
Polyphenolic Acid Changes in Stem Cuttings of Rosa Cultivars in Relation to Phenological Stage and Rooting Enhancers

Abstract
:1. Introduction
2. Materials and Methods
2.1. Stock Plants and Experiment Establishment
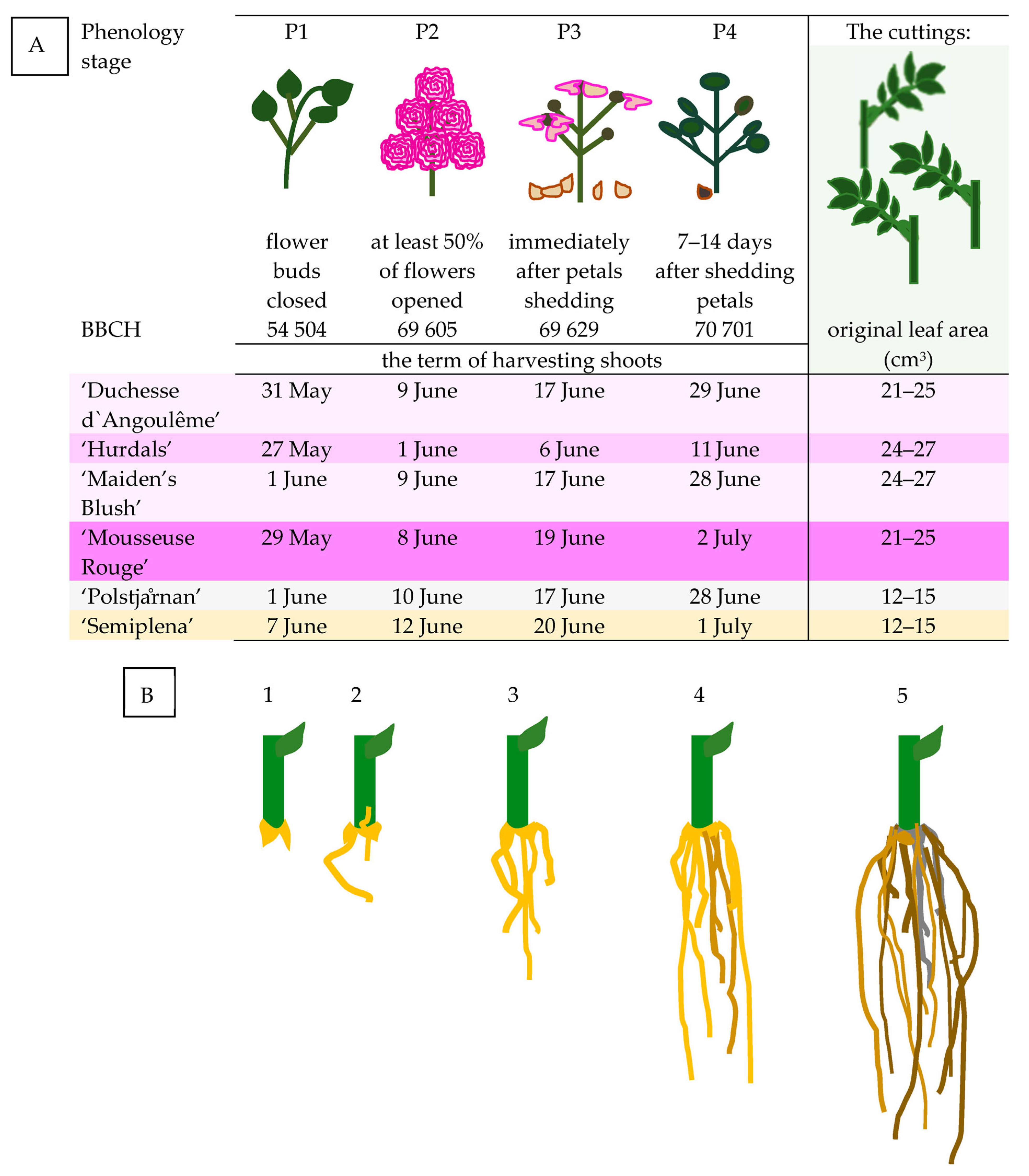
- (A)
- ‘Duchesse d’Angoulême’, Hybrid Gallica (Jean-Pierre Vibert, France 1821);
- (B)
- ‘Hurdals’, Alba/Hybrid Villosa (Germany/Norway; unknown origin);
- (C)
- ‘Maiden’s Blush’, Alba (unknown origin, before 1400);
- (D)
- ‘Mousseuse Rouge’, Moss (unknown origin, before 1842);
- (E)
- Rosa beggeriana ‘Polstjårnan’ (Wasastjarna, Finland 1937) (unknown origin, Valdemar Petersen);
- (F)
- Rosa helenae ‘Semiplena’ (Figure 1).
- P1—54 504, just before of flowering (flower buds closed);
- P2—69 605, full flowering (at least 50% of flowers opened);
- P3—69 629, end of flowering (immediately after petal shedding);
2.2. Assessment of Growth Parameters in Rose Cuttings
2.3. Biochemical Analysis
2.4. Statistical Analysis
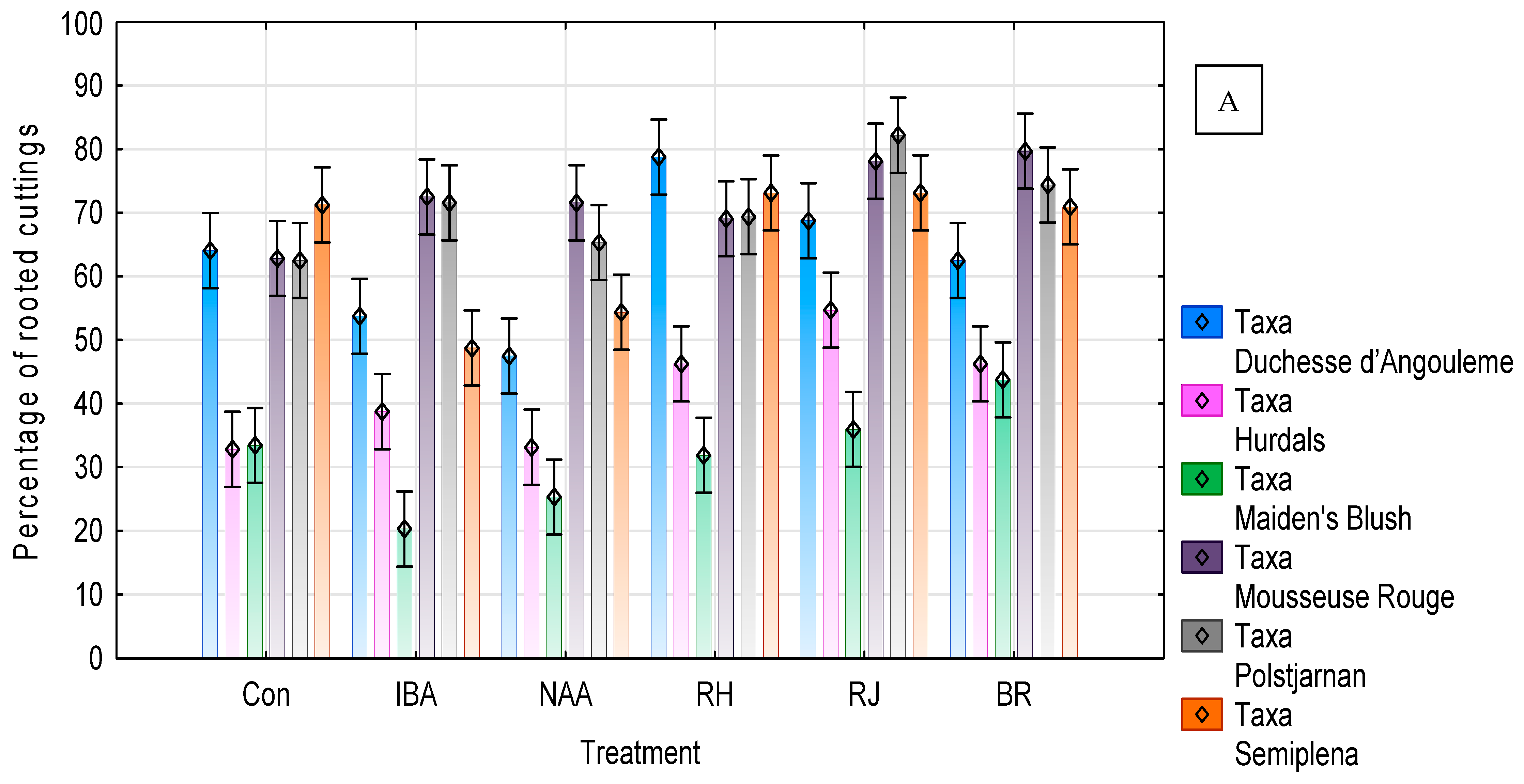
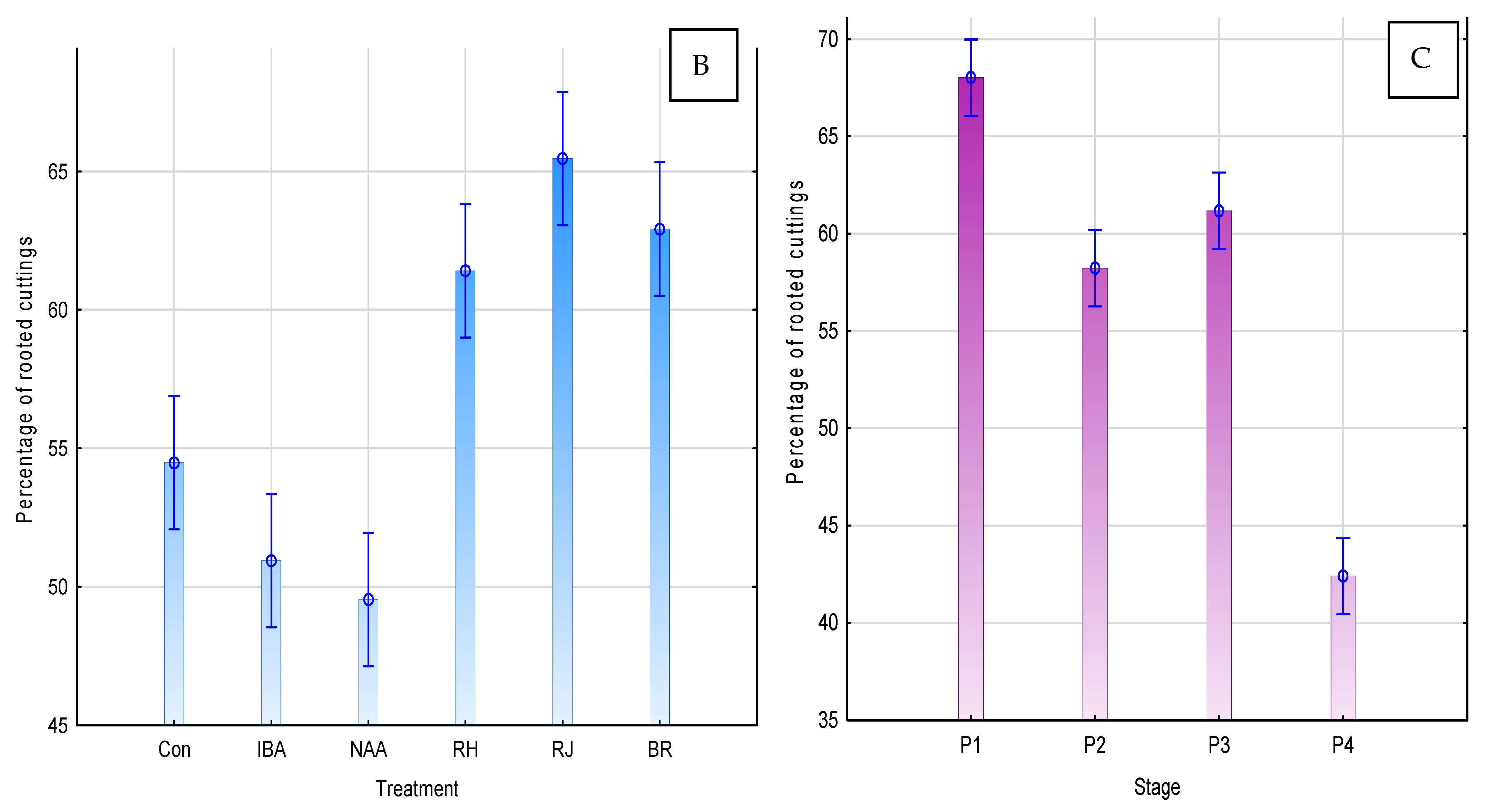
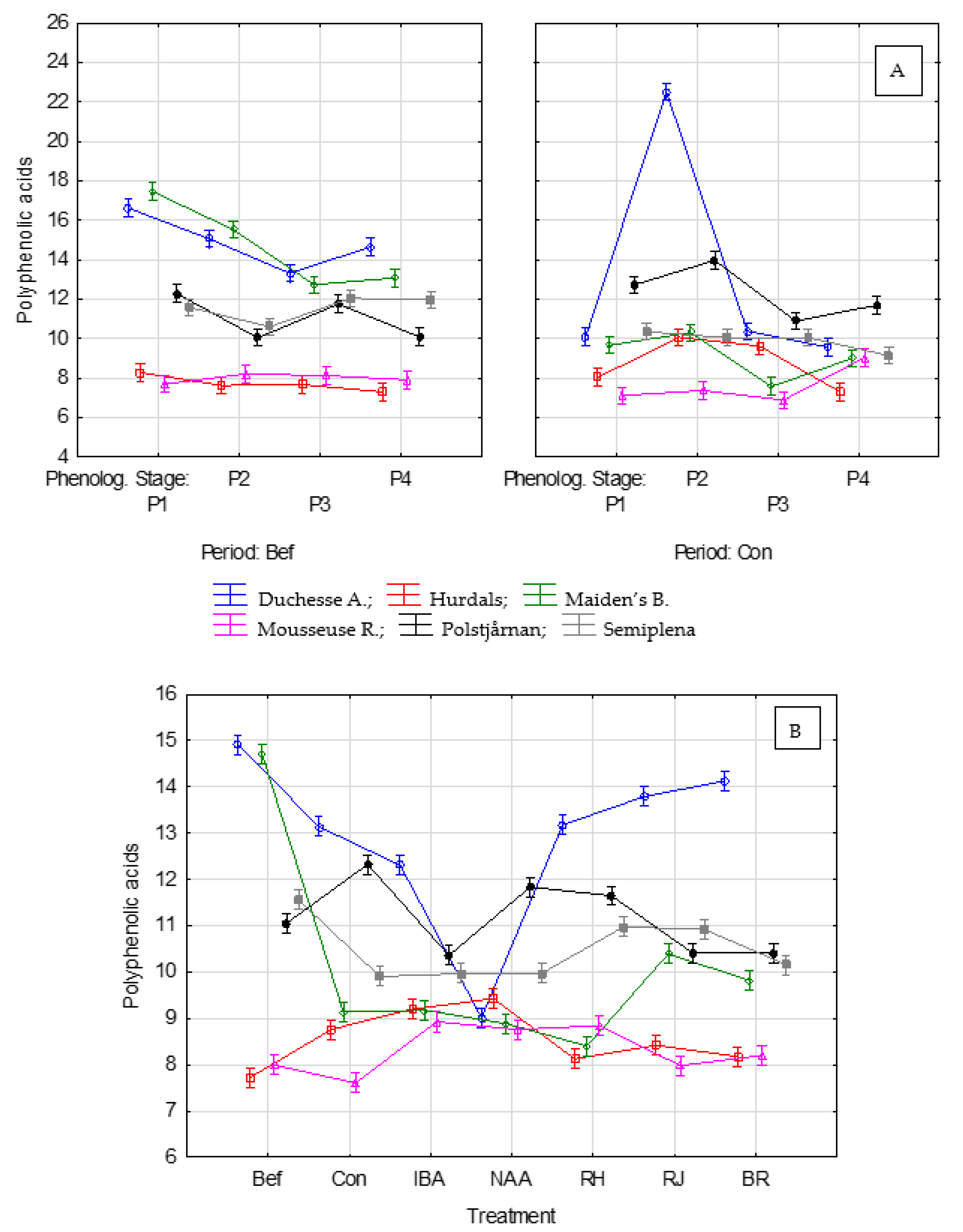
3. Results
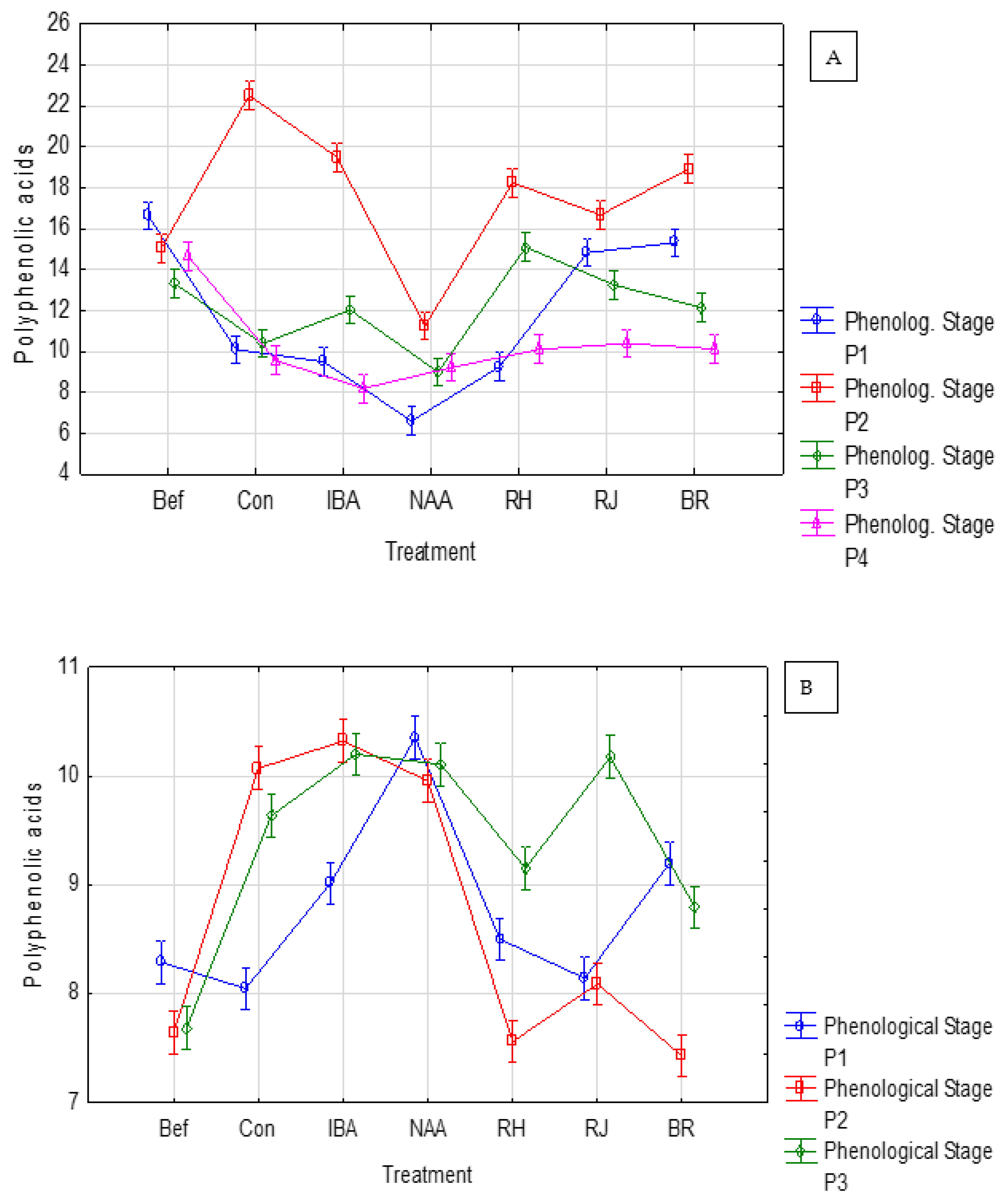
3.1. Rosa ‘Duchesse d’Angoulême’ Response
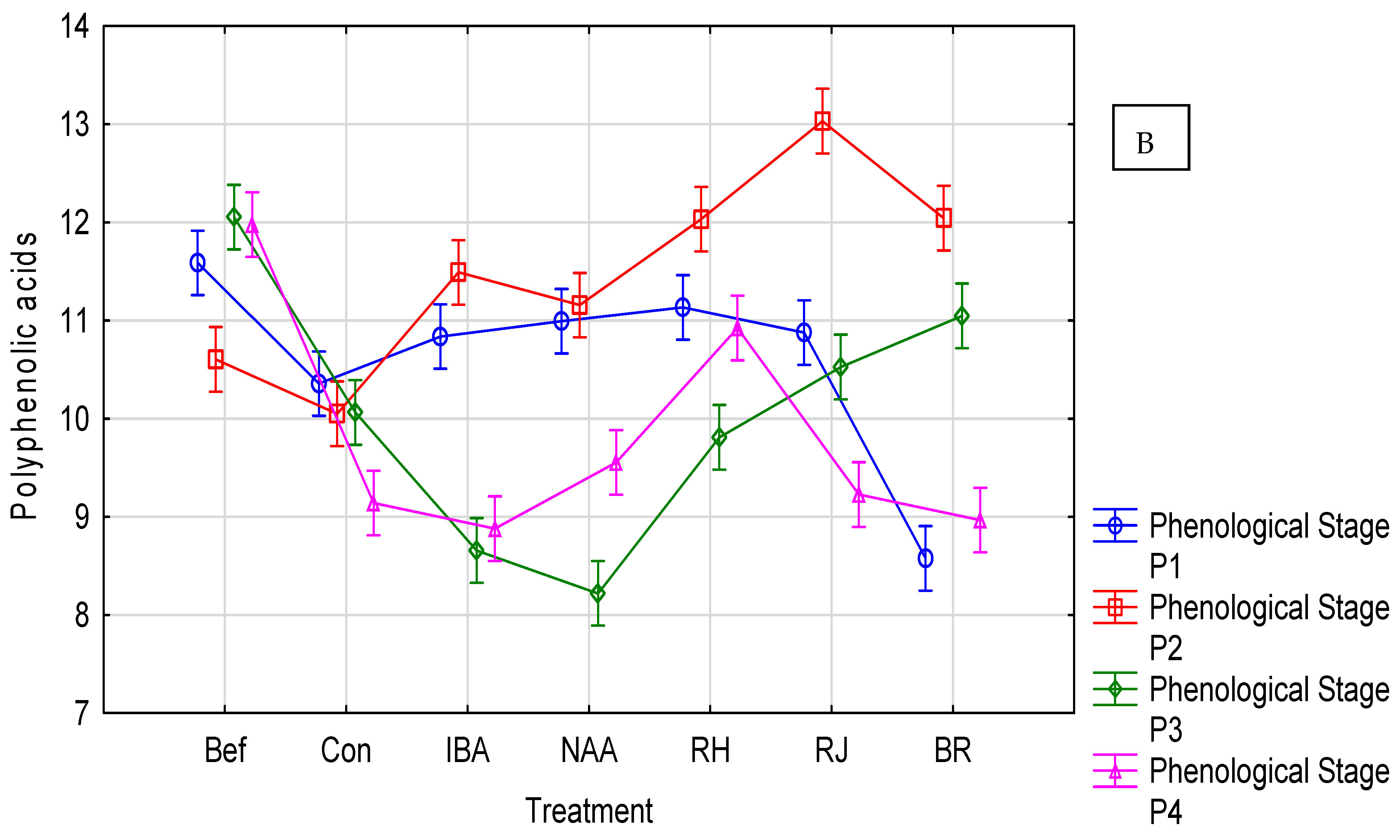
3.2. Rosa ‘Hurdal’ Response
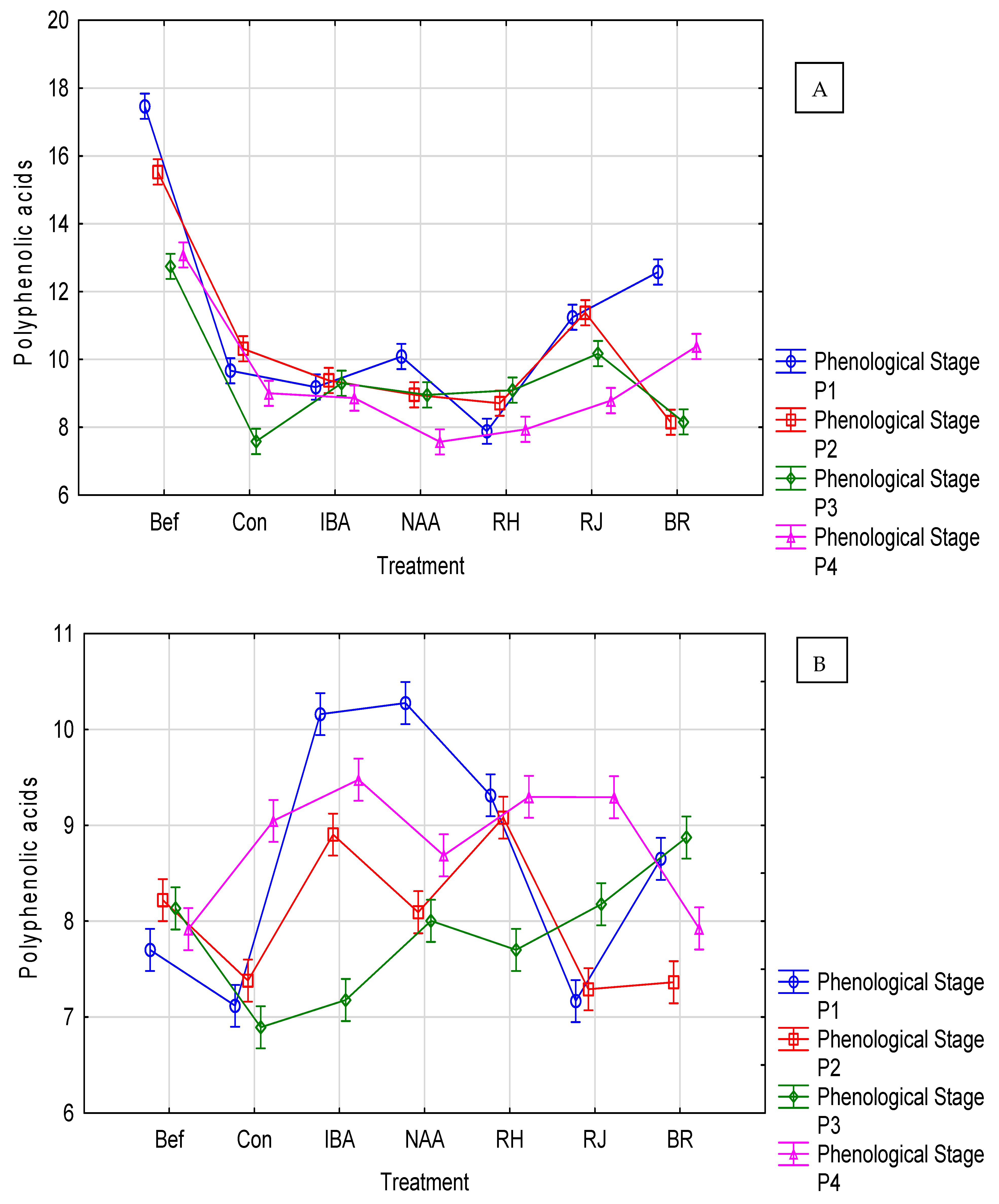
3.3. Rosa ‘Maiden’s Blush’ Response
3.4. Rosa ‘Mousseuse Rouge’ Response
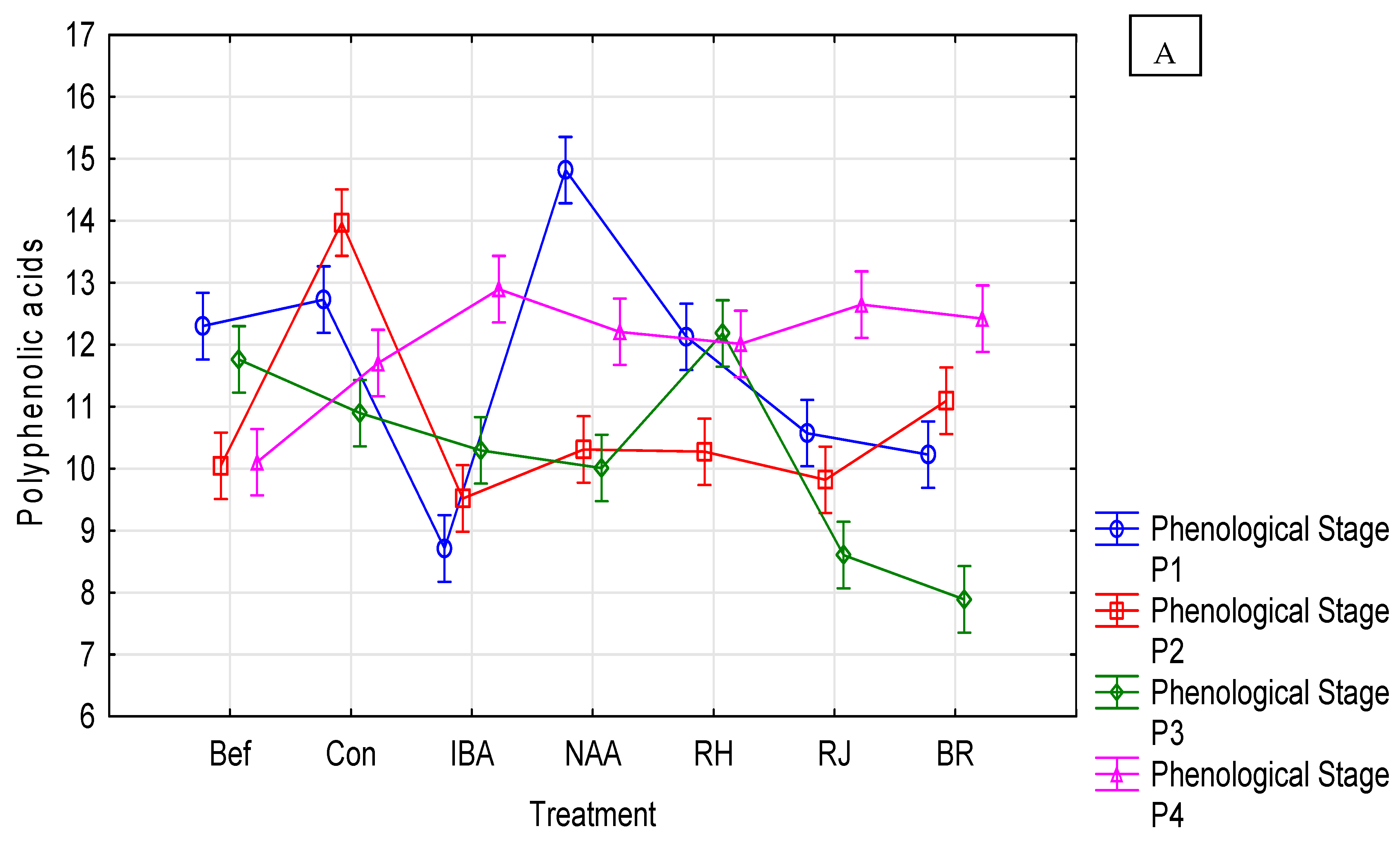
3.5. Rosa beggeriana ‘Polstjårnan’ Response
3.6. Rosa helenae ‘Semiplena’ Response
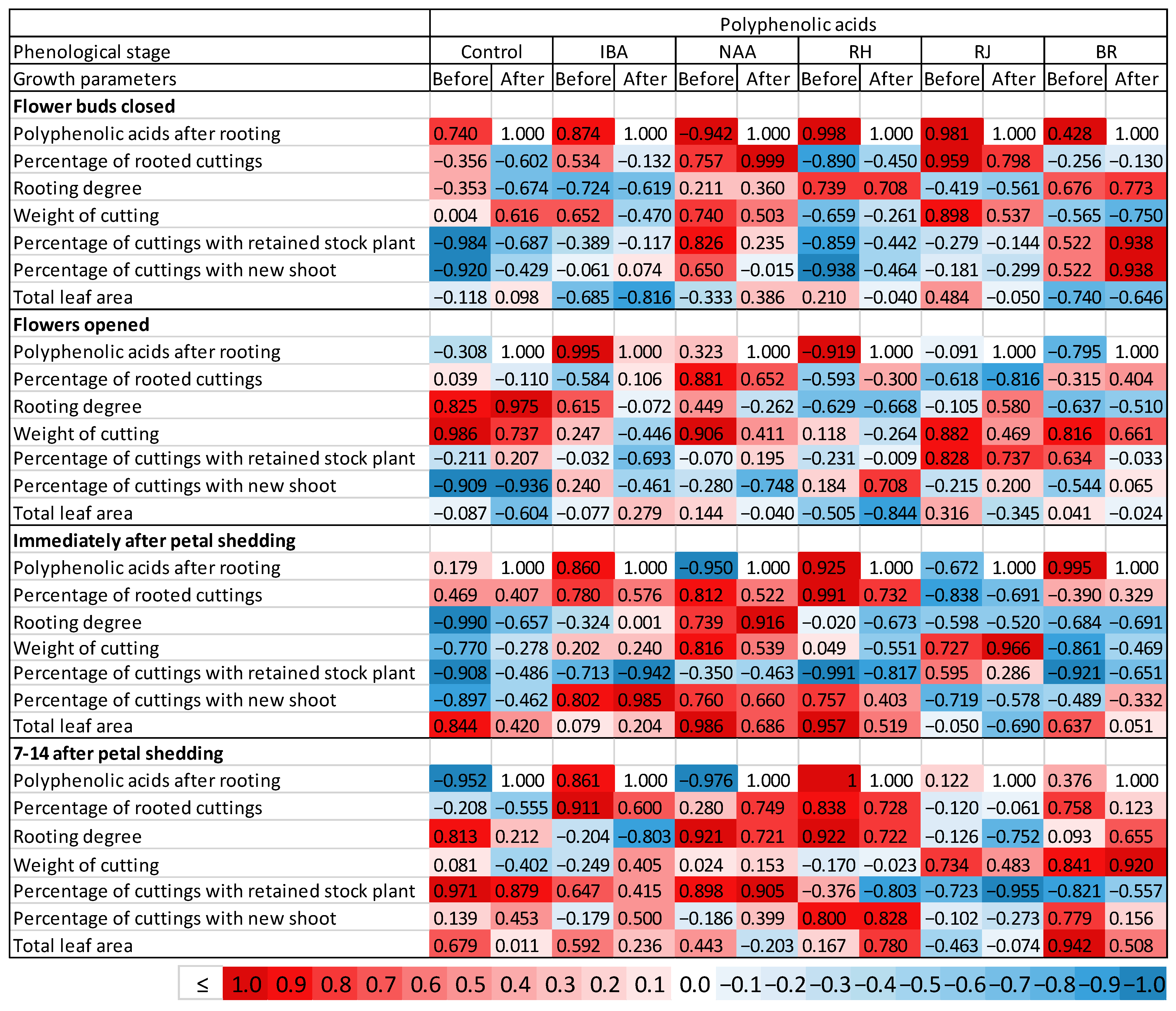
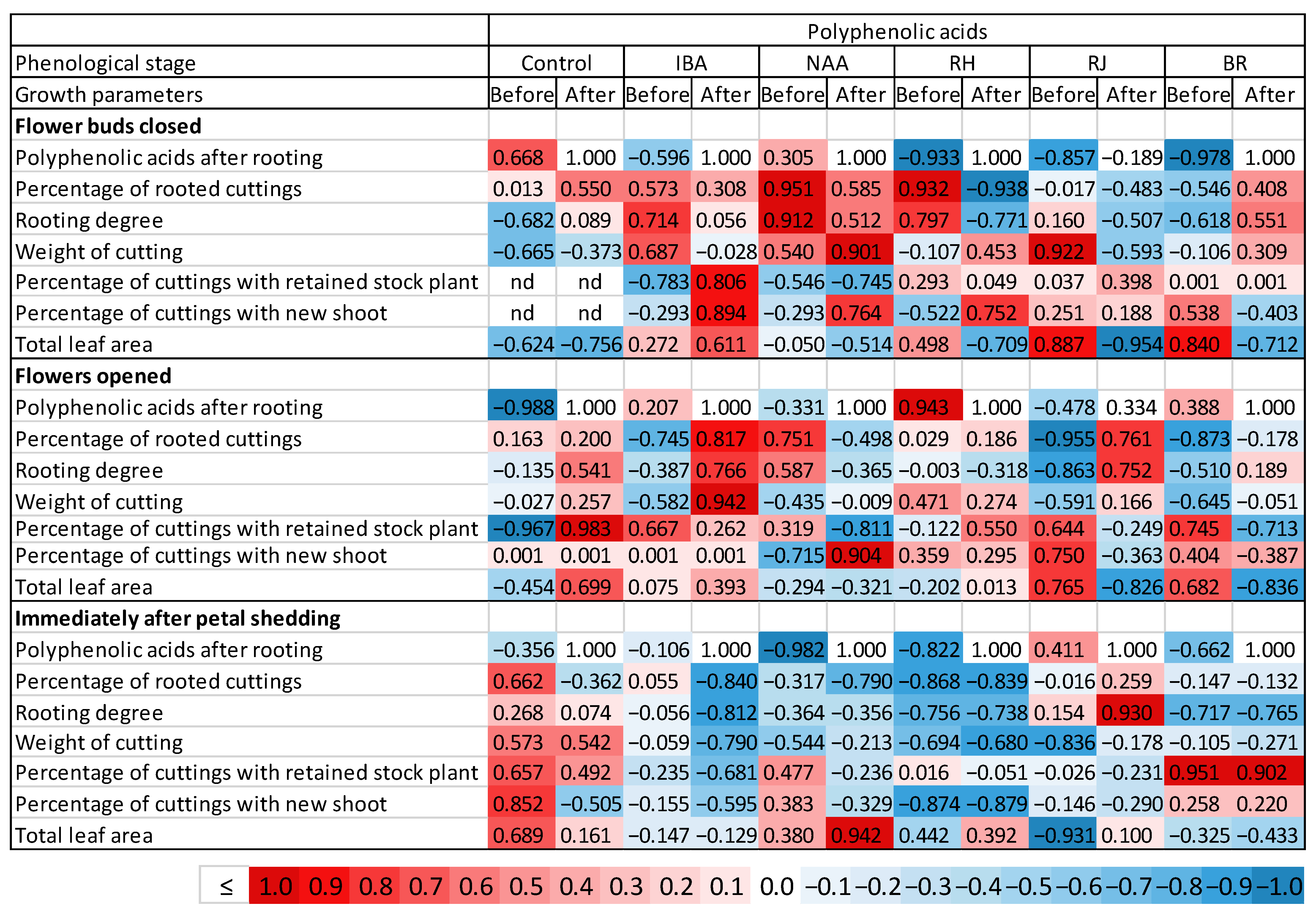
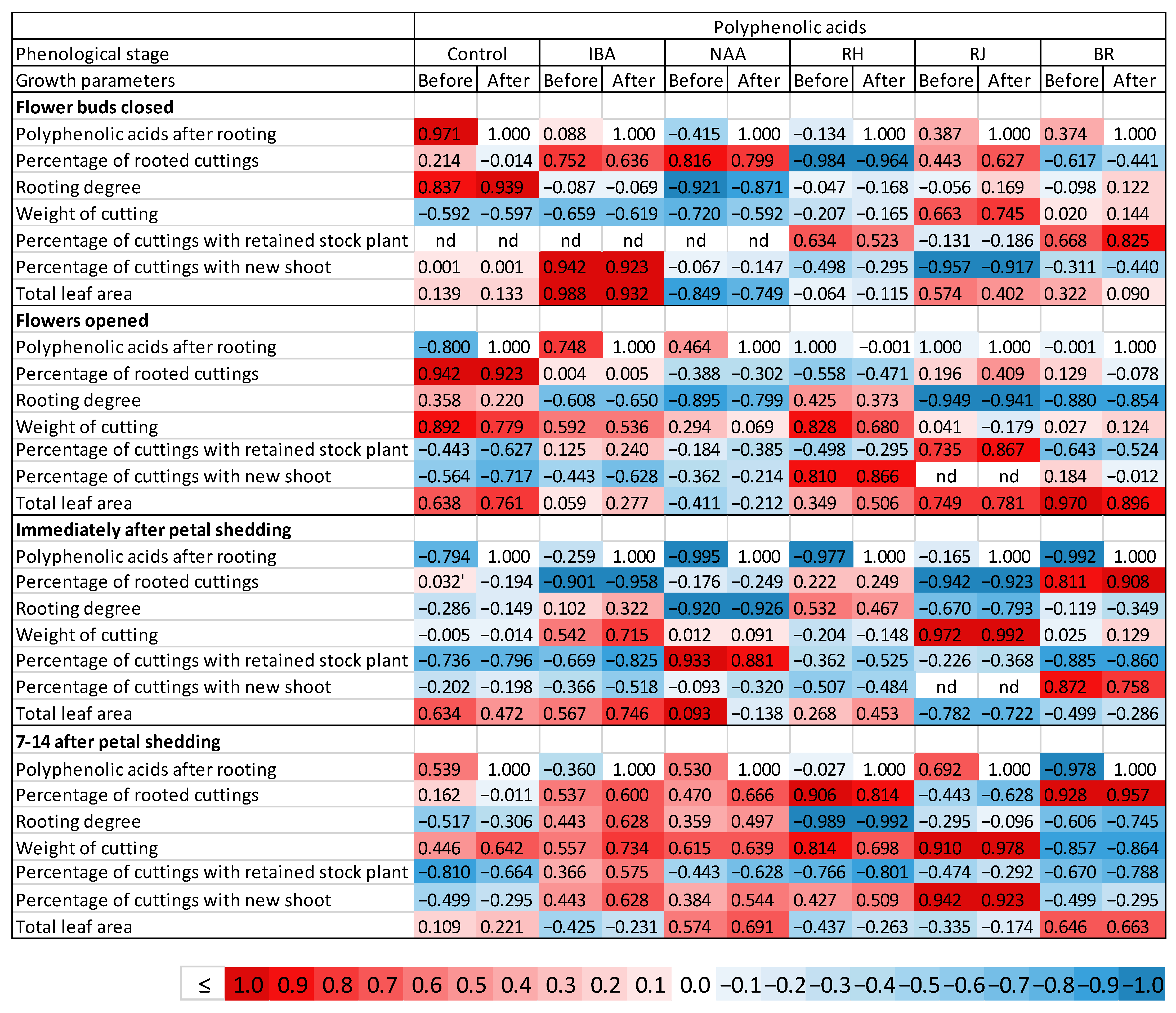
3.7. Correlation between Polyphenolic Acids and the Effectiveness of Rhizogenesis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pivetta, K.F.L.; Pereira, F.M.; Banzatto, D.A.; Grabiano, T.T. Effect of type of cutting and indolbutyric acid on the rooting of rose (Rosa sp. ‘Red Succes’) leaf cuttings during two seasons. Acta Hortic. 1999, 482, 333–338. [Google Scholar] [CrossRef]
- Hoşafçi, H.; Arslan, N.; Sarihan, E.O. Propagation of dog rose (Rosa canina L.) plants by softwood cuttings. Acta Hortic. 2005, 690, 139–142. [Google Scholar] [CrossRef]
- Ercişli, S.; Eşitken, A.; Anapali, O.; Şahin, U. Effects of substrate and IBA-concentration on adventitious root formation on hardwood cuttings of Rosa dumalis. Acta Hortic. 2005, 751, 149–152. [Google Scholar] [CrossRef]
- Cárdenaz-Navarro, R.; López-Perez, L. Vegetative propagation of rose: Effects of substrate, light and leaf persistence. Sci. Agropecu. 2011, 2, 203–211. [Google Scholar] [CrossRef]
- Monder, M.J.; Niedzielski, M.; Woliński, K. The pivotal role of phenological stages enhanced by plant origin preparations in the process of rhizogenesis of Rosa ‘Hurdal’ stem cuttings. Agriculture 2022, 12, 158. [Google Scholar] [CrossRef]
- Pihlajaniemi, H.; Siurainen, M.; Rautio, P.; Laine, K.; Peteri, S.L.; Huttunen, S. Field evaluation of phenology and success of hardy, micropropagated old shrub roses in northern Finland. Acta Agric. Scand. B Soil Plant Sci. 2005, 55, 275–286. [Google Scholar] [CrossRef]
- Monder, M.J.; Niedzielski, M.; Woliński, K.; Pacholczak, A. The impact of seasonal changes in plant tissue on rhizogenesis of stem cuttings of the once flowering roses. Not. Bot. Horti Agrobot. 2016, 44, 92–99. [Google Scholar] [CrossRef]
- Monder, M.J.; Kozakiewicz, P.; Jankowska, A. Effect of anatomical structure of shoots in different flowering phase on rhizogenesis of once-blooming roses. Not. Bot. Horti Agrobot. 2017, 45, 408–416. [Google Scholar] [CrossRef]
- Monder, M.J.; Niedzielski, M.; Woliński, K. Effect of phenological stage and rooting enhancers on physiological parameters in stem cuttings in the process of rhizogenesis of Rosa × alba ‘Maiden’s Blush’. Agriculture 2020, 10, 572. [Google Scholar] [CrossRef]
- Mhimdi, M.; Pérez-Pérez, J.M. Understanding of adventitious root formation: What can we learn from comparative genetics? Front. Plant Sci. 2020, 11, 582020. [Google Scholar] [CrossRef]
- Atkinson, J.A.; Rasmussen, A.; Traini, R.; Voß, U.; Sturrock, C.; Mooney, S.J.; Wells, D.M.; Bennet, M.J. Branching out in roots: Uncovering form, function, and regulation. Plant Physiol. 2014, 166, 538–550. [Google Scholar] [CrossRef] [PubMed]
- Lakehal, A.; Chaabouni, S.; Cavel, E.; Le Hir, R.; Ranjan, A.; Raneshan, Z.; Novák, O.; Păcurar, D.I.; Perrone, I.; Jobert, F.; et al. A molecular framework for the control of adventitious rooting by the TIR1/AFB2-Aux/IAA-dependent auxin signaling in Arabidopsis. Mol. Plant 2019, 12, 1499–1514. [Google Scholar] [CrossRef] [PubMed]
- Costa, C.T.; Almeida, M.R.; Ruedell, C.M.; Schwambach, J.; Maraschin, F.S.; Fett-Neto, A.G. When stress and development go hand in hand: Main hormonal controls of adventitious rooting in cuttings. Front. Plant Sci. 2013, 4, 133. [Google Scholar] [CrossRef]
- Okoro, O.O.; Grace, J. The physiology of rooting Populus cuttings. Physiol. Plant. 1976, 36, 133–138. [Google Scholar] [CrossRef]
- Monder, M.J.; Pacholczak, A. Preparations of plant origin enhance carbohydrate content in plant tissues of rooted cuttings of rambler roses: Rosa beggeriana ‘Polstjärnan’ and Rosa helenae ‘Semiplena’. Acta Agric. Scand. B Soil Plant Sci. 2018, 68, 189–198. [Google Scholar] [CrossRef]
- Monder, M.J.; Pacholczak, A. Rhizogenesis and concentration of carbohydrates in cuttings harvested at different phenological stages of once-blooming rose shrubs and treated with rooting stimulants. Biol. Agric. Hortic. 2020, 36, 55–73. [Google Scholar] [CrossRef]
- Bhattacharya, N.C. Enzyme activities during adventitious rooting. In Adventitious Root Formation in Cuttings; Davis, T.D., Haissig, B.E., Sankhala, N., Eds.; Dioscorides Press: Portland, OR, USA, 1988; pp. 88–101. [Google Scholar]
- Fu, Z.; Xu, P.; He, S.; Teixeira da Silva, J.A.; Tanaka, M. Dynamic changes in enzyme activities and phenolic content during in vitro rooting of tree peony (Paeonia suffruticosa Andr.) plantlets. Intern. J. Sci. Tech. 2011, 5, 252–265. [Google Scholar]
- Šamec, D.; Karalija, E.; Šola, I.; Vujčić Bok, V.; Salopek-Sondi, B. The role of polyphenols in abiotic stress response: The influence of molecular structure. Plants 2021, 10, 118. [Google Scholar] [CrossRef]
- Goleniowski, M.E.; Bonfill, M.; Cusido, R.; Palazon, J. Phenolic acids. In Natural Products; Ramawat, K., Mérillon, J.M., Eds.; Springer Science and Business Media: Berlin/Heidelberg, Germany, 2013; pp. 1951–1973. [Google Scholar]
- Bistgani, Z.E.; Hashemi, M.; Dacosta, M.; Craker, L.; Maggi, F.; Morshedloo, M.R. Effect of salinity stress on the physiological characteristics, phenolic compounds and antioxidant activity of Thymus vulgaris L. and Thymus daenensis Celak. Ind. Crops Prod. 2019, 135, 311–320. [Google Scholar] [CrossRef]
- Zhou, P.; Li, Q.; Liu, G.; Xu, N.; Yang, Y.; Zeng, W.; Chen, A.; Wang, S. Integrated analysis of transcriptomic and metabolomic data reveals critical metabolic pathways involved in polyphenol biosynthesis in Nicotiana tabacum under chilling stress. Funct. Plant Biol. 2018, 46, 30–43. [Google Scholar] [CrossRef]
- Sharma, A.; Thakur, S.; Kumar, V.; Kanwar, M.K.; Kesavan, A.K.; Thukral, A.K.; Bhardwaj, R.; Alam, P.; Ahmad, P. Pre-sowing seed treatment with 24-epibrassinolide ameliorates pesticide stress in Brassica juncea L. through the modulation of stress markers. Front. Plant Sci. 2016, 7, 1569. [Google Scholar] [CrossRef] [PubMed]
- Almeida, L.F.P.; Gomes, S.M.; Vasconcelos, R.G.; Yamanishi, O.K.; Alencar, C.M. Anatomy and histochemistry of araticum (Annona crassiflora Mart.) on three Annonaceae rootstocks. Am. J. Plant Sci. 2014, 5, 2440–2447. [Google Scholar] [CrossRef]
- van Loon, L.C. Systemic induced resistance. In Mechanisms of Resistance to Plant Diseases; Slusarenko, A.J., Fraser, R.S.S., Van Loon, L.C., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2000; pp. 521–574. [Google Scholar]
- Mandal, S.M.; Chakraborty, D.; Dey, S. Phenolic acids act as signaling molecules in plant-microbe symbioses. Plant Signal. Behav. 2010, 5, 359–368. [Google Scholar] [CrossRef] [PubMed]
- Gawrońska, H.; Przybysz, A.; Szalacha, E.; Słowiński, A. Physiological and molecular mode of action of Asahi SL biostimulator under optimal and stress conditions. In Biostimulators in Modern Agriculture: General Aspects; Gawrońska, H., Ed.; Wieś Jutra: Warszawa, Poland, 2008; pp. 54–76. [Google Scholar]
- Günes, T. Peroxidase and IAA-oxidase activities during rooting in cuttings of three poplar species. Turk. J. Bot. 2000, 24, 97–101. [Google Scholar]
- Davis, T.D.; Haissig, B.E.; Sankhla, N. Adventitious root formation in cuttings. In Advances in Plant Sciences Series; Discorides Press: Portland, OR, USA, 1988. [Google Scholar]
- Przybysz, A.; Gawrońska, H.; Gajc-Wolska, J. Biological mode of action of a nitrophenolates-based biostimulant: Case study. Front. Plant Sci. 2014, 5, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Michalak, A. Phenolic compounds and their antioxidant activity in plants growing under heavy metal stress. Pol. J. Environ. Stud. 2006, 15, 523–530. [Google Scholar]
- Mandal, S.M.; Mandal, M.; Das, A.K.; Pati, B.R.; Ghosh, A.K. Stimulation of indoleacetic acid production in a Rhizobium isolate of Vigna mungo by root nodule phenolic acids. Arch. Microbiol. 2009, 191, 389–393. [Google Scholar] [CrossRef]
- Hartmann, H.T.; Kester, D.E.; Davies, F.T.; Geneve, R.L. Plant Propagation, Principles and Practices; Prentice-Hall: Englewood Cliffs, NJ, USA, 2011. [Google Scholar]
- European Union. Directive 2009/128/EC of the European Parliament and of the Council of 21 October 2009 Establishing a Framework for Community Action to Achieve the Sustainable Use of Pesticides. Off. J. Eur. Union 2009, L309, 71–86. [Google Scholar]
- European Union. Regulation (EC) No 1107/2009 of the European Parliament and of the Council of 21 October 2009 Concerning the Placing of Plant Protection Products on the Market and Repealing Council Directives 79/117/EEC and 91/414/EEC. Off. J. Eur. Union 2009, L309, 1–50. [Google Scholar]
- USDA. United States Department of Agriculture. National Organic Program. 2017. Available online: https://www.ams.usda.gov/about-ams/programs-offices/national-organic-program (accessed on 16 March 2017).
- OMRI. Organic Materials Review Institute. OMRI Products List, Web Edition. Organic Materials Review Institute. Available online: https://www.omri.org/omri-lists (accessed on 28 July 2021).
- Rathore, S.S.; Chaudhary, D.R.; Boricha, G.N.; Ghosh, A.; Bhatt, B.P.; Zadope, S.T.; Patolia, J.S. Effect of seaweed extract on growth, yield and nutrient uptake of soybean (Glycine max) under rainfed conditions. S. Afr. J. Bot. 2009, 75, 351–355. [Google Scholar] [CrossRef]
- Pacholczak, A.; Szydło, W.; Petelewicz, P.; Szulczyk, K. The effect if Algaminoplant on rhizogenesis in stem cuttings of Physocarpus opulifolius ‘Dart’s Gold’ and ‘Red Baron’. Acta Sci. Pol. Hortorum Cultus 2013, 12, 105–116. [Google Scholar]
- Monder, M.J.; Kozakiewicz, P.; Jankowska, A. The role of plant origin preparations and phenological stage in anatomy structure changes in the rhizogenesis of Rosa ‘Hurdal’. Front. Plant Sci. 2021, 12, 696998. [Google Scholar] [CrossRef] [PubMed]
- Monder, M.J.; Pacholczak, A. Rhizogenesis and contents of polyphenolic acids in cuttings of old rose cultivars treated with rooting stimulants. Acta Hortic. 2019, 1232, 99–104. [Google Scholar] [CrossRef]
- Canna Continental. Bio Rhizotonic. 2021. Available online: https://www.canna.ca/biorhizotonic (accessed on 28 July 2021).
- BioBizz Worldwide, B.V. Root JuiceTM. 2017. Available online: http://www.biobizz.com/products/#root%c2%b7juice (accessed on 19 January 2017).
- T.A. Terra Aquatica. Pro Roots. Homepage General Hydroponics Europe/T.A. Terra Aquatica. 2021. Available online: https://www.eurohydro.com/proroots (accessed on 1 July 2021).
- Certificaat Bio Roots. No: C8008445INP-01.2013. 2014. Available online: /http://www.eurohydro.com/pdf/certif-bioboosters.pdf/ (accessed on 19 January 2017).
- Nardi, S.; Pizzeghello, D.; Schaivon, M.; Ertani, A. Plant biostimulants: Physiological responses induced by protein hydrolyzed-based products and humic substances in plant metabolism. Sci. Agric. 2015, 73, 18–23. [Google Scholar] [CrossRef]
- Gustavsson, L.A. Rosen Leksikon; Rosinante Forlag A/S.: Kopenhaga, Denmark, 1999. [Google Scholar]
- Ginova, A.; Tsvetkov, I.; Kondakova, V. Rosa damascena Mill.—An overview for evaluation of propagation methods. Bulg. J. Agric. Sci. 2012, 18, 514–556. [Google Scholar]
- Monder, M.J.; Niedzielski, M.; Woliński, K. Effect of rooting preparations on protein, chlorophyll and carotenoid content in leaves of Rosa gallica ‘Duchesse d`Angoulême’ cuttings. Dendrobiology 2014, 72, 29–40. [Google Scholar] [CrossRef]
- Winkelmann, T.; Spethmann, W.; Seegert, A. Propagation of 285-year-old alley linden (Tilia × vulgaris) trees via long cuttings. Eur. J. Hortic. Sci. 2020, 85, 160–168. [Google Scholar] [CrossRef]
- Monder, M.J.; Kozakiewicz, P.; Jankowska, A. Anatomical structure changes in stem cuttings of rambler roses induced with plant origin preparations. Sci. Hortic. 2019, 255, 242–254. [Google Scholar] [CrossRef]
- Druege, U.; Hilo, A.; Pérez-Pérez, J.M.; Klopotek, Y.; Acosta, M.; Shahinnia, F.; Zerche, S.; Franken, P.; Hajirezaei, M.R. Molecular and physiological control of adventitious rooting in cuttings: Phytohormone action meets resource allocation. Ann. Bot. 2019, 123, 929–949. [Google Scholar] [CrossRef]
- Quest-Ritson, C.; Quest-Ritson, B. The Royal Horticultural Society Encyclopedia of Roses; Penguin Company: London, UK, 2013. [Google Scholar]
- Rosenverzeichnis Rosarium Sangerhausen; 3Auflage: Sangerhausen, Germany, 1988.
- Meier, U.; Bleiholder, H.; Brumme, H.; Bruns, E.; Mehring, B.; Proll, T.; Wiegand, J. Phenological growth stages of roses (Rosa sp.): Codification and description according to the BBCH scale. Ann. Appl. Biol. 2008, 154, 231–238. [Google Scholar] [CrossRef]
- (PTF) Polskie Towarzystwo Farmakologiczne. Farmakopea Polska (Polish Pharmacopoeia V); PTF: Warsaw, Poland, 1999; Volume 5, pp. 445–446. [Google Scholar]
- Wójcik, A.R.; Laudański, Z. Planowanie i Wnioskowanie Statystyczne w Doświadczalnictwie. (Statistical Planning and Concluding in Experimental Works); PWN: Warsaw, Poland, 1989; p. 130. [Google Scholar]
- Denaxa, N.-K.; Tsafouros, A.; Roussos, P.A. Chapter 11—Role of phenolic compounds in adventitious root formation. In Plant Biology, Sustainability and Climate Change, Environmental, Physiological and Chemical Controls of Adventitious Rooting in Cuttings; Husen, A., Ed.; Academic Press: Cambridge, MA, USA, 2022; pp. 251–288. [Google Scholar] [CrossRef]
- Das, S.; Sultana, K.W.; Chandra, I. Adventitious rhizogenesis in Basilicum polystachyon (L.) Moench callus and HPLC analysis of phenolic acids. Acta Physiol. Plant. 2021, 43, 146. [Google Scholar] [CrossRef]
- Kumari, A.; Gogna, M.; Mehta, S.; Husen, A. Chapter 18—Adventitious root formation in cuttings and effects of maturation. In Plant Biology, Sustainability and Climate Change, Environmental, Physiological and Chemical Controls of Adventitious Rooting in Cuttings; Husen, A., Ed.; Academic Press: Cambridge, MA, USA, 2022; pp. 397–420. [Google Scholar] [CrossRef]
- Pacholczak, A.; Nowakowska, K.; Pietkiewicz, S. The effects of synthetic auxin and a seaweed-based biostimulator on physiological aspects of rhizogenesis in ninebark stem cuttings. Not. Bot. Horti Agrobot. 2016, 44, 85–91. [Google Scholar] [CrossRef]
- Pacholczak, A.; Szydło, W.; Pijus, J. Effect of Asahi SL preparation on rhizogenesis in stem cuttings of Physocarpus opulifolius ‘Dart’s Gold’ and ‘Diabolo’. Ann. WULS Hortic. Landsc. Archit. 2010, 31, 11–17. [Google Scholar]
- Schuetz, M.; Benske, A.; Smith, R.A.; Watanabe, Y.; Tobimatsu, Y.; Ralph, J.; Demura, T.; Ellis, B.; Samuels, A.L. Laccases direct lignification in the discrete secondary cell wall domains of protoxylem. Plant Physiol. 2014, 166, 798–807. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Luo, L.; Zheng, L. Lignins: Biosynthesis and biological functions in plants. Int. J. Mol. Sci. 2018, 19, 335. [Google Scholar] [CrossRef] [PubMed]
- Osterc, G.; Štefančič, A.; Solar, A.; Štampar, F. Potential involvement of flavonoids in the rooting response of chestnut hybrid (Castanea crenata × Castanea sativa) clones. Aust. J. Exp. Agric. 2007, 47, 96–102. [Google Scholar] [CrossRef]
- Pacholczak, A.; Petelewicz, P.; Jagiełło-Kubiec, K.; Ilczuk, A. The effect of two biopreparations on rhizogenesis in stem cuttings of Cotinus coggygria Scop. Eur. J. Hortic. Sci. 2015, 80, 183–189. [Google Scholar] [CrossRef]
- Cwalina-Ambroziak, B.; Amarowicz, R. Effects of biological and fungicidal environmental protection on chemical composition of tomato and red pepper fruits. Pol. J. Environ. Stud. 2012, 21, 831–836. [Google Scholar]
- Pacholczak, A.; Szydło, W.; Zagórska, K.; Petelewicz, P. The effect of biopreparations on the rooting of stem cuttings in Cotinus coggygria ‘Young Lady’. Ann. WULS 2012, 33, 33–41. [Google Scholar]
- Rosłon, W.; Osińska, E.; Bączek, K.; Węglarz, Z. The influence of organic-mineral fertilizers on field and raw materials quality of chosen plants of the Lamiaceae family from organic cultivation. Acta Sci. Pol. Hortorum Cultus 2011, 10, 147–158. [Google Scholar]
- Akila, N.; Jeyadoss, T. The potential of seaweed liquid fertilizer on the growth and antioxidant enhancement of Helianthus annuus L. Orient. J. Chem. 2010, 26, 1353–1360. [Google Scholar]
- Osterc, G.; Štampar, F. Initial cutting length modifies polyphenol profile in Castanea cuttings during the root formation process. Eur. J. Hortic. Sci. 2008, 73, 201–204. [Google Scholar]
- Khan, W.; Rayrath, U.P.; Subramanian, S.; Jithesh, M.N.; Rayorath, P.; Hodges, D.M.; Critchley, A.T.; Craigie, J.S.; Norrie, J.; Prithiviraj, B. Seaweed extracts as biostimulants of plant growth and development. J. Plant Growth Regul. 2009, 28, 386–399. [Google Scholar] [CrossRef]
- Crouch, I.J.; Van Staden, J. Evidence for the presence of plant growth regulators in commercial seaweed products. Plant Growth Regul. 1993, 13, 21–29. [Google Scholar] [CrossRef]
- Thorsen, M.K.; Woodward, M.; McKenzie, B.M. Kelp (Laminaria digitata) increases germination and affects rooting and plant vigour in crops and native plants from an arable grassland in the Outer Hebrides, Scotland. J. Coast. Conserv. 2010, 14, 239–247. [Google Scholar] [CrossRef]
- Stirk, W.A.; Van Staden, J. Comparison of cytokinin- and auxin-like activity in some commercially used seaweed extracts. J. Appl. Phycol. 1997, 8, 503–508. [Google Scholar] [CrossRef]
- Deolu-Ajayi, A.O.; van der Meer, I.M.; van der Werf, A.; Karlova, R. The power of seaweeds as plant biostimulants to boost cropproduction under abiotic stress. Plant Cell Environ. 2022, 45, 2537–2553. [Google Scholar] [CrossRef]

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
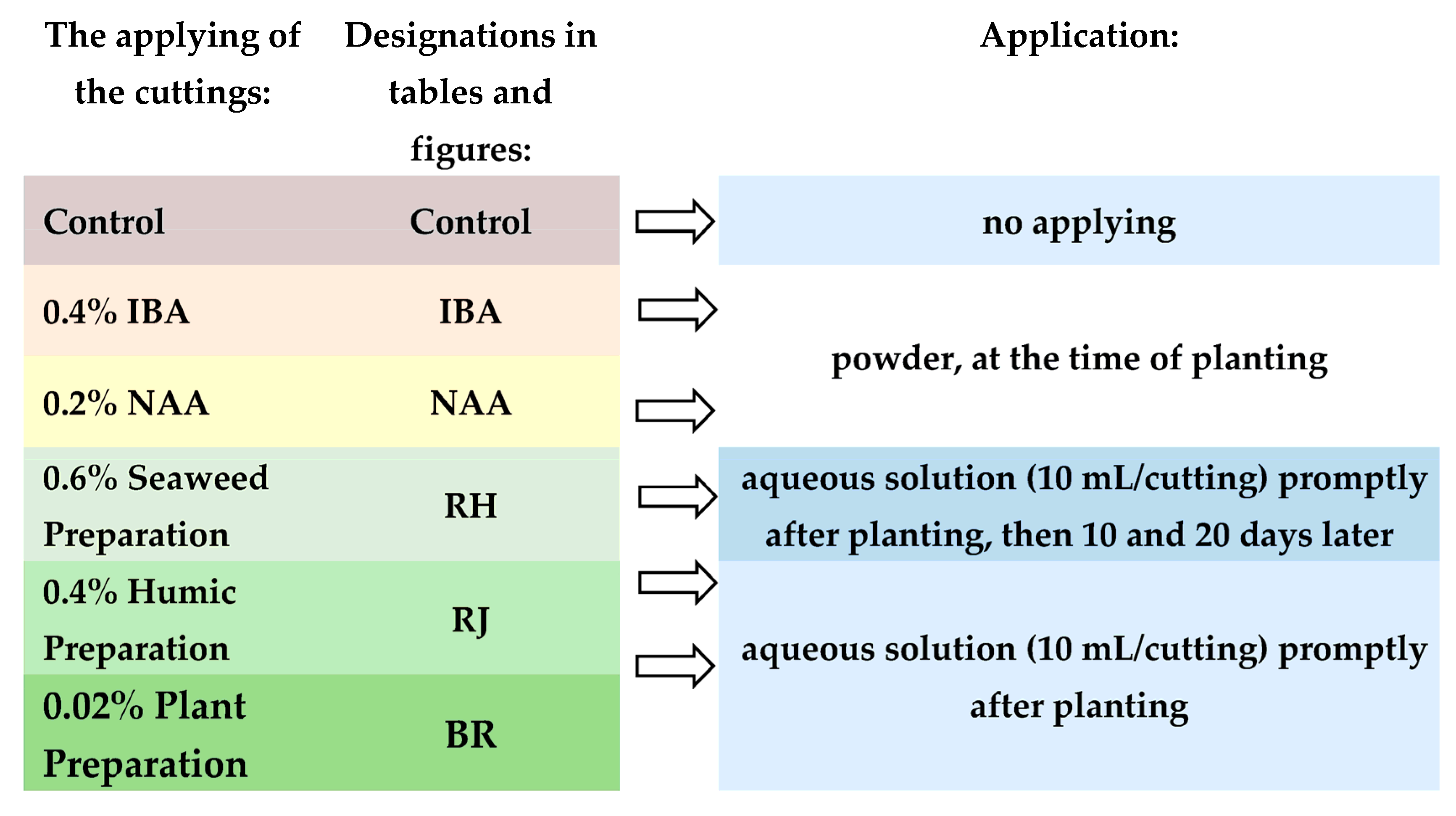
| Name in Work | Trade Name | Contentc | Notification |
|---|---|---|---|
| IBA | Ukorzeniacz Aaqua (Himal, Poland) | 0.4% indolebutyric acid (IBA) | Commercial rooting powder |
| NAA | Ukorzeniacz Baqua (Himal, Poland) | 0.2% naphthalene acetic acid (NAA) | Commercial rooting powder |
| Seaweed Preparation | Bio Rhizotonic [42] | organic matter, seaweed extracts (N 0.6%, P 0.2%, K 0.6% vitamins e.g., B1, B2; other natural active components) | Organic Materials Review Institute [37] |
| Humic Preparation | Root JuiceTM [43] | humic acids and seaweed extracts (N 0.1%, P2O5 0.1%, K2O 0.1%, Mg 0.03%, Fe 0.013%, Mn 0.002%, Zn 0.004%, B 0.025%, Cu 0.001%) | Organic Materials Review Institute [37], National Organic Program (NOP); Control Union Certified EU; Good Soil Quality Mark; Point Vert; Clean Green Certified |
| Plant Preparation | Bio Roots [44] | organic matter 84%, seaweed species extracts 10%, fruit oil up to 1%; humic acids 1%; pectinate 1%; sodium alginate 3%; amino acids, oligosaccharins | Regulation EC No 834/2007 on organic agriculture. Certificaat Bio Roots. No: C8008445INP-01.2013.2 [45] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Monder, M.J.; Pacholczak, A. Polyphenolic Acid Changes in Stem Cuttings of Rosa Cultivars in Relation to Phenological Stage and Rooting Enhancers. Agronomy 2023, 13, 1405. https://doi.org/10.3390/agronomy13051405
Monder MJ, Pacholczak A. Polyphenolic Acid Changes in Stem Cuttings of Rosa Cultivars in Relation to Phenological Stage and Rooting Enhancers. Agronomy. 2023; 13(5):1405. https://doi.org/10.3390/agronomy13051405
Chicago/Turabian StyleMonder, Marta Joanna, and Andrzej Pacholczak. 2023. "Polyphenolic Acid Changes in Stem Cuttings of Rosa Cultivars in Relation to Phenological Stage and Rooting Enhancers" Agronomy 13, no. 5: 1405. https://doi.org/10.3390/agronomy13051405