
Transgene-Free Genome Editing for Biotic and Abiotic Stress Resistance in Sugarcane: Prospects and Challenges
 , , , ,
, , , ,
Abstract
:1. Introduction
2. Biotic Stresses Affecting Sugarcane
2.1. Major Diseases
2.2. Major Insect Pests
3. Abiotic Stress in Sugarcane
3.1. Drought
3.2. Salt
3.3. Cold and Heat
3.4. Waterlogging
3.5. Heavy Metals
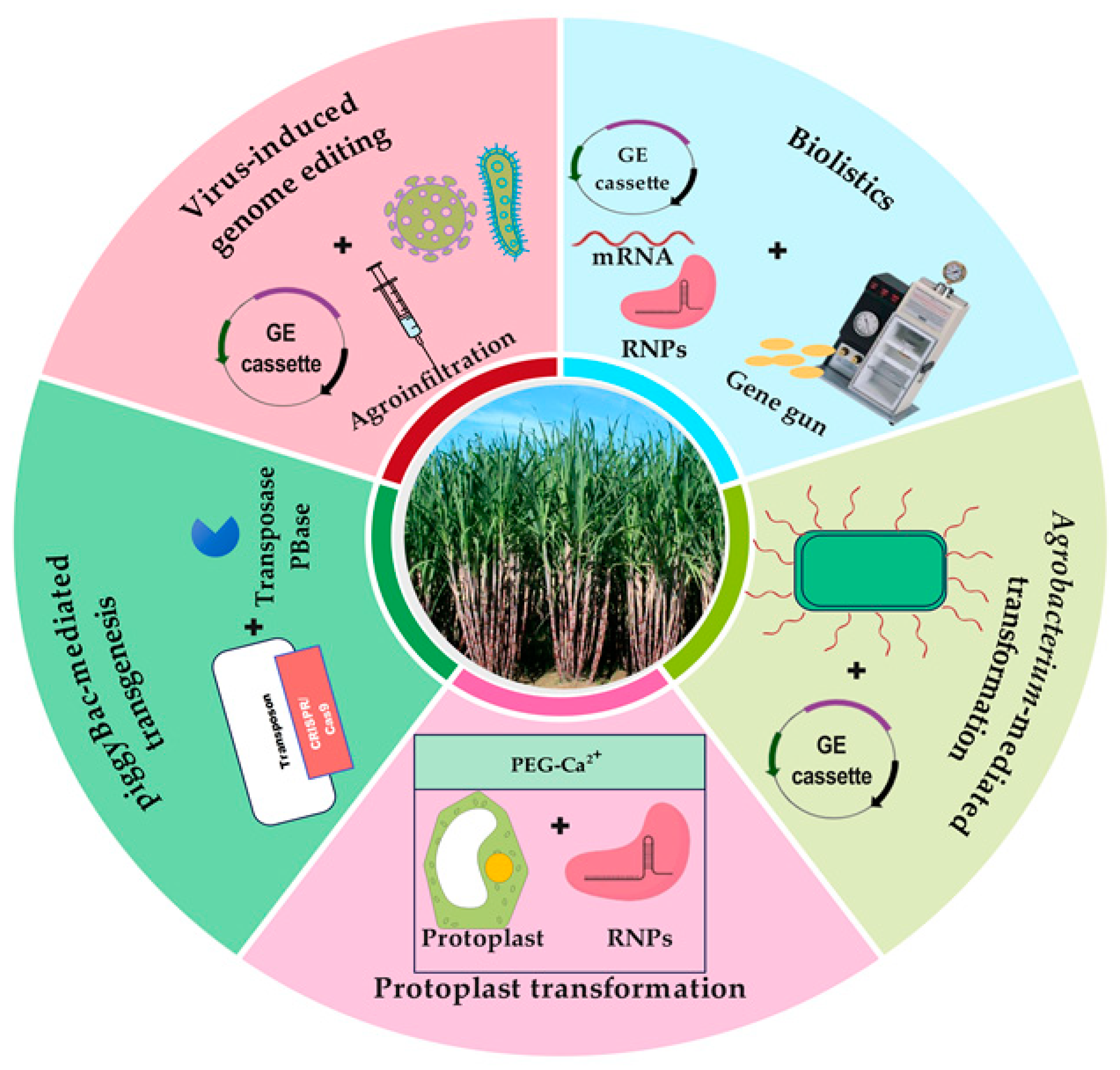
4. Transgene-Free Genome Editing: Approaches and Applications
4.1. Virus-Induced Genome Editing
4.2. Particle Bombardment/Biolistics
4.3. Agrobacterium-Mediated Transformation
4.4. Protoplast Transformation
4.5. piggyBac-Mediated Transgenesis
4.6. Grafting and Mobile RNA-Mediated Genome Editing
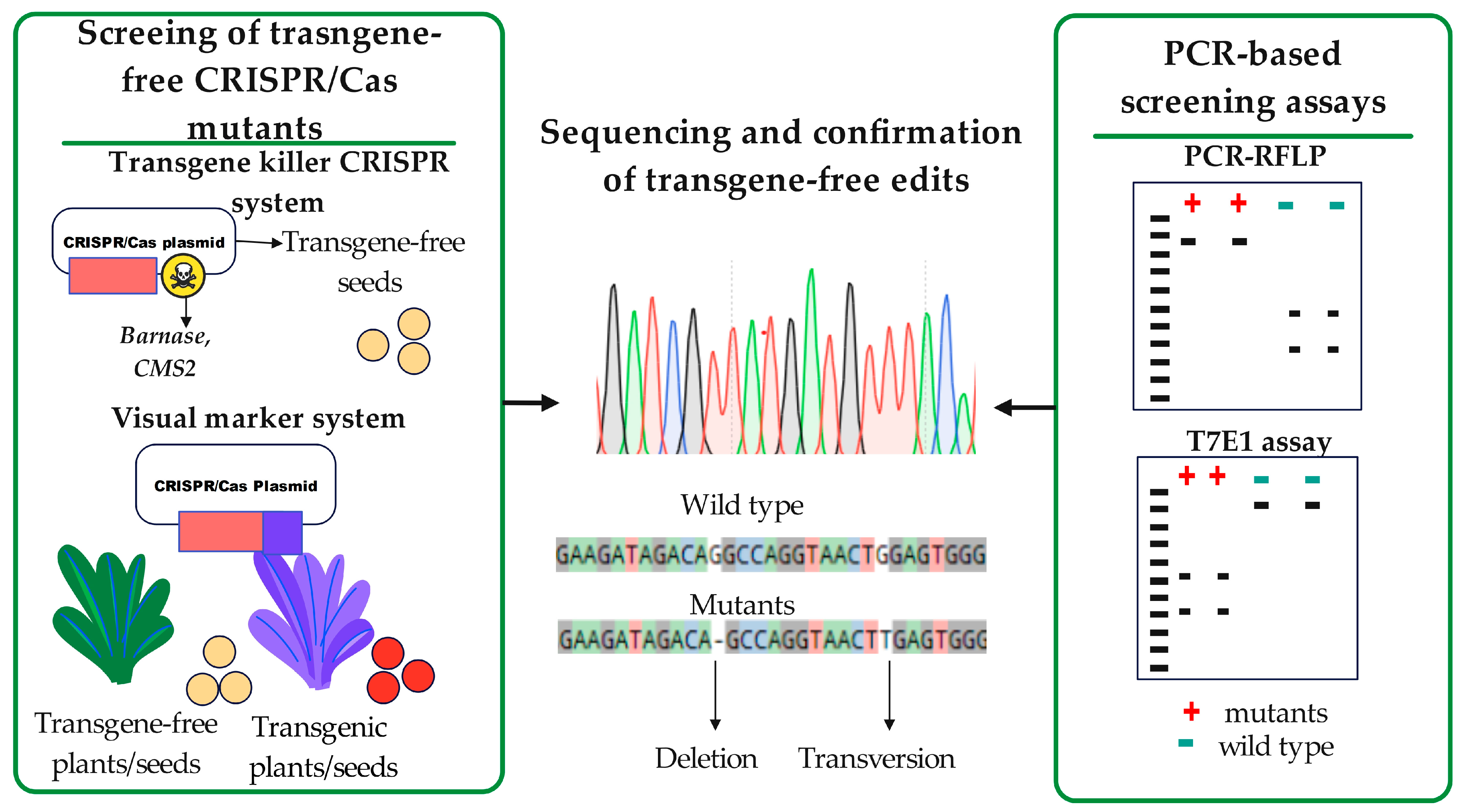
5. Screening of Transgene-Free GE Lines
6. Candidate Genes for Genome Editing in Sugarcane for Biotic and Abiotic Stress—Lessons from Crop Plants
7. Regulatory Issues in Commercializing GE Plants
8. Challenges in Transgene-Free Genome Editing in Sugarcane
9. Conclusions and Future Prospects
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Solomon, S. Sugarcane by-products based industries in India. Sugar Tech 2011, 13, 408–416. [Google Scholar] [CrossRef]
- Statista. Available online: https://www.statista.com/statistics/495973/sugar-production-worldwide/ (accessed on 10 February 2023).
- UNFPA. Available online: https://pdp.unfpa.org/apps/0aeda6af00dd4544ba50452da2dda474/explore (accessed on 10 February 2023).
- Viswanathan, R.; Rao, G. Disease scenario and management of major sugarcane diseases in India. Sugar Tech 2011, 13, 336–353. [Google Scholar] [CrossRef]
- Geetha, M.; Kalyanasundaram, M.; Jayaraj, J.; Shanthi, M.; Vijayashanthi, V.; Hemalatha, D.; Karthickraja, K. Pests of sugarcane. In Pests and Their Management; Omkar, Ed.; Springer: Singapore, 2018; pp. 241–310. [Google Scholar]
- Suganthi, M.; Muthukrishnan, P.; Chinnusamy, C. Crop weed competition in sugarcane-a review. Agric. Rev. 2019, 40, 239–242. [Google Scholar] [CrossRef]
- Lingle, S.E.; Wiegand, C.L. Soil salinity and sugarcane juice quality. Field Crops Res. 1997, 54, 259–268. [Google Scholar] [CrossRef]
- Rietz, D.; Haynes, R. Effect of irrigation-induced salinity and sodicity on sugarcane yield. Proc. S. Afr. Sugar Technol. Assoc. 2002, 76, 173–185. [Google Scholar]
- Jain, R.; Shrivastava, A.; Solomon, S.; Yadav, R. Low temperature stress-induced biochemical changes affect stubble bud sprouting in sugarcane (Saccharum spp. hybrid). Plant Growth Regul. 2007, 53, 17–23. [Google Scholar] [CrossRef]
- Lakshmanan, P.; Robinson, N. Stress physiology: Abiotic stresses. In Sugarcane: Physiology, Biochemistry, and Functional Biology; Moore, P., Botha, F., Eds.; John Wiley & Sons Ltd.: Chichester, UK, 2013; pp. 411–434. [Google Scholar]
- Budeguer, F.; Enrique, R.; Perera, M.F.; Racedo, J.; Castagnaro, A.P.; Noguera, A.S.; Welin, B. Genetic transformation of sugarcane, current status and future prospects. Front. Plant Sci. 2021, 12, 2467. [Google Scholar] [CrossRef]
- Abdallah, N.A.; Prakash, C.S.; McHughen, A.G. Genome editing for crop improvement: Challenges and opportunities. GM Crops Food 2015, 6, 183–205. [Google Scholar] [CrossRef]
- Wang, C.; Shen, L.; Fu, Y.; Yan, C.; Wang, K. A simple CRISPR/Cas9 system for multiplex genome editing in rice. J. Genet. Genom. 2015, 42, 703–706. [Google Scholar] [CrossRef]
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A programmable dual-RNA–guided DNA endonuclease in adaptive bacterial immunity. Science 2012, 337, 816–821. [Google Scholar] [CrossRef]
- Jiang, W.; Yang, B.; Weeks, D.P. Efficient CRISPR/Cas9-mediated gene editing in Arabidopsis thaliana and inheritance of modified genes in the T2 and T3 generations. PLoS ONE 2014, 9, e99225. [Google Scholar] [CrossRef] [PubMed]
- Chapman, J.R.; Taylor, M.R.; Boulton, S.J. Playing the end game: DNA double-strand break repair pathway choice. Mol. Cell 2012, 47, 497–510. [Google Scholar] [CrossRef]
- Rodgers, K.; McVey, M. Error-prone repair of DNA double-strand breaks. J. Cell. Physiol. 2016, 231, 15–24. [Google Scholar] [CrossRef]
- Pannunzio, N.R.; Watanabe, G.; Lieber, M.R. Nonhomologous DNA end-joining for repair of DNA double-strand breaks. J. Biol. Chem. 2018, 293, 10512–10523. [Google Scholar] [CrossRef]
- Viswanathan, R. Fusarium diseases affecting sugarcane production in India. Indian Phytopathol. 2020, 73, 415–424. [Google Scholar] [CrossRef]
- Bhuiyan, S.A.; Magarey, R.C.; McNeil, M.D.; Aitken, K.S. Sugarcane smut, caused by Sporisorium scitamineum, a major disease of sugarcane: A contemporary review. Phytopathology 2021, 111, 1905–1917. [Google Scholar] [CrossRef]
- Comstock, J. Ratoon stunting disease. Sugar Tech 2002, 4, 1–6. [Google Scholar] [CrossRef]
- Hoy, J. Sugarcane leaf scald distribution, symptomatology. Plant Dis. 1994, 78, 1083–1087. [Google Scholar] [CrossRef]
- Lin, L.-H.; Ntambo, M.S.; Rott, P.C.; Wang, Q.-N.; Lin, Y.-H.; Fu, H.-Y.; Gao, S.-J. Molecular detection and prevalence of Xanthomonas albilineans, the causal agent of sugarcane leaf scald, in China. Crop Protect. 2018, 109, 17–23. [Google Scholar] [CrossRef]
- Viswanathan, R.; Chinnaraja, C.; Karuppaiah, R.; Ganesh Kumar, V.; Jenshi Rooba, J.; Malathi, P. Genetic diversity of sugarcane grassy shoot (SCGS)-phytoplasmas causing grassy shoot disease in India. Sugar Tech 2011, 13, 220–228. [Google Scholar] [CrossRef]
- Fitch, M.; Lehrer, A.T.; Komor, E.; Moore, P.H. Elimination of Sugarcane yellow leaf virus from infected sugarcane plants by meristem tip culture visualized by tissue blot immunoassay. Plant Pathol. 2001, 50, 676–680. [Google Scholar] [CrossRef]
- Viswanathan, R.; Chinnaraja, C.; Malathi, P.; Gomathi, R.; Rakkiyappan, P.; Neelamathi, D.; Ravichandran, V. Impact of Sugarcane yellow leaf virus (ScYLV) infection on physiological efficiency and growth parameters of sugarcane under tropical climatic conditions in India. Acta Physiol. Plant 2014, 36, 1805–1822. [Google Scholar] [CrossRef]
- Chinnaraja, C.; Viswanathan, R. Variability in yellow leaf symptom expression caused by the Sugarcane yellow leaf virus and its seasonal influence in sugarcane. Phytoparasitica 2015, 43, 339–353. [Google Scholar] [CrossRef]
- Comstock, J. Yellow leaf syndrome appears on the United States mainland. Sugar J. 1994, 56, 33–35. [Google Scholar]
- Viswanathan, R.; Balamuralikrishnan, M. Impact of mosaic infection on growth and yield of sugarcane. Sugar Tech 2005, 7, 61–65. [Google Scholar] [CrossRef]
- Deshumkh, G.; Sawant, D. Studies on Symptomatology of Sorghum and Sugarcane Mosaic Virus. J. Plant Dis. Sci. 2008, 3, 116–117. [Google Scholar]
- Lu, G.; Wang, Z.; Xu, F.; Pan, Y.-B.; Grisham, M.P.; Xu, L. Sugarcane mosaic disease: Characteristics, identification and control. Microorganisms 2021, 9, 1984. [Google Scholar] [CrossRef] [PubMed]
- Avasthy, P.; Tiwari, N. (Eds.) The Shoot Borer, Chilo Infuscatellus Snellen; Sugarcane Breeding Institute: Coimbatore, India, 1986; pp. 69–92.
- Srikanth, J.; Salin, K.; Jayanthi, R. Sugarcane Pests and Their Management; Sugarcane Breeding Institute: Coimbatore, India, 2012.
- Easwaramoorthy, S.; Nandagopal, V. Life tables of internode borer, Chilo sacchariphagus indicus (K.), on resistant and susceptible varieties of sugarcane. Int. J. Pest Manag. 1986, 32, 221–228. [Google Scholar] [CrossRef]
- Mukunthan, N. New aspects in the biology of sugarcane top borer, Scirpophaga (Nivella F.) excerptalis Wlk. Entomon 1985, 10, 235–238. [Google Scholar]
- Sankar, M.; Jaigeetha, S.; Rao, M.S. Exploitation of biocontrol agents, Trichogramma chilonis and Tetrastichus howardi on yield improvement in sugarcane at EID Parry (India) Ltd., Sugar Mill Command Areas. Int. J. Innov. Res. Dev. 2014, 3, 314–318. [Google Scholar]
- Bhavani, B.; Rao, C.V.N. Management of sugarcane white fly (Aleurolobus barodensis Mask.) in North coastal districts of Andhra Pradesh, India. Int. J. Social Sci. Interdis. Res. 2013, 2, 112–115. [Google Scholar]
- Askarianzadeh, A.; Minaeimoghadam, M. Biology, natural enemies and damage of the sugarcane whitefly (Neomaskellia andropogonis)(Homoptera: Aleyrodidae) in Iran. Int. J. Trop. Insect Sci. 2018, 38, 381–386. [Google Scholar] [CrossRef]
- Sosa Jr, O. Effect of white grub (Coleoptera: Scarabaeidae) infestation on sugarcane yields. J. Econ. Entomol. 1984, 77, 183–185. [Google Scholar] [CrossRef]
- Liu, X.; Zhu, X.; Pan, Y.; Li, S.; Liu, Y.; Ma, Y. Agricultural drought monitoring: Progress, challenges, and prospects. J. Geogr. Sci. 2016, 26, 750–767. [Google Scholar] [CrossRef]
- Mall, A.; Misra, V.; Pathak, A.; Srivastava, S. Breeding for Drought Tolerance in Sugarcane: Indian Perspective. Sugar Tech 2022, 24, 1625–1635. [Google Scholar] [CrossRef]
- Narwade, A.V.; Bhagat, K.; Patil, D.; Singh, Y.; Kumari, A.; Ban, Y.; Thakare, H.; Singh, C. Abiotic stress responses in sugarcane. In Challenges and Prospective of Plant Abiotic Stress; Pasala, R., Ed.; Today & Tomorrow’s Printers and Publishers: Delhi, India, 2016; pp. 419–446. [Google Scholar]
- Ferreira, T.H.; Tsunada, M.S.; Bassi, D.; Araújo, P.; Mattiello, L.; Guidelli, G.V.; Righetto, G.L.; Gonçalves, V.R.; Lakshmanan, P.; Menossi, M. Sugarcane water stress tolerance mechanisms and its implications on developing biotechnology solutions. Front. Plant Sci. 2017, 8, 1077. [Google Scholar] [CrossRef]
- Inman-Bamber, N.; Lakshmanan, P.; Park, S. Sugarcane for water-limited environments: Theoretical assessment of suitable traits. Field Crops Res. 2012, 134, 95–104. [Google Scholar] [CrossRef]
- Sage, R.F.; Peixoto, M.M.; Sage, T.L. Photosynthesis in sugarcane. In Sugarcane: Physiology, Biochemistry, and Functional Biology; Moore, P., Botha, F., Eds.; John Wiley & Sons Ltd.: Chichester, UK, 2013; pp. 121–154. [Google Scholar]
- Ogbaga, C.C.; Amir, M.; Bano, H.; Chater, C.C.; Jellason, N.P. Clarity on frequently asked questions about drought measurements in plant physiology. Sci. Afr. 2020, 8, e00405. [Google Scholar] [CrossRef]
- Rai, M.K.; Kalia, R.K.; Singh, R.; Gangola, M.P.; Dhawan, A. Developing stress tolerant plants through in vitro selection—an overview of the recent progress. Environ. Exp. Bot. 2011, 71, 89–98. [Google Scholar] [CrossRef]
- Huang, D.; Wu, W.; Abrams, S.R.; Cutler, A.J. The relationship of drought-related gene expression in Arabidopsis thaliana to hormonal and environmental factors. J. Exp. Bot. 2008, 59, 2991–3007. [Google Scholar] [CrossRef]
- Azevedo, R.A.; Carvalho, R.F.; Cia, M.C.; Gratão, P.L. Sugarcane under pressure: An overview of biochemical and physiological studies of abiotic stress. Trop. Plant Biol. 2011, 4, 42–51. [Google Scholar] [CrossRef]
- Patade, V.Y.; Suprasanna, P.; Bapat, V.A. Effects of salt stress in relation to osmotic adjustment on sugarcane (Saccharum officinarum L.) callus cultures. Plant Growth Regul. 2008, 55, 169–173. [Google Scholar] [CrossRef]
- Sengar, K.; Sengar, R.; Singh, A. Biotechnological and genomic analysis for salinity tolerance in sugarcane. Int. J. Biotechnol. Bioeng. Res. 2013, 4, 407–414. [Google Scholar]
- Polash, M.A.S.; Sakil, M.A.; Hossain, M.A. Plants responses and their physiological and biochemical defense mechanisms against salinity: A review. Trop. Plant Res. 2019, 6, 250–274. [Google Scholar] [CrossRef]
- Meena, M.R.; Kumar, R.; Chinnaswamy, A.; Karuppaiyan, R.; Kulshreshtha, N.; Ram, B. Current breeding and genomic approaches to enhance the cane and sugar productivity under abiotic stress conditions. 3 Biotech 2020, 10, 440. [Google Scholar] [CrossRef]
- Das, K.; Roychoudhury, A. Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Front. Environ. Sci. 2014, 2, 53. [Google Scholar] [CrossRef]
- Rehman, S.; Muhammad, K.; Novaes, E.; Que, Y.; Din, A.; Islam, M.; Porto, A.; Sajid, M.; Ullah, N.; Iqsa, S. Expression analysis of transcription factors in sugarcane during cold stress. Braz. J. Biol. 2021, 83, e242603. [Google Scholar] [CrossRef]
- Hussain, S.; Khaliq, A.; Mehmood, U.; Qadir, T.; Saqib, M.; Iqbal, M.A.; Hussain, S. Sugarcane production under changing climate: Effects of environmental vulnerabilities on sugarcane diseases, insects and weeds. In Sugarcane Production-Agronomic, Scientific and Industrial Perspectives; IntechOpen: Rijeka, Croatia, 2018; pp. 1–17. [Google Scholar]
- Gomathi, R.; Chandran, K.; Gururaja Rao, P.; Rakkiyappan, P. Effect of Waterlogging in Sugarcane and Its Management; Sugarcane Breeding Institute: Coimbatore, India, 2010.
- Gomathi, R.; Gururaja Rao, P.; Chandran, K.; Selvi, A. Adaptive responses of sugarcane to waterlogging stress: An over view. Sugar Tech 2015, 17, 325–338. [Google Scholar] [CrossRef]
- Riyazuddin, R.; Nisha, N.; Ejaz, B.; Khan, M.I.R.; Kumar, M.; Ramteke, P.W.; Gupta, R. A comprehensive review on the heavy metal toxicity and sequestration in plants. Biomolecules 2022, 12, 43. [Google Scholar] [CrossRef]
- Kumar, A.; Aery, N. Impact, metabolism, and toxicity of heavy metals in plants. In Plant Responses to Xenobiotics; Singh, A., Prasad, S.M., Singh, R.P., Eds.; Springer: Singapore, 2016; pp. 141–176. [Google Scholar]
- Tiwari, S.; Lata, C. Heavy metal stress, signaling, and tolerance due to plant-associated microbes: An overview. Front. Plant Sci. 2018, 9, 452. [Google Scholar] [CrossRef] [PubMed]
- Baig, M.A.; Qamar, S.; Ali, A.A.; Ahmad, J.; Qureshi, M.I. Heavy metal toxicity and tolerance in crop plants. In Contaminants in Agriculture: Sources, Impacts and Management; Naeem, M., Ansari, A.A., Gill, S.S., Eds.; Springer: Cham, Switzerland, 2020; pp. 201–216. [Google Scholar]
- Hassan, T.U.; Bano, A.; Naz, I. Alleviation of heavy metals toxicity by the application of plant growth promoting rhizobacteria and effects on wheat grown in saline sodic field. Int. J. Phytoremediation 2017, 19, 522–529. [Google Scholar] [CrossRef]
- Panuccio, M.R.; Sorgonà, A.; Rizzo, M.; Cacco, G. Cadmium adsorption on vermiculite, zeolite and pumice: Batch experimental studies. J. Environ. Manag. 2009, 90, 364–374. [Google Scholar] [CrossRef]
- Zhang, C.; Liu, S.; Li, X.; Zhang, R.; Li, J. Virus-induced gene editing and its applications in plants. Int. J. Mol. Sci. 2022, 23, 10202. [Google Scholar] [CrossRef] [PubMed]
- Baltes, N.J.; Gil-Humanes, J.; Cermak, T.; Atkins, P.A.; Voytas, D.F. DNA replicons for plant genome engineering. Plant Cell 2014, 26, 151–163. [Google Scholar] [CrossRef]
- Čermák, T.; Baltes, N.J.; Čegan, R.; Zhang, Y.; Voytas, D.F. High-frequency, precise modification of the tomato genome. Genome Biol. 2015, 16, 232. [Google Scholar] [CrossRef]
- Butler, N.M.; Atkins, P.A.; Voytas, D.F.; Douches, D.S. Generation and inheritance of targeted mutations in potato (Solanum tuberosum L.) using the CRISPR/Cas system. PLoS ONE 2015, 10, e0144591. [Google Scholar] [CrossRef]
- Dahan-Meir, T.; Filler-Hayut, S.; Melamed-Bessudo, C.; Bocobza, S.; Czosnek, H.; Aharoni, A.; Levy, A.A. Efficient in planta gene targeting in tomato using geminiviral replicons and the CRISPR/Cas9 system. Plant J. 2018, 95, 5–16. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Lu, Y.; Botella, J.R.; Mao, Y.; Hua, K.; Zhu, J.-k. Gene targeting by homology-directed repair in rice using a geminivirus-based CRISPR/Cas9 system. Mol Plant. 2017, 10, 1007–1010. [Google Scholar] [CrossRef]
- Gil-Humanes, J.; Wang, Y.; Liang, Z.; Shan, Q.; Ozuna, C.V.; Sánchez-León, S.; Baltes, N.J.; Starker, C.; Barro, F.; Gao, C. High-efficiency gene targeting in hexaploid wheat using DNA replicons and CRISPR/Cas9. Plant J. 2017, 89, 1251–1262. [Google Scholar] [CrossRef] [PubMed]
- Yin, K.; Han, T.; Liu, G.; Chen, T.; Wang, Y.; Yu, A.Y.L.; Liu, Y. A geminivirus-based guide RNA delivery system for CRISPR/Cas9 mediated plant genome editing. Sci. Rep. 2015, 5, 14926. [Google Scholar] [CrossRef]
- Lei, J.; Dai, P.; Li, Y.; Zhang, W.; Zhou, G.; Liu, C.; Liu, X. Heritable gene editing using FT mobile guide RNAs and DNA viruses. Plant Methods 2021, 17, 20. [Google Scholar] [CrossRef]
- Cody, W.B.; Scholthof, H.B.; Mirkov, T.E. Multiplexed gene editing and protein overexpression using a tobacco mosaic virus viral vector. Plant Physiol. 2017, 175, 23–35. [Google Scholar] [CrossRef]
- Hu, J.; Li, S.; Li, Z.; Li, H.; Song, W.; Zhao, H.; Lai, J.; Xia, L.; Li, D.; Zhang, Y. A barley stripe mosaic virus-based guide RNA delivery system for targeted mutagenesis in wheat and maize. Mol. Plant Pathol. 2019, 20, 1463–1474. [Google Scholar] [CrossRef]
- Chiong, K.T.; Cody, W.B.; Scholthof, H.B. RNA silencing suppressor-influenced performance of a virus vector delivering both guide RNA and Cas9 for CRISPR gene editing. Sci. Rep. 2021, 11, 6769. [Google Scholar] [CrossRef]
- Ali, Z.; Eid, A.; Ali, S.; Mahfouz, M.M. Pea early-browning virus-mediated genome editing via the CRISPR/Cas9 system in Nicotiana benthamiana and Arabidopsis. Virus Res. 2018, 244, 333–337. [Google Scholar] [CrossRef]
- Jiang, N.; Zhang, C.; Liu, J.Y.; Guo, Z.H.; Zhang, Z.Y.; Han, C.G.; Wang, Y. Development of Beet necrotic yellow vein virus-based vectors for multiple-gene expression and guide RNA delivery in plant genome editing. Plant Biotechnol. J. 2019, 17, 1302–1315. [Google Scholar] [CrossRef]
- Mei, Y.; Beernink, B.M.; Ellison, E.E.; Konečná, E.; Neelakandan, A.K.; Voytas, D.F.; Whitham, S.A. Protein expression and gene editing in monocots using foxtail mosaic virus vectors. Plant Direct 2019, 3, e00181. [Google Scholar] [CrossRef]
- Zhang, X.; Kang, L.; Zhang, Q.; Meng, Q.; Pan, Y.; Yu, Z.; Shi, N.; Jackson, S.; Zhang, X.; Wang, H. An RNAi suppressor activates in planta virus–mediated gene editing. Funct. Integr. Genom. 2020, 20, 471–477. [Google Scholar] [CrossRef]
- Ariga, H.; Toki, S.; Ishibashi, K. Potato virus X vector-mediated DNA-free genome editing in plants. Plant Cell Physiol. 2020, 61, 1946–1953. [Google Scholar] [CrossRef]
- Uranga, M.; Aragonés, V.; Selma, S.; Vázquez-Vilar, M.; Orzáez, D.; Daròs, J.A. Efficient Cas9 multiplex editing using unspaced sgRNA arrays engineering in a Potato virus X vector. Plant J. 2021, 106, 555–565. [Google Scholar] [CrossRef]
- Uranga, M.; Vazquez-Vilar, M.; Orzáez, D.; Daròs, J.-A. CRISPR-Cas12a genome editing at the whole-plant level using two compatible RNA virus vectors. CRISPR J. 2021, 4, 761–769. [Google Scholar] [CrossRef]
- Calvache, C.; Vazquez-Vilar, M.; Selma, S.; Uranga, M.; Fernández-del-Carmen, A.; Daròs, J.A.; Orzáez, D. Strong and tunable anti-CRISPR/Cas activities in plants. Plant Biotechnol. J. 2022, 20, 399–408. [Google Scholar] [CrossRef]
- Gao, Q.; Xu, W.Y.; Yan, T.; Fang, X.D.; Cao, Q.; Zhang, Z.J.; Ding, Z.H.; Wang, Y.; Wang, X.B. Rescue of a plant cytorhabdovirus as versatile expression platforms for planthopper and cereal genomic studies. New Phytol. 2019, 223, 2120–2133. [Google Scholar] [CrossRef]
- Ma, X.; Zhang, X.; Liu, H.; Li, Z. Highly efficient DNA-free plant genome editing using virally delivered CRISPR–Cas9. Nat. Plants 2020, 6, 773–779. [Google Scholar] [CrossRef]
- Liu, J.-J.; Orlova, N.; Oakes, B.L.; Ma, E.; Spinner, H.B.; Baney, K.L.; Chuck, J.; Tan, D.; Knott, G.J.; Harrington, L.B. CasX enzymes comprise a distinct family of RNA-guided genome editors. Nature 2019, 566, 218–223. [Google Scholar] [CrossRef]
- Pausch, P.; Al-Shayeb, B.; Bisom-Rapp, E.; Tsuchida, C.A.; Li, Z.; Cress, B.F.; Knott, G.J.; Jacobsen, S.E.; Banfield, J.F.; Doudna, J.A. CRISPR-CasΦ from huge phages is a hypercompact genome editor. Science 2020, 369, 333–337. [Google Scholar] [CrossRef]
- Uranga, M.; Daròs, J.A. Tools and targets: The dual role of plant viruses in CRISPR–Cas genome editing. Plant Genome 2022, e20220. [Google Scholar] [CrossRef]
- Pereira, A.R.; Barbieri, V.; Nova, N.A.V. Climatic conditioning of flowering induction in sugarcane. Agric. Meteorol. 1983, 29, 103–110. [Google Scholar] [CrossRef]
- Moore, P.H.; Nuss, K. Flowering and flower synchronization. In Sugarcane Improvement Through Breeding; Heinz, D.J., Ed.; Elsevier Science: Amsterdam, The Netherlands, 1987; Volume 11, pp. 273–311. [Google Scholar]
- Ellison, E.E.; Nagalakshmi, U.; Gamo, M.E.; Huang, P.-j.; Dinesh-Kumar, S.; Voytas, D.F. Multiplexed heritable gene editing using RNA viruses and mobile single guide RNAs. Nat. Plants 2020, 6, 620–624. [Google Scholar] [CrossRef]
- Mohan, C.; Ashwin Narayan, J.; Esterling, M.; Yau, Y.-Y. Current transformation methods for genome–editing applications in energy crop sugarcane. In Climate Change, Photosynthesis and Advanced Biofuels: The Role of Biotechnology in the Production of Value-added Plant Bio-products; Kumar, A., Yau, Y., Ogita, S., Scheibe, R., Eds.; Springer: Singapore, 2020; pp. 369–388. [Google Scholar]
- Laforest, L.C.; Nadakuduti, S.S. Advances in delivery mechanisms of CRISPR gene-editing reagents in plants. Front. Genome Ed. 2022, 4, 1. [Google Scholar] [CrossRef]
- Tsanova, T.; Stefanova, L.; Topalova, L.; Atanasov, A.; Pantchev, I. DNA-free gene editing in plants: A brief overview. Biotechnol. Biotechnol. Equip. 2021, 35, 131–138. [Google Scholar] [CrossRef]
- Svitashev, S.; Schwartz, C.; Lenderts, B.; Young, J.; Cigan, M. Genome editing in maize directed by CRISPR-Cas9 ribonucleoprotein complexes. Nat. Commun. 2016, 7, 13274. [Google Scholar] [CrossRef]
- Banakar, R.; Eggenberger, A.L.; Lee, K.; Wright, D.A.; Murugan, K.; Zarecor, S.; Lawrence-Dill, C.J.; Sashital, D.G.; Wang, K. High-frequency random DNA insertions upon co-delivery of CRISPR-Cas9 ribonucleoprotein and selectable marker plasmid in rice. Sci. Rep. 2019, 9, 19902. [Google Scholar] [CrossRef] [PubMed]
- Banakar, R.; Schubert, M.; Collingwood, M.; Vakulskas, C.; Eggenberger, A.L.; Wang, K. Comparison of CRISPR-Cas9/Cas12a ribonucleoprotein complexes for genome editing efficiency in the rice phytoene desaturase (OsPDS) gene. Rice 2020, 13, 4. [Google Scholar] [CrossRef] [PubMed]
- Liang, Z.; Chen, K.; Zhang, Y.; Liu, J.; Yin, K.; Qiu, J.-L.; Gao, C. Genome editing of bread wheat using biolistic delivery of CRISPR/Cas9 in vitro transcripts or ribonucleoproteins. Nat. Protoc. 2018, 13, 413–430. [Google Scholar] [CrossRef] [PubMed]
- Oz, M.T.; Altpeter, A.; Karan, R.; Merotto, A.; Altpeter, F. CRISPR/Cas9-mediated multi-allelic gene targeting in sugarcane confers herbicide tolerance. Front. Genome Ed. 2021, 3, 673566. [Google Scholar] [CrossRef]
- Eid, A.; Mohan, C.; Sanchez, S.; Wang, D.; Altpeter, F. Multiallelic, targeted mutagenesis of magnesium chelatase with CRISPR/Cas9 provides a rapidly scorable phenotype in highly polyploid sugarcane. Front. Genome Ed. 2021, 3, 654996. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Xing, J.; Mi, Q.; Yang, W.; Xiang, H.; Xu, L.; Zeng, W.; Wang, J.; Deng, L.; Jiang, J. Highly efficient transgene-free genome editing in tobacco using an optimized CRISPR/Cas9 system, pOREU3TR. Plant Sci. 2023, 326, 111523. [Google Scholar] [CrossRef]
- Chen, L.; Li, W.; Katin-Grazzini, L.; Ding, J.; Gu, X.; Li, Y.; Gu, T.; Wang, R.; Lin, X.; Deng, Z. A method for the production and expedient screening of CRISPR/Cas9-mediated non-transgenic mutant plants. Hortic. Res. 2018, 5, 13. [Google Scholar] [CrossRef]
- Veillet, F.; Perrot, L.; Chauvin, L.; Kermarrec, M.-P.; Guyon-Debast, A.; Chauvin, J.-E.; Nogué, F.; Mazier, M. Transgene-free genome editing in tomato and potato plants using agrobacterium-mediated delivery of a CRISPR/Cas9 cytidine base editor. Int. J. Mol. Sci. 2019, 20, 402. [Google Scholar] [CrossRef] [PubMed]
- Danilo, B.; Perrot, L.; Mara, K.; Botton, E.; Nogué, F.; Mazier, M. Efficient and transgene-free gene targeting using Agrobacterium-mediated delivery of the CRISPR/Cas9 system in tomato. Plant Cell Rep. 2019, 38, 459–462. [Google Scholar] [CrossRef] [PubMed]
- Bánfalvi, Z.; Csákvári, E.; Villányi, V.; Kondrák, M. Generation of transgene-free PDS mutants in potato by Agrobacterium-mediated transformation. BMC Biotechnol. 2020, 20, 25. [Google Scholar] [CrossRef] [PubMed]
- Mayavan, S.; Subramanyam, K.; Arun, M.; Rajesh, M.; Kapil Dev, G.; Sivanandhan, G.; Jaganath, B.; Manickavasagam, M.; Selvaraj, N.; Ganapathi, A. Agrobacterium tumefaciens-mediated in planta seed transformation strategy in sugarcane. Plant Cell Rep. 2013, 32, 1557–1574. [Google Scholar] [CrossRef] [PubMed]
- Mayavan, S.; Subramanyam, K.; Jaganath, B.; Sathish, D.; Manickavasagam, M.; Ganapathi, A. Agrobacterium-mediated in planta genetic transformation of sugarcane setts. Plant Cell Rep. 2015, 34, 1835–1848. [Google Scholar] [CrossRef]
- Murovec, J.; Guček, K.; Bohanec, B.; Avbelj, M.; Jerala, R. DNA-free genome editing of Brassica oleracea and B. rapa protoplasts using CRISPR-Cas9 ribonucleoprotein complexes. Front. Plant Sci. 2018, 9, 1594. [Google Scholar] [CrossRef]
- Liu, W.; Rudis, M.R.; Cheplick, M.H.; Millwood, R.J.; Yang, J.-P.; Ondzighi-Assoume, C.A.; Montgomery, G.A.; Burris, K.P.; Mazarei, M.; Chesnut, J.D. Lipofection-mediated genome editing using DNA-free delivery of the Cas9/gRNA ribonucleoprotein into plant cells. Plant Cell Rep. 2020, 39, 245–257. [Google Scholar] [CrossRef]
- Sant’Ana, R.R.A.; Caprestano, C.A.; Nodari, R.O.; Agapito-Tenfen, S.Z. PEG-delivered CRISPR-Cas9 ribonucleoproteins system for gene-editing screening of maize protoplasts. Genes 2020, 11, 1029. [Google Scholar] [CrossRef]
- Toda, E.; Koiso, N.; Takebayashi, A.; Ichikawa, M.; Kiba, T.; Osakabe, K.; Osakabe, Y.; Sakakibara, H.; Kato, N.; Okamoto, T. An efficient DNA-and selectable-marker-free genome-editing system using zygotes in rice. Nat. Plants 2019, 5, 363–368. [Google Scholar] [CrossRef]
- Wu, S.; Zhu, H.; Liu, J.; Yang, Q.; Shao, X.; Bi, F.; Hu, C.; Huo, H.; Chen, K.; Yi, G. Establishment of a PEG-mediated protoplast transformation system based on DNA and CRISPR/Cas9 ribonucleoprotein complexes for banana. BMC Plant Biol. 2020, 20, 425. [Google Scholar] [CrossRef]
- Sidorov, V.; Wang, D.; Nagy, E.D.; Armstrong, C.; Beach, S.; Zhang, Y.; Groat, J.; Yang, S.; Yang, P.; Gilbertson, L. Heritable DNA-free genome editing of canola (Brassica napus L.) using PEG-mediated transfection of isolated protoplasts. In Vitro Cell. Dev. Biol. Plant 2022, 58, 447–456. [Google Scholar] [CrossRef]
- Lin, C.-S.; Hsu, C.-T.; Yuan, Y.-H.; Zheng, P.-X.; Wu, F.-H.; Cheng, Q.-W.; Wu, Y.-L.; Wu, T.-L.; Lin, S.; Yue, J.-J. DNA-free CRISPR-Cas9 gene editing of wild tetraploid tomato Solanum peruvianum using protoplast regeneration. Plant Physiol. 2022, 188, 1917–1930. [Google Scholar] [CrossRef] [PubMed]
- Andersson, M.; Turesson, H.; Olsson, N.; Fält, A.S.; Ohlsson, P.; Gonzalez, M.N.; Samuelsson, M.; Hofvander, P. Genome editing in potato via CRISPR-Cas9 ribonucleoprotein delivery. Physiol. Plant. 2018, 164, 378–384. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Gartland, K.; Davey, M.; Sotak, R.; Gartland, J.; Mulligan, B.; Power, J.; Cocking, E. Transformation of sugarcane protoplasts by direct uptake of a selectable chimaeric gene. Plant Cell Rep. 1987, 6, 297–301. [Google Scholar] [CrossRef]
- Rathus, C.; Birch, R.G. Stable transformation of callus from electroporated sugarcane protoplasts. Plant Sci. 1992, 82, 81–89. [Google Scholar] [CrossRef]
- Chowdhury, M.; Vasil, I.K. Stably transformed herbicide resistant callus of sugarcane via microprojectile bombardment of cell suspension cultures and electroporation of protoplasts. Plant Cell Rep. 1992, 11, 494–498. [Google Scholar] [CrossRef]
- Arencibia, A.; Vázquez, R.I.; Prieto, D.; Téllez, P.; Carmona, E.R.; Coego, A.; Hernández, L.; De la Riva, G.A.; Selman-Housein, G. Transgenic sugarcane plants resistant to stem borer attack. Mol. Breed. 1997, 3, 247–255. [Google Scholar] [CrossRef]
- Wang, Q.; Yu, G.; Chen, Z.; Han, J.; Hu, Y.; Wang, K. Optimization of protoplast isolation, transformation and its application in sugarcane (Saccharum spontaneum L). Crop J. 2021, 9, 133–142. [Google Scholar] [CrossRef]
- Nishizawa-Yokoi, A.; Toki, S. A piggyBac-mediated transgenesis system for the temporary expression of CRISPR/Cas9 in rice. Plant Biotechnol. J. 2021, 19, 1386–1395. [Google Scholar] [CrossRef]
- He, Y.; Mudgett, M.; Zhao, Y. Advances in gene editing without residual transgenes in plants. Plant Physiol. 2022, 188, 1757–1768. [Google Scholar] [CrossRef]
- Yang, L.; Machin, F.; Wang, S.; Saplaoura, E.; Kragler, F. Heritable transgene-free genome editing in plants by grafting of wild-type shoots to transgenic donor rootstocks. Nat. Biotechnol. 2023, 1–10. [Google Scholar] [CrossRef]
- Melnyk, C.W.; Meyerowitz, E.M. Plant grafting. Curr. Biol. 2015, 25, R183–R188. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Zhu, M.; Wang, L.; Wu, J.; Wang, Q.; Wang, R.; Zhao, Y. Programmed self-elimination of the CRISPR/Cas9 construct greatly accelerates the isolation of edited and transgene-free rice plants. Mol Plant. 2018, 11, 1210–1213. [Google Scholar] [CrossRef]
- Yubing, H.; Min, Z.; Lihao, W.; Junhua, W.; Qiaoyan, W.; Rongchen, W.; Yunde, Z. Improvements of TKC technology accelerate isolation of transgene-free CRISPR/Cas9-edited rice plants. Rice Sci. 2019, 26, 109–117. [Google Scholar] [CrossRef]
- Yang, N.; Yan, L.; Zheng, Z.; Zhang, Y.; Zhan, H.; Tian, Y.; Zhang, T.; Li, R.; Gong, X.; Xu, M. Editing gene families by CRISPR/Cas9: Accelerating the isolation of multiple transgene-free null mutant combinations with much reduced labor-intensive analysis. Plant Biotechnol. J. 2022, 20, 241. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Chen, J.; Dai, X.; Zhang, D.; Zhao, Y. An effective strategy for reliably isolating heritable and Cas9-free Arabidopsis mutants generated by CRISPR/Cas9-mediated genome editing. Plant Physiol. 2016, 171, 1794–1800. [Google Scholar] [CrossRef]
- He, Y.; Wang, R.; Dai, X.; Zhao, Y. On improving CRISPR for editing plant genes: Ribozyme-mediated guide RNA production and fluorescence-based technology for isolating transgene-free mutants generated by CRISPR. Prog. Mol. Biol. Transl. Sci. 2017, 149, 151–166. [Google Scholar]
- Aliaga-Franco, N.; Zhang, C.; Presa, S.; Srivastava, A.K.; Granell, A.; Alabadí, D.; Sadanandom, A.; Blázquez, M.A.; Minguet, E.G. Identification of transgene-free CRISPR-edited plants of rice, tomato, and Arabidopsis by monitoring DsRED fluorescence in dry seeds. Front. Plant Sci. 2019, 10, 1150. [Google Scholar] [CrossRef]
- Yan, Y.; Zhu, J.; Qi, X.; Cheng, B.; Liu, C.; Xie, C. Establishment of an efficient seed fluorescence reporter-assisted CRISPR/Cas9 gene editing in maize. J. Integr. Plant Biol. 2021, 63, 1671–1680. [Google Scholar] [CrossRef]
- Wang, J.; Chen, H. A novel CRISPR/Cas9 system for efficiently generating Cas9-free multiplex mutants in Arabidopsis. Abiotech 2020, 1, 6–14. [Google Scholar] [CrossRef]
- Liu, Y.; Zeng, J.; Yuan, C.; Guo, Y.; Yu, H.; Li, Y.; Huang, C. Cas9-PF, an early flowering and visual selection marker system, enhances the frequency of editing event occurrence and expedites the isolation of genome-edited and transgene-free plants. Plant Biotechnol. J. 2019, 17, 1191. [Google Scholar] [CrossRef]
- He, Y.; Zhu, M.; Wu, J.; Ouyang, L.; Wang, R.; Sun, H.; Yan, L.; Wang, L.; Xu, M.; Zhan, H. Repurposing of anthocyanin biosynthesis for plant transformation and genome editing. Front. Genome Ed. 2020, 2, 607982. [Google Scholar] [CrossRef] [PubMed]
- Peng, C.; Zheng, M.; Ding, L.; Chen, X.; Wang, X.; Feng, X.; Wang, J.; Xu, J. Accurate detection and evaluation of the gene-editing frequency in plants using droplet digital PCR. Front. Plant Sci. 2020, 11, 610790. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chi, X.; Feng, L.; Wu, X.; Qi, X. Improvement of multiplex semi-nested PCR system for screening of rare mutations by high-throughput sequencing. BioTechniques 2019, 67, 294–298. [Google Scholar] [CrossRef]
- Zhang, B.; Zhou, J.; Li, M.; Wei, Y.; Wang, J.; Wang, Y.; Shi, P.; Li, X.; Huang, Z.; Tang, H. Evaluation of CRISPR/Cas9 site-specific function and validation of sgRNA sequence by a Cas9/sgRNA-assisted reverse PCR technique. Anal. Bioanal. Chem. 2021, 413, 2447–2456. [Google Scholar] [CrossRef]
- Kamiya, Y.; Abe, F.; Mikami, M.; Endo, M.; Kawaura, K. A rapid method for detection of mutations induced by CRISPR/Cas9-based genome editing in common wheat. Plant Biotechnol. 2020, 37, 247–251. [Google Scholar] [CrossRef]
- Ueta, R.; Abe, C.; Watanabe, T.; Sugano, S.S.; Ishihara, R.; Ezura, H.; Osakabe, Y.; Osakabe, K. Rapid breeding of parthenocarpic tomato plants using CRISPR/Cas9. Sci. Rep. 2017, 7, 507. [Google Scholar] [CrossRef]
- Kim, D.; Alptekin, B.; Budak, H. CRISPR/Cas9 genome editing in wheat. Funct. Integr. Genom. 2018, 18, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Sentmanat, M.F.; Peters, S.T.; Florian, C.P.; Connelly, J.P.; Pruett-Miller, S.M. A survey of validation strategies for CRISPR-Cas9 editing. Sci. Rep. 2018, 8, 888. [Google Scholar] [CrossRef] [PubMed]
- Zong, Y.; Wang, Y.; Li, C.; Zhang, R.; Chen, K.; Ran, Y.; Qiu, J.-L.; Wang, D.; Gao, C. Precise base editing in rice, wheat and maize with a Cas9-cytidine deaminase fusion. Nat. Biotechnol. 2017, 35, 438–440. [Google Scholar] [CrossRef]
- Pan, C.; Ye, L.; Qin, L.; Liu, X.; He, Y.; Wang, J.; Chen, L.; Lu, G. CRISPR/Cas9-mediated efficient and heritable targeted mutagenesis in tomato plants in the first and later generations. Sci. Rep. 2016, 6, 24765. [Google Scholar] [CrossRef]
- Li, M.; Liang, Z.; Zeng, Y.; Jing, Y.; Wu, K.; Liang, J.; He, S.; Wang, G.; Mo, Z.; Tan, F. De novo analysis of transcriptome reveals genes associated with leaf abscission in sugarcane (Saccharum officinarum L.). BMC Genom. 2016, 17, 195. [Google Scholar] [CrossRef] [PubMed]
- Wanderley-Nogueira, A.; Soares-Cavalcanti, N.; Morais, D.; Belarmino, L.; Barbosa Da Silva, A.; Benko-Iseppon, A. Abundance and diversity of resistance genes in the sugarcane transcriptome revealed by in silico analysis. Gen. Mol. Res. 2007, 6, 866–889. [Google Scholar]
- Sharma, R.; Tamta, S. Red rot resistant gene characterization using RGAP markers among sugarcane cultivars resistant and susceptible to the red rot disease. 3 Biotech 2017, 7, 306. [Google Scholar] [CrossRef]
- Raju, G.; Shanmugam, K.; Kasirajan, L. High-throughput sequencing reveals genes associated with high-temperature stress tolerance in sugarcane. 3 Biotech 2020, 10, 198. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Chen, Y.; Feng, A.; Zou, W.; Wang, D.; Lin, P.; Chen, Y.; You, C.; Que, Y.; Su, Y. The allene oxide synthase gene family in sugarcane and its involvement in disease resistance. Ind. Crops Prod. 2023, 192, 116136. [Google Scholar] [CrossRef]
- Zhu, K.; Huang, C.; Phan, T.-T.; Yang, L.-T.; Zhang, B.-Q.; Xing, Y.-X.; Li, Y.-R. Overexpression of SoACLA-1 gene confers drought tolerance improvement in sugarcane. Plant Mol. Biol. Rep. 2021, 39, 489–500. [Google Scholar] [CrossRef]
- Li, J.; Phan, T.-T.; Li, Y.-R.; Xing, Y.-X.; Yang, L.-T. Isolation, transformation and overexpression of sugarcane SoP5CS gene for drought tolerance improvement. Sugar Tech 2018, 20, 464–473. [Google Scholar] [CrossRef]
- Ferreira, T.H.; Gentile, A.; Vilela, R.D.; Costa, G.G.L.; Dias, L.I.; Endres, L.; Menossi, M. microRNAs associated with drought response in the bioenergy crop sugarcane (Saccharum spp.). PLoS ONE 2012, 7, e46703. [Google Scholar] [CrossRef]
- Lin, S.; Chen, T.; Qin, X.; Wu, H.; Khan, M.A.; Lin, W. Identification of microrna families expressed in sugarcane leaves subjected to drought stress and the targets thereof. Pak. J. Agric. Sci. 2014, 51, 925–934. [Google Scholar]
- Andrade, L.M.; Benatti, T.R.; Nobile, P.M.; Goldman, M.H.; Figueira, A.; Marin, A.L.A.; dos Santos Brito, M.; da Silva, J.; Creste, S. Characterization, isolation and cloning of sugarcane genes related to drought stress. BMC Proceedings BioMed Cent. 2014, 8, 110. [Google Scholar]
- Carnavale Bottino, M.; Rosario, S.; Grativol, C.; Thiebaut, F.; Rojas, C.A.; Farrineli, L.; Hemerly, A.S.; Ferreira, P.C.G. High-throughput sequencing of small RNA transcriptome reveals salt stress regulated microRNAs in sugarcane. PLoS ONE 2013, 8, e59423. [Google Scholar] [CrossRef]
- Murugan, N.; Palanisamy, V.; Channappa, M.; Ramanathan, V.; Ramaswamy, M.; Govindakurup, H.; Chinnaswamy, A. Genome-Wide In Silico Identification, Structural Analysis, Promoter Analysis, and Expression Profiling of PHT Gene Family in Sugarcane Root under Salinity Stress. Sustainability 2022, 14, 15893. [Google Scholar] [CrossRef]
- Trujillo, L.E.; Menéndez, C.; Ochogavía, M.E.; Hernández, I.; Borrás, O.; Rodríguez, R.; Coll, Y.; Arrieta, J.G.; Banguela, A.; Ramírez, R. Engineering drought and salt tolerance in plants using SodERF3, a novel sugarcane ethylene responsive factor. Biotecnol. Apl. 2009, 26, 168–171. [Google Scholar]
- Shingote, P.R.; Kawar, P.G.; Pagariya, M.C.; Kuhikar, R.S.; Thorat, A.S.; Babu, K. SoMYB18, a sugarcane MYB transcription factor improves salt and dehydration tolerance in tobacco. Acta Physiol. Plant 2015, 37, 217. [Google Scholar] [CrossRef]
- Begcy, K.; Mariano, E.D.; Lembke, C.G.; Zingaretti, S.M.; Souza, G.M.; Araújo, P.; Menossi, M. Overexpression of an evolutionarily conserved drought-responsive sugarcane gene enhances salinity and drought resilience. Ann. Bot. 2019, 124, 691–700. [Google Scholar] [CrossRef]
- Begcy, K.; Mariano, E.D.; Gentile, A.; Lembke, C.G.; Zingaretti, S.M.; Souza, G.M.; Menossi, M. A novel stress-induced sugarcane gene confers tolerance to drought, salt and oxidative stress in transgenic tobacco plants. PLoS ONE 2012, 7, e44697. [Google Scholar] [CrossRef]
- Jin-Long, G.; Li-Ping, X.; Jing-Ping, F.; Ya-Chun, S.; Hua-Ying, F.; You-Xiong, Q.; Jing-Sheng, X. A novel dirigent protein gene with highly stem-specific expression from sugarcane, response to drought, salt and oxidative stresses. Plant Cell Rep. 2012, 31, 1801–1812. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Liu, Z.; Zhao, H.; Deng, X.; Su, Y.; Li, R.; Chen, B. Overexpression of sugarcane ScDIR genes enhances drought tolerance in Nicotiana benthamiana. Int. J. Mol. Sci. 2022, 23, 5340. [Google Scholar] [CrossRef]
- Liu, F.; Huang, N.; Wang, L.; Ling, H.; Sun, T.; Ahmad, W.; Muhammad, K.; Guo, J.; Xu, L.; Gao, S. A novel L-ascorbate peroxidase 6 gene, ScAPX6, plays an important role in the regulation of response to biotic and abiotic stresses in sugarcane. Front. Plant Sci. 2018, 8, 2262. [Google Scholar] [CrossRef]
- Park, J.-W.; Benatti, T.R.; Marconi, T.; Yu, Q.; Solis-Gracia, N.; Mora, V.; Da Silva, J.A. Cold responsive gene expression profiling of sugarcane and Saccharum spontaneum with functional analysis of a cold inducible Saccharum homolog of NOD26-like intrinsic protein to salt and water stress. PLoS ONE 2015, 10, e0125810. [Google Scholar] [CrossRef]
- Ramasamy, M.; Damaj, M.B.; Vargas-Bautista, C.; Mora, V.; Liu, J.; Padilla, C.S.; Irigoyen, S.; Saini, T.; Sahoo, N.; DaSilva, J.A. A sugarcane G-protein-coupled receptor, ShGPCR1, confers tolerance to multiple abiotic stresses. Front. Plant Sci. 2021, 12, 745891. [Google Scholar] [CrossRef]
- Wu, Q.; Chen, Y.; Zou, W.; Pan, Y.-B.; Lin, P.; Xu, L.; Grisham, M.P.; Ding, Q.; Su, Y.; Que, Y. Genome-wide characterization of sugarcane catalase gene family identifies a ScCAT1 gene associated disease resistance. Int. J. Biol. Macromol. 2023, 232, 123398. [Google Scholar] [CrossRef] [PubMed]
- Asnaghi, C.; Roques, D.; Ruffel, S.; Kaye, C.; Hoarau, J.-Y.; Telismart, H.; Girard, J.-C.; Raboin, L.-M.; Risterucci, A.-M.; Grivet, L. Targeted mapping of a sugarcane rust resistance gene (Bru 1) using bulked segregant analysis and AFLP markers. Theor. Appl. Genet. 2004, 108, 759–764. [Google Scholar] [CrossRef] [PubMed]
- Zafar, S.A.; Zaidi, S.S.E.A.; Gaba, Y.; Singla-Pareek, S.L.; Dhankher, O.P.; Li, X.; Mansoor, S.; Pareek, A. Engineering abiotic stress tolerance via CRISPR/Cas-mediated genome editing. J. Exp. Bot. 2020, 71, 470–479. [Google Scholar] [CrossRef]
- Li, Y.; Wu, X.; Zhang, Y.; Zhang, Q. CRISPR/Cas genome editing improves abiotic and biotic stress tolerance of crops. Front. Genome Ed. 2022, 4, 987817. [Google Scholar] [CrossRef]
- Wang, Y.; Zafar, N.; Ali, Q.; Manghwar, H.; Wang, G.; Yu, L.; Ding, X.; Ding, F.; Hong, N.; Wang, G. CRISPR/Cas Genome Editing Technologies for Plant Improvement against Biotic and Abiotic Stresses: Advances, Limitations, and Future Perspectives. Cells 2022, 11, 3928. [Google Scholar] [CrossRef] [PubMed]
- Ogata, T.; Ishizaki, T.; Fujita, M.; Fujita, Y. CRISPR/Cas9-targeted mutagenesis of OsERA1 confers enhanced responses to abscisic acid and drought stress and increased primary root growth under nonstressed conditions in rice. PLoS ONE 2020, 15, e0243376. [Google Scholar] [CrossRef] [PubMed]
- Santosh Kumar, V.; Verma, R.K.; Yadav, S.K.; Yadav, P.; Watts, A.; Rao, M.; Chinnusamy, V. CRISPR-Cas9 mediated genome editing of drought and salt tolerance (OsDST) gene in indica mega rice cultivar MTU1010. Physiol. Mol. Biol. Plants 2020, 26, 1099–1110. [Google Scholar] [CrossRef]
- Usman, B.; Nawaz, G.; Zhao, N.; Liao, S.; Liu, Y.; Li, R. Precise editing of the OsPYL9 gene by RNA-guided Cas9 nuclease confers enhanced drought tolerance and grain yield in rice (Oryza sativa L.) by regulating circadian rhythm and abiotic stress responsive proteins. Int. J. Mol. Sci. 2020, 21, 7854. [Google Scholar] [CrossRef] [PubMed]
- Lim, C.; Kang, K.; Shim, Y.; Yoo, S.-C.; Paek, N.-C. Inactivating transcription factor OsWRKY5 enhances drought tolerance through abscisic acid signaling pathways. Plant Physiol. 2022, 188, 1900–1916. [Google Scholar] [CrossRef]
- Illouz-Eliaz, N.; Nissan, I.; Nir, I.; Ramon, U.; Shohat, H.; Weiss, D. Mutations in the tomato gibberellin receptors suppress xylem proliferation and reduce water loss under water-deficit conditions. J. Exp. Bot. 2020, 71, 3603–3612. [Google Scholar] [CrossRef]
- Wang, L.; Chen, L.; Li, R.; Zhao, R.; Yang, M.; Sheng, J.; Shen, L. Reduced drought tolerance by CRISPR/Cas9-mediated SlMAPK3 mutagenesis in tomato plants. J. Agric. Food Chem. 2017, 65, 8674–8682. [Google Scholar] [CrossRef] [PubMed]
- Abdallah, N.A.; Elsharawy, H.; Abulela, H.A.; Thilmony, R.; Abdelhadi, A.A.; Elarabi, N.I. Multiplex CRISPR/Cas9-mediated genome editing to address drought tolerance in wheat. GM Crops Food 2022, 1–17. [Google Scholar] [CrossRef]
- Mohr, T.; Horstman, J.; Gu, Y.Q.; Elarabi, N.I.; Abdallah, N.A.; Thilmony, R. CRISPR-Cas9 Gene Editing of the Sal1 Gene Family in Wheat. Plants 2022, 11, 2259. [Google Scholar] [CrossRef] [PubMed]
- Zhong, X.; Hong, W.; Shu, Y.; Li, J.; Liu, L.; Chen, X.; Islam, F.; Zhou, W.; Tang, G. CRISPR/Cas9 mediated gene-editing of GmHdz4 transcription factor enhances drought tolerance in soybean (Glycine max [L.] Merr.). Front. Plant Sci. 2022, 13, 988505. [Google Scholar] [CrossRef]
- Badhan, S.; Ball, A.S.; Mantri, N. First report of CRISPR/Cas9 mediated DNA-free editing of 4CL and RVE7 genes in chickpea protoplasts. Int. J. Mol. Sci. 2021, 22, 396. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.S.; Kong, J.; Tao, R.; Ahmed, T.; Alamin, M.; Alotaibi, S.S.; Abdelsalam, N.R.; Xu, J.-H. CRISPR/Cas9 mediated knockout of the OsbHLH024 transcription factor improves salt stress resistance in rice (Oryza sativa L.). Plants 2022, 11, 1184. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Chen, Z.; Li, P.; Xu, H.; Liu, K.; Zha, W.; Li, S.; Chen, J.; Yang, G.; Huang, J. Development of novel rice germplasm for salt-tolerance at seedling stage using CRISPR-Cas9. Sustainability 2022, 14, 2621. [Google Scholar] [CrossRef]
- Yue, E.; Cao, H.; Liu, B. OsmiR535, a potential genetic editing target for drought and salinity stress tolerance in Oryza sativa. Plants 2020, 9, 1337. [Google Scholar] [CrossRef]
- Bouzroud, S.; Gasparini, K.; Hu, G.; Barbosa, M.A.M.; Rosa, B.L.; Fahr, M.; Bendaou, N.; Bouzayen, M.; Zsögön, A.; Smouni, A. Down regulation and loss of auxin response factor 4 function using CRISPR/Cas9 alters plant growth, stomatal function and improves tomato tolerance to salinity and osmotic stress. Genes 2020, 11, 272. [Google Scholar] [CrossRef]
- Tran, M.T.; Doan, D.T.H.; Kim, J.; Song, Y.J.; Sung, Y.W.; Das, S.; Kim, E.J.; Son, G.H.; Kim, S.H.; Van Vu, T. CRISPR/Cas9-based precise excision of SlHyPRP1 domain (s) to obtain salt stress-tolerant tomato. Plant Cell Rep. 2021, 40, 999–1011. [Google Scholar] [CrossRef]
- Wang, T.; Xun, H.; Wang, W.; Ding, X.; Tian, H.; Hussain, S.; Dong, Q.; Li, Y.; Cheng, Y.; Wang, C. Mutation of GmAITR genes by CRISPR/Cas9 genome editing results in enhanced salinity stress tolerance in soybean. Front. Plant Sci. 2021, 2752. [Google Scholar] [CrossRef]
- Chu, C.; Huang, R.; Liu, L.; Tang, G.; Xiao, J.; Yoo, H.; Yuan, M. The rice heavy-metal transporter OsNRAMP1 regulates disease resistance by modulating ROS homoeostasis. Plant, Cell Environ. 2022, 45, 1109–1126. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.D.; Huang, S.; Yamaji, N.; Zhang, W.; Ma, J.F.; Zhao, F.J. OsNRAMP1 transporter contributes to cadmium and manganese uptake in rice. Plant Cell Environ. 2020, 43, 2476–2491. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.-Z.; Chen, M.-X.; Yu, L.-J.; Xie, L.-J.; Yuan, L.-B.; Qi, H.; Xiao, M.; Guo, W.; Chen, Z.; Yi, K. OsARM1, an R2R3 MYB transcription factor, is involved in regulation of the response to arsenic stress in rice. Front. Plant Sci. 2017, 8, 1868. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Wang, C.; Liu, P.; Lei, C.; Hao, W.; Gao, Y.; Liu, Y.-G.; Zhao, K. Enhanced rice blast resistance by CRISPR/Cas9-targeted mutagenesis of the ERF transcription factor gene OsERF922. PLoS ONE 2016, 11, e0154027. [Google Scholar] [CrossRef]
- Ma, J.; Chen, J.; Wang, M.; Ren, Y.; Wang, S.; Lei, C.; Cheng, Z. Disruption of OsSEC3A increases the content of salicylic acid and induces plant defense responses in rice. J. Exp. Bot. 2018, 69, 1051–1064. [Google Scholar] [CrossRef]
- Zhou, Y.; Xu, S.; Jiang, N.; Zhao, X.; Bai, Z.; Liu, J.; Yao, W.; Tang, Q.; Xiao, G.; Lv, C. Engineering of rice varieties with enhanced resistances to both blast and bacterial blight diseases via CRISPR/Cas9. Plant Biotechnol. J. 2022, 20, 876–885. [Google Scholar] [CrossRef]
- Nekrasov, V.; Wang, C.; Win, J.; Lanz, C.; Weigel, D.; Kamoun, S. Rapid generation of a transgene-free powdery mildew resistant tomato by genome deletion. Sci. Rep. 2017, 7, 482. [Google Scholar] [CrossRef]
- Santillán Martínez, M.I.; Bracuto, V.; Koseoglou, E.; Appiano, M.; Jacobsen, E.; Visser, R.G.; Wolters, A.-M.A.; Bai, Y. CRISPR/Cas9-targeted mutagenesis of the tomato susceptibility gene PMR4 for resistance against powdery mildew. BMC Plant Biol. 2020, 20, 284. [Google Scholar] [CrossRef]
- Wang, Y.; Cheng, X.; Shan, Q.; Zhang, Y.; Liu, J.; Gao, C.; Qiu, J.-L. Simultaneous editing of three homoeoalleles in hexaploid bread wheat confers heritable resistance to powdery mildew. Nat. Biotechnol. 2014, 32, 947–951. [Google Scholar] [CrossRef]
- Wan, D.-Y.; Guo, Y.; Cheng, Y.; Hu, Y.; Xiao, S.; Wang, Y.; Wen, Y.-Q. CRISPR/Cas9-mediated mutagenesis of VvMLO3 results in enhanced resistance to powdery mildew in grapevine (Vitis vinifera). Hortic. Res. 2020, 7, 116. [Google Scholar] [CrossRef]
- Sun, Q.; Lin, L.; Liu, D.; Wu, D.; Fang, Y.; Wu, J.; Wang, Y. CRISPR/Cas9-mediated multiplex genome editing of the BnWRKY11 and BnWRKY70 genes in Brassica napus L. Int. J. Mol. Sci. 2018, 19, 2716. [Google Scholar] [CrossRef]
- Fister, A.S.; Landherr, L.; Maximova, S.N.; Guiltinan, M.J. Transient expression of CRISPR/Cas9 machinery targeting TcNPR3 enhances defense response in Theobroma cacao. Front. Plant Sci. 2018, 9, 268. [Google Scholar] [CrossRef] [PubMed]
- Králová, M.; Bergougnoux, V.; Frébort, I. CRISPR/Cas9 genome editing in ergot fungus Claviceps purpurea. J. Biotechnol. 2021, 325, 341–354. [Google Scholar] [CrossRef] [PubMed]
- Arulganesh, T.; Kumam, Y.; Kumar, K.; Arul, L.; Kokiladevi, E.; Nakeeran, S.; Varanavasiappan, S.; Manonmani, S.; Sudhakar, D. Genome editing of elite rice cultivar CO51 for bacterial leaf blight resistance. Electron. J. Plant Breed. 2021, 12, 1060–1068. [Google Scholar]
- Ortigosa, A.; Gimenez-Ibanez, S.; Leonhardt, N.; Solano, R. Design of a bacterial speck resistant tomato by CRISPR/Cas9-mediated editing of Sl JAZ 2. Plant Biotechnol. J. 2019, 17, 665–673. [Google Scholar] [CrossRef]
- Thomazella, D.P.D.T.; Seong, K.; Mackelprang, R.; Dahlbeck, D.; Geng, Y.; Gill, U.S.; Qi, T.; Pham, J.; Giuseppe, P.; Lee, C.Y. Loss of function of a DMR6 ortholog in tomato confers broad-spectrum disease resistance. Proc. Natl. Acad. Sci. USA 2021, 118, e2026152118. [Google Scholar] [CrossRef]
- Tripathi, J.N.; Ntui, V.O.; Ron, M.; Muiruri, S.K.; Britt, A.; Tripathi, L. CRISPR/Cas9 editing of endogenous banana streak virus in the B genome of Musa spp. overcomes a major challenge in banana breeding. Commun. Biol. 2019, 2, 46. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, J.N.; Ntui, V.O.; Shah, T.; Tripathi, L. CRISPR/Cas9-mediated editing of DMR6 orthologue in banana (Musa spp.) confers enhanced resistance to bacterial disease. Plant Biotechnol. J. 2021, 19, 1291. [Google Scholar] [CrossRef]
- Kumam, Y.; Rajadurai, G.; Kumar, K.; Varanavasiappan, S.; Reddy, M.K.; Krishnaveni, D.; Mangrauthia, S.K.; Raveendran, M.; Arul, L.; Kokiladevi, E. Genome editing of indica rice ASD16 for imparting resistance against rice tungro disease. J. Plant Biochem. Biotechnol. 2022, 31, 880–893. [Google Scholar] [CrossRef]
- Macovei, A.; Sevilla, N.R.; Cantos, C.; Jonson, G.B.; Slamet-Loedin, I.; Čermák, T.; Voytas, D.F.; Choi, I.R.; Chadha-Mohanty, P. Novel alleles of rice eIF4G generated by CRISPR/Cas9-targeted mutagenesis confer resistance to Rice tungro spherical virus. Plant Biotechnol. J. 2018, 16, 1918–1927. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Ma, S.; Hu, P.; Ji, Y.; Sun, F. Genome editing of rice eIF4G loci confers partial resistance to rice black-streaked dwarf virus. Viruses 2021, 13, 2100. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Kong, M.; Yang, F.; Zhu, J.; Qi, X.; Weng, J.; Di, D.; Xie, C. Targeted generation of Null Mutants in ZmGDIα confers resistance against maize rough dwarf disease without agronomic penalty. Plant Biotechnol. J. 2022, 20, 803. [Google Scholar] [CrossRef]
- Lucioli, A.; Tavazza, R.; Baima, S.; Fatyol, K.; Burgyan, J.; Tavazza, M. CRISPR-Cas9 Targeting of the eIF4E1 Gene Extends the Potato Virus Y Resistance Spectrum of the Solanum tuberosum L. cv. Desirée. Front. Microbiol. 2022, 13, 873930. [Google Scholar] [CrossRef] [PubMed]
- Noureen, A.; Khan, M.Z.; Amin, I.; Zainab, T.; Mansoor, S. CRISPR/Cas9-mediated targeting of susceptibility factor eIF4E-enhanced resistance against potato virus Y. Front. Genet. 2022, 13, 922019. [Google Scholar]
- Ishikawa, M.; Yoshida, T.; Matsuyama, M.; Kouzai, Y.; Kano, A.; Ishibashi, K. Tomato brown rugose fruit virus resistance generated by quadruple knockout of homologs of TOBAMOVIRUS MULTIPLICATION1 in tomato. Plant Physiol. 2022, 189, 679–686. [Google Scholar] [CrossRef]
- Ghorbani Faal, P.; Farsi, M.; Seifi, A.; Mirshamsi Kakhki, A. Virus-induced CRISPR-Cas9 system improved resistance against tomato yellow leaf curl virus. Mol. Biol. Rep. 2020, 47, 3369–3376. [Google Scholar] [CrossRef]
- Zhang, P.; Du, H.; Wang, J.; Pu, Y.; Yang, C.; Yan, R.; Yang, H.; Cheng, H.; Yu, D. Multiplex CRISPR/Cas9-mediated metabolic engineering increases soya bean isoflavone content and resistance to soya bean mosaic virus. Plant Biotechnol. J. 2020, 18, 1384–1395. [Google Scholar] [CrossRef]
- Chandrasekaran, J.; Brumin, M.; Wolf, D.; Leibman, D.; Klap, C.; Pearlsman, M.; Sherman, A.; Arazi, T.; Gal-On, A. Development of broad virus resistance in non-transgenic cucumber using CRISPR/Cas9 technology. Mol. Plant Pathol. 2016, 17, 1140–1153. [Google Scholar] [CrossRef]
- Yu, K.; Liu, Z.; Gui, H.; Geng, L.; Wei, J.; Liang, D.; Lv, J.; Xu, J.; Chen, X. Highly efficient generation of bacterial leaf blight-resistant and transgene-free rice using a genome editing and multiplexed selection system. BMC Plant Biol. 2021, 21, 197. [Google Scholar] [CrossRef]
- Ahmad, A.; Ghouri, M.Z.; Munawar, N.; Ismail, M.; Ashraf, S.; Aftab, S.O. Regulatory, ethical, and social aspects of CRISPR crops. In CRISPR Crops: The Future of Food Security; Ahmad, A., Khan, S.H., Khan, Z., Eds.; Springer: Singapore, 2021; pp. 261–287. [Google Scholar]
- Gupta, S.; Kumar, A.; Patel, R.; Kumar, V. Genetically modified crop regulations: Scope and opportunity using the CRISPR-Cas9 genome editing approach. Mol. Biol. Rep. 2021, 48, 4851–4863. [Google Scholar] [CrossRef] [PubMed]
- Sprink, T.; Eriksson, D.; Schiemann, J.; Hartung, F. Regulatory hurdles for genome editing: Process-vs. product-based approaches in different regulatory contexts. Plant Cell Rep. 2016, 35, 1493–1506. [Google Scholar] [CrossRef]
- Ahmad, A.; Munawar, N.; Khan, Z.; Qusmani, A.T.; Khan, S.H.; Jamil, A.; Ashraf, S.; Ghouri, M.Z.; Aslam, S.; Mubarik, M.S. An outlook on global regulatory landscape for genome-edited crops. Int. J. Mol. Sci. 2021, 22, 11753. [Google Scholar] [CrossRef] [PubMed]
- Podevin, N.; Davies, H.V.; Hartung, F.; Nogué, F.; Casacuberta, J.M. Site-directed nucleases: A paradigm shift in predictable, knowledge-based plant breeding. Trends Biotechnol. 2013, 31, 375–383. [Google Scholar] [CrossRef] [PubMed]
- Zannoni, L. Evolving regulatory landscape for genome-edited plants. CRISPR J. 2019, 2, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Verma, P.; Tandon, R.; Yadav, G.; Gaur, V. Structural aspects of DNA repair and recombination in crop improvement. Front. Genet. 2020, 1103. [Google Scholar] [CrossRef] [PubMed]
- Van de Wiel, C.; Schaart, J.; Lotz, L.; Smulders, M. New traits in crops produced by genome editing techniques based on deletions. Plant Biotechnol. Rep. 2017, 11, 1–8. [Google Scholar] [CrossRef]
- Buchholzer, M.; Frommer, W.B. An increasing number of countries regulate genome editing in crops. New Phytol. 2022, 237, 12–15. [Google Scholar] [CrossRef]
- Meena, D.M.R.; Reddy, G.; Kumar, R.; Pandey, S.; Hemaprabha, G. Recent Advances in Sugarcane Genomics, Physiology, and Phenomics for Superior Agronomic Traits. Front. Genet. 2022, 1523. [Google Scholar] [CrossRef]
- Mohan, C.; Easterling, M.; Yau, Y.-Y. Gene editing technologies for sugarcane improvement: Opportunities and limitations. Sugar Tech 2022, 1–17. [Google Scholar] [CrossRef]
- Hussin, S.H.; Liu, X.; Li, C.; Diaby, M.; Jatoi, G.H.; Ahmed, R.; Imran, M.; Iqbal, M.A. An Updated Overview on Insights into Sugarcane Genome Editing via CRISPR/Cas9 for Sustainable Production. Sustainability 2022, 14, 12285. [Google Scholar] [CrossRef]
- Labun, K.; Montague, T.G.; Krause, M.; Torres Cleuren, Y.N.; Tjeldnes, H.; Valen, E. CHOPCHOP v3: Expanding the CRISPR web toolbox beyond genome editing. Nucleic Acids Res. 2019, 47, W171–W174. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Ding, Y.; Zhou, Y.; Jin, W.; Xie, K.; Chen, L.-L. CRISPR-P 2.0: An improved CRISPR-Cas9 tool for genome editing in plants. Mol Plant. 2017, 10, 530–532. [Google Scholar] [CrossRef] [PubMed]
- Riaño-Pachón, D.M.; Mattiello, L. Draft genome sequencing of the sugarcane hybrid SP80-3280. F1000Research 2017, 6. [Google Scholar] [CrossRef]
- Garsmeur, O.; Droc, G.; Antonise, R.; Grimwood, J.; Potier, B.; Aitken, K.; Jenkins, J.; Martin, G.; Charron, C.; Hervouet, C. A mosaic monoploid reference sequence for the highly complex genome of sugarcane. Nat. Commun. 2018, 9, 2638. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhang, X.; Tang, H.; Zhang, Q.; Hua, X.; Ma, X.; Zhu, F.; Jones, T.; Zhu, X.; Bowers, J. Allele-defined genome of the autopolyploid sugarcane Saccharum spontaneum L. Nat. Genet. 2018, 50, 1565–1573. [Google Scholar] [CrossRef]
- Joyce, P.; Kuwahata, M.; Turner, N.; Lakshmanan, P. Selection system and co-cultivation medium are important determinants of Agrobacterium-mediated transformation of sugarcane. Plant Cell Rep. 2010, 29, 173–183. [Google Scholar] [CrossRef]
- Metje-Sprink, J.; Menz, J.; Modrzejewski, D.; Sprink, T. DNA-free genome editing: Past, present and future. Front. Plant Sci. 2019, 9, 1957. [Google Scholar] [CrossRef]
- Gazaffi, R.; Oliveira, K.; Souza, A.; Garcia, A. The importance of the germplasm in developing agro-energetic profile sugarcane cultivars. In Sugar Cane Bioethanol: R&D for Productivity and Sustainability; Cortez, L.A.B., Ed.; Blucher: Sao Paulo, Brazil, 2010; pp. 333–343. [Google Scholar]



{kind=link}
{kind=link}
| Stress | Candidate Genes | References |
|---|---|---|
| Drought | SoACLA-1 | [150] |
| Drought | SoP5CS | [151] |
| Drought | miRNAs | [152,153] |
| Drought | ScLoX, Dehydrin. Dirigent-jacalin | [154] |
| Salinity | miRNAs | [155] |
| Salinity | ShPHT | [156] |
| Drought and salinity | SodERF3 | [157] |
| Drought and salinity | SoMYB18 | [158] |
| Drought and salinity | Scdr2 | [159] |
| Drought, salinity and oxidative | Scdr1 | [160] |
| Drought, salinity and oxidative | ScDir | [161,162] |
| Drought, salinity and oxidative | ScAPX6 | [163] |
| Salinity and low temperature | SspNIP2 | [164] |
| Drought, salinity and cold | ShGPCR1 | [165] |
| Whip smut | ScCAT1 | [166] |
| Rust | Bru1 | [167] |
| Stress | Crop | Target Gene | References |
|---|---|---|---|
| Drought | Rice | OsERA1 | [171] |
| OsDST | [172] | ||
| OsPYL9 | [173] | ||
| OsWRKY5 | [174] | ||
| Tomato | SlGID1 | [175] | |
| SlMAPK3 | [176] | ||
| Wheat | TaSal1 | [177] | |
| Sal1 | [178] | ||
| Soybean | GmHdz4 | [179] | |
| Chickpea | 4CL and RVE7 | [180] | |
| Salinity | Rice | OsbHLH024 | [181] |
| OsRR22 | [182] | ||
| OsmiR535 | [183] | ||
| Tomato | SlARF4 | [184] | |
| SlHyPRP1 | [185] | ||
| Soybean | GmAITR | [186] | |
| Heavy metals | Rice | OsNRAMP5 | [187] |
| OsNRAMP1 | [188] | ||
| OsARM1 | [189] | ||
| Fungus | |||
| Rice Blast | Rice | OsERF922 | [190] |
| Rice | OsSEC3A | [191] | |
| Rice | Bsr-d1, Pi21 and ERF922 | [192] | |
| Powdery mildew | Tomato | SlML | [193] |
| Tomato | SlPMR4 | [194] | |
| Wheat | TaMLO-A1 | [195] | |
| Grapevine | VvMOL3 | [196] | |
| Stem rot | Rapeseed | BnWRKY70 | [197] |
| Black pod | Cacao | TcNPR3 | [198] |
| Ergot | Rye | pyr4 and TrpE | [199] |
| Bacteria | |||
| Bacterial leaf blight | Rice | Pi21 or ERF922 | [192] |
| OsSWEET14 | [200] | ||
| Bacterial leaf spot | Tomato | SlJAZ2 | [201] |
| Sldmr6-1 | [202] | ||
| Banana Xanthomonas wilt | Musa balbisiana | LRR, WAK2, WAK5, Vicilin, RPM1, PR1, NPR1 | [203] |
| Banana | MusaDMR6 | [204] | |
| Bacterial speck | Tomato | SLDMR6-1 | [202] |
| Virus | |||
| Rice tungro spherical virus | Rice | eIF4G | [205,206] |
| Rice black streaked dwarf virus | Rice | eIF4G | [207] |
| Rice black-streaked dwarf virus | Maize | ZmGDIα | [208] |
| Potato virus Y | Potato | eIF4E1 | [209] |
| eIF4E | [210] | ||
| Tomato brown rugose fruit virus | Tomato | TOM1 | [211] |
| Tomato yellow leaf curl virus | Tomato | IR, CP of virus | [212] |
| Soybean mosaic virus | Soybean | GmF3H1, GmF3H2 and GmFNSII-1 | [213] |
| Cucumber vein yellowing virus, Zucchini yellow mosaic virus; Papaya ring spot mosaic virus- | Cucumber | eIF4G | [214] |
| Banana streak virus | Banana | ORF1, ORF2,ORF3 of virus | [203] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Surya Krishna, S.; Harish Chandar, S.R.; Ravi, M.; Valarmathi, R.; Lakshmi, K.; Prathima, P.T.; Manimekalai, R.; Viswanathan, R.; Hemaprabha, G.; Appunu, C. Transgene-Free Genome Editing for Biotic and Abiotic Stress Resistance in Sugarcane: Prospects and Challenges. Agronomy 2023, 13, 1000. https://doi.org/10.3390/agronomy13041000
Surya Krishna S, Harish Chandar SR, Ravi M, Valarmathi R, Lakshmi K, Prathima PT, Manimekalai R, Viswanathan R, Hemaprabha G, Appunu C. Transgene-Free Genome Editing for Biotic and Abiotic Stress Resistance in Sugarcane: Prospects and Challenges. Agronomy. 2023; 13(4):1000. https://doi.org/10.3390/agronomy13041000
Chicago/Turabian StyleSurya Krishna, Sakthivel, S R Harish Chandar, Maruthachalam Ravi, Ramanathan Valarmathi, Kasirajan Lakshmi, Perumal Thirugnanasambandam Prathima, Ramaswamy Manimekalai, Rasappa Viswanathan, Govindkurup Hemaprabha, and Chinnaswamy Appunu. 2023. "Transgene-Free Genome Editing for Biotic and Abiotic Stress Resistance in Sugarcane: Prospects and Challenges" Agronomy 13, no. 4: 1000. https://doi.org/10.3390/agronomy13041000