

Effects of Planting Pre-Germinated Buds on Stand Establishment in Sugarcane

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Greenhouse Experiment Setup
2.2. Field Experiment Setup
2.3. Meteorological Conditions
2.4. Growth Measurements
2.5. Statistical Analysis
3. Results and Discussion
3.1. Tiller Count
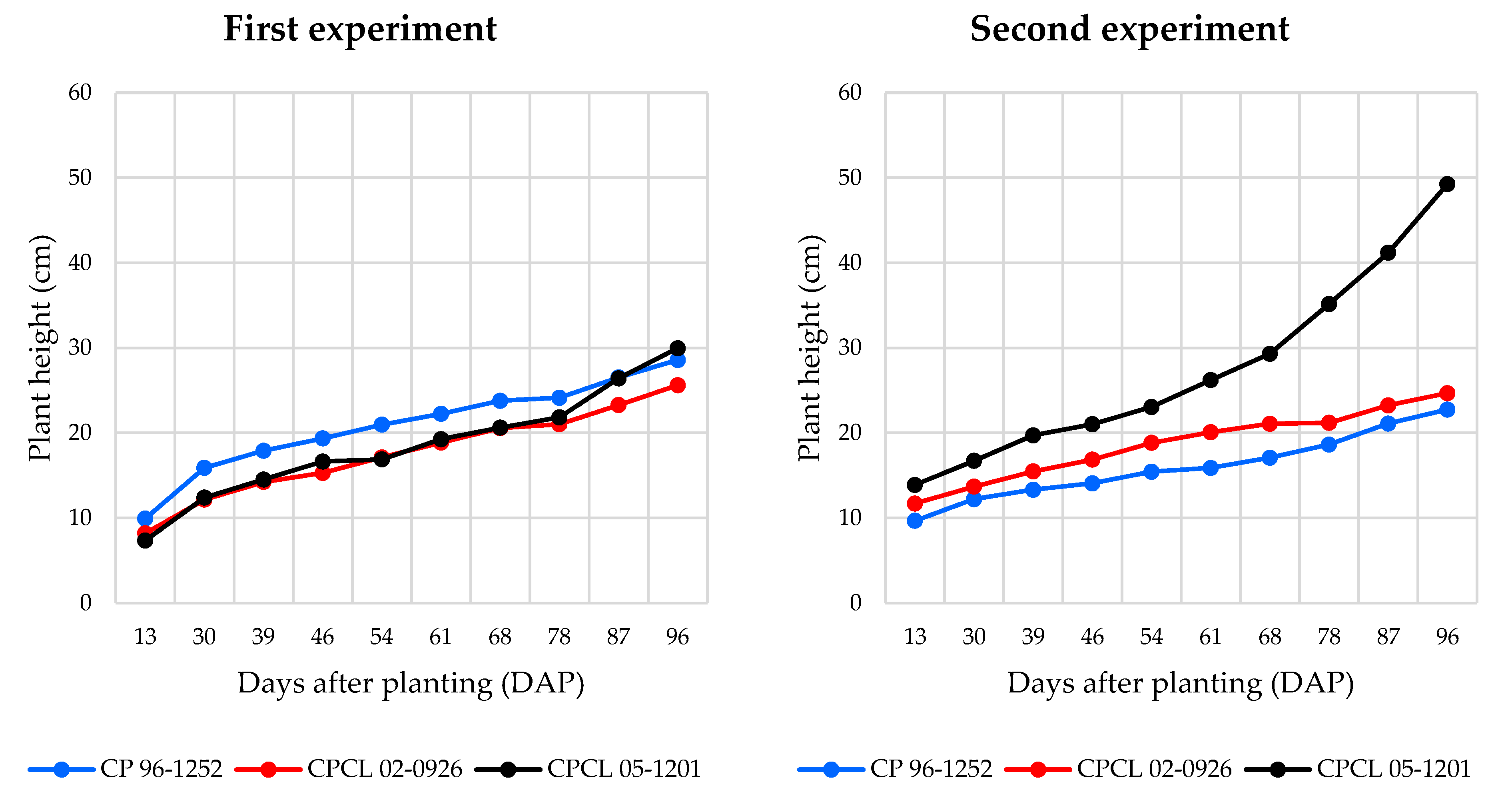
3.2. Primary Shoot Height
3.3. SPAD
3.4. Shoot Dry Biomass
3.5. Root Dry Biomass
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- USDA-NASS. Crop Production; National Agricultural Statistics Service (NASS), Agricultural Statistics Board, United States Department of Agriculture (USDA): Washington, DC, USA, February 2022. Available online: https://www.nass.usda.gov/Publications/Todays_Reports/reports/crop0922.pdf (accessed on 10 September 2022).
- Sandhu, H.S.; Miller, J.D.; Gilbert, R.A.; Odero, D.C. Sugarcane Botany: A Brief View; University of Florida IFAS Extension: Gainesville, FL, USA, 2019; SS-AGR-234; Available online: https://edis.ifas.ufl.edu/publication/SC034 (accessed on 10 September 2022).
- Zeiger, E.; Taiz, L.; Moller, I.M.; Murphy, A. Plant Physiology, 6th ed.; Sinauer: Sunderland, MA, USA, 2014. [Google Scholar]
- Kochhar, S.L.; Gujral, S.K. Plant Physiology: Theory and Applications, 2nd ed.; Cambridge University Press: Cambridge, UK, 2020. [Google Scholar]
- Clements, H.F. Flowering of Sugarcane: Mechanics and Control; Hawaii Agricultural Experiment Station, University of Hawaii: Honolulu, HI, USA, 1975; Volume 92. [Google Scholar]
- Otto, R.; Machado, B.A.; da Silva, A.C.M.; de Castro, S.G.Q.; Lisboa, I.P. Sugarcane pre-sprouted seedlings: A novel method for sugarcane establishment. Field Crop. Res. 2022, 275, 108336. [Google Scholar] [CrossRef]
- dos Santos, S.G.; Chaves, V.A.; Ribeiro, F.D.S.; Alves, G.C.; Reis, V.M. Rooting and growth of pre-germinated sugarcane seedlings inoculated with diazotrophic bacteria. Appl. Soil Ecol. 2019, 133, 12–23. [Google Scholar] [CrossRef]
- De Oliveira, H.P.; De Melo, R.O.; Baldotto, M.A.; Andrade, M.A.; Baldotto, L.E.B. Performance of pre-sprouted sugarcane seedlings in response to the application of humic acid and plant growth-promoting bacteria. Semin. Ciênc. Agrár. 2018, 39, 1365–1370. [Google Scholar] [CrossRef]
- Gírio, L.A.D.S.; da Silva, R.P.; de Menezes, P.C.; Carneiro, F.M.; Zerbato, C.; Ormond, A.T.S. Quality of multi-row harvesting in sugarcane plantations established from pre-sprouted seedlings and billets. Ind. Crop. Prod. 2019, 142, 111831. [Google Scholar] [CrossRef]
- Glynn, N.C.; Zhao, D.; Comstock, J.C.; Milligan, S.B.; Glaz, B.; Edmé, S.J.; Davidson, R.W.; Gilbert, R.A.; Sood, S.; D’Hont, A.; et al. Registration of ‘CPCL 02-0926′ Sugarcane. J. Plant Regist. 2013, 7, 164–171. [Google Scholar] [CrossRef]
- Edme, S.; Tai, P.; Glaz, B.; Gilbert, R.; Miller, J.; Davidson, J.; Dunckelman, J.; Comstock, J. Registration of ‘CP 96-1252′ Sugarcane. Crop. Sci. 2005, 45, 423–424. [Google Scholar] [CrossRef]
- Edmé, S.J.; Davidson, R.W.; Zhao, D.; Comstock, J.C.; Sandhu, H.S.; Glaz, B.; Milligan, S.; Hu, C.-J.; Sood, S.; McCorkle, K.; et al. Registration of ‘CPCL 05-1201′ Sugarcane. J. Plant Regist. 2016, 10, 14–21. [Google Scholar] [CrossRef]
- Sandhu, H.; Davidson, W. Sugarcane Cultivars Descriptive Fact Sheet: CP 96-1252, CP 01-1372 and CP 00-1101. EDIS 2017. Available online: https://edis.ifas.ufl.edu/publication/SC102 (accessed on 10 September 2022). [CrossRef]
- Whiteman, P.; Bull, T.; Glasziou, K. The Physiology of Sugar-Cane VI. Effects of Temperature, Light, and Water on Set Germination and Early Growth of Saccharum Spp. Aust. J. Biol. Sci. 1963, 16, 416–428. [Google Scholar] [CrossRef] [Green Version]
- Glasziou, K.; Bull, T.; Hatch; Whiteman, P. Physiology of Sugar-Cane VII. Effects of Temperature, Photoperiod Duration, and Diurnal and Seasonal Temperature Changes on Growth and Ripening. Aust. J. Biol. Sci. 1965, 18, 53–66. [Google Scholar] [CrossRef]
- Matsuoka, S. Sugarcane tillering and ratooning: Key factors for a profitable cropping. Sugarcane Prod. Cultiv. Uses 2012, 5, 137–157. [Google Scholar]
- Shrivastava, A.K.; Solomon, S.; Rai, R.K.; Singh, P.; Chandra, A.; Jain, R.; Shukla, S.P. Physiological Interventions for Enhancing Sugarcane and Sugar Productivity. Sugar Tech 2015, 17, 215–226. [Google Scholar] [CrossRef]
- Chattha, M.; Ali, A.; Chattha, M. Influence of planting techniques on growth and yield of spring planted sugarcane (Saccharum officinarum L.). Pak. J. Agric. Sci. 2007, 44, 452–455. [Google Scholar]
- Sandhu, H.; Davidson, W. Sugarcane Cultivar Descriptive Fact Sheet: CPCL 02-0926 and CP 05-1526. EDIS 2019, 2019, 5. [Google Scholar] [CrossRef]
- Shrivastava, A.K. Yield decline and its management in sugarcane. In Management and Agro-Industrial Imperatives; Solomon, S., Grewal, S.S., Li, Y., Magarey, R.C., Rao, G.P., Eds.; International Book Distributing Co.: Lucknow, India, 2004; pp. 321–345. [Google Scholar]
- Bendig, J.; Bolten, A.; Bennertz, S.; Broscheit, J.; Eichfuss, S.; Bareth, G. Estimating Biomass of Barley Using Crop Surface Models (CSMs) Derived from UAV-Based RGB Imaging. Remote Sens. 2014, 6, 10395–10412. [Google Scholar] [CrossRef] [Green Version]
- Zhao, D.; Irey, M.; LaBorde, C.; Hu, C. Identifying Physiological and Yield-Related Traits in Sugarcane and Energy Cane. Agron. J. 2017, 109, 927–937. [Google Scholar] [CrossRef]
- Mearaji, H.S.; Ansari, A.; Igdelou, N.K.M.; Lajayer, B.A.; Mohammad, P. Phytohormones and Abiotic Stresses Roles of Phytohormones in Plants under Abiotic Stresses. Nader Khadem Moghadam Igdelou, Behnam Asgari Lajayer, and Mohammad Pessarakli. In Handbook of Plant and Crop Physiology, 4th ed.; Mohammad, P., Ed.; CRC Press: Boca Raton, FL, USA, 2021; pp. 175–213. [Google Scholar]
- Clark, A.; Landolt, W.; Bucher, J.; Strasser, R. Beech (Fagus sylvatica) response to ozone exposure assessed with a chlorophyll a fluorescence performance index. Environ. Pollut. 2000, 109, 501–507. [Google Scholar] [CrossRef]
- Kohila, S.; Gomathi, R. Adaptive physiological and biochemical response of sugarcane genotypes to high-temperature stress. Indian J. Plant Physiol. 2018, 23, 245–260. [Google Scholar] [CrossRef] [Green Version]
- Silva, M.D.A.; Jifon, J.L.; Sharma, V.; Da Silva, J.A.G.; Caputo, M.M.; Damaj, M.B.; Guimarães, E.R.; Ferro, M.I.T. Use of Physiological Parameters in Screening Drought Tolerance in Sugarcane Genotypes. Sugar Tech 2011, 13, 191–197. [Google Scholar] [CrossRef]
- Silva, M.D.A.; Jifon, J.L.; Da Silva, J.A.G.; Dos Santos, C.M.; Sharma, V. Relationships between physiological traits and productivity of sugarcane in response to water deficit. J. Agric. Sci. 2014, 152, 104–118. [Google Scholar] [CrossRef]
- Silva, M.D.A.; Jifon, J.L.; Dos Santos, C.M.; Jadoski, C.J.; Da Silva, J.A.G. Photosynthetic capacity and water use efficiency in sugarcane genotypes subject to water deficit during early growth phase. Braz. Arch. Biol. Technol. 2013, 56, 735–748. [Google Scholar] [CrossRef] [Green Version]
- Dinh, T.H.; Takaragawa, H.; Watanabe, K.; Nakabaru, M.; Kawamitsu, Y. Leaf Photosynthesis Response to Change of Soil Moisture Content in Sugarcane. Sugar Tech 2019, 21, 949–958. [Google Scholar] [CrossRef]
- Jaroenseng, S.; Wongmaneeroj, A.; Amkhab, S. Results of studies the possible correlations between SPAD value and total nitrogen contents in the leaves of sugarcane (Saccharum officinarum L.). J. ISSAAS [Int. Society Southeast Asian Agric. Sci.] 2013, 16, 136. [Google Scholar]
- Bell, M.; Garside, A. Shoot and stalk dynamics and the yield of sugarcane crops in tropical and subtropical Queensland, Australia. Field Crop. Res. 2005, 92, 231–248. [Google Scholar] [CrossRef]
- Otto, R.; Franco, H.C.J.; Faroni, C.E.; Vitti, A.C.; de Oliveira, E.C.A.; Sermarini, R.A.; Trivelin, P.C.O. The Role of Nitrogen Fertilizers in Sugarcane Root Biomass under Field Conditions. Agric. Sci. 2014, 05, 1527–1538. [Google Scholar] [CrossRef] [Green Version]
- Milligan, S.B.; Gravois, K.A.; Bischoff, K.P.; Martin, F.A. Crop Effects on Genetic Relationships Among Sugarcane Traits. Crop. Sci. 1990, 30, 927–931. [Google Scholar] [CrossRef]
- Smith, D.; Inman-Bamber, N.; Thorburn, P. Growth and function of the sugarcane root system. Field Crop. Res. 2005, 92, 169–183. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| First Greenhouse Experiment | |||||
|---|---|---|---|---|---|
| Fixed Effects | Number of Tillers | Primary Shoot Height (cm) | SPAD | Shoot Dry Biomass (g) | Root Dry Biomass (g) |
| Cultivar | 0.1378 | 0.0012 * | 0.0002 * | 0.0598 | 0.0452 * |
| BD level | 0.0100 * | 0.1325 | 0.3501 | 0.0001 * | 0.1393 |
| Cultivar × BD level | 0.9154 | 0.5797 | 0.0584 * | 0.1185 | 0.9582 |
| Second Greenhouse Experiment | |||||
| Cultivar | 0.1687 | <0.0001 * | 0.8773 | 0.1521 | <0.0001 * |
| BD level | 0.1012 | 0.9216 | 0.9224 | 0.1942 | 0.0033 * |
| Cultivar × BD level | 0.9022 | 0.8500 | 0.9240 | 0.3438 | 0.7147 |
| Effect | Treatment | Tillers Pot−1 | Shoot Dry Biomass (g) |
|---|---|---|---|
| Bud germination level | Lalas | 4.61 a | 27.47 a |
| Pop-eyes | 3.21 b | 19.80 b | |
| Ungerminated | 2.94 b | 17.74 b |
| Treatment | 36 DAP | 43 DAP | 53 DAP | 95 DAP |
|---|---|---|---|---|
| Lalas | 82,075 a | 103,872 a | 113,021 a | 240,304 a |
| Pop-eyes | 37,135 b | 47,630 b | 52,474 b | 128,620 b |
| Ungerminated | 29,332 b | 41,172 b | 47,092 b | 157,960 b |
| Effect | Treatment | SPAD |
|---|---|---|
| Cultivar | CP 96-1252 | 31.98 b |
| CPCL 05-1201 | 33.36 b | |
| CPCL 02-0926 | 38.11 a |
| Cultivar | Bud Germination Level | SPAD |
|---|---|---|
| CP 96-1252 | Lalas | 34.67 bcd |
| Pop-eyes | 30.23 d | |
| Ungerminated | 31.04 d | |
| CPCL 05-1201 | Lalas | 36.18 abc |
| Pop-eyes | 30.43 d | |
| Ungerminated | 33.48 cd | |
| CPCL 02-0926 | Lalas | 35.86 abc |
| Pop-eyes | 39.90 a | |
| Ungerminated | 38.57 ab |
| Effect | Treatment | Shoot Dry Biomass (g) |
|---|---|---|
| Bud germination level | Lalas | 27.47 a |
| Pop-eyes | 19.80 b | |
| Ungerminated | 17.74 b |
| Effect | Treatment | Root Dry Biomass (g) |
|---|---|---|
| Bud germination level | Lalas | 13.40 a |
| Pop-eyes | 11.78 a | |
| Ungerminated | 8.78 b |
| Effect | Treatment | Root Dry Biomass (g) | |
|---|---|---|---|
| 1st Experiment | 2nd Experiment | ||
| Cultivar | CP 96-1252 | 15.74 b | 17.06 c |
| CPCL 05-1201 | 19.63 ab | 12.03 b | |
| CPCL 02-0926 | 10.51 a | 14.85 a | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Madala, H.V.; Lesmes-Vesga, R.A.; Odero, C.D.; Sharma, L.K.; Sandhu, H.S. Effects of Planting Pre-Germinated Buds on Stand Establishment in Sugarcane. Agronomy 2023, 13, 1001. https://doi.org/10.3390/agronomy13041001
Madala HV, Lesmes-Vesga RA, Odero CD, Sharma LK, Sandhu HS. Effects of Planting Pre-Germinated Buds on Stand Establishment in Sugarcane. Agronomy. 2023; 13(4):1001. https://doi.org/10.3390/agronomy13041001
Chicago/Turabian StyleMadala, Hima Varsha, Ricardo A. Lesmes-Vesga, Calvin D. Odero, Lakesh K. Sharma, and Hardev S. Sandhu. 2023. "Effects of Planting Pre-Germinated Buds on Stand Establishment in Sugarcane" Agronomy 13, no. 4: 1001. https://doi.org/10.3390/agronomy13041001