.; von der Fecht-Bartenbach, J.; Friml, J.; Lohmann, J.U.; Neuhäuser, B.; Ludewig, U. Auxin-Modulated Root Growth Inhibition in Arabidopsis thaliana Seedlings with Ammonium as the Sole Nitrogen Source. Funct. Plant Biol. 2014, 42, 239–251. [Google Scholar] [CrossRef]
- Geiger, D.; Maierhofer, T.; Al-Rasheid, K.A.S.; Scherzer, S.; Mumm, P.; Liese, A.; Ache, P.; Wellmann, C.; Marten, I.; Grill, E. Stomatal Closure by Fast Abscisic Acid Signaling Is Mediated by the Guard Cell Anion Channel SLAH3 and the Receptor RCAR1. Sci. Signal. 2011, 4, ra32. [Google Scholar] [CrossRef] [PubMed]
- Flores, T.; Todd, C.D.; Tovar-Mendez, A.; Dhanoa, P.K.; Correa-Aragunde, N.; Hoyos, M.E.; Brownfield, D.M.; Mullen, R.T.; Lamattina, L.; Polacco, J.C. Arginase-Negative Mutants of Arabidopsis Exhibit Increased Nitric Oxide Signaling in Root Development. Plant Physiol. 2008, 147, 1936–1946. [Google Scholar] [CrossRef] [PubMed]
- Wimalasekera, R.; Villar, C.; Begum, T.; Scherer, G.F.E. COPPER AMINE OXIDASE1 (CuAO1) of Arabidopsis Thaliana Contributes to Abscisic Acid-and Polyamine-Induced Nitric Oxide Biosynthesis and Abscisic Acid Signal Transduction. Mol. Plant 2011, 4, 663–678. [Google Scholar] [CrossRef] [PubMed]
- Ariz, I.; Asensio, A.C.; Zamarreño, A.M.; García-Mina, J.M.; Aparicio-Tejo, P.M.; Moran, J.F. Changes in the C/N Balance Caused by Increasing External Ammonium Concentrations Are Driven by Carbon and Energy Availabilities during Ammonium Nutrition in Pea Plants: The Key Roles of Asparagine Synthetase and Anaplerotic Enzymes. Physiol. Plant. 2013, 148, 522–537. [Google Scholar] [CrossRef] [PubMed]
- Jeong, B.R. Ammonium and Nitrate Nutrition of Selected Bedding Plants. Ph.D. Thesis, Colorado State University, Fort Collins, CO, USA, 1991. [Google Scholar]
- Jeong, B.R.; Lee, C.W.; Daley, L.S. Use of Non-destructive In vivo Spectroscopic Analyses in Determining Leaf Chlorophyll Contents and Light Absorption Characteristics of Bedding Plants Grown with Different N Source. HortScience 1990, 25, 1098G–1098. [Google Scholar] [CrossRef]
- Jeong, B.R.; Lee, C.W. Influence of Ammonium, Nitrate, and Chloride on Solution PH and Ion Uptake by Ageratum and Salvia in Hydroponic Culture. J. Plant Nutr. 1996, 19, 1343–1360. [Google Scholar] [CrossRef]
- M’rah Helali, S.; Nebli, H.; Kaddour, R.; Mahmoudi, H.; Lachaâl, M.; Ouerghi, Z. Influence of Nitrate—Ammonium Ratio on Growth and Nutrition of Arabidopsis thaliana. Plant Soil 2010, 336, 65–74. [Google Scholar] [CrossRef]
- Masakapalli, S.K.; Kruger, N.J.; Ratcliffe, R.G. The Metabolic Flux Phenotype of Heterotrophic Arabidopsis Cells Reveals a Complex Response to Changes in Nitrogen Supply. Plant J. 2013, 74, 569–582. [Google Scholar] [CrossRef]
- Marino, D.; Ariz, I.; Lasa, B.; Santamaría, E.; Fernández-Irigoyen, J.; González-Murua, C.; Aparicio Tejo, P.M. Quantitative Proteomics Reveals the Importance of Nitrogen Source to Control Glucosinolate Metabolism in Arabidopsis thaliana and Brassica oleracea. J. Exp. Bot. 2016, 67, 3313–3323. [Google Scholar] [CrossRef]
- Fuertes-Mendizábal, T.; González-Torralba, J.; Arregui, L.M.; González-Murua, C.; González-Moro, M.B.; Estavillo, J.M. Ammonium as Sole N Source Improves Grain Quality in Wheat. J. Sci. Food Agric. 2013, 93, 2162–2171. [Google Scholar] [CrossRef]
- Cramer, M.D.; Lewis, O.A.M. The Influence of Nitrate and Ammonium Nutrition on the Growth of Wheat (Triticum aestivum) and Maize (Zea mays) Plants. Ann. Bot. 1993, 72, 359–365. [Google Scholar] [CrossRef]
- Raab, T.K.; Terry, N. Nitrogen Source Regulation of Growth and Photosynthesis in Beta vulgaris L. Plant Physiol. 1994, 105, 1159–1166. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.X.; Li, C.J.; Zhang, F.S. Transpiration, Potassium Uptake and Flow in Tobacco as Affected by Nitrogen Forms and Nutrient Levels. Ann. Bot. 2005, 95, 991–998. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Kaldenhoff, R.; Uehlein, N.; Sattelmacher, B.; Brueck, H. Relationship between Water and Nitrogen Uptake in Nitrate-and Ammonium-supplied Phaseolus vulgaris L. Plants. J. Plant Nutr. Soil Sci. 2007, 170, 73–80. [Google Scholar] [CrossRef]
- Gao, L.; Liu, M.; Wang, M.; Shen, Q.; Guo, S. Enhanced Salt Tolerance under Nitrate Nutrition Is Associated with Apoplast Na+ Content in Canola (Brassica napus L.) and Rice (Oryza sativa L.) Plants. Plant Cell Physiol. 2016, 57, 2323–2333. [Google Scholar] [CrossRef]
- Malagoli, M.; Dal Canal, A.; Quaggiotti, S.; Pegoraro, P.; Bottacin, A. Differences in Nitrate and Ammonium Uptake between Scots Pine and European Larch. Plant Soil 2000, 221, 1–3. [Google Scholar] [CrossRef]
- Horchani, F.; Hajri, R.; Aschi-Smiti, S. Effect of Ammonium or Nitrate Nutrition on Photosynthesis, Growth, and Nitrogen Assimilation in Tomato Plants. J. Plant Nutr. Soil Sci. 2010, 173, 610–617. [Google Scholar] [CrossRef]
- Guo, S.; Chen, G.; Zhou, Y.; Shen, Q. Ammonium Nutrition Increases Photosynthesis Rate under Water Stress at Early Development Stage of Rice (Oryza sativa L.). Plant Soil 2007, 296, 115–124. [Google Scholar] [CrossRef]
- Barber, S.A. Soil Nutrient Bioavailability: A Mechanistic Approach; John Wiley & Sons: Hoboken, NJ, USA, 1995. [Google Scholar]
- Ahmed, M.; Rauf, M.; Akhtar, M.; Mukhtar, Z.; Saeed, N.A. Hazards of Nitrogen Fertilizers and Ways to Reduce Nitrate Accumulation in Crop Plants. Environ. Sci. Pollut. Res. 2020, 27, 17661–17670. [Google Scholar] [CrossRef]
- Jeong, B.; Lee, C.W. Optimum Concentrations of NH4+, NO3−, and NH4++ NO3− for Petunia hybrida Grown in Peat-Lite and Rockwool Media. Hortic. Environ. Biotechnol. 2007, 48, 325–331. [Google Scholar]
- Zhang, Y.; Lv, H.; Wang, D.; Deng, J.; Song, W.; Makeen, K.; Shen, Q.; Xu, G. Partial Nitrate Nutrition Amends Photosynthetic Characteristics in Rice (Oryza sativa L. var. Japonica) Differing in Nitrogen Use Efficiency. Plant Growth Regul. 2011, 63, 235–242. [Google Scholar] [CrossRef]
- Zhang, F.; Kang, S.; Li, F.; Zhang, J. Growth and Major Nutrient Concentrations in Brassica campestris Supplied with Different NH4+/NO3− Ratios. J. Integr. Plant Biol. 2007, 49, 455–462. [Google Scholar] [CrossRef]
- Jesus, G.L.D.; Pauletti, V.; Zawadneak, M.A.C.; Cuquel, F.L. Strawberry Quality Affected by the Nitrate: Ammonium Ratio in the Nutrient Solution. Rev. Mex. Cienc. Agrícolas 2021, 12, 753–763. [Google Scholar]
- Zhang, J.; Lv, J.; Dawuda, M.M.; Xie, J.; Yu, J.; Li, J.; Zhang, X.; Tang, C.; Wang, C.; Gan, Y. Appropriate Ammonium-Nitrate Ratio Improves Nutrient Accumulation and Fruit Quality in Pepper (Capsicum annuum L.). Agronomy 2019, 9, 683. [Google Scholar] [CrossRef]
- Zhu, Y.; Qi, B.; Hao, Y.; Liu, H.; Sun, G.; Chen, R.; Song, S. Appropriate NH4+/NO3–Ratio Triggers Plant Growth and Nutrient Uptake of Flowering Chinese Cabbage by Optimizing the PH Value of Nutrient Solution. Front. Plant Sci. 2021, 12, 656144. [Google Scholar] [CrossRef]
- Song, J.; Yang, J.; Jeong, B.R. Decreased Solution PH and Increased K+ Uptake Are Related to Ammonium Tolerance in Hydroponically Cultured Plants. Horticulturae 2022, 8, 228. [Google Scholar] [CrossRef]
- Song, J.; Yang, J.; Jeong, B.R. Growth, Quality, and Nitrogen Assimilation in Response to High Ammonium or Nitrate Supply in Cabbage (Brassica campestris L.) and Lettuce (Lactuca sativa L.). Agronomy 2021, 11, 2556. [Google Scholar] [CrossRef]
- Narayan, O.P.; Kumar, P.; Yadav, B.; Dua, M.; Johri, A.K. Sulfur Nutrition and Its Role in Plant Growth and Development. Plant Signal. Behav. 2022, 2030082. [Google Scholar] [CrossRef] [PubMed]
- Chien, S.H.; Gearhart, M.M.; Villagarcía, S. Comparison of Ammonium Sulfate with Other Nitrogen and Sulfur Fertilizers in Increasing Crop Production and Minimizing Environmental Impact: A Review. Soil Sci. 2011, 176, 327–335. [Google Scholar] [CrossRef]
- Megda, M.X.V.; Mariano, E.; Leite, J.M.; Megda, M.M.; Trivelin, P.C.O. Ammonium Chloride as an Alternative Source of Nitrogen for Sugarcane during Two Consecutive Cycles. Pesqui. Agropecuária Bras. 2019, 54, e00329. [Google Scholar] [CrossRef]
- Ishikawa, N.; Ishioka, G.; Yanaka, M.; Takata, K.; Murakami, M. Effects of Ammonium Chloride Fertilizer and Its Application Stage on Cadmium Concentrations in Wheat (Triticum aestivum L.) Grain. Plant Prod. Sci. 2015, 18, 137–145. [Google Scholar] [CrossRef]
- Wang, X.-X.; Liu, S.; Zhang, S.; Li, H.; Maimaitiaili, B.; Feng, G.; Rengel, Z. Localized Ammonium and Phosphorus Fertilization Can Improve Cotton Lint Yield by Decreasing Rhizosphere Soil PH and Salinity. Field Crops Res. 2018, 217, 75–81. [Google Scholar] [CrossRef]
- Eid, M.A.M.; Abdel-Salam, A.A.; Salem, H.M.; Mahrous, S.E.; Seleiman, M.F.; Alsadon, A.A.; Solieman, T.H.I.; Ibrahim, A.A. Interaction Effects of Nitrogen Source and Irrigation Regime on Tuber Quality, Yield, and Water Use Efficiency of Solanum tuberosum L. Plants 2020, 9, 110. [Google Scholar] [CrossRef] [PubMed]
- Abd-Elrahman, S.; Fahim, M.; El-Morshedy, R.; Abdrabbo, M.; Hashem, F. Appropriate Ammonium-Nitrate Ratio Improves Vegetative Growth and Yield of Eggplant under Water Stress Conditions. Middle East J. Agric. Res. 2022, 11, 89–102. [Google Scholar]
- Sanz-Cobena, A.; Lassaletta, L.; Aguilera, E.; del Prado, A.; Garnier, J.; Billen, G.; Iglesias, A.; Sanchez, B.; Guardia, G.; Abalos, D. Strategies for Greenhouse Gas Emissions Mitigation in Mediterranean Agriculture: A Review. Agric. Ecosyst. Environ. 2017, 238, 5–24. [Google Scholar] [CrossRef]
- Cui, L.; Li, D.; Wu, Z.; Xue, Y.; Xiao, F.; Zhang, L.; Song, Y.; Li, Y.; Zheng, Y.; Zhang, J. Effects of Nitrification Inhibitors on Soil Nitrification and Ammonia Volatilization in Three Soils with Different PH. Agronomy 2021, 11, 1674. [Google Scholar] [CrossRef]
- Johnson, C.R.; Jarrell, W.M.; Menge, J.A. Influence of Ammonium: Nitrate Ratio and Solution PH on Mycorrhizal Infection, Growth and Nutrient Composition of Chrysanthemum morifolium var. Circus. Plant Soil 1984, 77, 151–157. [Google Scholar] [CrossRef]
- Shaviv, A.; Hagin, J. Interaction of Ammonium and Nitrate Nutrition with Potassium in Wheat. Fertil. Res. 1988, 17, 137–146. [Google Scholar] [CrossRef]
- Tabatabaei, S.J.; Fatemi, L.S.; Fallahi, E. Effect of Ammonium: Nitrate Ratio on Yield, Calcium Concentration, and Photosynthesis Rate in Strawberry. J. Plant Nutr. 2006, 29, 1273–1285. [Google Scholar] [CrossRef]
- Lobit, P.; López-Pérez, L.; Cárdenas-Navarro, R.; Castellanos-Morales, V.C.; Ruiz-Corro, R. Effect of Ammonium/Nitrate Ratio on Growth and Development of Avocado Plants under Hydroponic Conditions. Can. J. Plant Sci. 2007, 87, 99–103. [Google Scholar] [CrossRef]
- Dong, C.; Lu, Y.; Zhu, Y.; Zhou, Y.; Xu, Y.; Shen, Q. Effect of Homogeneous and Heterogeneous Supply of Nitrate and Ammonium on Nitrogen Uptake and Distribution in Tomato Seedlings. Plant Growth Regul. 2012, 68, 271–280. [Google Scholar] [CrossRef]
- Liu, G.; Du, Q.; Li, J. Interactive Effects of Nitrate-Ammonium Ratios and Temperatures on Growth, Photosynthesis, and Nitrogen Metabolism of Tomato Seedlings. Sci. Hortic. 2017, 214, 41–50. [Google Scholar] [CrossRef]
- Khalaj, M.A.; Kiani, S.; Khoshgoftarmanesh, A.H.; Amoaghaie, R. Growth, Quality, and Physiological Characteristics of Gerbera (Gerbera jamesonii L.) Cut Flowers in Response to Different NO3−: NH4+ Ratios. Hortic. Environ. Biotechnol. 2017, 58, 313–323. [Google Scholar] [CrossRef]
- Mantovani, C.; Prado, R.M.; Pivetta, K.F.L. Impact of Nitrate and Ammonium Ratio on Nutrition and Growth of Two Epiphytic Orchids. An. Acad. Bras. Ciências 2018, 90, 3423–3431. [Google Scholar] [CrossRef]
- Shang, H.Q.; Shen, G.M. Effect of Ammonium/Nitrate Ratio on Pak Choi (Brassica chinensis L.) Photosynthetic Capacity and Biomass Accumulation under Low Light Intensity and Water Deficit. Photosynthetica 2018, 56, 1039–1046. [Google Scholar] [CrossRef]
- Boschiero, B.N.; Mariano, E.; Azevedo, R.A.; Trivelin, P.C.O. Influence of Nitrate-Ammonium Ratio on the Growth, Nutrition, and Metabolism of Sugarcane. Plant Physiol. Biochem. 2019, 139, 246–255. [Google Scholar] [CrossRef]
- Assimakopoulou, A.; Salmas, I.; Kounavis, N.; Bastas, A.I.; Michopoulou, V.; Michail, E. The Impact of Ammonium to Nitrate Ratio on the Growth and Nutritional Status of Kale. Not. Bot. Horti Agrobot. Cluj-Napoca 2019, 47, 848–859. [Google Scholar] [CrossRef]
- BDR, M.F.; Widiayani, N.; Anshori, M.F. Ratio of Ammonium and Nitrate to Response of Chinese Broccoli Variety (Brassica oleracea var. Alboglabra) in Hydroponic Culture. IOP Conf. Ser. Earth Environ. Sci. 2020, 575, 12124. [Google Scholar]
- Carr, N.F.; Boaretto, R.M.; Mattos Jr, D. Coffee Seedlings Growth under Varied NO3−: NH4+ Ratio: Consequences for Nitrogen Metabolism, Amino Acids Profile, and Regulation of Plasma Membrane H+-ATPase. Plant Physiol. Biochem. 2020, 154, 11–20. [Google Scholar] [CrossRef]
- Li, S.; Zhang, H.; Wang, S.; Shi, L.; Xu, F.; Wang, C.; Cai, H.; Ding, G. The Rapeseed Genotypes with Contrasting NUE Response Discrepantly to Varied Provision of Ammonium and Nitrate by Regulating Photosynthesis, Root Morphology, Nutritional Status, and Oxidative Stress Response. Plant Physiol. Biochem. 2021, 166, 348–360. [Google Scholar] [CrossRef]
- Xu, J.; Fang, Y.; Tavakkoli, E.; Pan, X.; Liao, F.; Chen, W.; Guo, W. Preferential Ammonium: Nitrate Ratio of Blueberry Is Regulated by Nitrogen Transport and Reduction Systems. Sci. Hortic. 2021, 288, 110345. [Google Scholar] [CrossRef]
- Söylemez, S. The Impact of Different Growth Media and Ammonium-Nitrate Ratio on Yield and Nitrate Accumulation in Lettuce (Lactuca sativa var. longifolia). Not. Bot. Horti Agrobot. Cluj Napoca 2021, 49, 12540. [Google Scholar] [CrossRef]
- Hu, L.; Wu, Y.; Dawuda, M.M.; Liao, W.; Lv, J.; Li, Y.; Yu, J.; Xie, J.; Feng, Z.; Zhang, G. Appropriate Ammonium/Nitrate Mitigates Low Light Stress in Brassica pekinensis by Regulating the Nitrogen Metabolism and Expression Levels of Key Proteins. J. Plant Growth Regul. 2021, 40, 574–593. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, X.; Liu, H.; Sun, G.; Song, S.; Chen, R. High NH4+/NO3− Ratio Inhibits the Growth and Nitrogen Uptake of Chinese Kale at the Late Growth Stage by Ammonia Toxicity. Horticulturae 2021, 8, 8. [Google Scholar] [CrossRef]
- Pérez, A.H.; Santiago, J.C.G.; Torres, V.R.; López, A.M.; Rangel, A.S.; Montejo, N.C. Nitrate/Ammonium Ratio Effect on the Growth, Yield and Foliar Anatomy of Grafted Tomato Plants. Hortic. Sci. 2021, 48, 80–89. [Google Scholar] [CrossRef]
- Noh, K.; Jeong, B.R. Silicon Supplementation Alleviates Adverse Effects of Ammonium on Ssamchoo Grown in Home Cultivation System. Plants 2022, 11, 2882. [Google Scholar] [CrossRef] [PubMed]
- Naseri, A.; Alirezalu, A.; Noruzi, P.; Alirezalu, K. The Effect of Different Ammonium to Nitrate Ratios on Antioxidant Activity, Morpho-Physiological and Phytochemical Traits of Moldavian Balm (Dracocephalum moldavica). Sci. Rep. 2022, 12, 16841. [Google Scholar] [CrossRef]
- Santana, M.M.; Gonzalez, J.M.; Cruz, C. Nitric Oxide Accumulation: The Evolutionary Trigger for Phytopathogenesis. Front. Microbiol. 2017, 8, 1947. [Google Scholar] [CrossRef]
- Gupta, K.J.; Brotman, Y.; Segu, S.; Zeier, T.; Zeier, J.; Persijn, S.T.; Cristescu, S.M.; Harren, F.J.M.; Bauwe, H.; Fernie, A.R. The Form of Nitrogen Nutrition Affects Resistance against Pseudomonas syringae pv. phaseolicola in Tobacco. J. Exp. Bot. 2013, 64, 553–568. [Google Scholar] [CrossRef]
- Wang, M.; Sun, Y.; Gu, Z.; Wang, R.; Sun, G.; Zhu, C.; Guo, S.; Shen, Q. Nitrate Protects Cucumber Plants against Fusarium oxysporum by Regulating Citrate Exudation. Plant Cell Physiol. 2016, 57, 2001–2012. [Google Scholar] [CrossRef]
- López-Berges, M.S.; Rispail, N.; Prados-Rosales, R.C.; Di Pietro, A. A Nitrogen Response Pathway Regulates Virulence Functions in Fusarium oxysporum via the Protein Kinase TOR and the BZIP Protein MeaB. Plant Cell 2010, 22, 2459–2475. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Crespo, E. NH4 Protects Tomato Plants against Pseudomonas syringae by Activation of Systemic Acquired Acclimation. J. Exp. Bot. 2015, 66, 6777–6790. [Google Scholar] [CrossRef] [PubMed]
- González-Hernández, A.I.; Fernández-Crespo, E.; Scalschi, L.; Hajirezaei, M.-R.; von Wirén, N.; García-Agustín, P.; Camañes, G. Ammonium Mediated Changes in Carbon and Nitrogen Metabolisms Induce Resistance against Pseudomonas syringae in Tomato Plants. J. Plant Physiol. 2019, 239, 28–37. [Google Scholar] [CrossRef]
- Chi, W.J.; Wang, Z.Y.; Liu, J.M.; Zhang, C.; Wu, Y.H.; Bai, Y.J. Ammonium Uptake and Assimilation Are Required for Rice Defense against Sheath Blight Disease. Cereal Res. Commun. 2019, 47, 98–110. [Google Scholar] [CrossRef]
- Evans, J.; Scott, B.J.; Lill, W.J. Manganese Tolerance in Subterranean Clover (Trifolium subterraneum L.) Genotypes Grown with Ammonium Nitrate or Symbiotic Nitrogen. Plant Soil 1987, 97, 207–215. [Google Scholar] [CrossRef]
- Klotz, F.; Horst, W.J. Effect of Ammonium-and Nitrate-Nitrogen Nutrition on Aluminium Tolerance of Soybean (Glycine max L.). Plant Soil 1988, 111, 59–65. [Google Scholar] [CrossRef]
- Zhao, X.Q.; Shen, R.F.; Sun, Q. Bin Ammonium under Solution Culture Alleviates Aluminum Toxicity in Rice and Reduces Aluminum Accumulation in Roots Compared with Nitrate. Plant Soil 2009, 315, 107–121. [Google Scholar] [CrossRef]
- Chen, Z.C.; Zhao, X.Q.; Shen, R.F. The Alleviating Effect of Ammonium on Aluminum Toxicity in Lespedeza bicolor Results in Decreased Aluminum-Induced Malate Secretion from Roots Compared with Nitrate. Plant Soil 2010, 337, 389–398. [Google Scholar] [CrossRef]
- Wu, Z.; Jiang, Q.; Yan, T.; Zhang, X.; Xu, S.; Shi, H.; Li, F.; Du, Y.; Du, R.; Hu, C. Ammonium Nutrition Mitigates Cadmium Toxicity in Rice (Oryza sativa L.) through Improving Antioxidase System and the Glutathione-Ascorbate Cycle Efficiency. Ecotoxicol. Environ. Saf. 2020, 189, 110010. [Google Scholar] [CrossRef]
- Zhang, L.-D.; Liu, X.; Wei, M.-Y.; Guo, Z.-J.; Zhao, Z.-Z.; Gao, C.-H.; Li, J.; Xu, J.-X.; Shen, Z.-J.; Zheng, H.-L. Ammonium Has Stronger Cd Detoxification Ability than Nitrate by Reducing Cd Influx and Increasing Cd Fixation in Solanum nigrum L. J. Hazard. Mater. 2022, 425, 127947. [Google Scholar] [CrossRef]
- Liu, J.; Wang, J.; Wang, Z.; Li, M.; Liang, C.; Yang, Y.; Li, D.; Wang, R. Alleviation of Iron Deficiency in Pear by Ammonium Nitrate and Nitric Oxide. BMC Plant Biol. 2022, 22, 434. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Crespo, E. NH4 Inducesantioxidant Cellular Machinery and Provides Resistance to Salt Stress in Citrus Plants. Trees 2014, 28, 1693–1704. [Google Scholar] [CrossRef]
- Miranda, R.D.S.; Mesquita, R.O.; Costa, J.H.; Alvarez-Pizarro, J.C.; Prisco, J.T.; Gomes-Filho, E. Integrative Control between Proton Pumps and SOS1 Antiporters in Roots Is Crucial for Maintaining Low Na+ Accumulation and Salt Tolerance in Ammonium-Supplied Sorghum Bicolor. Plant Cell Physiol. 2017, 58, 522–536. [Google Scholar] [CrossRef] [PubMed]
- Hessini, K.; Issaoui, K.; Ferchichi, S.; Saif, T.; Abdelly, C.; Siddique, K.H.M.; Cruz, C. Interactive Effects of Salinity and Nitrogen Forms on Plant Growth, Photosynthesis and Osmotic Adjustment in Maize. Plant Physiol. Biochem. 2019, 139, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Tzortzakis, N.; Pitsikoulaki, G.; Stamatakis, A.; Chrysargyris, A. Ammonium to Total Nitrogen Ratio Interactive Effects with Salinity Application on Solanum lycopersicum Growth, Physiology, and Fruit Storage in a Closed Hydroponic System. Agronomy 2022, 12, 386. [Google Scholar] [CrossRef]
- Gao, Y.; Li, Y.; Yang, X.; Li, H.; Shen, Q.; Guo, S. Ammonium Nutrition Increases Water Absorption in Rice Seedlings (Oryza sativa L.) under Water Stress. Plant Soil 2010, 331, 193–201. [Google Scholar] [CrossRef]
- Cao, X.; Zhong, C.; Zhu, C.; Zhu, L.; Zhang, J.; Wu, L.; Jin, Q. Ammonium Uptake and Metabolism Alleviate PEG-Induced Water Stress in Rice Seedlings. Plant Physiol. Biochem. 2018, 132, 128–137. [Google Scholar] [CrossRef]
- Liu, Y.; Bai, L.; Sun, M.; Wang, J.; Li, S.; Miao, L.; Yan, Y.; He, C.; Yu, X.; Li, Y. Adaptation of Cucumber Seedlings to Low Temperature Stress by Reducing Nitrate to Ammonium during It’s Transportation. BMC Plant Biol. 2021, 21, 189. [Google Scholar] [CrossRef]
- Collado-González, J.; Piñero, M.C.; Otalora, G.; López-Marín, J.; Del Amor, F.M. Assessing Optimal Nitrate/Ammonium-Ratios in Baby-Leaf Lettuce to Enhance the Heat Stress Tolerance under Elevated CO2 Conditions. PLoS ONE 2022, 17, e0278309. [Google Scholar] [CrossRef]
- Kumar, P.; Eriksen, R.L.; Simko, I.; Shi, A.; Mou, B. Insights into Nitrogen Metabolism in the Wild and Cultivated Lettuce as Revealed by Transcriptome and Weighted Gene Co-Expression Network Analysis. Sci. Rep. 2022, 12, 9852. [Google Scholar] [CrossRef]
- Li, Y.; Wang, M.; Teng, K.; Dong, D.; Liu, Z.; Zhang, T.; Han, L. Transcriptome Profiling Revealed Candidate Genes, Pathways and Transcription Factors Related to Nitrogen Utilization and Excessive Nitrogen Stress in Perennial Ryegrass. Sci. Rep. 2022, 12, 3353. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, B.; Liu, F.; Luo, P.; Wang, Y.; Liu, D.; Wu, X.; Zhang, Z.; Wu, J. Transcriptomic and Physiological Analysis Revealed the Ammonium Tolerance Mechanisms of Myriophyllum aquaticum. Environ. Exp. Bot. 2021, 187, 104462. [Google Scholar] [CrossRef]
- Goel, P.; Sharma, N.K.; Bhuria, M.; Sharma, V.; Chauhan, R.; Pathania, S.; Swarnkar, M.K.; Chawla, V.; Acharya, V.; Shankar, R. Transcriptome and Co-Expression Network Analyses Identify Key Genes Regulating Nitrogen Use Efficiency in Brassica juncea L. Sci. Rep. 2018, 8, 7451. [Google Scholar] [CrossRef]
- Wang, W.; Li, R.; Zhu, Q.; Tang, X.; Zhao, Q. Transcriptomic and Physiological Analysis of Common Duckweed Lemna minor Responses to NH4+ Toxicity. BMC Plant Biol. 2016, 16, 92. [Google Scholar] [CrossRef]
- Li, W.; Xiang, F.; Zhong, M.; Zhou, L.; Liu, H.; Li, S.; Wang, X. Transcriptome and Metabolite Analysis Identifies Nitrogen Utilization Genes in Tea Plant (Camellia sinensis). Sci. Rep. 2017, 7, 1693. [Google Scholar] [CrossRef]
- Díaz-Silva, M.; Maldonado, J.; Veloso, P.; Delgado, N.; Silva, H.; Gallardo, J.A. RNA-Seq Analysis and Transcriptome Assembly of Salicornia neei Reveals a Powerful System for Ammonium Detoxification. Electron. J. Biotechnol. 2022, 58, 70–81. [Google Scholar] [CrossRef]
- Tschoep, H.; Gibon, Y.; Carillo, P.; Armengaud, P.; Szecowka, M.; Nunes-Nesi, A.; Fernie, A.R.; Koehl, K.; Stitt, M. Adjustment of Growth and Central Metabolism to a Mild but Sustained Nitrogen-limitation in Arabidopsis. Plant. Cell Environ. 2009, 32, 300–318. [Google Scholar] [CrossRef]
- Lin, Z.-H.; Chen, C.-S.; Zhong, Q.-S.; Ruan, Q.-C.; Chen, Z.-H.; You, X.-M.; Shan, R.-Y.; Li, X.-L. The GC-TOF/MS-Based Metabolomic Analysis Reveals Altered Metabolic Profiles in Nitrogen-Deficient Leaves and Roots of Tea Plants (Camellia sinensis). BMC Plant Biol. 2021, 21, 506. [Google Scholar] [CrossRef]
- González-Moro, M.B.; González-Moro, I.; De la Peña, M.; Estavillo, J.M.; Aparicio-Tejo, P.M.; Marino, D.; González-Murua, C.; Vega-Mas, I. A Multi-Species Analysis Defines Anaplerotic Enzymes and Amides as Metabolic Markers for Ammonium Nutrition. Front. Plant Sci. 2021, 11, 632285. [Google Scholar] [CrossRef]
- Buoso, S.; Tomasi, N.; Arkoun, M.; Maillard, A.; Jing, L.; Marroni, F.; Pluchon, S.; Pinton, R.; Zanin, L. Transcriptomic and Metabolomic Profiles of Zea mays Fed with Urea and Ammonium. Physiol. Plant. 2021, 173, 935–953. [Google Scholar] [CrossRef]
- Xun, Z.; Guo, X.; Li, Y.; Wen, X.; Wang, C.; Wang, Y. Quantitative Proteomics Analysis of Tomato Growth Inhibition by Ammonium Nitrogen. Plant Physiol. Biochem. 2020, 154, 129–141. [Google Scholar] [CrossRef]
- Setién, I.; Fuertes-Mendizabal, T.; González, A.; Aparicio-Tejo, P.M.; González-Murua, C.; González-Moro, M.B.; Estavillo, J.M. High Irradiance Improves Ammonium Tolerance in Wheat Plants by Increasing N Assimilation. J. Plant Physiol. 2013, 170, 758–771. [Google Scholar] [CrossRef]
- Vega-Mas, I.; Marino, D.; Sanchez-Zabala, J.; Gonzalez-Murua, C.; Estavillo, J.M.; González-Moro, M.B. CO2 Enrichment Modulates Ammonium Nutrition in Tomato Adjusting Carbon and Nitrogen Metabolism to Stomatal Conductance. Plant Sci. 2015, 241, 32–44. [Google Scholar] [CrossRef]
- Zheng, X.; He, K.; Kleist, T.; Chen, F.; Luan, S. Anion Channel SLAH3 Functions in Nitrate-dependent Alleviation of Ammonium Toxicity in Arabidopsis. Plant. Cell Environ. 2015, 38, 474–486. [Google Scholar] [CrossRef]
- Etesami, H.; Jeong, B.R. Silicon (Si): Review and Future Prospects on the Action Mechanisms in Alleviating Biotic and Abiotic Stresses in Plants. Ecotoxicol. Environ. Saf. 2018, 147, 881–896. [Google Scholar] [CrossRef]
- Campos, C.N.S.; Silva Júnior, G.B.D.; Prado, R.D.M.; David, C.H.O.D.; Souza Junior, J.P.D.; Teodoro, P.E. Silicon Mitigates Ammonium Toxicity in Plants. Agron. J. 2020, 112, 635–647. [Google Scholar] [CrossRef]
- Bittsánszky, A.; Pilinszky, K.; Gyulai, G.; Komives, T. Overcoming Ammonium Toxicity. Plant Sci. 2015, 231, 184–190. [Google Scholar] [CrossRef]
- Campos, C.N.S.; de Mello Prado, R.; Roque, C.G.; de Lima Neto, A.J.; Marques, L.J.P.; Chaves, A.P.; Cruz, C.A. Use of Silicon in Mitigating Ammonium Toxicity in Maize Plants. Am. J. Plant Sci. 2015, 6, 1780. [Google Scholar] [CrossRef]
- Campos, C.N.S.; de Mello Prado, R.; Caione, G. Silicon and Excess Ammonium and Nitrate in Cucumber Plants. African J. Agric. Res. 2016, 11, 276–283. [Google Scholar]
- Barreto, R.F.; Júnior, A.A.S.; Maggio, M.A.; de Mello Prado, R. Silicon Alleviates Ammonium Toxicity in Cauliflower and in Broccoli. Sci. Hortic. 2017, 225, 743–750. [Google Scholar] [CrossRef]
- Silva Júnior, G.B.D.; Prado, R.M.; Campos, C.; Agostinho, F.B.; Silva, S.L.O.; Santos, L.C.N.; González, L.C. Silicon Mitigates Ammonium Toxicity in Yellow Passionfruit Seedlings. Chil. J. Agric. Res. 2019, 79, 425–434. [Google Scholar] [CrossRef]
- Viciedo, D.O.; de Mello Prado, R.; Lizcano Toledo, R.; dos Santos, L.C.N.; Calero Hurtado, A.; Nedd, L.L.T.; Castellanos Gonzalez, L. Silicon Supplementation Alleviates Ammonium Toxicity in Sugar Beet (Beta vulgaris L.). J. Soil Sci. Plant Nutr. 2019, 19, 413–419. [Google Scholar] [CrossRef]
- Viciedo, D.O.; de Mello Prado, R.; Lizcano Toledo, R.; Salas Aguilar, D.; Dos Santos, L.C.N.; Calero Hurtado, A.; Peña Calzada, K.; Betancourt Aguilar, C. Physiological Role of Silicon in Radish Seedlings under Ammonium Toxicity. J. Sci. Food Agric. 2020, 100, 5637–5644. [Google Scholar] [CrossRef] [PubMed]
- De Souza Junior, J.P.; Prado, R.D.M.; de Morais, T.; Chagas, B.; Frazão, J.J.; dos Santos Sarah, M.M.; de Oliveira, K.R.; de Paula, R.C. Silicon Fertigation and Salicylic Acid Foliar Spraying Mitigate Ammonium Deficiency and Toxicity in Eucalyptus spp. Clonal Seedlings. PLoS ONE 2021, 16, e0250436. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Yang, J.; Jeong, B.R. Silicon Mitigates Ammonium Toxicity in Cabbage (Brassica campestris L. ssp. Pekinensis) ‘Ssamchu.’. Front. Sustain. Food Syst. 2022, 6, 922666. [Google Scholar] [CrossRef]
- Song, J.; Yang, J.; Jeong, B.R. Alleviation of Ammonium Toxicity in Salvia splendens ‘Vista Red’with Silicon Supplementation. Toxics 2022, 10, 446. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Mechanism of uptake and assimilation of nitrate and ammonium ions in plants. The transport of nitrate occurs through a nitrate transporter (NRT) followed by its conversion into nitrite by nitrate reductase (NR) in the cytoplasm and its subsequent reduction into ammonium ions by nitrite reductase (NiR) in the chloroplast. The nitrate transporters act as H+/NO3− symporters that depend on the proton-motive force produced by the plasma membrane H+-ATPases. According to the hypothesis postulated by Coskun et al. [19], ammonium (NH4+) is taken up as the non-electrogenic flux of NH3 through the plasma membrane, mediated by aquaporins (AQP). Ammonium is also transported through high-affinity ammonium transporters (AMT) and to some extent through the non-specific cation channels (NSCC) or K+ channels. The ammonium ion that enters the cell directly goes to the GS/GOGAT cycle, where NH4+ is assimilated into glutamine (Gln) by glutamine synthetase and glutamate by glutamate synthase (GOGAT) in the chloroplast. Glutamine can also be converted into asparagine (Asn) via asparagine synthetase (ASN). The enzyme glutamate dehydrogenase (GDH) yields ammonium and 2-oxoglutarate (2-OG), a product that links nitrogen metabolism with photorespiration, which involves three cell organelles, such as chloroplast, peroxisome, and mitochondria. The photorespiratory product, glyoxylate from chloroplast is converted into glycine by the action of glutamate: glyoxylate aminotransferase (GGT) in peroxisome, while two molecules of glycine are converted into one molecule of serine by the combined action of glycine decarboxylase (GDC) and serine hydroxymethyltransferase (SHMT) in mitochondria.
Figure 1.
Mechanism of uptake and assimilation of nitrate and ammonium ions in plants. The transport of nitrate occurs through a nitrate transporter (NRT) followed by its conversion into nitrite by nitrate reductase (NR) in the cytoplasm and its subsequent reduction into ammonium ions by nitrite reductase (NiR) in the chloroplast. The nitrate transporters act as H+/NO3− symporters that depend on the proton-motive force produced by the plasma membrane H+-ATPases. According to the hypothesis postulated by Coskun et al. [19], ammonium (NH4+) is taken up as the non-electrogenic flux of NH3 through the plasma membrane, mediated by aquaporins (AQP). Ammonium is also transported through high-affinity ammonium transporters (AMT) and to some extent through the non-specific cation channels (NSCC) or K+ channels. The ammonium ion that enters the cell directly goes to the GS/GOGAT cycle, where NH4+ is assimilated into glutamine (Gln) by glutamine synthetase and glutamate by glutamate synthase (GOGAT) in the chloroplast. Glutamine can also be converted into asparagine (Asn) via asparagine synthetase (ASN). The enzyme glutamate dehydrogenase (GDH) yields ammonium and 2-oxoglutarate (2-OG), a product that links nitrogen metabolism with photorespiration, which involves three cell organelles, such as chloroplast, peroxisome, and mitochondria. The photorespiratory product, glyoxylate from chloroplast is converted into glycine by the action of glutamate: glyoxylate aminotransferase (GGT) in peroxisome, while two molecules of glycine are converted into one molecule of serine by the combined action of glycine decarboxylase (GDC) and serine hydroxymethyltransferase (SHMT) in mitochondria.

Figure 2.
The major physiological changes and symptoms associated with ammonium toxicity and tolerance as well as the potential strategies to improve ammonium tolerance are illustrated. The ammonium toxicity is affected by various environmental factors and the extent of toxicity varies among species.
Figure 2.
The major physiological changes and symptoms associated with ammonium toxicity and tolerance as well as the potential strategies to improve ammonium tolerance are illustrated. The ammonium toxicity is affected by various environmental factors and the extent of toxicity varies among species.

Figure 3.
Extracellular and intracellular ammonium sensing events in plants. In low ammonium conditions, the crucial threonine residue T460 in the C-terminal domain of AMT1;1 is dephosphorylated allowing the ammonium transport pores to open and allows for ammonium uptake. AMT1;3 sends a signal to encourage lateral root development when roots are exposed to localized ammonium, which leads to ammonium scavenging. AMT1;1 is rendered inactive by phosphorylation of its C-terminal domain in response to increased ammonium availability. Elevated cytosolic ammonium levels cause AMT1;3 to be clustered and internalized into the cytoplasm by the endocytic pathway, which further stops the uptake of ammonia. The tonoplast-localized receptor-like kinase CAP1 facilitates the compartmentation of ammonium into vacuoles mediated through aquaporins (AQP), which protects cells from ammonium toxicity.
Figure 3.
Extracellular and intracellular ammonium sensing events in plants. In low ammonium conditions, the crucial threonine residue T460 in the C-terminal domain of AMT1;1 is dephosphorylated allowing the ammonium transport pores to open and allows for ammonium uptake. AMT1;3 sends a signal to encourage lateral root development when roots are exposed to localized ammonium, which leads to ammonium scavenging. AMT1;1 is rendered inactive by phosphorylation of its C-terminal domain in response to increased ammonium availability. Elevated cytosolic ammonium levels cause AMT1;3 to be clustered and internalized into the cytoplasm by the endocytic pathway, which further stops the uptake of ammonia. The tonoplast-localized receptor-like kinase CAP1 facilitates the compartmentation of ammonium into vacuoles mediated through aquaporins (AQP), which protects cells from ammonium toxicity.

Table 2.
Alleviation of ammonium toxicity using silicon supplementation in various plant species.
| PLANT SPECIES | Mode of Si Application in the Cultivation System | Form and Concentration of Si Used | Mitigating Effects of Si under Ammonium Toxicity | Reference |
|---|---|---|---|---|
| Zea mays L. | In the nutrient solution of the hydroponics system | 10 mmol L−1 K2SiO3 | Increased N accumulation, shoot, and root dry mass production | [150] |
| Lycopersicon esculentum Mill. | In the nutrient solution of the hydroponics system | 1 mmol L−1 monosilicic acid | Improved N and Si use efficiency, increased leaf area, root area, dry biomass, total chlorophyll, carotenoids, and reduced oxidative stress | [22] |
| Cucumis sativus L. var. Hokushin and Tsubasa | In the nutrient solution of the hydroponics system | 0, 1, and 10 mmol L−1 K2SiO3 | Greater dry mass, N accumulation, green color index (GCI), and increased nitrate reductase activity | [151] |
| Brassica oleracea var. Botrytis cv Barcelona and Brassica oleracea var. Italica) cv BRO 68 | In the nutrient solution of the hydroponics system | 2 mmol L−1 silicon using stabilized sodium and potassium silicate (114.9 g L−1 Si and 18.9 g L−1 K2O). | Increased growth characteristics, physical integrity, total chlorophyll content, stomatal conductance, water use efficiency, accumulation of ions (N, K, Ca, Mg, and Si), and reduced electrolyte leakage | [152] |
| Passiflora edulis Sims f. flavicarpa O. Deg. | In the nutrient solution of the vermiculate cultivation | 2 mmol L−1 K2SiO3 | Greater stem and root diameter, root length, dry matter, leaf green color index (GCI), increased accumulation of ions (N, Si, K, Ca, and Mg), improved NUE, and reduced electrolyte leakage | [153] |
| Beta vulgaris L. | In the nutrient solution of the vermiculate cultivation in growth chamber | 28.5 g L−1 H4SiO4 | Improved photosynthesis, stomatal conductance, transpiration, dry mass production, and accumulation of N and Si in roots | [154] |
| Raphanus sativus L. | In the nutrient solution of the hydroponics system | 2 mmol L−1 K2SiO3 | Improved photosynthesis, transpiration, stomatal conductance, water-use efficiency, and higher total dry biomass | [155] |
| Eucalyptus urophylla x Eucalyptus grandis | Fertigation of Si in vermiculate cultivation | 2 mmol L−1 K2SiO3 | Increased dry mass production and improved the photosynthetic apparatus by decreasing fluorescence, and improving the quantum efficiency of photosystem II | [156] |
| Brassica campestris L. ssp. pekinensis | In the nutrient solution of the hydroponics system | 0.0 and 1.0 me·L−1 K2SiO3 | Improved growth parameters, photosynthesis, stomatal conductance, cation accumulation (Ca, Mg, K, and Si) antioxidant enzyme activities, and N assimilation enzyme activities | [157] |
| Salvia splendens ‘Vista Red’ | In the nutrient solution of the hydroponics substrate system | 0.0 and 1.0 me·L−1 K2SiO3 | Improved growth parameters, photosynthesis, stomatal conductance, cation accumulation (Ca, Mg, K, and Si) antioxidant enzyme activities, and N assimilation enzyme activities as well as reduced accumulation of ROS, and malondialdehyde (MDA) | [158] |
| Brassica lee ssp. namai Ssamchoo ‘Chunssamhwang51’ | In the nutrient solution of the hydroponics system | 0.0 and 10.7 me·L−1 K2SiO3 | Increased shoot fresh weight, chlorophyll content, leaf area, vitamin C, and antioxidant enzyme activities | [108] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Shilpha, J.; Song, J.; Jeong, B.R. Ammonium Phytotoxicity and Tolerance: An Insight into Ammonium Nutrition to Improve Crop Productivity. Agronomy 2023, 13, 1487. https://doi.org/10.3390/agronomy13061487
AMA Style
Shilpha J, Song J, Jeong BR. Ammonium Phytotoxicity and Tolerance: An Insight into Ammonium Nutrition to Improve Crop Productivity. Agronomy. 2023; 13(6):1487. https://doi.org/10.3390/agronomy13061487
Chicago/Turabian StyleShilpha, Jayabalan, Jinnan Song, and Byoung Ryong Jeong. 2023. "Ammonium Phytotoxicity and Tolerance: An Insight into Ammonium Nutrition to Improve Crop Productivity" Agronomy 13, no. 6: 1487. https://doi.org/10.3390/agronomy13061487
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.