1. Introduction
Arbuscular mycorrhizal fungi (AMF) in the soil can colonize the roots of most terrestrial plants, thus establishing a reciprocal symbiosis with plants [
1,
2,
3]. AMF are not strictly specific and can therefore form mycorrhizal structures with most plants [
4]. An AMF strain can colonize various plants, and a plant can be colonized by a variety of AMF [
5]. Nevertheless, AMF play an irreplaceable role in regulating plant interspecific competition and the soil nutrient cycle [
6,
7]. AMF can regulate the absorption of soil nutrients by plants, enhance stress resistance, stabilize soil structure, improve soil physical and chemical properties, and improve a plant’s competitiveness and survival rate [
8,
9].
AMF have diverse effects on a host plant’s growth, including improvement, inhibition, and no significant effect [
10]. Jifon et al. [
11] reported that
Rhizoglomus intraradices inoculation increased the growth of
Citrus aurantium plants at suitable CO
2 concentrations, while an inhibition of plant growth under mycorrhization conditions was found at high CO
2 concentrations. The growth changes of tomatoes inoculated with
R. intraradices depended on the tomato varieties, in which the fungus improved plant growth and enhanced antioxidant enzyme activities in the Momotaro variety but not the Rodeo variety [
12]. In addition, different AMF species have diverse effects on the same host plant. For example, in
Vicia villosa plants,
R.
intraradices showed a prominent positive effect on plant growth, root architecture, and sugar concentrations among
Diversispora spurca,
Funneliformis mosseae, and
R.
intraradices [
13]. The competitive ratio contribution of
F. mosseae to P was higher than that of
R. intraradices in the maize–rape intercropping [
14]. Yao et al. [
15] also found that inoculation with
F. mosseae and
Glomus versiforme significantly improved plant growth in maize, soybean, and white clover but not wheat, along with the decreasing order of maize > soybean > white clover > wheat in terms of mycorrhizal dependence. Earlier studies revealed that AMF roles in maize are dependent on the soil conditions, fungal isolated environment, and tillage systems [
16,
17,
18]. These results suggest that the responses between mycorrhizal fungi and plants are variable and complex.
Soil aggregates are an important component of soil structure, as they influence processes such as soil organic carbon (SOC) mineralization and microbial activity [
19,
20]. The formation of soil aggregates is influenced by various biotic and abiotic factors [
21]. The distribution and stability of soil aggregates are commonly used to assess soil structure [
22]. Macroaggregates (>0.25 mm) are formed and stabilized by soil organic matter, root exudates, and microorganisms [
20]. AMF make important contributions to changing soil fertility, structure, and stability [
23,
24]. AMF enhance soil structure through the entanglement of their hyphae and the release of glomalin-related soil protein (GRSP), which, thus, promotes the formation and stability of soil aggregates [
25]. In Lane Late navel orange, field inoculation with mycorrhizal fungi changed soil nutrient levels to some extent, dependent on the AMF species [
26]. For example,
D.
spurca did not change soil nitrate nitrogen levels, but it inhibited soil available K levels, and increased soil Olsen-P and ammonium nitrogen levels;
D.
versiformis inhibited the levels of ammonium nitrogen, nitrate nitrogen, and available K levels, but increased the levels of Olsen-P in soil. These AMF inoculations, however, collectively improved soil aggregate stability in Lane Late navel orange, as compared with non-AMF control. In Newhall navel orange,
D.
versiformis and
D.
spurca increased soil Olsen-P levels and aggregate stability [
27]. Inoculation with
F.
mosseae,
D.
spurca, and
R.
intraradices significantly reduced soil Olsen-P and available K levels in vetch [
13]. As a result, AMF inoculation can change soil nutrient levels and soil aggregate stability and distribution, but whether this effect also depends on the host plant species has not been clarified.
Marigold (
Tagetes erecta L.) is an ornamental plant used in medicine and cosmetics due to its rich lutein [
28]. Maize (
Zea mays L.) is an important food crop [
29]. White clover (
Trifolium repens L.) is an important perennial legume forage [
30,
31]. Vetch (
Vicia villosa Roth.) is an annual herb in the legumes, which has the characteristics of increasing soil fertility and being a green fertilizer [
13]. The present study assessed the responses of the four plants to a popular arbuscular mycorrhizal fungus (
Rhizoglomus intraradices) in terms of plant growth, leaf gas exchange, sugar content, and soil fertility, aggregate distribution, and stability.
4. Discussion
AMF improve plant growth by promoting the uptake of nutrients and water from the soil and improving root growth in the host plant [
13]. The degree of mycorrhizal dependence is an important factor in assessing the maximum plant growth of mycorrhizal plants in given soil conditions [
42]. In this study, the growth performances of marigold, maize, clover, and vetch were improved to different degrees after inoculation with
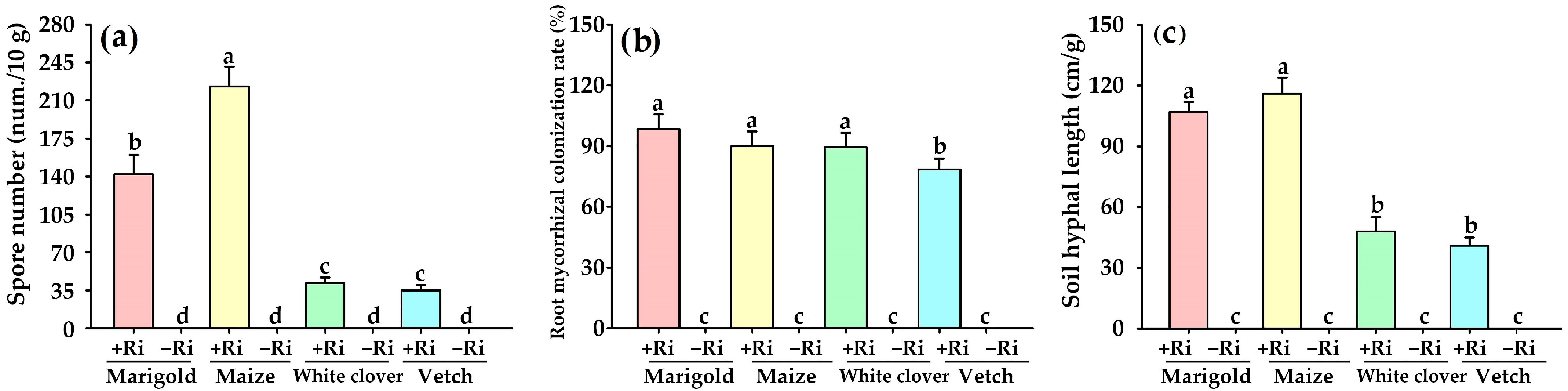
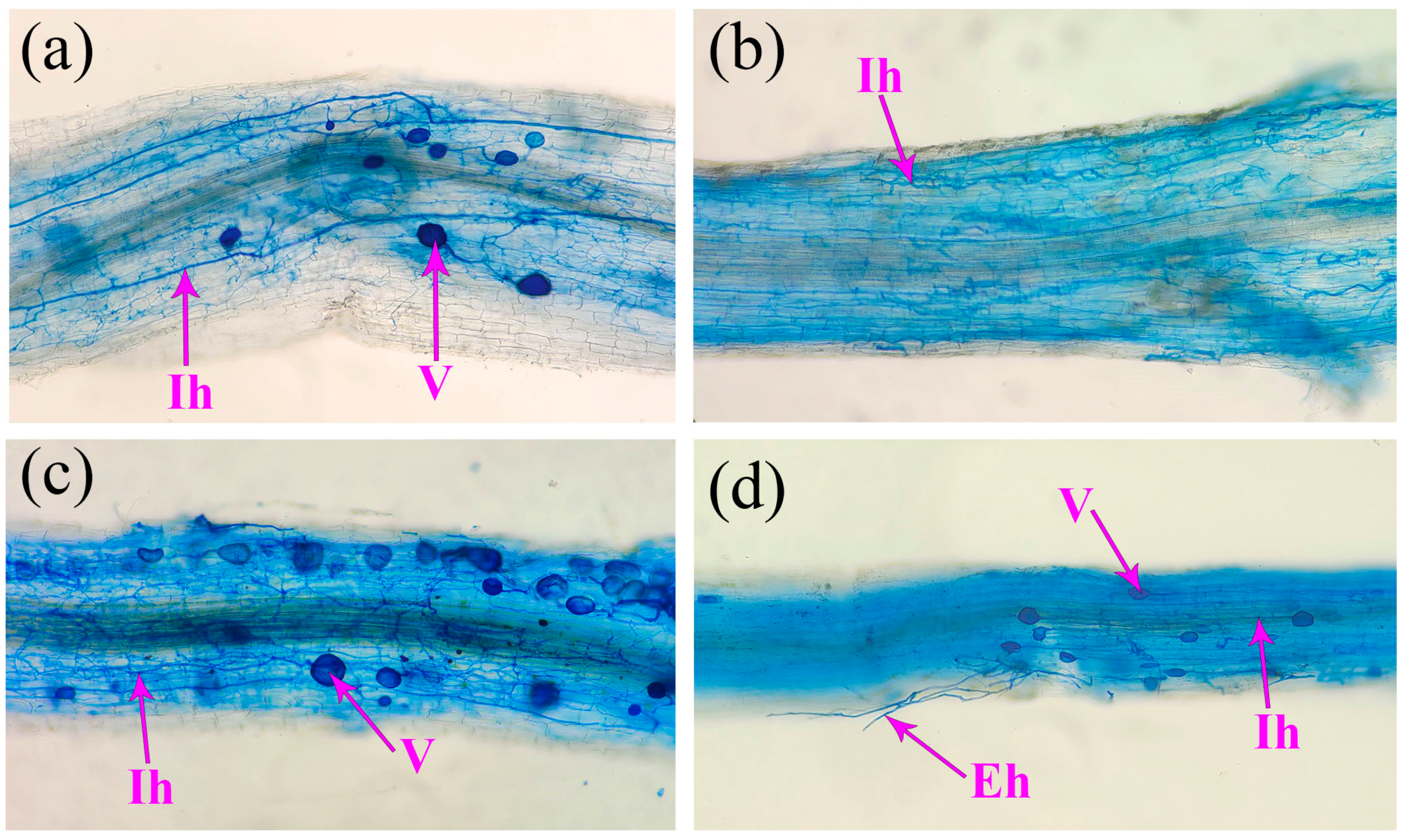
R. intraradices, with marigold showing the most pronounced effect. This indicates that AMF-improved plant growth is dependent on the host plant species. In addition, the root AMF colonization rate, soil hyphal length, and soil spore density are important indicators to evaluate the symbiotic status between AMF and host plants [
7,
43]. Our results indicated that mycorrhizal development was variable among the four plant species, with marigold and maize showing better mycorrhizal development than white clover and vetch. This is consistent with the results of Du [
44] inoculating
G.
mosseae on five greenery plants (
Ligustrum lucidum,
Viburnum odoratissimum,
Osmanthus fragrans,
Galium ordoratum, and
Paeonia lactiflora), showing the selectivity between mycorrhizal fungi and hosts. Nevertheless, the growth-promoting effect was not significant in maize after inoculation with
R. intraradices. This result may be due to the fact that maize is a field crop, which has limited root growth under potted conditions and thus affects the mycorrhizal function [
45]. On the other hand, it is related to the number of host root hairs, such as maize, which has dense root hairs and is less dependent on mycorrhizae [
46]. Soil nutrient levels and environmental conditions also influence AMF to improve plant growth [
16,
18].
Root morphology is a critical trait describing the variability of roots in response to environmental conditions [
47], of which root length indicates the spatial range of root absorption, root surface area reflects the root-to-soil contact area, and root diameter affects water transfer efficiency [
48]. This study showed that inoculation with
R. intraradices improved to some extent root morphological variables, such as total length, area, diameter, and volume in marigold, root diameter in maize, root surface area and diameter in white clover, and root total length, surface area, and diameter in vetch. This is in agreement with the results of Qu et al. [
49] on
Zenia insignis plants colonized by
F.
mosseae and
R. intraradices. In fact, earlier findings revealed that AMF improved root development in host plants by promoting root auxins, cytokinins, and polyamine levels [
47,
50]. Such better root morphology in mycorrhizal versus non-mycorrhizal plants would support the expansion of root nutrient uptake and improve water conduction efficiency.
This study demonstrated the variable effect of
R.
intraradices on the leaf gas exchange of host plants, with no significant changes in marigold and vetch, increased stomatal conductance and transpiration rate in maize, and increased net photosynthetic rate and stomatal conductance in white clover. Such results are in agreement with the findings of Frosi et al. [
51] in woody plants and Sonal et al. [
52] in maize. In a meta-analysis, the effect of AMF on leaf gas exchange variables of plants was regulated by various factors, including plant species, growth habits, soil types, experimental conditions, soil moisture, and salinity levels [
53]. Goicoechea et al. [
54] found that the effect of mycorrhizae on leaf gas exchange was associated with the balance between abscisic acid and cytokinins by AMF. However, whether such an association also appeared in the present study remains to be investigated.
In roots, the presence of mycorrhizal symbionts usually triggers the cleavage of sucrose in the host to glucose and fructose, which are then converted to other types of glycogen for mycorrhizal growth [
13,
55]. Approximately 4–20% of photosynthetic products are transferred from the host to the mycorrhiza [
56]. In the present study,
R. intraradices distinctly increased the Chi of marigold and white clover only. The root AMF colonization rate was significantly and positively correlated with the Chi (
r = 0.41,
p < 0.05), which may be due to the increase in the host’s Fe and Mg acquisition under mycorrhization conditions [
57]. Correspondingly, inoculated marigold presented significantly higher leaf glucose than uninoculated plants.
R. intraradices inoculation also triggered elevated sucrose in the leaves and roots of maize, fructose and glucose concentrations in the roots, and sucrose concentrations in the leaves and roots of white clover and vetch. Thus, mycorrhizal plants could provide a carbon source into the roots for mycorrhizal development. In fact, higher chlorophyll levels of mycorrhizal versus non-mycorrhizal plants are associated with higher rates of photosynthesis and carbon sequestration, thus maintaining AMF–plant symbiosis [
58].
Another important function of AMF is to improve the root microenvironment, including root secretions, soil fertility, and soil structure [
13]. Although
R. intraradices did not significantly alter rhizosphere NH
4+-N and NO
3−-N levels, it significantly increased soil available K levels of marigold and Olsen-P levels of white clover, while it decreased available K levels of white clover and vetch. Meng et al. [
55] also found that
F.
mosseae reduced soil available K levels of trifoliate orange but promoted soil Olsen-P levels. He et al. [
13] also observed a decrease in soil available K and Olsen-P in vetch after
R. intraradices colonization. As a consequence, the effect of
R. intraradices on soil fertility is variable. The elevation of soil Olsen-P in white clover after AMF inoculation may imply that acid phosphatases of both mycorrhizal fungi and hosts are secreted into the soil to decompose organic P [
58].
SOC is a marker of soil fertility, as well as a key component of biosphere stability and sustainability [
19]. In most cases, AMF accelerate SOC formation and sequestration and also provide physical protection for SOC [
59,
60]. In our study, the SOC of marigold, maize, and white clover significantly increased after inoculation with
R. intraradices, and there was no significant change in vetch. Additionally, there was a significantly positive correlation between root mycorrhizal colonization and SOC. The increase in SOC is derived from AMF secretion, mycorrhizal extraradical hyphal turnover (the dominant pathway), and carbon input from fine root turnover [
61], which may be a key factor influencing SOC storage capacity after inoculation with
R. intraradices [
62].
The present study showed that a relatively high EE-GRSP level was observed in the rhizosphere of maize plants inoculated with
R. intraradices, and higher DE-GRSP and T-GRSP levels in mycorrhizal marigold and vetch, varying by GRSP species and host plants. Holátko et al. [
63] also concluded that there was not a direct relationship between GRSP levels and the abundance of AMF because GRSP is derived from proteins of both mycorrhizal and non-mycorrhizal fungal origin [
64]. In our study, the root colonization rate, soil hyphal length, and spore density were significantly positively correlated with DE-GRSP and T-GRSP levels, plus there was a positive correlation between EE-GRSP and soil hyphal length and spore density. However, an important function of GRSP is to glue WSAs and stabilize WSAs [
23], and mycorrhizal extraradical hyphae are also involved in the gluing of WSA for improved soil structure [
25,
65,
66]. In this study, inoculation with
R.
intraradices significantly improved WSA distribution at a certain size level in the host plant, which in turn improved the stability of WSA in marigold, white clover, and vetch. This is in agreement with the results obtained by Xu et al. [
67] in tomato and maize. Correlation analysis also revealed that the root AMF colonization rate was significantly positively correlated with WSA in 0.5–4 mm, plus there was a significantly positive correlation between MWD and root colonization, soil hyphal length, soil spore density, SOC, DE-GRSP, and T-GRSP. Better soil WSA distribution and stability in mycorrhizal versus non-mycorrhizal plants is the comprehensive result of mycorrhizae, GRSP, and SOC. How GRSP stabilizes WSA should be further deciphered in the future. In addition, soil WSA stability is important for maintaining soil structure, porosity, erosion resistance, and water-holding capacity [
68]. Therefore, mycorrhizal plants have a great root microenvironment, which is particularly important for plant growth and stress tolerance [
41].

 ,
, 





{kind=link}
{kind=link}
{kind=link}