
Influence of Environmental Factors on Some Biochemical and Physiological Indicators in Grapevine from Copou Vineyard, Iasi, Romania

 , ,
, ,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
3. Results
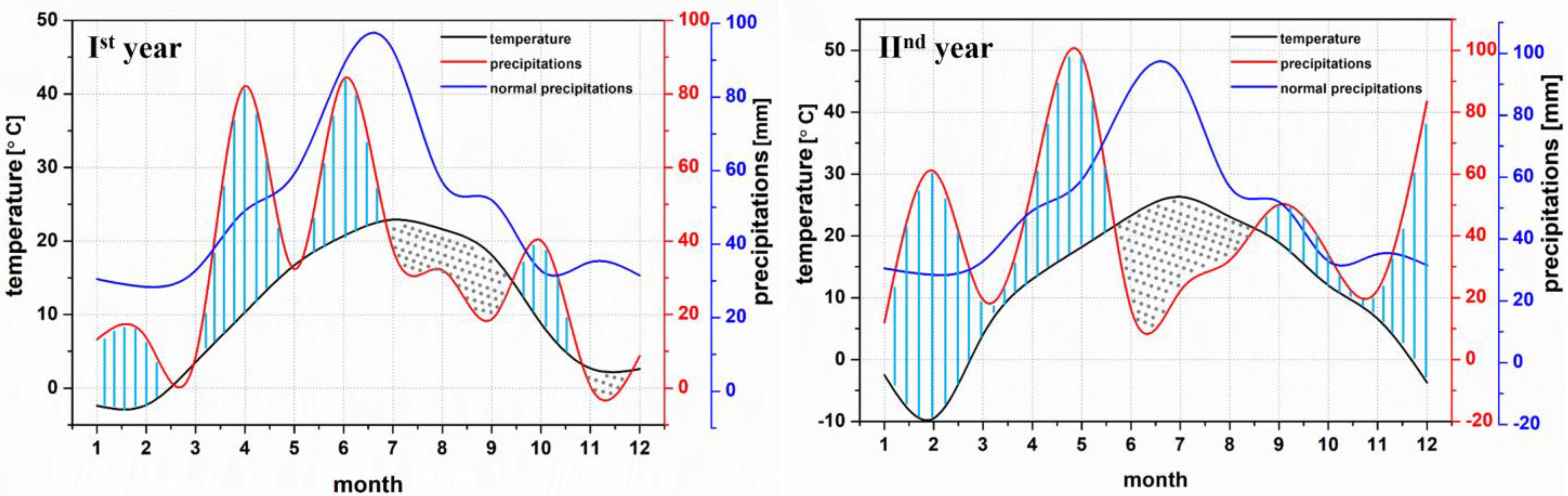
3.1. Meteorological Conditions
3.2. Total Anthocyanin Concentrations
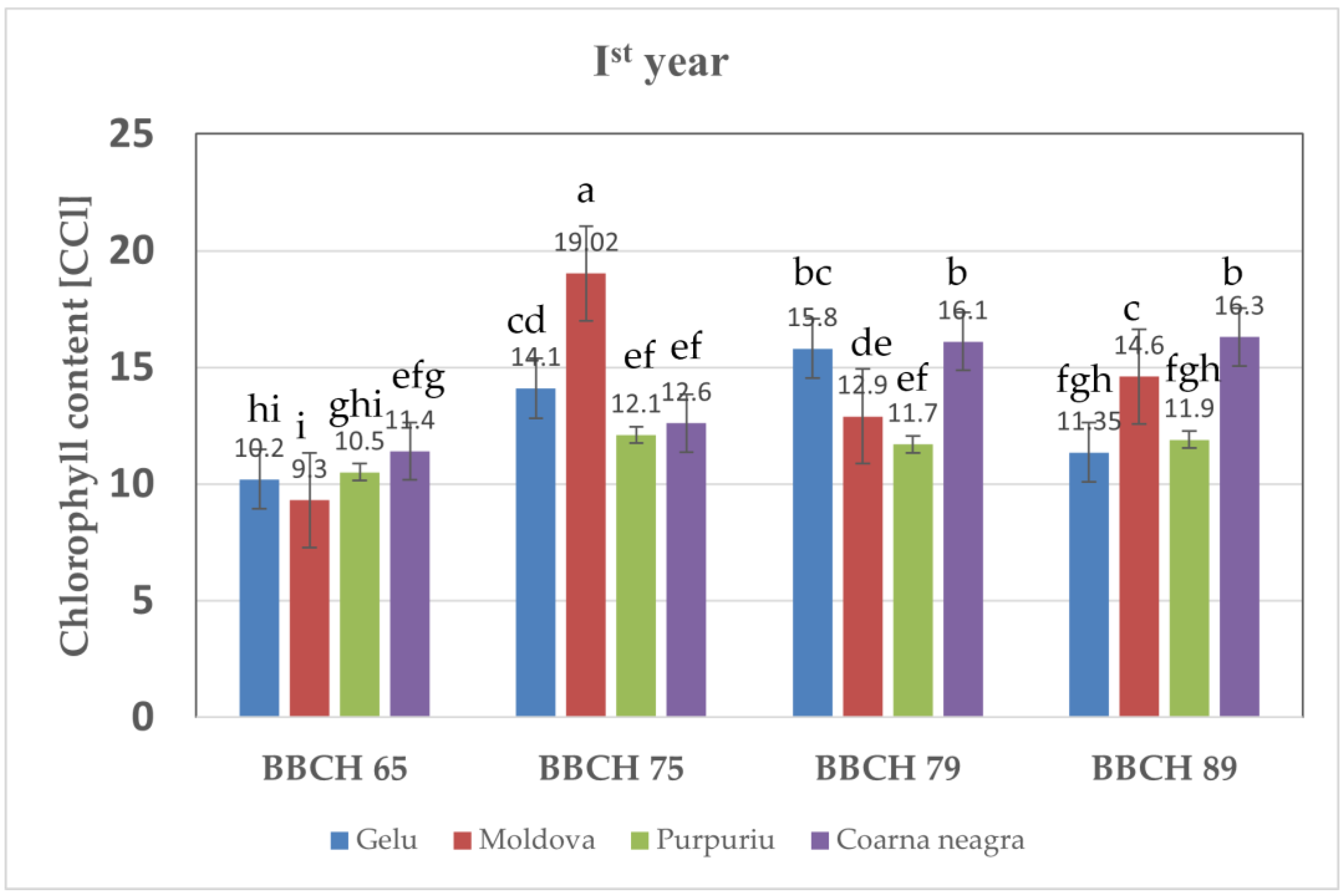
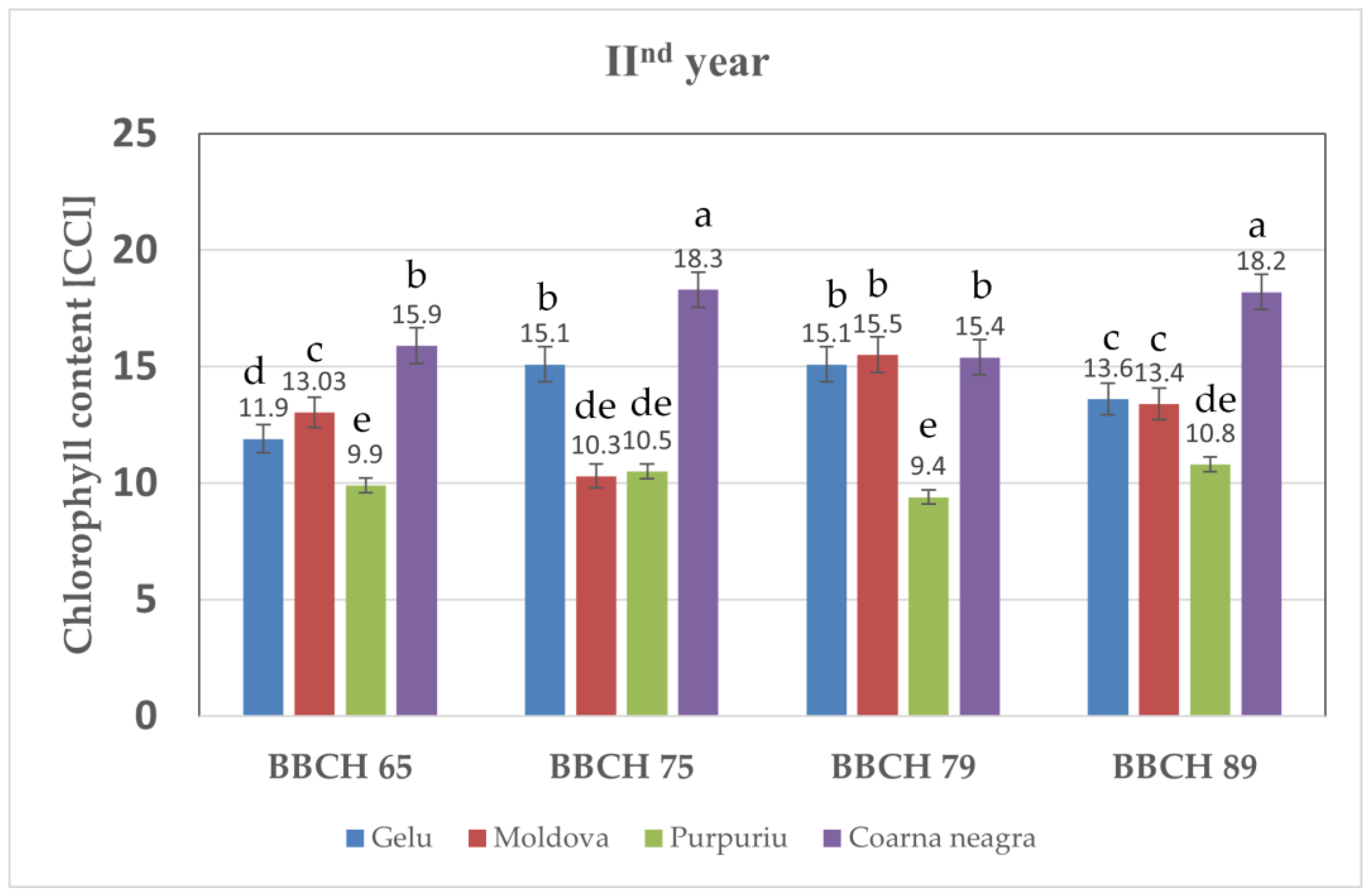
3.3. Total Chlorophyll Content
3.4. LC Pro Photosynthetic Indicators
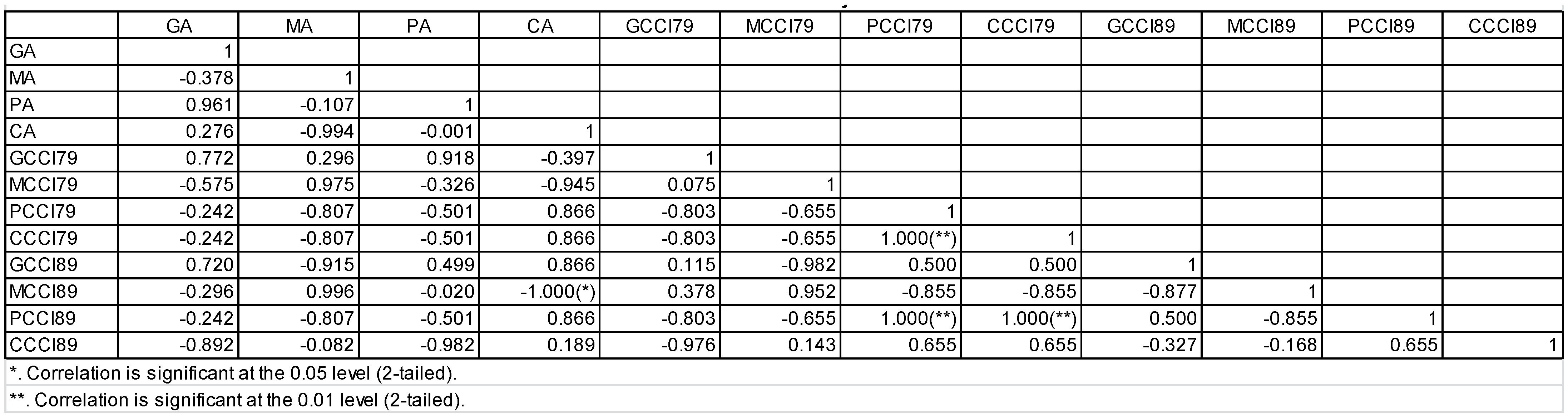
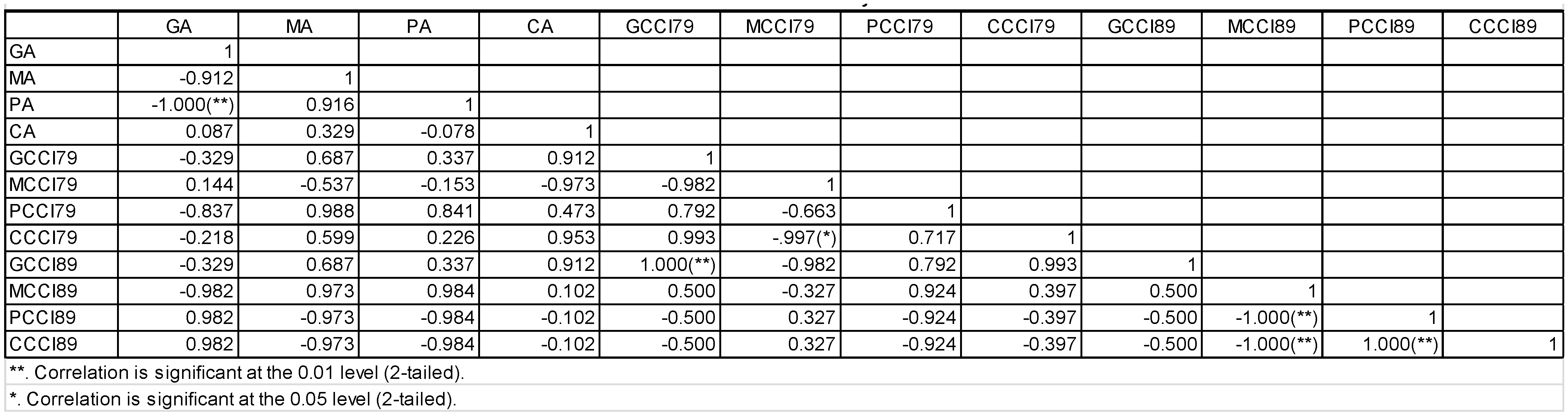
3.5. Pearson Correlation Test
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Keller, M. Managing grapevines to optimise fruit development in a challenging environment: A climate change primer for viticulturists. Aust. J. Grape Wine Res. 2010, 16, 56–69. [Google Scholar] [CrossRef]
- Schultz, H.R. Global climate change, sustainability, and some challenges for grape and wine production. J. Wine Econ. 2016, 11, 181–200. [Google Scholar] [CrossRef]
- Reinth, M.; Vigneron, N.; Darriet, P.; Sweetman, C.; Burbidge, C.; Bonghi, C.; Walker, R.P.; Famiani, F.; Castellarin, S.D. Grape berry secondary metabolites and their modulation by abiotic factors in a climate change scenario—A review. Front. Plant Sci. 2021, 12, 643258. [Google Scholar] [CrossRef] [PubMed]
- De Morales-Castilla, I.; Cortazar-Atauri, I.G.; Cook, B.I.; Wolkovich, E.M. Diversity buffers winegrowing regions from climate change losses. Proc. Natl. Acad. Sci. USA 2020, 117, 2864–2869. [Google Scholar] [CrossRef]
- Corets-Flores, J.; Hernandez-Esquivel, K.B.; Gonzales-Rodriguez, A.; Ibarra-Maniquez, G. Flowering phenology, growth forms, and pollination syndromes in tropical dry forest species: Influence of phylogeny and abiotic factors. Am. J. Bot. 2016, 104, 39–49. [Google Scholar] [CrossRef] [Green Version]
- Cramer, G.R. Abiotic stress and plant responses from the whole vine to the genes. Aus. J. Grape Wine Res. 2010, 16, 86–93. [Google Scholar] [CrossRef]
- Gao, L.; Wang, W.; Li, H.; Li, H.; Yang, Y.; Zheng, H.; Tao, J. Anthocyanin accumulation in grape berry flesh is associated with an alternative splicing variant of VvMYBA1. Plant Physiol. Biochem. 2022, 195, 1–13. [Google Scholar] [CrossRef]
- Brouillard, R. Chemical structure of anthocyanins. In Anthocyanins as Food Colors; Markakis, P., Ed.; Academic Press: London, UK, 1982; pp. 1–38. [Google Scholar]
- He, F.; Mu, L.; Yan, G.-L.; Liang, N.-N.; Pan, Q.-H.; Wang, J.; Reeves, M.J.; Duan, C.-Q. Biosynthesis of Anthocyanins and Their Regulation in Colored Grapes. Molecules 2010, 15, 9057–9091. [Google Scholar] [CrossRef] [Green Version]
- Sadras, V.O.; Moran, M.A. Elevated temperature decouples anthocyanins and sugars in berries of Shiraz and Cabernet Franc. Aust. J. Grape Wine Res. 2012, 18, 115–122. [Google Scholar] [CrossRef]
- Sadras, V.O.; Moran, M.A.; Bonada, M. Effects of elevated temperature in grapevine. I berry sensory traits. Aust. J. Grape Wine Res. 2013, 19, 95–106. [Google Scholar] [CrossRef]
- Yan, Y.; Song, C.; Falginella, L.; Castellarin, S.D. Day temperature has a stronger effect than night temperature on anthocyanin and flavonol accumulation in “merlot” (Vitis vinifera L.) grapes during ripening. Front. Plant Sci. 2020, 11, 1095. [Google Scholar] [CrossRef]
- Theodorou, N.; Nikolaou, N.; Zioziou, E.; Kyraleou, M.; Stamatina, K.; Kotseridis, Y.; Koundouras, S. Anthocyanin content and composition in four red winegrape cultivars (Vitis vinifera L.) under variable irrigation. OENO One 2019, 53, 39–51. [Google Scholar] [CrossRef]
- Alatzas, A.; Theocharis, S.; Miliordos, D.-E.; Leontaridou, K.; Kanellis, A.K.; Kotseridis, Y.; Hatzopoulos, P.; Koundouras, S. The Effect of Water Deficit on Two Greek Vitis vinifera L. Cultivars: Physiology, Grape Composition and Gene Expression during Berry Development. Plants 2021, 10, 1947. [Google Scholar] [CrossRef]
- Barbagallo, M.G.; Vesco, G.; Di Lorenzo, R.; Lo Bianco, R.; Pisciotta, A. Soil and Regulated Deficit Irrigation Affect Growth, Yield and Quality of ‘Nero d’Avola’ Grapes in a Semi-Arid Environment. Plants 2021, 10, 641. [Google Scholar] [CrossRef]
- Castellarin, S.D.; Pfeiffer, A.; Sivilotti, P.; Degan, M.; Peterlunger, E.; Di Gaspero, G. Transcriptional regulation of anthocyanin biosynthesis in ripening fruits of grapevine under seasonal water deficit. Plant Cell Environ. 2007, 30, 1381–1399. [Google Scholar] [CrossRef] [Green Version]
- Bakuradze, T.; Tausend, A.; Galan, J.; Groh, I.A.M.; Berry, D.; Tur, J.A.; Marko, D.; Richling, E. Antioxidative activity and health benefits of anthocyanin-rich fruit juice in healthy volunteers. Free Radic. Res. 2019, 53, 1045–1055. [Google Scholar] [CrossRef]
- De Rosas, M.I.; Martinez, L.; Duran, M.; Malovini, E.; Cavagnaro, J.B. Anthocyanins in nutrition: Biochemistry and health benefits. In Psychiatry and Neuroscience Update; Gargiulo, P.A., Mesones Arroyo, H.L., Eds.; Springer Nature: Cham, Switzerland, 2019; pp. 143–152. [Google Scholar] [CrossRef]
- Khoo, H.E.; Azlan, A.; Tang, S.T.; Lim, S.M. Anthocyanidins and anthocyanins: Colored pigments as food, pharmaceutical ingredients, and the potential health benefits. Food Nutr. Res. 2017, 61, 1361779. [Google Scholar] [CrossRef] [Green Version]
- Smeriglio, A.; Barreca, D.; Bellocoo, E.; Trombetta, D. Chemistry, pharmacology and health benefits of anthocyanins. Phytother. Res. 2016, 30, 1265–1286. [Google Scholar] [CrossRef]
- Mozos, I.; Flangea, C.; Vlad, D.C.; Gug, C.; Mozos, C.; Stoian, D.; Luca, C.T.; Horbańczuk, J.O.; Horbańczuk, O.K.; Atanasov, A.G. Effects of Anthocyanins on Vascular Health. Biomolecules 2021, 11, 811. [Google Scholar] [CrossRef]
- Paissoni, M.A.; Segade, S.R.; Carrero-Carralero, C.; Montanini, C.; Giacosa, S.; Rolle, L. Role of anthocyanin traits on the impact of oenological tannins addition in the first stage if winegrape skin simulated maceration. Food Chem. 2020, 320, 126633. [Google Scholar] [CrossRef]
- Abi-Habib, E.; Vernhet, A.; Roi, S.; Carrillo, S.; Veran, F.; Ducasse, M.A.; Poncet-Legrand, C. Diffusion of phenolic compounds during a model maceration in winemaking: Role of flesh and seeds. J. Sci. Food Agric. 2022, 103, 12331. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Zhang, Y.; Zhu, L.; Huang, Y.; Lu, J. Influence of growing season on phenolic compounds and antioxidant properties of grape berries from vines grown in subtropical climate. J. Agric. Food Chem. 2011, 59, 1078–1086. [Google Scholar] [CrossRef] [PubMed]
- Biniari, K.; Xenaki, M.; Daskalakis, I.; Rusjan, D.; Bouza, D.; Stavrakaki, M. Polyphenolic compounds and antioxidants of skin and berry grapes of Greek Vitis vinifera cultivars in relation to climate conditions. Food Chem. 2020, 307, 125518. [Google Scholar] [CrossRef] [PubMed]
- Cui, Z.H.; Bi, W.L.; Hao, X.Y.; Li, P.M.; Duan, Y.; Walker, M.A.; Xu, Y.; Wang, Q.C. Drought stress enhances up-regulation of anthocyanin biosynthesis in Grapevine leafroll-associated virus 3- infected in vitro grapevine (Vitis vinifera) Leaves. Plant Dis. 2017, 101, 1606–1615. [Google Scholar] [CrossRef] [Green Version]
- Venios, X.; Korkas, E.; Nisiotou, A.; Banilas, G. Grapevine Responses to Heat Stress and Global Warming. Plants 2020, 9, 1754. [Google Scholar] [CrossRef]
- Lee, J.; Durst, R.W.; Wrolstad, R.E. Determination of total monomeric anthocyanin pigment content of fruit juices, beverages, natural colorants, and wines by the pH differential method: Collaborative study. J. AOAC Int. 2005, 88, 1269–1278. [Google Scholar] [CrossRef] [Green Version]
- Balik, J.; Kumsta, M. Evaluation of colour content in grapes originating from south Moravia. Czech J. Food Sci. 2008, 26, S18–S24. [Google Scholar] [CrossRef] [Green Version]
- Meier, U. (Ed.) Growth Stages of Mono- and Dicotyledonous Plants; Julius Kuhn-Institute: Quedlinburg, Germany, 2018; pp. 93–96. [Google Scholar] [CrossRef]
- Douthe, C.; Medrano, H.; Tortosa, I.; Escalona, J.M.; Hernandez-Montes, E.; Pou, A. Whole-plant water use in field grown grapevine: Seasonal and environmental effects on water and carbon balance. Front. Plant. Sci. 2018, 9, 1540. [Google Scholar] [CrossRef]
- Jiang, W.; Li, N.; Zhang, D.; Meinhardt, L.; Cao, B.; Li, Y.; Song, L. Elevated temperature and drought stress significantly affect fruit quality and activity of anthocyanin-related enzymes in jujube (Ziziphus jujuba Mill. Cv.’Lingwuchangzao’). PLoS ONE 2020, 15, 241491. [Google Scholar] [CrossRef]
- Ma, D.; Sun, D.; Wang, C.; Li, Y.; Guo, T. Expression of flavonoid biosynthesis genes and accumulation of flavonoid in wheat leaves in response to drought stress. Plant Physiol. Biochem. 2014, 80, 60–66. [Google Scholar] [CrossRef]
- Lungoci, C.; Motrescu, I.; Filipov, F.; Jitareanu, C.D.; Teliban, G.-C.; Ghitau, C.S.; Puiu, I.; Robu, T. The Impact of Salinity Stress on Antioxidant Response and Bioactive Compounds of Nepeta cataria L. Agronomy 2022, 12, 562. [Google Scholar] [CrossRef]
- Hinojosa-Gómez, J.; San Martín-Hernández, C.; Heredia, J.B.; León-Félix, J.; Osuna-Enciso, T.; Muy-Rangel, M.D. Anthocyanin Induction by Drought Stress in the Calyx of Roselle Cultivars. Molecules 2020, 25, 1555. [Google Scholar] [CrossRef] [Green Version]
- Yamane, T.; Jeong, S.T.; Goto-Yamamoto, N.; Koshita, Y.; Kobayashi, S. Effects of temperature on anthocyanin biosynthesis in grape berry skins. Am. J. Enol. Vitic. 2006, 57, 54–59. [Google Scholar] [CrossRef]
- Blancquaert, E.H.; Oberholster, A.; Da-Silva, J.M.R.; Deloire, A.J. Effects of abiotic factors on phenolic compounds in the grape berry—A review. S. Afr. J. Enol. Vitic. 2019, 40, 1–14. [Google Scholar] [CrossRef]
- Hunag, Z.; Wang, B.; Williams, P.; Pace, R.D. Identification of anthocyanins in muscadine grapes with HLPC-ESI-MS. LWT Food Sci. Technol. 2009, 42, 819–824. [Google Scholar] [CrossRef]
- Liang, Z.; Wu, B.; Fan, P.; Duan, W.; Zheng, X.; Liu, C.; Li, S. Anthocyanin composition and content in grape berry skin in Vitis germplasm. Food Chem. 2008, 111, 837–844. [Google Scholar] [CrossRef]
- Nile, S.H.; Kim, D.H.; Keum, Y.S. Determination of anthocyanin content and antioxidant capacity of different grape varieties. Ciencia Tec. Vitiv. 2015, 30, 60–68. [Google Scholar] [CrossRef]
- Demir, K.; Erdinc, B. Compositional differences in phenolic compounds and anthocyanin contents of some table and wine grape (V. vinifera L.) varieties from Turkey. Oxid. Commun. 2017, 40, 648–656. [Google Scholar]
- Hatfield, J.L.; Dold, C. Water-use efficiency: Advances and Challenges in a Changing Climate. Front. Plant. Sci. 2019, 10, 103. [Google Scholar] [CrossRef] [Green Version]
- Greer, D.H. Temperature-dependent responses of the photosynthetic and chlorophyll fluorescence attributes of apple (Malus domestica) leaves during a sustained high temperature event. Plant Physiol. Biochem. 2015, 97, 139–146. [Google Scholar] [CrossRef]
- Luo, H.B.; Ma, L.; Xi, H.F.; Duan, W.; Li, S.H.; Loescher, W.; Wang, J.F.; Wang, L.J. Photosynthetic responses to heat treatments at different temperatures and following recovery in grapevine (Vitis amurensis L.) leaves. PLoS ONE 2011, 6, 23033. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| January | February | March | April | May | June | July | August | September | October | November | December | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Temperature (°C) | ||||||||||||
| Monthly average | −2.4 | −2.3 | 3.5 | 10.3 | 16.7 | 20.7 | 22.9 | 21.6 | 18.1 | 8.8 | 2.7 | 2.6 |
| Normal average | −3.1 | −1.2 | 3.4 | 10.4 | 16.3 | 19.7 | 21.2 | 20.5 | 15.8 | 10.1 | 4.4 | −0.8 |
| Difference | 0.7 | −1.1 | 0.1 | −0.1 | 0.4 | 1.0 | 1.7 | 1.1 | 2.3 | −1.3 | −1.7 | 3.4 |
| Monthly minimum | −16.6 | −13.1 | −15.7 | 1.5 | 3.8 | 10.7 | 10.4 | 10.2 | 4.9 | −3.7 | −7.9 | −10.2 |
| Monthly maximum | 10.3 | 15.3 | 21.6 | 24.7 | 31.6 | 34.6 | 35.5 | 33.1 | 31.9 | 28.5 | 14.3 | 15.6 |
| Relative humidity of the air (%) | ||||||||||||
| Monthly average | 93 | 79 | 73 | 67 | 67 | 68 | 69 | 59 | 66 | 77 | 82 | 86 |
| Normal average | 86 | 83 | 78 | 70 | 67 | 69 | 70 | 70 | 74 | 78 | 84 | 87 |
| Difference | 7 | −4 | −5 | −3 | 0 | −1 | −1 | −11 | −8 | −1 | −2 | −1 |
| Rainfall (mm) | ||||||||||||
| Monthly total | 13.2 | 13.7 | 8.4 | 82.2 | 32.3 | 84.1 | 37.8 | 32.1 | 18.7 | 40.2 | 0.6 | 8.7 |
| Normal value | 30.5 | 28.4 | 32.8 | 49.1 | 59.1 | 88.7 | 82.8 | 56.9 | 52.0 | 32.8 | 35.1 | 31.5 |
| Difference | −17.3 | −14.7 | −24.4 | 33.1 | −26.8 | −4.6 | −45.0 | −24.8 | −33.3 | 7.4 | −34.5 | −22.8 |
| January | February | March | April | May | June | July | August | September | October | November | December | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Temperature (°C) | ||||||||||||
| Monthly average | −2.5 | −9.5 | 4.0 | 13.0 | 18.2 | 23.3 | 26.3 | 23.1 | 18.9 | 12.0 | 6.6 | −3.7 |
| Normal average | −3.1 | −1.2 | 3.4 | 10.4 | 16.3 | 19.7 | 21.2 | 20.5 | 15.8 | 10.1 | 4.4 | −0.8 |
| Difference | +0.6 | −8.3 | +0.6 | +2.6 | +1.9 | +3.2 | +5.1 | +2.6 | +3.1 | +1.9 | +2.2 | −2.9 |
| Monthly minimum | −19.2 | −20.4 | −18.5 | −0.3 | 4.4 | 6.5 | 8.3 | 11.5 | 0.1 | −1.0 | −3.9 | −7.0 |
| Monthly maximum | 10.8 | 11.3 | 18.9 | 28.6 | 31.4 | 32.1 | 32.1 | 32.9 | 31.9 | 28.0 | 16.4 | 13.2 |
| Relative humidity of the air (%) | ||||||||||||
| Monthly average | 85 | 83 | 72 | 69 | 68 | 58 | 53 | 59 | 65 | 82 | 87 | 92 |
| Normal average | 86.4 | 83.4 | 77.9 | 70.4 | 67.8 | 69.4 | 70.9 | 70.6 | 73.8 | 78.0 | 84.5 | 86.8 |
| Difference | −1.4 | −0.4 | −5.9 | −1.4 | +0.2 | −11.4 | −17.9 | −11.6 | −8.8 | +4.0 | +2.5 | +5.2 |
| Rainfall (mm) | ||||||||||||
| Monthly total | 12.0 | 61.0 | 19.4 | 56.2 | 98.2 | 16.3 | 22.2 | 32.1 | 50.1 | 34.0 | 22.5 | 83.5 |
| Normal value | 30,5 | 28,4 | 32.8 | 49.1 | 59.1 | 88.7 | 82.8 | 56.9 | 52.0 | 32.8 | 35.1 | 31.5 |
| Difference | −18,5 | +32,6 | −13.4 | +7.1 | +39.1 | −72.4 | −60.6 | −24.8 | −1.9 | +1.2 | −10.3 | +52 |
| Variety | pH = 1 | pH = 4.5 | Conc.* [mg/L] | Average Conc. [mg/L] | Conc. per Dry Matter [mg/g] | |||
|---|---|---|---|---|---|---|---|---|
| A520 | A700 | A520 | A700 | |||||
| 1st year | Gelu | 0.416 | 0.017 | 0.125 | 0.019 | 134.13 | 133.06 f | 1.673 f |
| 0.414 | 0.019 | 0.125 | 0.019 | 132.30 | ||||
| 0.417 | 0.019 | 0.126 | 0.018 | 132.75 | ||||
| Moldova | 2.103 | 0.018 | 0.358 | 0.023 | 801.12 | 806.92 b | 7.482 b | |
| 2.106 | 0.02 | 0.344 | 0.022 | 807.53 | ||||
| 2.107 | 0.021 | 0.334 | 0.022 | 812.11 | ||||
| Purpuriu | 1.067 | 0.021 | 0.208 | 0.019 | 392.32 | 389.11 d | 5.713 d | |
| 1.057 | 0.023 | 0.214 | 0.023 | 385.91 | ||||
| 1.062 | 0.023 | 0.212 | 0.023 | 389.11 | ||||
| Coarna neagra | 0.371 | 0.035 | 0.175 | 0.038 | 38.54 | 38.47 h | 0.375 h | |
| 0.374 | 0.038 | 0.176 | 0.039 | 38.54 | ||||
| 0.37 | 0.038 | 0.171 | 0.037 | 38.34 | ||||
| 2nd year | Gelu | 0.708 | 0.014 | 0.172 | 0.013 | 244.91 | 245.22 e | 2.491 e |
| 0.709 | 0.013 | 0.17 | 0.013 | 246.74 | ||||
| 0.703 | 0.013 | 0.17 | 0.013 | 243.99 | ||||
| Moldova | 1.972 | 0.01 | 0.253 | 0.012 | 1545.39 | 1547.49 a | 12.399 a | |
| 1.953 | 0.01 | 0.239 | 0.012 | 1540.90 | ||||
| 1.965 | 0.01 | 0.234 | 0.012 | 1556.17 | ||||
| Purpuriu | 0.723 | 0.01 | 0.149 | 0.01 | 515.43 | 511.83 c | 5.480 c | |
| 0.697 | 0.009 | 0.149 | 0.01 | 492.98 | ||||
| 0.737 | 0.01 | 0.15 | 0.01 | 527.10 | ||||
| Coarna neagra | 0.306 | 0.012 | 0.089 | 0.012 | 99.33 | 101.93 g | 1.167 g | |
| 0.317 | 0.013 | 0.092 | 0.013 | 103.00 | ||||
| 0.317 | 0.013 | 0.091 | 0.013 | 103.45 | ||||
| PAR (µMOL M−2S−1) | Tleaf (°C) | TEXT (°C) | CI (PPM) | E (µMOL H2O M−2S−1) | AN (µMOL CO2 M−2S−1) | WUEINST = A/E | ||
|---|---|---|---|---|---|---|---|---|
| Gelu | BBCH 65 | 277.00 ± 1.1 b | 31.99 ± 0.07 ab | 29.72 ± 0.08 a | 362.00 ± 8.30 c | 1.28 b | 0.04 ef | 0.03 fg |
| Gelu | BBCH 75 | 114.22 ± 1.7 h | 29.64 ± 0.11 cd | 27.44 ± 0.05 b | 347.20 ± 6.25 e | 0.60 d | 0.45 d | 0.74 e |
| Gelu | BBCH 89 | 120.15 ± 1.5 g | 27.70 ± 0.12 e | 25.75 ± 0.07 c | 397.15 ± 7.23 a | 0.36 fg | −0.02 f | −0.04 h |
| Moldova | BBCH 65 | 520.40 ± 2.2 a | 32.01 ± 0.05 ab | 30.28 ± 0.09 a | 339.30 ± 6.38 g | 0.79 c | 0.07 ef | 0.09 fg |
| Moldova | BBCH 75 | 120.18 ± 1.7 g | 27.87 ± 0.09 e | 25.45 ± 0.10 c | 342.77 ± 7.98 f | 0.57 de | 0.78 c | 1.38 d |
| Moldova | BBCH 89 | 172.50 ± 1.5 c | 28.90 ± 0.11 de | 26.70 ± 0.07 bc | 360.65 ± 9.20 c | 0.87 c | 0.76 c | 0.87 e |
| Purpuriu | BBCH 65 | 152.67 ± 0.9 e | 33.48 ± 0.08 a | 30.37 ± 0.12 a | 355.44 ± 7.24 d | 1.86 a | 0.47 d | 0.25 f |
| Purpuriu | BBCH 75 | 94.04 ± 1.2 i | 28.26 ± 0.14 de | 26.13 ± 0.09 bc | 331.17 ± 3.25 i | 0.55 d | 2.15 a | 3.89 a |
| Purpuriu | BBCH 89 | 156.45 ± 0.8 d | 28.80 ± 0.10 de | 26.75 ± 0.08 bc | 368.70 ± 2.65 b | 0.48 ef | 0.41 d | 0.85 e |
| Coarna neagra | BBCH 65 | 156.00 ± 1.4 d | 30.89 ± 0.11 bc | 29.63 ± 0.11 s | 316.89 ± 9.33 j | 0.43 ef | 0.72 c | 1.65 c |
| Coarna neagra | BBCH 75 | 126.42 ± 1.1 f | 27.92 ± 0.18 e | 25.90 ± 0.10 c | 337.32 ± 8.21 h | 0.36 fg | 1.09 b | 3.03 b |
| Coarna neagra | BBCH 89 | 119.40 ± 0.8 g | 28.40 ± 0.12 d | 26.80 ± 0.08 bc | 355.90 ± 10.05 d | 0.24 g | 0.17 e | 0.71 e |
| PAR (µMOL M−2S−1) | Tleaf (°C) | TEXT (°C) | CI (PPM) | E (µMOL H2O M−2S−1) | AN (µMOL CO2 M−2S−1) | WUEINST = A/E | ||
|---|---|---|---|---|---|---|---|---|
| Gelu | BBCH 65 | 156.8 ± 5.3 e | 29.20 ± 0.05 ab | 31.00 ± 0.05 b | 348.1 ± 13.58 i | 0.7 bcd | 3.8 d | 5.43 de |
| Gelu | BBCH 75 | 172.14 ± 2.1 d | 25.03 ± 0.08 de | 25.29 ± 0.04 cd | 496.94 ± 12.20 a | 0.64 bcd | 3.13 e | 4.89 e |
| Gelu | BBCH 89 | 116.4 ± 1.4 i | 27.1 ± 0.05 cd | 24.80 ± 0.11 d | 373.9 ± 12.05 e | 0.5 cde | 0.3 i | 1.4 h |
| Moldova | BBCH 65 | 172.5 ± 2.5 d | 30.77 ± 0.10 a | 32.72 ± 0.08 a | 407.07 ± 11.04 c | 0.49 cde | 2.98 f | 6.08 cd |
| Moldova | BBCH 75 | 89.78 ± 1.3 k | 26.41 ± 0.05 de | 26.58 ± 0.10 c | 365.38 ± 11.25 f | 0.69 bc | 8.56 a | 12.41 a |
| Moldova | BBCH 89 | 181.6 ± 0.9 c | 29.0 ± 0.05 b | 26.6 ± 0.07 cd | 329.10 ± 11.98 j | 1.24 a | 1.31 h | 2.17 g |
| Purpuriu | BBCH 65 | 92.09 ± 1.6 j | 30.19 ± 0.07 a | 30.36 ± 0.05 b | 348.52 ± 12.55 ih | 0.45 e | 5.17 c | 11.4 a |
| Purpuriu | BBCH 75 | 218.41 ± 1.5 b | 26.38 ± 0.12 de | 26.82 ± 0.09 c | 352.88 ± 13.05 d | 0.61 b | 5.76 b | 9.44 b |
| Purpuriu | BBCH 89 | 130.8 ± 2.0 f | 28.8 ± 0.15 b | 26.80 ± 0.10 cd | 397.7 ± 11.20 b | 0.43 de | −0.04 j | −0.13 i |
| Coarna neagra | BBCH 65 | 283.87 ± 1.3 a | 29.38 ± 0.05 ab | 30.06 ± 0.11 b | 461.49 ± 1.51 b | 0.4 cde | 2.53 g | 6.33 c |
| Coarna neagra | BBCH 75 | 64.23 ± 1.4 l | 26.39 ± 0.04 de | 26.51 ± 0.05 cd | 396.65 ± 1.05 d | 0.70 b | 2.58 g | 3.69 f |
| Coarna neagra | BBCH 89 | 119.4 ± 1.5 g | 28.4 ± 0.08 bc | 26.8 ± 0.07 c | 355.9 ± 1.35 g | 0.24 e | 0.17 i | 0.63 h |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marta, A.E.; Slabu, C.; Covasa, M.; Motrescu, I.; Lungoci, C.; Jitareanu, C.D. Influence of Environmental Factors on Some Biochemical and Physiological Indicators in Grapevine from Copou Vineyard, Iasi, Romania. Agronomy 2023, 13, 886. https://doi.org/10.3390/agronomy13030886
Marta AE, Slabu C, Covasa M, Motrescu I, Lungoci C, Jitareanu CD. Influence of Environmental Factors on Some Biochemical and Physiological Indicators in Grapevine from Copou Vineyard, Iasi, Romania. Agronomy. 2023; 13(3):886. https://doi.org/10.3390/agronomy13030886
Chicago/Turabian StyleMarta, Alina Elena, Cristina Slabu, Mihaela Covasa, Iuliana Motrescu, Constantin Lungoci, and Carmenica Doina Jitareanu. 2023. "Influence of Environmental Factors on Some Biochemical and Physiological Indicators in Grapevine from Copou Vineyard, Iasi, Romania" Agronomy 13, no. 3: 886. https://doi.org/10.3390/agronomy13030886