
The Changes in Rhizosphere Metabolome and Microbiota Are the Main Direct Obstacles to Continuous Cropping in Tobacco (Nicotiana tabacum L.)
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
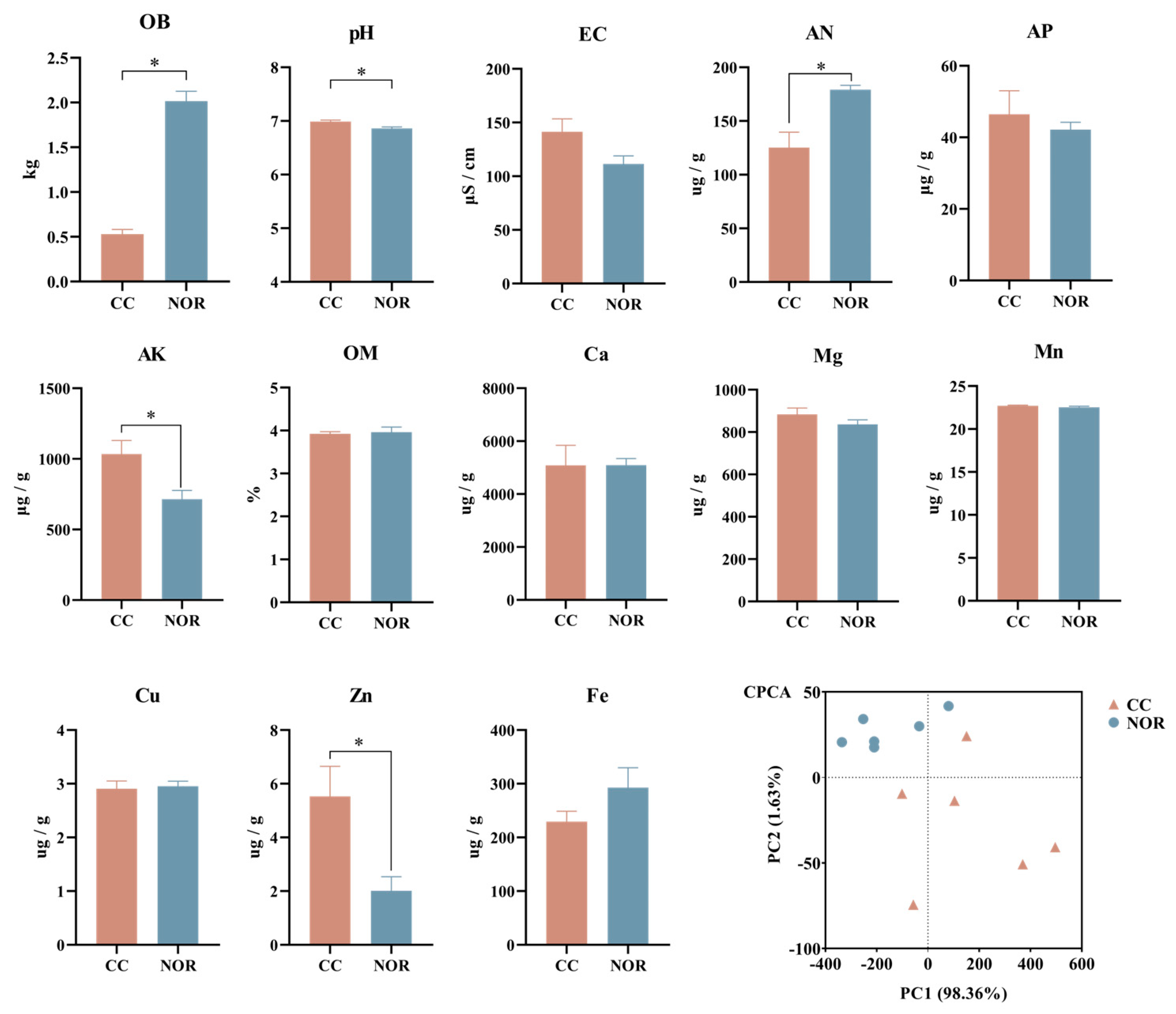
2.2. Soil Physicochemical Indicators
2.3. Microbial Community by Illumina MiSeq Sequencing
2.4. Chemical Composition of Rhizosphere Soil by GC-MS
2.5. Statistics and Analysis
3. Results
3.1. The Biomass of Continuous Cropping Obstacle and Normal Tobacco
3.2. Soil Physicochemical Properties of Continuous Cropping Obstacle and Normal Tobacco
3.3. Rhizosphere Soil Microbial Communities
3.4. Rhizosphere Soil Metabolomics
3.5. Multiple Linear Regression Model
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Kessler, A.; Baldwin, I.T. Defensive function of herbivore-induced plant volatile emissions in nature. Science 2001, 291, 2141–2144. [Google Scholar] [CrossRef] [PubMed]
- Chae, L.; Kim, T.; Nilo-Poyanco, R.; Rhee, S.Y. Genomic signatures of specialized metabolism in plants. Science 2014, 344, 510–513. [Google Scholar] [CrossRef]
- Huang, A.C.; Jiang, T.; Liu, Y.X.; Bai, Y.C.; Reed, J.; Qu, B.; Goossens, A.; Nützmann, H.-W.; Bai, Y.; Osbourn, A. A specialized metabolic network selectively modulates Arabidopsis root microbiota. Science 2019, 364, eaau6389. [Google Scholar] [CrossRef]
- Li, J.; Halitschke, R.; Li, D.; Paetz, C.; Su, H.; Heiling, S.; Xu, S.; Baldwin, I.T. Controlled hydroxylations of diterpenoids allow for plant chemical defense without autotoxicity. Science 2021, 371, 255–260. [Google Scholar] [CrossRef] [PubMed]
- Van der Putten, W.H.; Bardgett, R.D.; Bever, J.D.; Bezemer, T.M.; Casper, B.B.; Fukami, T.; Kardol, P.; Klironomos, J.N.; Kulmatiski, A.; Schweitzer, J.A. Plant–soil feedbacks: The past, the present and future challenges. J. Ecol. 2013, 101, 265–276. [Google Scholar] [CrossRef]
- Reynolds, H.L.; Packer, A.; Bever, J.D.; Clay, K. Grassroots ecology: Plant–microbe–soil interactions as drivers of plant community structure and dynamics. Ecology 2003, 84, 2281–2291. [Google Scholar] [CrossRef] [Green Version]
- Packer, A.; Clay, K. Soil pathogens and spatial patterns of seedling mortality in a temperate tree. Nature 2000, 404, 278–281. [Google Scholar] [CrossRef]
- Bever, J.D. Soil community feedback and the coexistence of competitors: Conceptual frameworks and empirical tests. New Phytol. 2003, 157, 465–473. [Google Scholar] [CrossRef] [Green Version]
- Cesarano, G.; Zotti, M.; Antignani, V.; Marra, R.; Scala, F.; Bonanomi, G. Soil sickness and negative plant-soil feedback: A reappraisal of hypotheses. J. Plant Pathol. 2017, 99, 545–570. [Google Scholar]
- Luo, L.; Guo, C.; Wang, L.; Zhang, J.; Deng, L.; Luo, K.; Huang, H.; Liu, Y.; Mei, X.; Zhu, S. Negative plant-soil feedback driven by re-assemblage of the rhizosphere microbiome with the growth of Panax notoginseng. Front. Microbiol. 2019, 10, 1597. [Google Scholar] [CrossRef] [Green Version]
- Bao, L.; Liu, Y.; Ding, Y.; Shang, J.; Wei, Y.; Tan, Y.; Zi, F. Interactions between phenolic acids and microorganisms in rhizospheric soil from continuous cropping of Panax notoginseng. Front. Microbiol. 2022, 13, 791603. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Wang, Y.Z.; Liu, Q.Z.; Zhang, Y.T.; Li, X.Y.; Li, H.Q.; Li, W.H. Phase changes of continuous cropping obstacles in strawberry (Fragaria × ananassa Duch.) production. Appl. Soil Ecol. 2020, 155, 103626. [Google Scholar] [CrossRef]
- Liu, J.; Xie, M.; Li, X.; Jin, H.; Yang, X.; Yan, Z.; Su, A.; Qin, B. Main allelochemicals from the rhizosphere soil of Saussurea lappa (Decne.) Sch. Bip. and their effects on plants’ antioxidase systems. Molecules 2018, 23, 2506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Z.; Han, M.; Hu, Y.; Li, Z.; Liu, C.; Wang, X.; Tian, Q.; Jiao, W.; Hu, J.; Liu, L.; et al. Effects of continuous cropping of sweet potato on the fungal community structure in rhizospheric soil. Front. Microbiol. 2019, 10, 2269. [Google Scholar] [CrossRef] [PubMed]
- Niu, J.; Chao, J.; Xiao, Y.; Chen, W.; Zhang, C.; Liu, X.; Rang, Z.; Yin, H.; Dai, L. Insight into the effects of different cropping systems on soil bacterial community and tobacco bacterial wilt rate. J. Basic Microbiol. 2017, 57, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Han, X.; Zhao, D.; Wei, K.; Yuan, Y.; Li, Y.; Liu, M.; Zhang, C.S. Exploring biocontrol agents from microbial keystone taxa associated to suppressive soil: A new attempt for a biocontrol strategy. Front. Plant Sci. 2021, 12, 407. [Google Scholar] [CrossRef] [PubMed]
- Roberts, T.L.; Ross, W.J.; Norman, R.J.; Slaton, N.A.; Wilson, C.E., Jr. Predicting nitrogen fertilizer needs for rice in arkansas using alkaline hydrolyzable-nitrogen. Soil Sci. Soc. Am. J. 2011, 75, 1161–1171. [Google Scholar] [CrossRef] [Green Version]
- Anderson, J.M.; Ingram, J. Tropical Soil Biology and Fertility: A Handbook of Methods, 2nd ed.; CAB International: Wallingford, UK, 1993; Volume 157, p. 221. [Google Scholar] [CrossRef]
- Liu, X.; Xu, J.; Zhang, M.; Zhou, B. Effects of land management change on spatial variability of organic matter and nutrients in paddy field: A case study of Pinghu, China. Environ. Manag. 2004, 34, 691–700. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Kaehler, B.D.; Bokulich, N.A.; McDonald, D.; Knight, R.; Caporaso, J.G.; Huttley, G.A. Species abundance information improves sequence taxonomy classification accuracy. Nat. Commun. 2019, 10, 4643. [Google Scholar] [CrossRef] [Green Version]
- Li, W.H.; Liu, Q.Z. Changes in fungal community and diversity in strawberry rhizosphere soil after 12 years in the greenhouse. J. Integr. Agric. 2019, 18, 677–687. [Google Scholar] [CrossRef]
- Li, W.H.; Liu, Q.Z.; Peng, C. Effect of long-term continuous cropping of strawberry on soil bacterial community structure and diversity. J. Integr. Agric. 2018, 17, 2570–2582. [Google Scholar] [CrossRef]
- Xie, X.; Pu, L.; Wang, Q.; Zhu, M.; Xu, Y.; Zhang, M. Response of soil physicochemical properties and enzyme activities to long-term reclamation of coastal saline soil, Eastern China. Sci. Total Environ. 2017, 607, 1419–1427. [Google Scholar] [CrossRef] [PubMed]
- Wilson, S.; Andrews, P.; Nair, T.S. Non-fumigant management of apple replant disease. Sci. Hortic. 2004, 102, 221–231. [Google Scholar] [CrossRef]
- Mendes, R.; Garbeva, P.; Raaijmakers, J.M. The rhizosphere microbiome: Significance of plant beneficial, plant pathogenic, and human pathogenic microorganisms. FEMS Microbiol. Rev. 2013, 37, 634–663. [Google Scholar] [CrossRef] [Green Version]
- Tan, G.; Liu, Y.; Peng, S.; Yin, H.; Meng, D.; Tao, J.; Gu, Y.; Li, J.; Yang, S.; Xiao, N. Soil potentials to resist continuous cropping obstacle: Three field cases. Environ. Res. 2021, 200, 111319. [Google Scholar] [CrossRef] [PubMed]
- Yuan, S.; Wang, L.; Wu, K.; Shi, J.; Wang, M.; Yang, X.; Shen, Q.; Shen, B. Evaluation of Bacillus-fortified organic fertilizer for controlling tobacco bacterial wilt in greenhouse and field experiments. Appl. Soil Ecol. 2014, 75, 86–94. [Google Scholar] [CrossRef]
- Van Bruggen, A.H.C.; Semenov, A.M. In search of biological indicators for soil health and disease suppression. Appl. Soil Ecol. 2000, 15, 13–24. [Google Scholar] [CrossRef]
- Venkatachalam, S.; Kannan, V.M.; Saritha, V.N.; Loganathachetti, D.S.; Mohan, M.; Krishnan, K.P. Bacterial diversity and community structure along the glacier foreland of Midtre Lovénbreen, Svalbard, Arctic. Ecol. Indic. 2021, 126, 107704. [Google Scholar] [CrossRef]
- Dams, R.I.; Paton, G.I.; Killham, K. Rhizoremediation of pentachlorophenol by Sphingobium chlorophenolicum ATCC 39723. Chemosphere 2007, 68, 864–870. [Google Scholar] [CrossRef]
- Xiao, R.; Zhu, W.; Zheng, Y.; Xu, S.; Lu, H. Active assimilators of soluble microbial products produced by wastewater anammox bacteria and their roles revealed by DNA-SIP coupled to metagenomics. Environ. Int. 2022, 164, 107265. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Zhu, G.; Tao, J.; Zhang, Y. Iron carriers promote biofilm formation and p-nitrophenol degradation. Chemosphere 2022, 293, 133601. [Google Scholar] [CrossRef]
- Wang, Y.; Du, L.; Liu, H.; Long, D.; Huang, M.; Wang, Y.; Huang, S.; Jin, D. Halosulfuron methyl did not have a significant effect on diversity and community of sugarcane rhizosphere microflora. J. Hazard. Mater. 2020, 399, 123040. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Liu, A.; Fu, W.; Peng, D.; Wang, G.; Ji, J.; Jin, C.; Guan, C. Tobacco-associated with Methylophilus sp. FP-6 enhances phytoremediation of benzophenone-3 through regulating soil microbial community, increasing photosynthetic capacity and maintaining redox homeostasis of plant. J. Hazard. Mater. 2022, 431, 128588. [Google Scholar] [CrossRef]
- Yoon, J.H.; Cho, Y.G.; Lee, S.T.; Suzuki, K.I.; Nakase, T.; Park, Y.H. Nocardioides nitrophenolicus sp. nov., a p-nitrophenol-degrading bacterium. Int. J. Syst. Evol. Microbiol. 1999, 49, 675–680. [Google Scholar] [CrossRef] [Green Version]
- Habe, H.; Miyakoshi, M.; Chung, J.; Kasuga, K.; Yoshida, T.; Nojiri, H.; Omori, T. Phthalate catabolic gene cluster is linked to the angular dioxygenase gene in Terrabacter sp. strain DBF63. Appl. Microbiol. Biotechnol. 2003, 61, 44–54. [Google Scholar] [CrossRef]
- Zhao, Y.X.; Guo, L.; Wang, L.; Jiang, N.D.; Chen, K.X.; Dai, Y.J. Biodegradation of the pyridinecarboxamide insecticide flonicamid by Microvirga flocculans and characterization of two novel amidases involved. Ecotoxicol. Environ. Saf. 2021, 220, 112384. [Google Scholar] [CrossRef]
- Gao, L.; Ma, Y.; Zhao, W.; Wei, Z.; Gleason, M.L.; Chen, H.; Hao, L.; Sun, G.; Zhang, R. Three new species of Cyphellophora (Chaetothyriales) associated with sooty blotch and flyspeck. PLoS ONE 2015, 10, e0136857. [Google Scholar] [CrossRef]
- Pistán, M.E.; Gutiérrez, S.A.; Schnittger, L.; Gardner, D.R.; Cholich, L.A.; Gonzalez, A.M. Localization of the fungal symbiont (Chaetothyriales) in Ipomoea carnea. Botany 2022, 100, 729–736. [Google Scholar] [CrossRef]
- Keinath, A.P. From Native Plants in Central Europe to Cultivated Crops Worldwide: The Emergence of Didymella bryoniae as a Cucurbit Pathogen. HortScience Horts 2011, 46, 532–535. [Google Scholar] [CrossRef] [Green Version]
- Abbott, S.P.; Sigler, L.; McAleer, R.; McGough, D.A.; Rinaldi, M.G.; Mizell, G. Fatal cerebral mycoses caused by the ascomycete Chaetomium strumarium. J. Clin. Microbiol. 1995, 33, 2692–2698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, P.; Guo, Y.; Li, Y.; Duan, T. Effects of dual inoculation of AMF and rhizobium on alfalfa (Medicago sativa) root rot caused by Microdochium tabacinum. Australas. Plant Pathol. 2018, 47, 195–203. [Google Scholar] [CrossRef]
- Levenfors, J.P.; Eberhard, T.H.; Levenfors, J.J.; Gerhardson, B.; Hökeberg, M. Biological control of snow mould (Microdochium nivale) in winter cereals by Pseudomonas brassicacearum, MA250. BioControl 2008, 53, 651–665. [Google Scholar] [CrossRef]
- Pinruan, U.; Unartngam, J.; Unartngam, A.; Piyaboon, O.; Sommai, S.; Khamsuntorn, P. Paramyrothecium eichhorniae sp. nov., causing leaf blight disease of water hyacinth from Thailand. Mycobiology 2022, 50, 12–19. [Google Scholar] [CrossRef]
- Zhang, Q.; Wijayawardene, N.N.; Hyde, K.D.; Wang, Y.; Pan, X.J. Xepicula yifeii sp. nov. caused a leaf blight of Lasia spinosa (Araceae) in South China karst. Eur. J. Plant Pathol. 2020, 158, 121–134. [Google Scholar] [CrossRef]
- Feng, Y.; Zhang, H.; Song, X.; Ge, T.; Zhu, J.; Zhou, C.; Cobb, K.; Yan, X.; Ruan, R.; Cheng, P. Microalgae as a potential conditioner for continuous cropping obstacles for taro (Colocasia esculenta L. Schott) production. J. Clean. Prod. 2022, 369, 133356. [Google Scholar] [CrossRef]
- Huiyong, Y.; Hongbo, L.; Guoming, S.; Sampietro, D.A.; Xinxin, G. Effects of allelochemicals from tobacco root exudates on seed germination and seedling growth of tobacco. Allelopath. J. 2014, 33, 107. [Google Scholar] [CrossRef]
- Chen, Y.D.; Zhu, Y.; Xin, J.P.; Zhao, C.; Tian, R.N. Succinic acid inhibits photosynthesis of Microcystis aeruginosa via damaging PSII oxygen-evolving complex and reaction center. Environ. Sci. Pollut. Res. 2021, 28, 58470–58479. [Google Scholar] [CrossRef]
- Muzaffar, S.; Ali, B.; Wani, N.A. Effect of catechol, gallic acid and pyrogallic acid on the germination, seedling growth and the level of endogenous phenolics in cucumber (Cucumis sativus L.). Int. J. Life Sci. Biotechnol. Pharma Res. 2012, 1, 50–55. [Google Scholar]
- Kawahigashi, H. Transgenic plants for phytoremediation of herbicides. Curr. Opin. Biotechnol. 2009, 20, 225–230. [Google Scholar] [CrossRef]
- Hirose, J. Diversity and evolution of integrative and conjugative elements involved in bacterial aromatic compound degradation and their utility in environmental remediation. Microorganisms 2023, 11, 438. [Google Scholar] [CrossRef]
- Jia, Z.H.; Yi, J.H.; Su, Y.R.; Shen, H. Autotoxic substances in the root exudates from continuous tobacco cropping. Allelopath. J. 2011, 27, 87–96. [Google Scholar]
- Bie, C.C.; Li, F.M.; Li, Y.Y.; Wang, Z.Y. Effects of allelochemical dibutyl phthalate on Gymnodinium breve reactive oxygen species. Huan Jing Ke Xue 2012, 33, 442–447. [Google Scholar]
- Bie, C.C.; Li, F.M.; Wang, Y.F.; Wang, H.Y.; Zhao, Y.H.; Zhao, W.; Wang, Z.Y. Mechanism of the inhibitory action of allelochemical dibutyl phthalate on algae Gymnodinium breve. Huan Jing Ke Xue 2012, 33, 228–232. [Google Scholar] [PubMed]
- Dampanaboina, L.; Yuan, N.; Mendu, V. Estimation of plant biomass lignin content using Thioglycolic Acid (TGA). J. Vis. Exp. 2021, 173, e62055. [Google Scholar] [CrossRef]
- Wang, J.; Shi, Y.; Zhang, M.; Li, Y.; Liu, Q.; Xie, S. Allelopathically inhibitory effects of 5, 4′-dihydroxyflavone on the growth, antioxidant system and photosynthesis of the bloom-forming cyanobacterium Microcystis aeruginosa. J. Appl. Phycol. 2021, 33, 3843–3852. [Google Scholar] [CrossRef]
- Iwashina, T. Flavonoid function and activity to plants and other organisms. Biol. Sci. Space 2003, 17, 24–44. [Google Scholar] [CrossRef] [Green Version]
- Einhellig, F.A. Mechanism of Action of Allelochemicals in Allelopathy; ACS Publications: Washington, DC, USA, 1995. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.N.; Liu, B.Y.; Ge, F.J.; He, Y.; Lu, Z.Y.; Zhou, Q.H.; Zhang, Y.Y.; Wu, Z.B.; Gao, Y.N.; Liu, B.Y. Joint effects of allelochemical nonanoic acid, N-phenyl-1-naphtylamine and caffeic acid on the growth of Microcystis aeruginosa. Allelopath. J. 2015, 35, 249–258. [Google Scholar]
- Chotsaeng, N.; Laosinwattana, C.; Charoenying, P. Herbicidal activities of some allelochemicals and their synergistic behaviors toward Amaranthus tricolor L. Molecules 2017, 22, 1841. [Google Scholar] [CrossRef] [Green Version]
- Claeys, H.; Van Landeghem, S.; Dubois, M.; Maleux, K.; Inzé, D. What Is Stress? Dose-response effects in commonly used in vitro stress assays. Plant Physiol. 2014, 165, 519–527. [Google Scholar] [CrossRef] [Green Version]
- Fincheira, P.; Parada, M.; Quiroz, A. Volatile organic compounds stimulate plant growing and seed germination of Lactuca sativa. J. Soil Sci. Plant Nutr. 2017, 17, 853–867. [Google Scholar] [CrossRef] [Green Version]
- Chen, D.; Zhou, Y.; Wang, M.; Mujtaba Munir, M.A.; Lian, J.; Yu, S.; Dai, K.; Yang, X. Succession pattern in soil micro-ecology under tobacco (Nicotiana tabacum L.) continuous cropping circumstances in Yunnan province of southwest China. Front. Microbiol. 2021, 12, 785110. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| M1 | M2 | M3 | M4 | M5 | M6 | M7 | M8 | M9 | M10 | ||
|---|---|---|---|---|---|---|---|---|---|---|---|
| OB | Pearson Correlation | −0.817 * | 0.271 | −0.350 | 0.500 | 0.354 | −0.008 | 0.410 | 0.465 | 0.383 | −0.324 |
| p-value | 0.001 | 0.394 | 0.265 | 0.098 | 0.258 | 0.981 | 0.185 | 0.128 | 0.219 | 0.304 | |
| Model | Unstandardized Coefficients | Standardized Coefficients | t | Sig. | Collinearity Statistics | |||
|---|---|---|---|---|---|---|---|---|
| B | Std. Error | Beta | Tolerance | VIF | ||||
| 1 | (Constant) | −0.127 | 0.141 | −0.900 | 0.389 | |||
| MPC1 | 1.219 | 0.248 | 0.841 | 4.909 | 0.001 | 1.000 | 1.000 | |
| 2 | (Constant) | 0.378 | 0.233 | 1.624 | 0.139 | |||
| MPC1 | 0.772 | 0.270 | 0.533 | 2.865 | 0.019 | 0.558 | 1.793 | |
| M1 | −0.690 | 0.277 | −0.463 | −2.491 | 0.034 | 0.558 | 1.793 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, F.; Yan, Y.; Dong, Q.; Jiang, C.; Zu, C.; Shen, J. The Changes in Rhizosphere Metabolome and Microbiota Are the Main Direct Obstacles to Continuous Cropping in Tobacco (Nicotiana tabacum L.). Agronomy 2023, 13, 964. https://doi.org/10.3390/agronomy13040964
Yu F, Yan Y, Dong Q, Jiang C, Zu C, Shen J. The Changes in Rhizosphere Metabolome and Microbiota Are the Main Direct Obstacles to Continuous Cropping in Tobacco (Nicotiana tabacum L.). Agronomy. 2023; 13(4):964. https://doi.org/10.3390/agronomy13040964
Chicago/Turabian StyleYu, Fei, Yifeng Yan, Qing Dong, Chaoqiang Jiang, Chaolong Zu, and Jia Shen. 2023. "The Changes in Rhizosphere Metabolome and Microbiota Are the Main Direct Obstacles to Continuous Cropping in Tobacco (Nicotiana tabacum L.)" Agronomy 13, no. 4: 964. https://doi.org/10.3390/agronomy13040964