
LANZA® Tedera Is Strongly Suppressed by Competition from Lolium multiflorum and Is Best Adapted to Light-Textured Soils

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Experimental Design
2.3. Phenology
2.4. Dry Mass, Morphological Traits, and Leaf Nutrients Content
2.5. Physiological Measurements
2.6. Statistical Analysis
3. Results
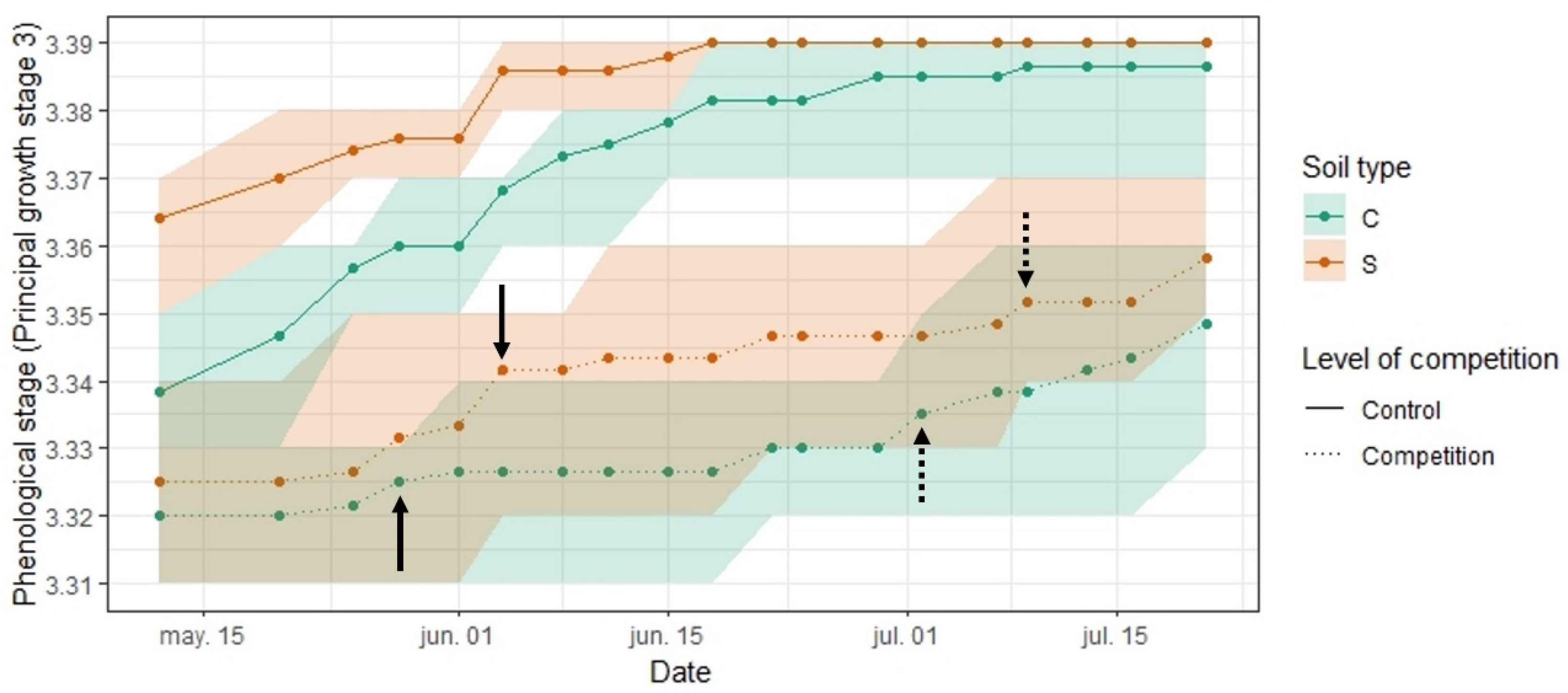
3.1. Effect of Competition and Soil Type on the Development of LANZA®
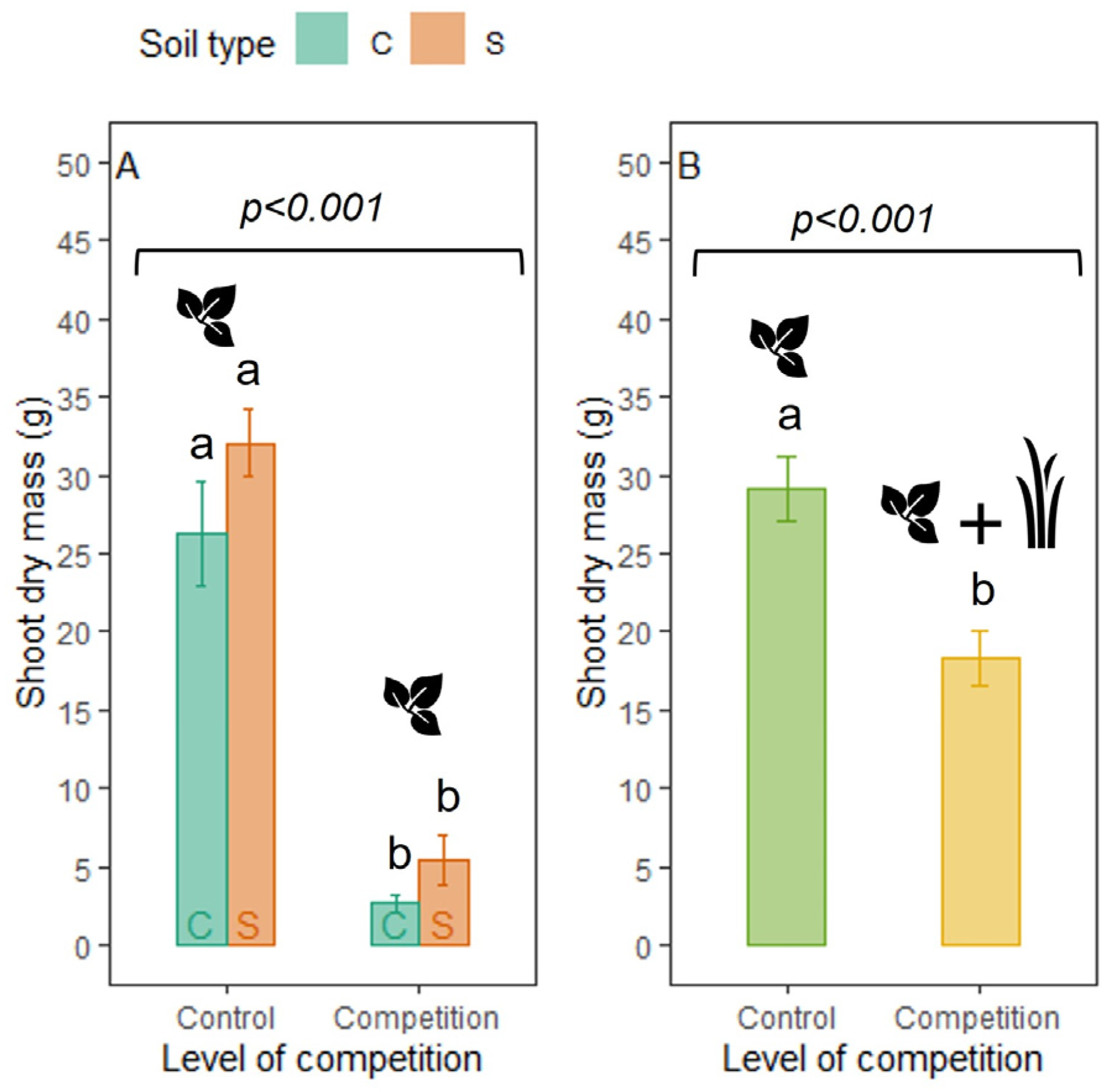
3.2. Effects of Competition and Soil Type on Dry Mass and Morphological Traits
3.3. Leaf Nutrients Content
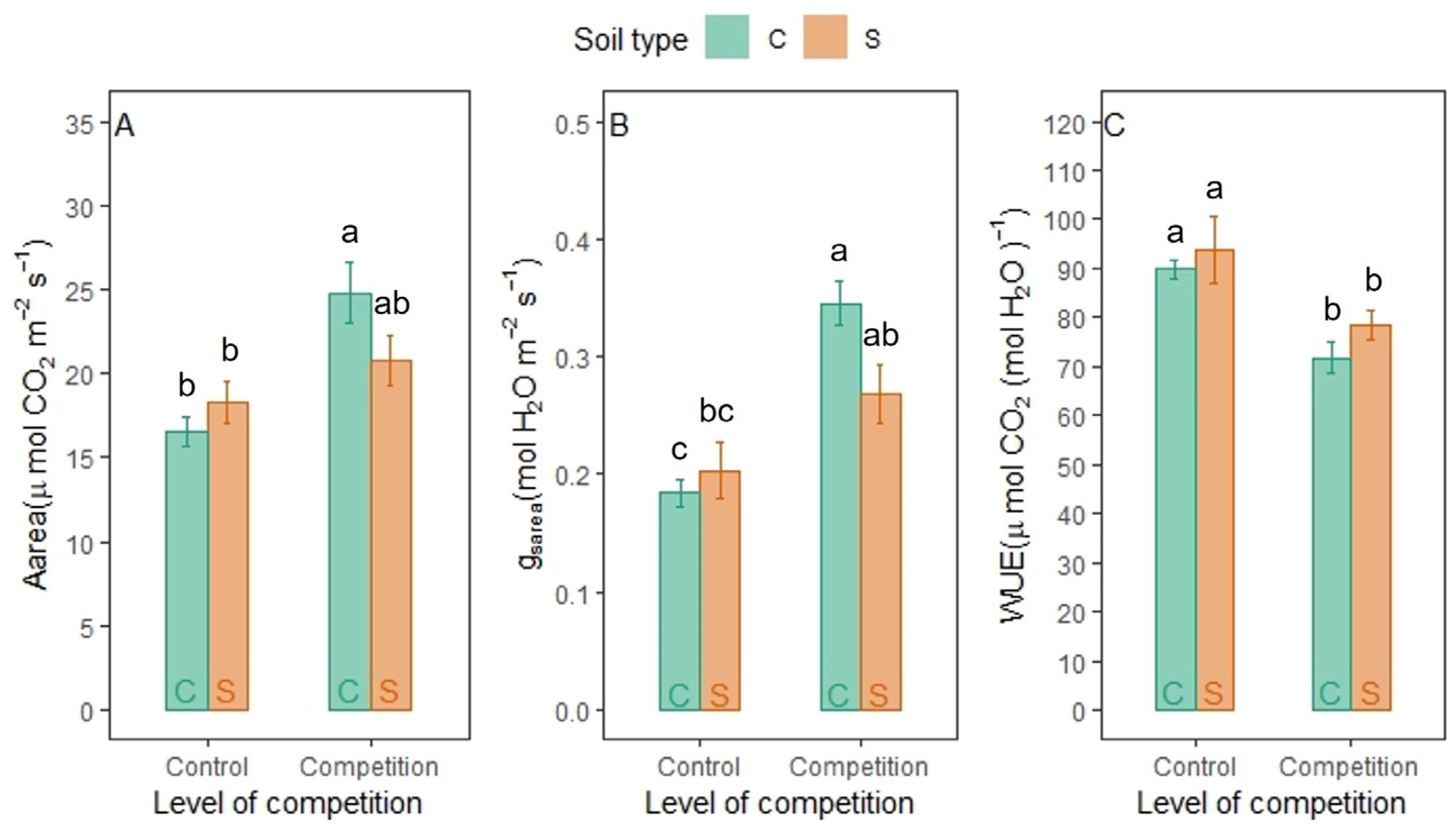
3.4. Physiological Response of LANZA® to Soil Type and Competition
4. Discussion
4.1. Effect of Competition
4.2. Effect of Soil Type
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cosentino, S.; Porqueddu, C.; Copani, V.; Patanè, C.; Testa, G.; Scordia, D.; Melis, R. European grasslands overview: Mediterranean region. In Proceedings of the 25th General Meeting of the European Grassland Federation: The Future of European Grasslands, Aberystwyth, UK, 7–11 September 2014; pp. 41–56. [Google Scholar]
- Bell, L.W.; Moore, A.D.; Thomas, D.T. Feed-base strategies that reduce risk of feed-gaps in livestock systems across Australia’s mixed farming zone. In Animal Production in Australia, Proceedings of the 31st Biennial Conference of the Australian Society of Animal Production, Adelaide, Australia, 5–7 July 2016; Australian Society of Animal Production (ASAP): Adelaide, Australia, 2016; pp. 1–2. [Google Scholar]
- Moore, A.D.; Bell, L.W.; Revell, D.K. Feed gaps in mixed-farming systems: Insights from the Grain & Graze program. Anim. Prod. Sci. 2009, 49, 736–748. [Google Scholar]
- Bell, L.W.; Robertson, M.J.; Revell, D.K.; Lilley, J.M.; Moore, A.D. Approaches for assessing some attributes of feed-base systems in mixed farming enterprises. Aust. J. Exp. Agric. 2008, 48, 789–798. [Google Scholar] [CrossRef]
- Giannakopoulos, C.; Le Sager, P.; Bindi, M.; Moriondo, M.; Kostopoulou, E.; Goodess, C.M. Climatic changes and associated impacts in the Mediterranean resulting from a 2 °C global warming. Glob. Planet. Chang. 2009, 68, 209–224. [Google Scholar] [CrossRef]
- Giorgi, F.; Lionello, P. Climate change projections for the Mediterranean region. Glob. Planet. Chang. 2008, 63, 90–104. [Google Scholar] [CrossRef]
- Ma, Z.; Liu, H.; Mi, Z.; Zhang, Z.; Wang, Y.; Xu, W.; Jiang, L.; He, J.-S. Climate warming reduces the temporal stability of plant community biomass production. Nat. Commun. 2017, 8, 15378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernández-Esteban, A.; López-Díaz, M.L.; Cáceres, Y.; Moreno, G. Are sown legume-rich pastures effective allies for the profitability and sustainability of Mediterranean dehesas? Agrofor. Syst. 2019, 93, 2047–2065. [Google Scholar] [CrossRef]
- Real, D.; Oldham, C.M.; Nelson, M.N.; Croser, J.; Castello, M.; Verbyla, A.; Pradhan, A.; Van Burgel, A.; Mendez, P.; Correal, E.; et al. Evaluation and breeding of tedera for Mediterranean climates in southern Australia. Crop Pasture Sci. 2014, 65, 1114–1131. [Google Scholar] [CrossRef]
- Thomas, D.T.; Flohr, B.M.; Monjardino, M.; Loi, A.; Llewellyn, R.S.; Lawes, R.A.; Norman, H.C. Selecting higher nutritive value annual pasture legumes increases the profitability of sheep production. Agric. Syst. 2021, 194, 103272. [Google Scholar] [CrossRef]
- Edwards, T.; Howieson, J.; Nutt, B.; Yates, R.; O’Hara, G.; Van Wyk, B.E. A ley-farming system for marginal lands based upon a self-regenerating perennial pasture legume. Agron. Sustain. Dev. 2019, 39, 13. [Google Scholar] [CrossRef] [Green Version]
- Moore, G.A.; Sanford, P.; Dolling, P.J.; Real, D. The challenges of developing resilient perennial pastures for a Mediterranean environment—A review for Western Australia. Crop Pasture Sci. 2021, 72, 613–633. [Google Scholar] [CrossRef]
- Rognli, O.A.; Pecetti, L.; Kovi, M.R.; Annicchiarico, P. Grass and legume breeding matching the future needs of European grassland farming. Grass Forage Sci. 2021, 76, 175–185. [Google Scholar] [CrossRef]
- Méndez, P.; Fernández, M.; Santos, A. Variedades de Bituminaria bituminosa (L.) Stirton (Leguminosae) en el archipélago canario. Pastos 1991, 20–21, 157–166. [Google Scholar]
- Foster, K.; Ryan, M.H.; Real, D.; Ramankutty, P.; Lambers, H. Drought resistance at the seedling stage in the promising fodder plant tedera (Bituminaria bituminosa var. albomarginata). Crop Pasture Sci. 2012, 63, 1034–1042. [Google Scholar] [CrossRef]
- Foster, K.; Lambers, H.; Real, D.; Ramankutty, P.; Cawthray, G.R.; Ryan, M.H. Drought resistance and recovery in mature Bituminaria bituminosa var. albomarginata. Ann. Appl. Biol. 2015, 166, 154–169. [Google Scholar] [CrossRef]
- Martínez-Fernández, D.; Walker, D.J.; Romero, P.; Martínez-Ballesta, M.C.; Correal, E. The Response of the Leguminous Fodder Plant Bituminaria bituminosa to Water Stress. J. Agron. Crop Sci. 2012, 198, 442–451. [Google Scholar] [CrossRef]
- Fernández-Habas, J.; Hidalgo-Fernández, M.T.; Leal-Murillo, J.R.; Méndez, P.; Quero, J.L.; Vanwalleghem, T.; Fernández-Rebollo, P. Effects of two water regimes on morphological traits, nutritive value and physiology of three Bituminaria bituminosa varieties from the Canary Islands. J. Agron. Crop Sci. 2021, 208, 413–426. [Google Scholar] [CrossRef]
- Ventura, M.R.; Castanon, J.I.R.; Mendez, P. Effect of season on tedera (Bituminaria bituminosa) intake by goats. Anim. Feed Sci. Technol. 2009, 153, 314–319. [Google Scholar] [CrossRef]
- Ventura, M.R.; Méndez, P.; Flores, M.P.; Rodriguez, R.; Castañon, J.I.R. Energy and protein content of Tedera. Analysis 1991, 82, 1988–1990. [Google Scholar]
- Finlayson, J.; Real, D.; Nordblom, T.; Revell, C.; Ewing, M.; Kingwell, R. Farm level assessments of a novel drought tolerant forage: Tedera (Bituminaria bituminosa C.H. Stirt var. albomarginata). Agric. Syst. 2012, 112, 38–47. [Google Scholar] [CrossRef]
- Oldham, C.M.; Wood, D.; Milton, J.; Real, D.; Vercoe, P.; Van Burgel, A.J. An animal house study on utilisation of fresh tedera (Bituminaria bituminosa var. albomarginata and crassiuscula) by Merino wethers. Anim. Prod. Sci. 2015, 55, 617–624. [Google Scholar] [CrossRef]
- Oldham, C.; Real, D.; Bailey, H.J.; Thomas, D.; Van Burgel, A.; Vercoe, P.; Correal, E.; Rios, S. Australian and Spanish scientists are collaborating in the domestication of tedera: Young Merino sheep grazing a monoculture of tedera in autumn showed preference for certain accessions but no signs of ill health. Crop Pasture Sci. 2013, 64, 399–408. [Google Scholar] [CrossRef]
- Real, D. Tedera (Bituminaria bituminosa). Plant Var. J. 2016, 29, 97. [Google Scholar]
- Real, D. Critical Agronomic Practices for Establishing the Recently Domesticated Perennial Herbaceous Forage Legume Tedera in Mediterranean-like Climatic Regions in Western Australia. Agronomy 2022, 12, 274. [Google Scholar] [CrossRef]
- Real, D.; Bennett, R.G.; Nazeri, N.K.; Weaver, D.M. Critical P, K and S Concentrations in Soil and Shoot Samples for Optimal Tedera Productivity and Nodulation. Agronomy 2022, 12, 1581. [Google Scholar] [CrossRef]
- Real, D.; Dhammu, H.; Moore, J.; Clegg, D.; van Burgel, A. Herbicide Tolerance Options for Weed Control in Lanza® Tedera. Agronomy 2022, 12, 1198. [Google Scholar] [CrossRef]
- Raeside, M.C.; Nie, Z.N.; Clark, S.G.; Partington, D.L.; Behrendt, R.; Real, D. Evaluation of tedera [(Bituminaria bituminosa (L.) C.H. Stirton var. albomarginata)] as a forage alternative for sheep in temperate southern Australia. Crop Pasture Sci. 2012, 63, 1135–1144. [Google Scholar] [CrossRef]
- Real, D.; Li, G.D.; Clark, S.; Albertsen, T.O.; Hayes, R.C.; Denton, M.D.; Dantuono, M.F.; Dear, B. Evaluation of perennial forage legumes and herbs in six Mediterranean environments. Chil. J. Agric. Res. 2011, 71, 357–369. [Google Scholar] [CrossRef] [Green Version]
- Bell, L.W.; Moore, G.A.; Ewing, M.A.; Bennett, S.J. Establishment and summer survival of the perennial legumes, Dorycnium hirsutum and D. rectum in Mediterranean environments. Aust. J. Exp. Agric. 2005, 45, 1245–1254. [Google Scholar] [CrossRef]
- Häring, D.A.; Scharenberg, A.; Heckendorn, F.; Dohme, F.; Lüscher, A.; Maurer, V.; Suter, D.; Hertzberg, H. Tanniferous forage plants: Agronomic performance, palatability and efficacy against parasitic nematodes in sheep. Renew. Agric. Food Syst. 2008, 23, 19–29. [Google Scholar] [CrossRef] [Green Version]
- Hogg, N.; Davis, J.K. What is hindering the adoption of new annual pasture legumes? Extension requirements to overcome these barriers. Ext. Farming Syst. J. 2009, 5, 29–38. [Google Scholar]
- Lewis, C.; Farquharson, R.; Leury, B.; Behrendt, R.; Clark, S. Economic analysis of improved perennial pasture systems. Aust. Farm Bus. Manag. J. 2012, 9, 37–56. [Google Scholar]
- Carruggio, F.; Onofri, A.; Impelluso, C.; Del Galdo, G.G.; Scopece, G.; Cristaudo, A. Seed dormancy breaking and germination in Bituminaria basaltica and B. bituminosa (fabaceae). Plants 2020, 9, 1110. [Google Scholar] [CrossRef] [PubMed]
- Pecetti, L.; Tava, A.; Pagnotta, M.; Russi, L. Variation in forage quality and chemical composition among Italian accessions of Bituminaria bituminosa (L.) Stirt. J. Sci. Food Agric. 2007, 87, 985–991. [Google Scholar] [CrossRef] [Green Version]
- Reyna-Bowen, L.; Fernandez-Rebollo, P.; Fernández-Habas, J.; Gómez, J.A. The influence of tree and soil management on soil organic carbon stock and pools in dehesa systems. Catena 2020, 190, 104511. [Google Scholar] [CrossRef]
- García, M.C.; García, A.D.; Gavilán, A.G.; Castellet, J.T. Estudio Edafológico de la Finca ‘Rabanales’ Córdoba, Spain; Departamento de Ciencias y Recursos Agrícolas y Forestales, Universidad de Córdoba: Córdoba, Spain, 1993. [Google Scholar]
- Eijkelkamp. Sandbox for pF-Determination. User Manual; Eijkelkamp: Giesbeek, The Netherlands, 2019. [Google Scholar]
- Eijkelkamp. 08.02 Sand/Kaolin Box. Operating Instructions; Eijkelkamp: Giesbeek, The Netherlands, 2016. [Google Scholar]
- Decagon. WP4 Dewpoint PotentialMeter Operator’s Manual Version 5; Decagon Devices, Inc.: Pullman, WA, USA, 2007. [Google Scholar]
- Van Genuchten, M.T. A closed-form equation for predicting the hydraulic conductivity of unsaturated soils. Soil Sci. Soc. Am. J. 1980, 44, 892–898. [Google Scholar] [CrossRef] [Green Version]
- Van Genuchten, M.T.; Leij, F.J.; Yates, S.R. The RETC Code for Quantifying the Hydraulic Functions of Unsaturated Soils. In Version 1.0. EPA Report 600/2-91/065; U.S. Salinity Laboratory: Riverside, CA, USA, 1991. [Google Scholar]
- Bleiholder, H.; Weber, E.; Lancashire, P.; Feller, C.; Buhr, L.; Hess, M.; Wicke, H.; Hack, H.; Meier, U.; Klose, R. Growth Stages of Mono-and Dicotyledonous Plants, BBCH Monograph; Federal Biological Research Centre for Agriculture and Forestry: Berlin/Braunschweig, Germany, 2001.
- Christensen, R.H.B. A Tutorial on Fitting Cumulative Link Mixed Models with clmm2 from the Ordinal Package. 1-18. Available online: https://cran.r-project.org/web/packages/ordinal/vignettes/clmm2_tutorial.pdf (accessed on 15 December 2022).
- Agresti, A. Categorical Data Analysis; John Wiley & Sons: Hoboken, NJ, USA, 2003. [Google Scholar]
- Christensen, R.H.B. Ordinal—Regression Models for Ordinal Data, R package version 2022.11-16. 2022. Available online: https://cran.r-project.org/web/packages/ordinal/index.html (accessed on 15 December 2022).
- Mangiafico, S.S. One-Way Repeated Ordinal Regression with CLMM. Available online: https://rcompanion.org/handbook/G_08.html (accessed on 7 June 2022).
- Hervé, M. Package ‘RVAideMemoire’: Testing and Plotting Procedures for Biostatistics. 2022. Available online: https://cran.r-project.org/web/packages/RVAideMemoire/index.html (accessed on 15 December 2022).
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: https://www.R-project.org/ (accessed on 7 November 2022).
- Bell, L.; Lawrence, J.; Jasper, S.; Guppy, C. Pasture legume production severely reduced when co-sown with winter forage cereals. Capturing Opportunities and Overcoming Obstacles in Australian Agronomy. In Proceedings of the 16th ASA Conference, Armidale, Australia, 14–18 October 2012; Australian Society of Agronomy Inc.: Armidale, Australia, 2012. [Google Scholar]
- Olea, L.; San Miguel-Ayanz, A. The Spanish dehesa. A traditional Mediterranean silvopastoral system linking production and nature conservation. In Sustainable Grassland Productivity, Proceedings of the 21st General Meeting of the European Grassland Federation, Badajoz, Spain, 3–6 April 2006; Lloveras, J., González-Rodríguez, A., Vázquez-Yáñez, O., Piñeiro, J., Santamaría, O., Olea, L., Poblaciones, M.J., Eds.; Sociedad Española para el Estudio de los Pastos: Badajoz, Spain, 2006; pp. 1–13. [Google Scholar]
- Evans, J. Photosynthesis and nitrogen relationships in leaves of C3 plants. Oecologia 1989, 78, 9–19. [Google Scholar] [CrossRef]
- Kiær, L.P.; Weisbach, A.N.; Weiner, J. Root and shoot competition: A meta-analysis. J. Ecol. 2013, 101, 1298–1312. [Google Scholar] [CrossRef]
- Lüscher, A.; Connolly, J.; Jacquard, P. Neighbour specificity between Lolium perenne and Trifolium repens from a natural pasture. Oecologia 1992, 91, 404–409. [Google Scholar] [CrossRef]
- Semchenko, M.; Abakumova, M.; Lepik, A.; Zobel, K. Plants are least suppressed by their frequent neighbours: The relationship between competitive ability and spatial aggregation patterns. J. Ecol. 2013, 101, 1313–1321. [Google Scholar] [CrossRef]
- Malisch, C.S.; Suter, D.; Studer, B.; Lüscher, A. Multifunctional benefits of sainfoin mixtures: Effects of partner species, sowing density and cutting regime. Grass Forage Sci. 2017, 72, 794–805. [Google Scholar] [CrossRef]
- Volaire, F.; Barkaoui, K.; Norton, M. Designing resilient and sustainable grasslands for a drier future: Adaptive strategies, functional traits and biotic interactions. Eur. J. Agron. 2014, 52, 81–89. [Google Scholar] [CrossRef] [Green Version]
- Myers, N.; Mittermeler, R.A.; Mittermeler, C.G.; Da Fonseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Plieninger, T.; Flinzberger, L.; Hetman, M.; Horstmannshoff, I.; Reinhard-Kolempas, M.; Topp, E.; Moreno, G.; Huntsinger, L. Dehesas as high nature value farming systems: A social-ecological synthesis of drivers, pressures, state, impacts, and responses. Ecol. Soc. 2021, 26, 23. [Google Scholar] [CrossRef]
- Real, D.; Department of Primary Industries and Regional Development (DPIRD), Perth, WA, Australia. Personal communication, 2021.
- Lavorel, S.; McIntyre, S.; Grigulis, K. Plant response to disturbance in a Mediterranean grassland: How many functional groups? J. Veg. Sci. 1999, 10, 661–672. [Google Scholar] [CrossRef]
- Levassor, C.; Ortega, M.; Peco, B. Seed bank dynamics of Mediterranean pastures subjected to mechanical disturbance. J. Veg. Sci. 1990, 1, 339–344. [Google Scholar] [CrossRef]
- Volaire, F. A unified framework of plant adaptive strategies to drought: Crossing scales and disciplines. Glob. Chang. Biol. 2018, 24, 2929–2938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foxx, A.J.; Fort, F. Root and shoot competition lead to contrasting competitive outcomes under water stress: A systematic review and meta-analysis. PLoS ONE 2019, 14, e0220674. [Google Scholar] [CrossRef] [Green Version]
- Ben-Noah, I.; Friedman, S.P. Aeration of clayey soils by injecting air through subsurface drippers: Lysimetric and field experiments. Agric. Water Manag. 2016, 176, 222–233. [Google Scholar] [CrossRef]
- Drew, M.C.; Lynch, J.M. Soil Anaerobiosis, Microorganisms, and Root Function. Annu. Rev. Phytopathol. 1980, 18, 37–66. [Google Scholar] [CrossRef]
- Friedman, S.P.; Naftaliev, B. A survey of the aeration status of drip-irrigated orchards. Agric. Water Manag. 2012, 115, 132–147. [Google Scholar] [CrossRef]
- Bhattarai, S.P.; Huber, S.; Midmore, D.J. Aerated subsurface irrigation water gives growth and yield benefits to zucchini, vegetable soybean and cotton in heavy clay soils. Ann. Appl. Biol. 2004, 144, 285–298. [Google Scholar] [CrossRef]
- Irving, L.J.; Sheng, Y.B.; Woolley, D.; Matthew, C. Physiological effects of waterlogging on two lucerne varieties grown under glasshouse conditions. J. Agron. Crop Sci. 2007, 193, 345–356. [Google Scholar] [CrossRef]
- Haling, R.E.; Yang, Z.; Shadwell, N.; Culvenor, R.A.; Stefanski, A.; Ryan, M.H.; Lambers, H.; Simpson, R.J. Root morphological traits that determine phosphorus-acquisition efficiency and critical external phosphorus requirement in pasture species. Funct. Plant Biol. 2016, 43, 815–826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, J.O.; Simpson, R.J.; Moore, A.D.; Chapman, D.F. Morphology and response of roots of pasture species to phosphorus and nitrogen nutrition. Plant Soil 2006, 286, 7–19. [Google Scholar] [CrossRef]
- Pang, J.; Ryan, M.H.; Tibbett, M.; Cawthray, G.R.; Siddique, K.H.M.; Bolland, M.D.A.; Denton, M.D.; Lambers, H. Variation in morphological and physiological parameters in herbaceous perennial legumes in response to phosphorus supply. Plant Soil 2010, 331, 241–255. [Google Scholar] [CrossRef]
- Pang, J.; Tibbett, M.; Denton, M.D.; Lambers, H.; Siddique, K.H.M.; Bolland, M.D.A.; Revell, C.K.; Ryan, M.H. Variation in seedling growth of 11 perennial legumes in response to phosphorus supply. Plant Soil 2010, 328, 133–143. [Google Scholar] [CrossRef]
- Edmeades, D.C.; Metherell, A.K.; Waller, J.E.; Roberts, A.H.C.; Morton, J.D. Defining the relationships between pasture production and soil P and the development of a dynamic P model for New Zealand pastures: A review of recent developments. N. Z. J. Agric. Res. 2006, 49, 207–222. [Google Scholar] [CrossRef]
- Moody, P.W.; Speirs, S.D.; Scott, B.J.; Mason, S.D. Soil phosphorus tests I: What soil phosphorus pools and processes do they measure? Crop Pasture Sci. 2013, 64, 461–468. [Google Scholar] [CrossRef]
- Saggar, S.; Hedley, M.J.; White, R.E.; Perrott, K.W.; Gregg, P.E.H.; Cornforth, I.S.; Sinclair, A.G. Development and evaluation of an improved soil test for phosphorus, 3: Field comparison of Olsen, Colwell and Resin soil P tests for New Zealand pasture soils. Nutr. Cycl. Agroecosyst. 1999, 55, 35–50. [Google Scholar] [CrossRef]
- Mendez, P.; Santos, A.; Correal, E.; Ríos, S. Agronomic traits as forage crops of nineteen populations of Bituminaria bituminosa. In Proceedings of the 21st General Meeting of the European Grassland Federation, Badajoz, Spain, 3–6 April 2006; Sociedad Española para el Estudio de los Pastos (SEEP): Madrid, Spain, 2006; pp. 300–302. [Google Scholar]
- Martínez-Fernández, D.; Walker, D.J.; Romero-Espinar, P.; Flores, P.; del Río, J.A. Physiological responses of Bituminaria bituminosa to heavy metals. J. Plant Physiol. 2011, 168, 2206–2211. [Google Scholar] [CrossRef]
- Walker, D.J.; Bernal, M.P.; Correal, E. The influence of heavy metals and mineral nutrient supply on Bituminaria bituminosa. Water Air Soil Pollut. 2007, 184, 335–345. [Google Scholar] [CrossRef]
- Martínez-Fernández, D. Respuestas Fisiológicas de Bituminaria Bituminosa Frente a Sequía y Metales Pesados. Ph.D. Thesis, Universidad Politécnica de Cartagena, Cartagena, Spain, 2012. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Units | Loamy Sand (S) | Clay (C) |
|---|---|---|---|
| Sand | % | 73 | 29 |
| Silt | % | 13 | 25 |
| Clay | % | 14 | 46 |
| Bulk density | g cm−3 | 1.48 | 1.02 |
| Soil pH in water (12.5) | U. pH | 7.1 | 8.12 |
| Cation exchange capacity | Meq 100 g−1 | 11 | 31 |
| Electrical conductivity 25 °C (1:5) | mS cm−1 | 0.16 | 0.22 |
| Oxidable Organic matter content | % | 1.97 | 3.21 |
| N (Kjeldahl) | % | 0.11 | 0.19 |
| Available P (Olsen) | mg kg−1 | 44 | 7 |
| Exchangeable K (NH4Cl) | mg kg−1 | 138 | 319 |
| Exchangeable Ca (NH4Cl) | mg kg−1 | 1.12 | 4.91 |
| Exchangeable Mg (NH4Cl) | mg kg−1 | 114 | 371 |
| Exchangeable Na (NH4Cl) | mg kg−1 | 102 | 118 |
| Saturated volumetric water content | cm3 cm−3 | 0.33 | 0.62 |
| Level of Competition | Soil Type | Thick Roots Dry Mass (g) | Thin Roots Dry Mass (g) | Thin Roots Proportion (g) |
|---|---|---|---|---|
| Control | S | 7.1 ± 0.9 a | 9.0 ± 1.5 a | 0.55 ± 0.04 a |
| C | 6.8 ± 1.1 a | 5.0 ± 0.8 b | 0.42 ± 0.01 b | |
| Competition | S | 1.0 ± 0.3 b | - | - |
| C | 0.4 ± 0.1 b | - | - |
| Level of Competition | Soil Type | N | P | C N−1 | N P−1 |
|---|---|---|---|---|---|
| Control | S | 2.10 ± 0.07 b | 0.42 ± 0.03 a | 20.7 ± 0.67 a | 5.16 ± 0.45 b |
| Control | C | 2.30 ± 0.14 b | 0.12 ± 0.02 b | 19.4 ± 0.95 a | 21.4 ± 2.58 a |
| Competition | S | 2.58 ± 0.16 a | 0.33 ± 0.06 a | 17.5 ± 0.97 b | 9.03 ± 1.46 b |
| Competition | C | 2.89 ± 0.37 a | 0.13 ± 0.01 b | 16.1 ± 1.86 b | 22.8 ± 2.36 a |
| Level of competition | Soil type | K | Ca | Mg | S |
| Control | S | 2.69 ± 0.22 n.s. | 1.29 ± 0.09 n.s. | 0.85 ± 0.04 a | 0.24 ± 0.01 d |
| Control | C | 2.16 ± 0.10 n.s. | 1.24 ± 0.17 n.s. | 0.52 ± 0.08 b | 0.27 ± 0.01 c |
| Competition | S | 2.63 ± 0.18 n.s. | 1.23 ± 0.13 n.s. | 0.68 ± 0.05 ab | 0.30 ± 0.02 b |
| Competition | C | 2.60 ± 0.18 n.s. | 1.19 ± 0.14 n.s. | 0.78 ± 0.08 a | 0.32 ± 0.01 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernández-Habas, J.; Real, D.; Vanwalleghem, T.; Fernández-Rebollo, P. LANZA® Tedera Is Strongly Suppressed by Competition from Lolium multiflorum and Is Best Adapted to Light-Textured Soils. Agronomy 2023, 13, 965. https://doi.org/10.3390/agronomy13040965
Fernández-Habas J, Real D, Vanwalleghem T, Fernández-Rebollo P. LANZA® Tedera Is Strongly Suppressed by Competition from Lolium multiflorum and Is Best Adapted to Light-Textured Soils. Agronomy. 2023; 13(4):965. https://doi.org/10.3390/agronomy13040965
Chicago/Turabian StyleFernández-Habas, Jesús, Daniel Real, Tom Vanwalleghem, and Pilar Fernández-Rebollo. 2023. "LANZA® Tedera Is Strongly Suppressed by Competition from Lolium multiflorum and Is Best Adapted to Light-Textured Soils" Agronomy 13, no. 4: 965. https://doi.org/10.3390/agronomy13040965