
Transcriptome Sequence Analysis of Defense Response of Resistant and Susceptible Bottle Gourd to Powdery Mildew
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Planting Material and Pathogen Infection
2.2. mRNA Extraction and Sequencing Data
2.3. GO and KEGG Analysis
2.4. Quantitative RT-PCR Assay
2.5. Regulatory Network Construction
3. Results
3.1. Changes in the Disease-Resistance Phenotype
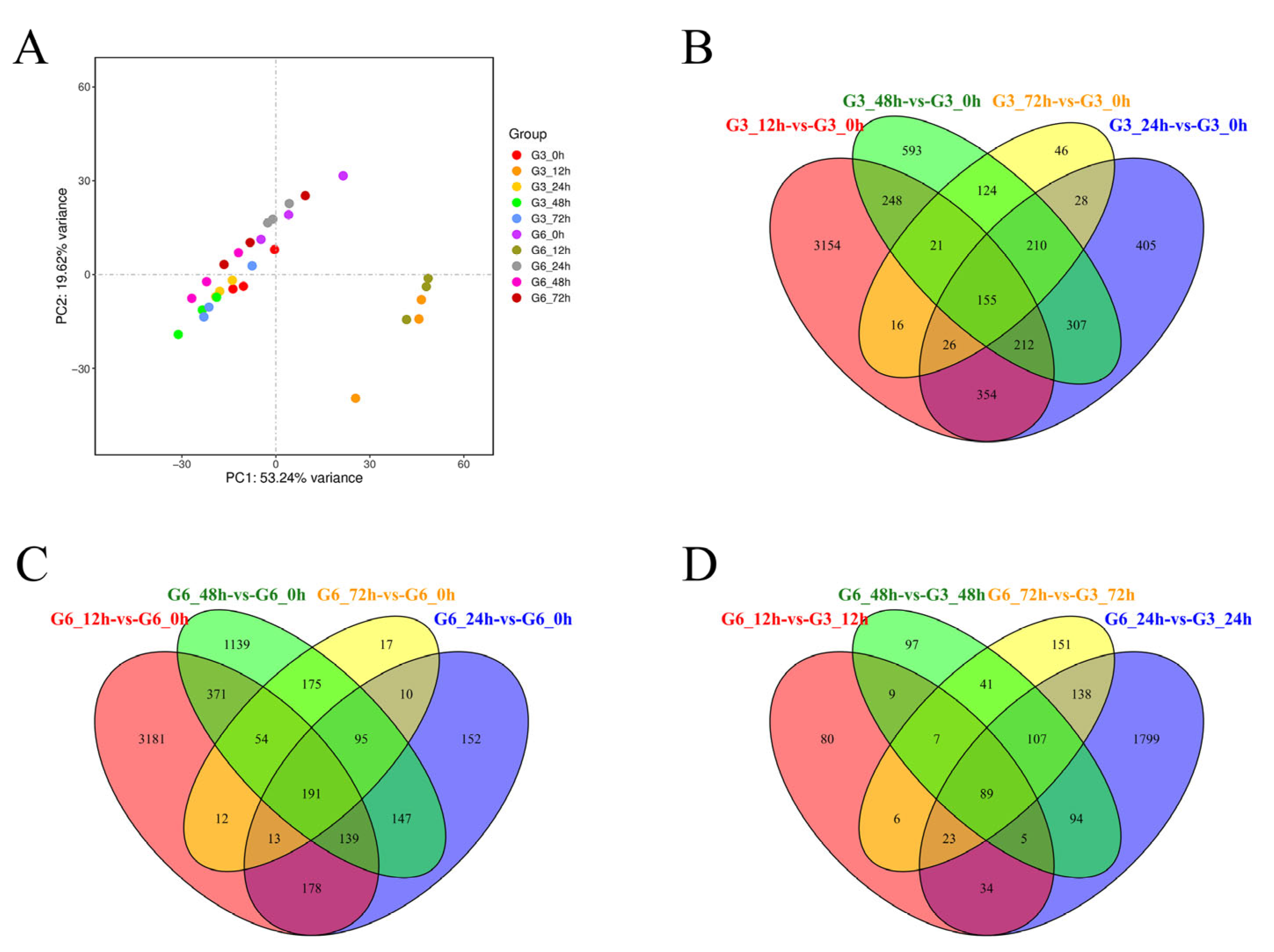
3.2. Overview of Sequencing Data and Comparison Results
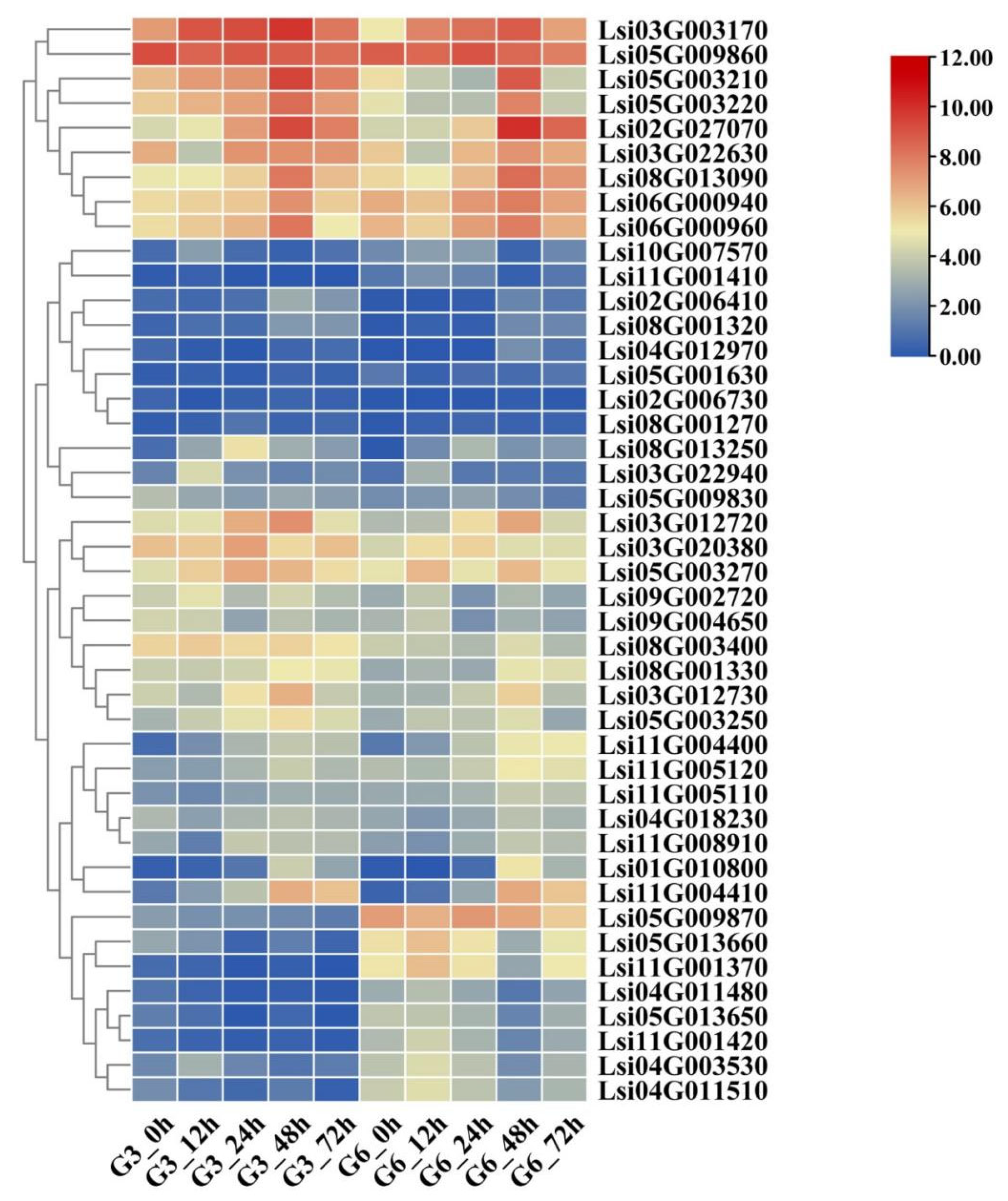
3.3. Identification and Expression Pattern of mRNA
3.4. GO Functional Analysis of Significantly Different Genes
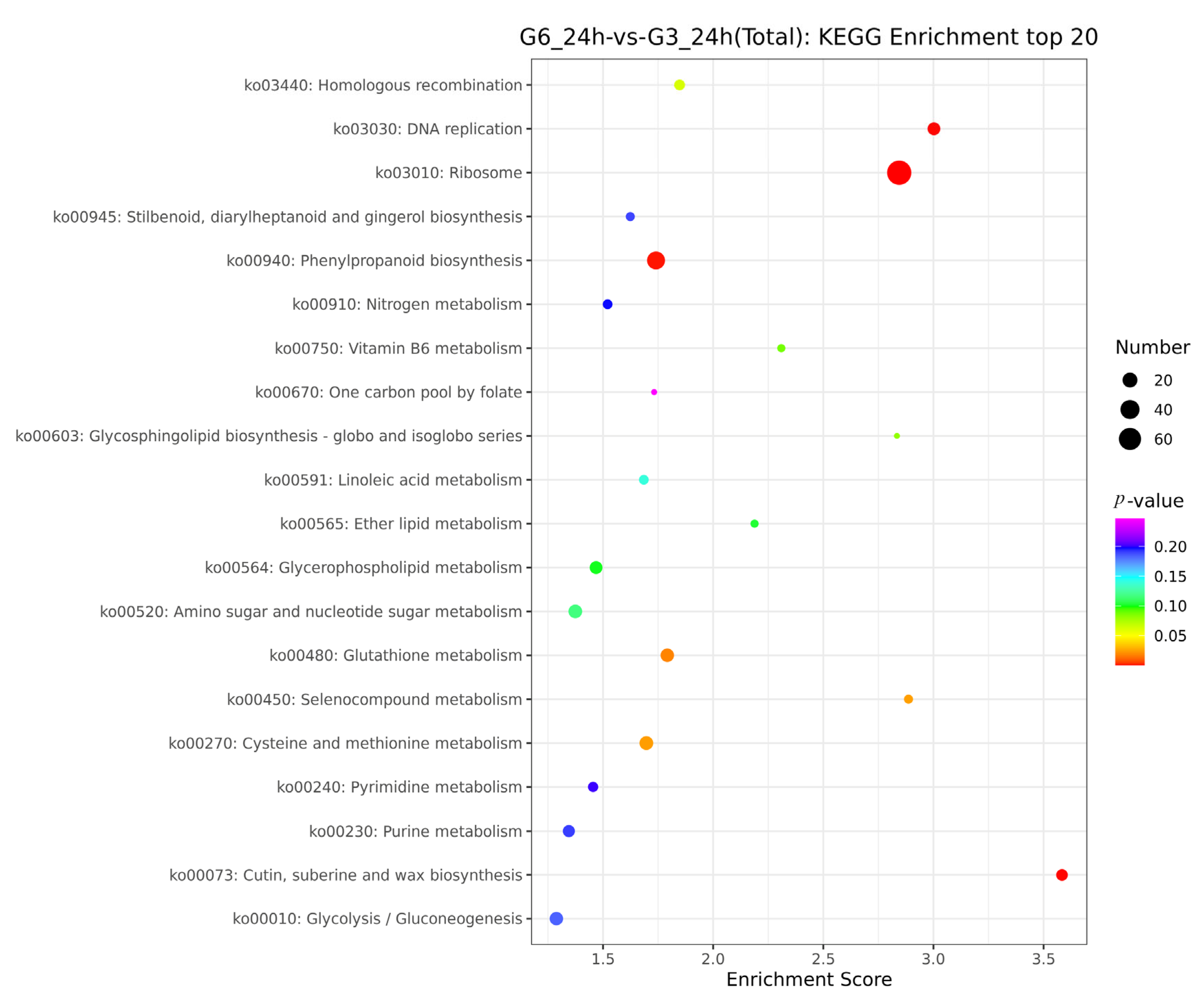
3.5. Analysis of the KEGG Pathway for Significantly Different Genes
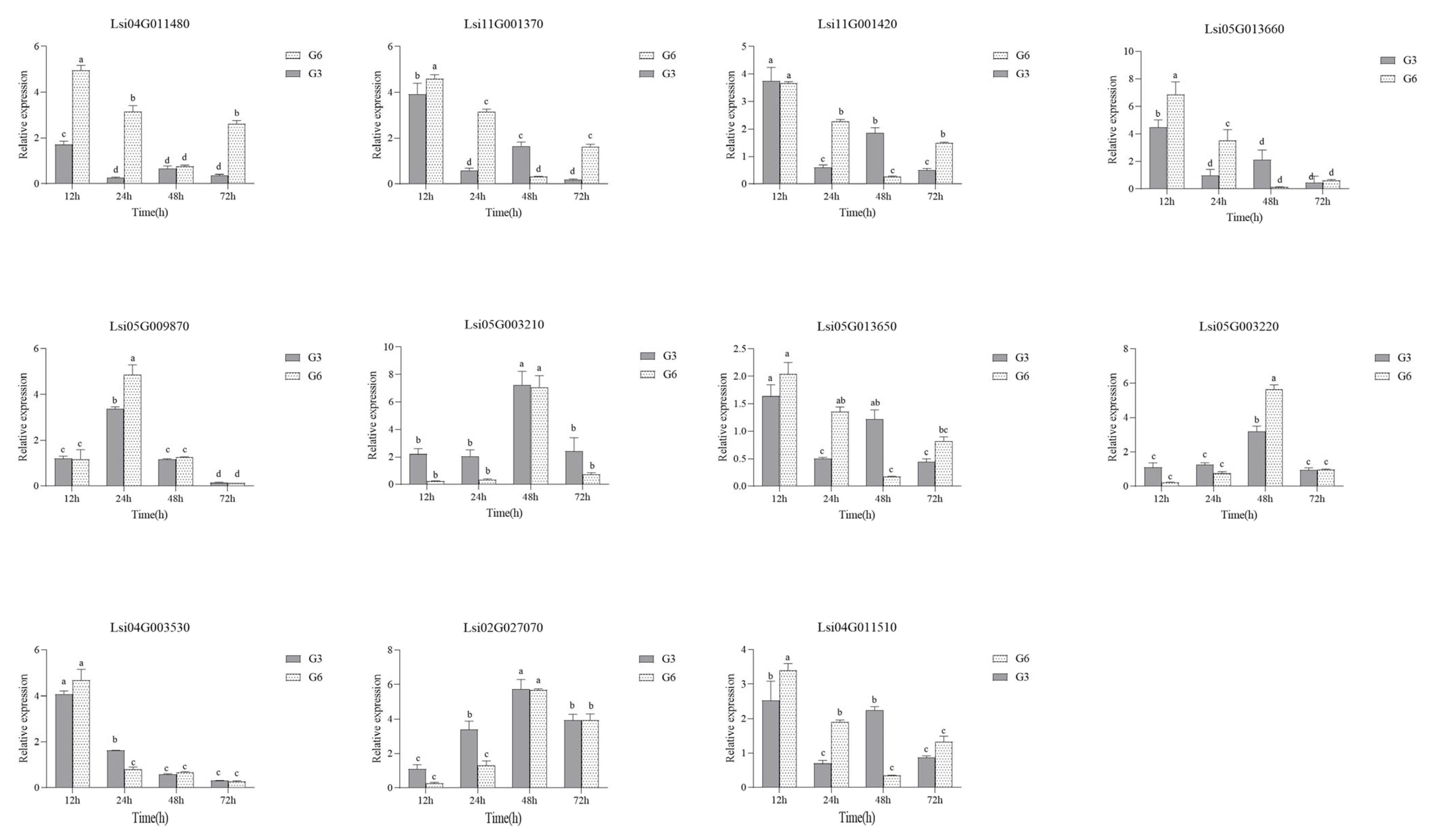
3.6. qRT-PCR Validation
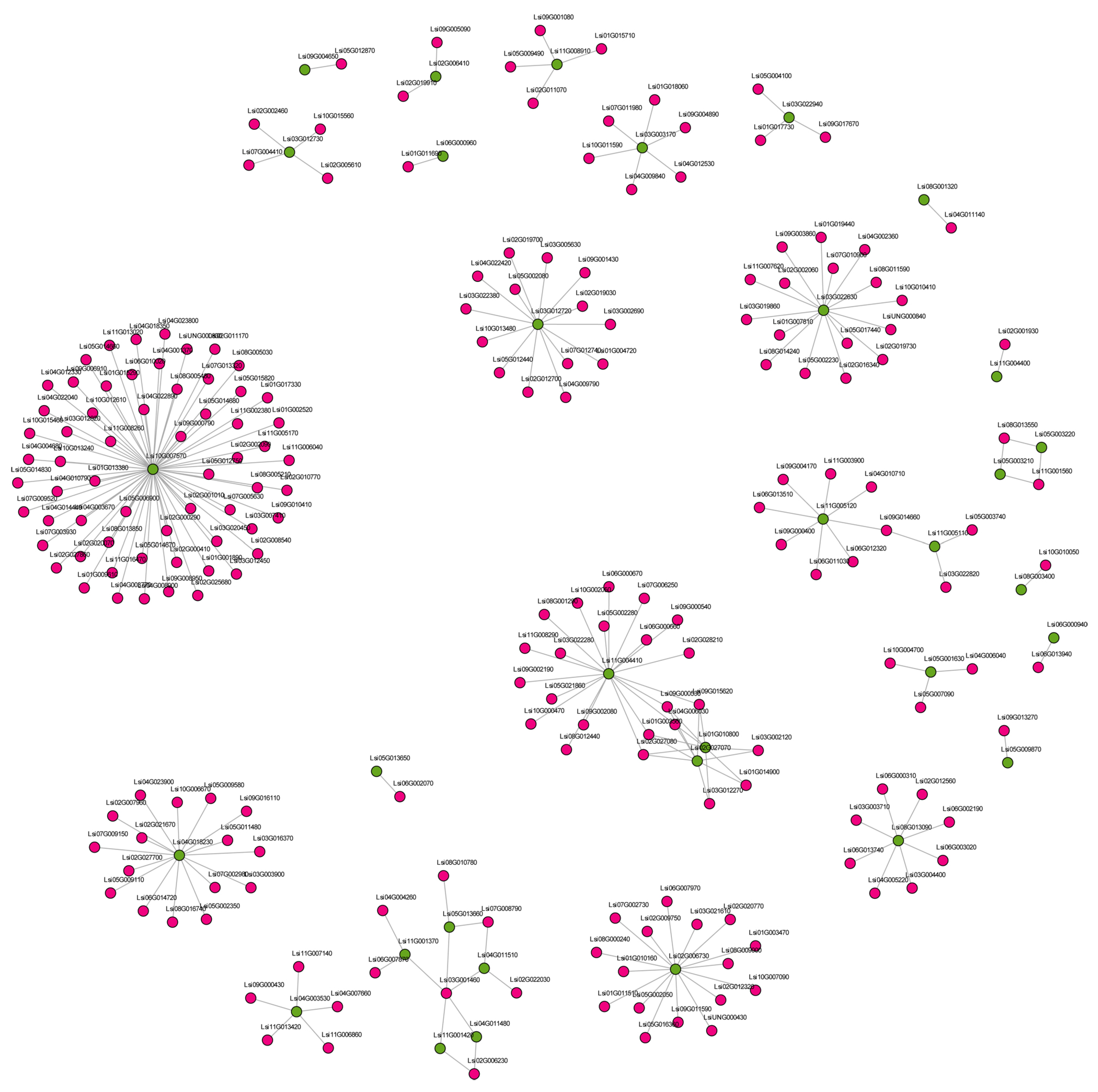
3.7. Regulatory Networks of Disease-Resistance Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Decker-Walters, D.S.; Wilkins-Ellert, M.; Chung, S.M.; Staub, J.E. Discovery and genetic assessment of wild bottle Gourd [Lagenaria siceraria (Mol.) Standley; Cucurbitaceae] from Zimbabwe. Econ. Bot. 2004, 58, 501–508. [Google Scholar] [CrossRef]
- Zahoor, M.; Ikram, M.; Nazir, N.; Naz, S.; Batiha, G.E.S.; Kamran, A.W.; Tomczyk, M.; Kabrah, A. A Comprehensive Review on the Medicinal Importance; Biological and Therapeutic Efficacy of Lagenaria siceraria (Mol.) (Bottle Gourd) Standley Fruit. Curr. Top. Med. Chem. 2021, 21, 1788–1803. [Google Scholar] [CrossRef]
- Chen, X.; Guo, P.; Wang, Z.; Liang, J.; Li, G.; He, W.; Zhen, A. Grafting improves growth and nitrogen-use efficiency by enhancing NO3− uptake, photosynthesis, and gene expression of nitrate transporters and nitrogen metabolizing enzymes in watermelon under reduced nitrogen application. Plant Soil 2022, 480, 305–327. [Google Scholar] [CrossRef]
- Sallaku, G.; Rewald, B.; Sanden, H.; Balliu, A. Scions impact biomass allocation and root enzymatic activity of rootstocks in grafted melon and watermelon plants. Front. Plant Sci. 2022, 13, hal-03793671. [Google Scholar] [CrossRef]
- Li, H.; Gong, S.Q.; Song, H.J.; Chu, X.; Sun, X.W.; Cai, Y.P. Research progress of pumpkin powdery mildew. Chin. Cucurbits Veg. 2021, 34, 8–11. [Google Scholar]
- Qin, J.; Huang, H.; Shang, X.H.; Guo, Y.Y.; Wang, Y.K.; Zhou, S.M. Advances of bottle gourd genetics basic and breeding. Chin. Cucurbits Veg. 2022, 35, 1–7. [Google Scholar]
- Robinson, R.W.; Decker-Walters, D.S. Cucurbits. Q. Rev. Biol. 1997, 73, 213. [Google Scholar]
- Wu, X.; Wang, Y.; Wu, X.; Xu, P.; Wang, B.; Lu, F.Z.; Li, G.J. Genome-wide Association Analysis of Powdery Mildew Resistance in BottleGourd (Lagenaria siceraria). Mol. Plant Breed. 2020, 18, 759–764. [Google Scholar]
- Cui, H.N.; Fan, C.; Ding, Z.; Wang, X.Z.; Tang, L.L.; Bi, Y.D.; Luan, F.S.; Gao, P. CmPMRl and CmPMrs are responsible for resistance to powdery mildew caused by Podosphaera xanthii race 1 in Melon. Theor. Appl. Genet. 2022, 135, 1209–1222. [Google Scholar] [CrossRef]
- Tek, M.I.; Calis, O.; Fidan, H.; Shah, M.D.; Celik, S.; Wani, S.H. CRISPR/Cas9 based mlo-mediated resistance against Podosphaera xanthii in cucumber (Cucumis sativus L.). Front. Plant Sci. 2022, 13, 5234. [Google Scholar] [CrossRef]
- Wang, L.P.; Wu, X.; Wang, B.; Xu, P.; Li, G.J. SCAR marker linked to resistance gene of powdery mildew in bottle gourd [Lagenariasiceraria (Molina) Standl] breeding line J083. J. Zhejiang Univ. 2011, 37, 119–124. [Google Scholar]
- Wang, J.; Yang, C.; Wu, X.; Wang, Y.; Wang, B.; Wu, X.; Lu, Z.; Li, G. Genome-wide characterization of NBS-LRR family genes and expression analysis under powdery mildew stress in Lagenaria siceraria. Physiol. Mol. Plant Pathol. 2022, 118, 101798. [Google Scholar] [CrossRef]
- Zhao, Z.; Dong, Y.; Wang, J.; Zhang, G.; Zhang, Z.; Zhang, A.; Wang, Z.; Ma, P.; Li, Y.; Zhang, X.; et al. Comparative transcriptome analysis of melon (Cucumis melo L.) reveals candidate genes and pathways involved in powdery mildew resistance. Sci. Rep. 2022, 12, 4936. [Google Scholar]
- Cao, Y.Y.; Diao, Q.N.; Chen, Y.Y.; Jin, H.J.; Zhang, Y.P.; Zhang, H.M. Development of KASP Markers and Identification of a QTL Underlying Powdery Mildew Resistance in Melon (Cucumis melo L.) by Bulked Segregant Analysis and RNA-Seq. Front. Plant Sci. 2021, 11, 1819. [Google Scholar]
- Cao, Y.Y.; Diao, Q.N.; Lu, S.J.; Zhang, Y.P.; Yao, D.W. Comparative transcriptomic analysis of powdery mildew resistant and susceptible melon inbred lines to identify the genes involved in the response to Podosphaera xanthii infection. Sci. Hortic. 2022, 304, 111305. [Google Scholar] [CrossRef]
- Sakata, Y.; Kubo, N.; Morishita, M.; Kitadani, E.; Sugiyama, M.; Hirai, M. QTL analysis of powdery mildew resistance in cucumber (Cucumis sativus L.). TAG. Theor. Appl. Genet. Theor. Angew. Genet. 2006, 112, 243–250. [Google Scholar] [CrossRef]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—A Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef]
- Anders, S.; Huber, W. Differential Expression of RNA-Seq Data at the Gene Level—The DESeq Package; European Molecular Biology Laboratory: Heidelberg, Germany, 2013. [Google Scholar]
- Zhang, X.L.; Zhang, J.S.; Lei, B.; Yu, J.; Zhao, D.G. Research Progress in Plant MLO Protein. Plant Physiol. J. 2018, 54, 1159–1171. [Google Scholar]
- Averyanov, A. Oxidative burst and plant disease resistance. Front Biosci. 2009, 1, 142–152. [Google Scholar]
- Zhang, S.; Liu, J.; Xu, B.; Zhou, J. Differential Responses of Cucurbita pepo to Podosphaera xanthii Reveal the Mechanism of Powdery Mildew Disease Resistance in Pumpkin. Front Plant Sci. 2021, 12, 633221. [Google Scholar] [CrossRef]
- Muro-Villanueva, F.; Mao, X.; Chapple, C. Linking phenylpropanoid metabolism, lignin deposition, and plant growth inhibition. Curr. Opin. Biotechnol. 2019, 56, 202–208. [Google Scholar] [CrossRef]
- Polonio, Á.; Pineda, M.; Bautista, R.; Martínez-Cruz, J.; Pérez-Bueno, M.L.; Barón, M.; Pérez-García, A. RNA-seq analysis and fluorescence imaging of melon powdery mildew disease reveal an orchestrated reprogramming of host physiology. Sci. Rep. 2019, 9, 7978. [Google Scholar] [CrossRef]
- Jogaiah, S.; Satapute, P.; De Britto, S.; Konappa, N.; Udayashankar, A.C. Exogenous priming of chitosan induces upregulation of phytohormones and resistance against cucumber powdery mildew disease is correlated with localized biosynthesis of defense enzymes. Int. J. Biol. Macromol. 2020, 162, 1825–1838. [Google Scholar] [CrossRef]
- Chen, B.-H.; Guo, W.-L.; Yang, H.-L.; Li, Q.-F.; Zhou, J.-G.; Li, X.-Z. Photosynthetic properties and biochemical metabolism of Cucurbita moschata genotypes following infection with powdery mildew. J. Plant Pathol. 2020, 102, 1021–1027. [Google Scholar] [CrossRef]
- Miao, L.X.; Jiang, M.; Zhang, Y.C.; Yang, X.F.; Zhang, H.Q.; Zhang, Z.F.; Wang, Y.Z.; Jiang, G.H. Genomic identification, phylogeny, and expression analysis of MLO genes involved in susceptibility to powdery mildew in Fragaria vesca. Genet. Mol. Res. 2016, 15. [Google Scholar] [CrossRef]
- Zhou, M.M. Cloning and Expression Analysis of Powdery Mildew Resistance Relative Genes in Momordica charantia L. Master’s Thesis, Hainan University, Hainan, China, 2019. [Google Scholar]
- Zhu, L.; Li, Y.; Li, J.; Yong, W.; Zhang, Z.; Wang, Y.; Wang, Z.; Hu, J.; Yang, L.; Sun, S. Genome-wide identification and analysis of the MLO gene families in three Cucurbita species. Czech J. Genet. Plant Breed. 2021, 57, 119–123. [Google Scholar] [CrossRef]
- Win, K.T.; Zhang, C.; Lee, S. Genome-wide identification and description of MLO family genes in pumpkin (Cucurbita maxima Duch.). Hortic. Environ. Biotechnol. 2018, 59, 397–410. [Google Scholar] [CrossRef]
- Fujita, K.; Inui, H. Review: Biological functions of major latex-like proteins in plants. Plant Sci. 2021, 306, 110856. [Google Scholar] [CrossRef]
- Kang, Y.; Tong, J.; Liu, W.; Jiang, Z.; Pan, G.; Ning, X.; Yang, X.; Zhong, M. Comprehensive Analysis of Major Latex-Like Protein Family Genes in Cucumber (Cucumis sativus L.) and Their Potential Roles in Phytophthora Blight Resistance. Int. J. Mol. Sci. 2023, 24, 784. [Google Scholar]
- Fujita, K.; Asuke, S.; Isono, E.; Yoshihara, R.; Uno, Y.; Inui, H. MLP-PG1, a major latex-like protein identified in Cucurbita pepo, confers resistance through the induction of pathogenesis-related genes. Planta 2021, 255, 10. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence |
|---|---|
| LsGAPDH | F: CCCAGGGGATATCTGCAGGG |
| R: CATGGTGTTTTCAATGGAACCA | |
| Lsi02G027070 | F: GAGACTCAACGACACAGGCA |
| R: GTGAGGAAGAAGGTGGTCCG | |
| Lsi05G003210 | F: CGGTATCCCACTAAAAGCAAAGC |
| R: ACTGAGCCTTTGGTTCCACC | |
| Lsi05G003220 | F: AGAGGTAAAAGAGAGGCTGGC |
| R: ACTGAGCTGCCATTGCTACC | |
| Lsi05G009870 | F: TTGCCACCAGGTGAAACCAT |
| R: TCAGCCAATGGAAGGGATTGA | |
| Lsi05G013660 | F: TGCTTGAGCTTTGTCACGGT |
| R: TCATGCCTTAGGAGCTTCAACA | |
| Lsi11G001370 | F: TGGAGGTGATTGGGACAACC |
| R: AACACATGTCCATCCCGTCC | |
| Lsi04G011480 | F: ATGTGGAAATTGAGGCTCCAG |
| R: CCCACTTTAGCCTTTCCATAGC | |
| Lsi05G013650 | F: GGGAAGTGTGGTTCACTGGT |
| R: GGTTCATCACGACCTCCTGT | |
| Ls11G001420 | F: AGTCGTGCCAAAGGGTCAAA |
| R: TCGATATCTTTGGTCACACCGA | |
| Ls04G003530 | F: GAGCACCAAACCCCATCTCC |
| R: CAGCCTTCTTGTTTGGACCG | |
| Lsi04G011510 | F: GGGAAAGGGAAGTGTAGCTC |
| R: ATCAAGGTCTCTGCCGACTTC |
| Total | G3 | G6 | ||||||
|---|---|---|---|---|---|---|---|---|
| 12 h | 24 h | 48 h | 72 h | 12 h | 24 h | 48 h | 72 h | |
| cinnamic acid biosynthetic process | √ | √ | √ | √ | √ | √ | √ | √ |
| L-phenylalanine catabolic process | √ | √ | √ | √ | √ | √ | √ | √ |
| alkaloid biosynthetic process | √ | √ | √ | √ | √ | |||
| glutamine metabolic process | √ | √ | ||||||
| response to auxin | √ | √ | √ | √ | √ | |||
| tyrosine metabolic process | √ | √ | √ | √ | ||||
| alanine metabolic process | √ | √ | ||||||
| response to oxidative stress | √ | √ | √ | √ | √ | √ | ||
| regulation of transcription, DNA-templated | √ | √ | √ | √ | √ | √ | ||
| hydrogen peroxide catabolic process | √ | √ | √ | √ | ||||
| cellular oxidant detoxification | √ | √ | √ | √ | √ | √ | ||
| serine family amino acid metabolic process | √ | √ | √ | √ | ||||
| protein phosphorylation | √ | √ | √ | √ | ||||
| cell wall macromolecule catabolic process | √ | √ | √ | |||||
| chitin catabolic process | √ | √ | √ | |||||
| recognition of pollen | √ | √ | √ | |||||
| chloroplast RNA processing | √ | √ | ||||||
| chloroplast RNA modification | √ | √ | ||||||
| starch catabolic process | √ | √ | ||||||
| RNA secondary structure unwinding | √ | √ | ||||||
| maturation of SSU-rRNA | √ | √ | ||||||
| protein refolding | √ | √ | ||||||
| microtubule-based movement | √ | √ | ||||||
| transmembrane transport | √ | √ | √ | √ | ||||
| metabolic process | √ | √ | √ | |||||
| oxidation-reduction process | √ | √ | √ | √ | √ | |||
| transport | √ | √ | √ | |||||
| aromatic compound biosynthetic process | √ | √ | ||||||
| Pathway ID | Pathway Name | ListHits | p Value |
|---|---|---|---|
| ko00250 | Alanine, aspartate, and glutamate metabolism | 25 | 1.53 × 10−5 |
| ko00360 | Phenylalanine metabolism | 20 | 0.000135356 |
| ko04712 | Circadian rhythm–plant | 19 | 0.000224721 |
| ko00410 | Beta-alanine metabolism | 16 | 0.001916155 |
| ko00511 | Other glycan degradation | 10 | 0.002438963 |
| ko00130 | Ubiquinone and other terpenoid–quinone biosynthesis | 17 | 0.00589103 |
| ko04075 | Plant hormone signal transduction | 66 | 0.005954455 |
| ko00062 | Fatty acid elongation | 11 | 0.006594654 |
| ko00920 | Sulfur metabolism | 13 | 0.010577539 |
| ko00940 | Phenylpropanoid biosynthesis | 51 | 0.013346325 |
| Pathway ID | Pathway Name | ListHits | p Value |
|---|---|---|---|
| ko00250 | Alanine, aspartate, and glutamate metabolism | 26 | 2.21 × 10−6 |
| ko03008 | Ribosome biogenesis in eukaryotes | 33 | 3.68 × 10−6 |
| ko00940 | Phenylpropanoid biosynthesis | 56 | 0.000457519 |
| ko04712 | Circadian rhythm–plant | 18 | 0.000470089 |
| ko00360 | Phenylalanine metabolism | 18 | 0.000869833 |
| ko00196 | Photosynthesis–antenna proteins | 11 | 0.515563734 |
| ko00906 | Carotenoid biosynthesis | 12 | 0.955349824 |
| ko00430 | Taurine and hypotaurine metabolism | 5 | 0.363705141 |
| ko03018 | RNA degradation | 24 | 0.369539462 |
| ko00760 | Nicotinate and nicotinamide metabolism | 8 | 0.699410569 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Z.; Wang, K.; Chen, C.; Tian, S.; Wu, J.; Li, J.; Kong, L.; Yang, X.; Zhang, C.; Li, Y.; et al. Transcriptome Sequence Analysis of Defense Response of Resistant and Susceptible Bottle Gourd to Powdery Mildew. Agronomy 2023, 13, 1406. https://doi.org/10.3390/agronomy13051406
Zhang Z, Wang K, Chen C, Tian S, Wu J, Li J, Kong L, Yang X, Zhang C, Li Y, et al. Transcriptome Sequence Analysis of Defense Response of Resistant and Susceptible Bottle Gourd to Powdery Mildew. Agronomy. 2023; 13(5):1406. https://doi.org/10.3390/agronomy13051406
Chicago/Turabian StyleZhang, Zhaohui, Kuanhong Wang, Chunhong Chen, Shoubo Tian, Jue Wu, Jianyong Li, Lingjuan Kong, Xiaofeng Yang, Changwei Zhang, Ying Li, and et al. 2023. "Transcriptome Sequence Analysis of Defense Response of Resistant and Susceptible Bottle Gourd to Powdery Mildew" Agronomy 13, no. 5: 1406. https://doi.org/10.3390/agronomy13051406