1. Introduction
Salinity is an environmental and/or abiotic stress and a challenge for wheat breeders, hindering the improvement of new cultivars [
1,
2] and wheat production [
3,
4]. Salinity affects more than 6% of the world’s farmland (about 800 million ha [
5], 20% of irrigated land [
3,
6]), and this ratio is forecast to increase to 50% of agricultural areas by 2050 [
7,
8].
Causes of salinization include natural factors such as global warming (climate change) and human activities, e.g., irrigation and drainage systems use, specifically in arid and semi-arid regions [
3,
9]. For instance, approximately 33% of the cultivated land area in Egypt is improperly irrigated with poor drainage practices. In particular, the Nile delta (the most cultivated area) represents 30–37% of the cultivated zones of the Nile River. Mohamed [
10] reported that reusing roughly 10 billion m
3 of drainage water adds to the soil salinity problem. This reflects limited water resources in Egypt and is considered another source of salinization. Soil and water salinity are considered constraints of food production worldwide [
3,
11,
12,
13]. Thus, salinity reduces plant yield [
14,
15] and the possibility of amending new land. In general, wheat is more sensitive to salinity than other field crops. Salinity inhibits plant growth and development, results in low production, or even causes crop failure when extremely severe [
12,
16].
Wheat (
Triticum aestivum L.) is a paramount cultivated cereal crop worldwide and plays a crucial role in food security, representing 765.76 million tons produced from 215.9 million ha [
11,
17]. For example, an approximately 1.3-million-hectare area of cultivated wheat in Egypt produces 9 million tons. This production example represents 50% self-sufficiency in a nation’s first importer of wheat globally.
Since saline soil reclamation is expensive, another option is using existing adapted genotypes or breeding more resilient genotypes. Nevertheless, this acquires adapted genotypes or breading of resilient genotypes to increase wheat production or maintain growth conditions (crop environment). For instance, Morsy et al. [
4] reported that the gypsum supply is suitable for this purpose. However, breeding salt-tolerant genotypes may be crucial for efficiency and economy [
13,
18]. Wheat breeders have a crucial role in improving varieties suitable for biotic and abiotic stresses. Phenotyping by the classical method depends on harvest index and grain yield [
19,
20], improves performance, carbon assimilation, and increases light interception, and consequently may enhance crop production. Hence, increasing the rate and duration of photosynthesis achieves high-yielding performance [
19,
21].
Salinity-induced osmotic and ionic stresses occur by means of reactive oxygen species (ROS). Reactive oxygen species can interact with unsaturated fatty acids to produce peroxidation of vital membrane lipids in plasmalemma; evidence suggests that membranes are the principal sites of saline harm to cells [
22]. Malondialdehyde (MDA) is a lipid peroxidation product that indicates membrane damage under salt stress conditions [
23]. Glycine betaine (GB) and many other osmolytes that are accumulated play essential roles in preventing cells from becoming damaged under stress. Increased GB has been considered to reflect the ability to cope with salinity stress through its role in osmotic adjustment [
24,
25]. Selection for such physiological traits is not common in breeding programs because it is time-consuming and requires effort to measure.
Remote sensing techniques are considered highly productive, precise, and accurate means of determining plant growth or plant vegetation, resulting from spectral reflectance indices (SRI) used to produce phenotypic data [
19]. This data assists breeders in selecting and releasing new cultivars with salt tolerance and high yielding. Spectroscopic measurements are extensively used for phenotyping crop growth by producing spectral reflectance indices alternatively with conventional methods [
26,
27,
28]. The benefit of this approach in comparison with the physiological method (estimated in the laboratory) is rapid assessment, precision, and non-destructive measurements [
28,
29]. To investigate plant vigor and performance, spectral reflectance indices (SRIs) rely on visible, near infrared, and shortwave infrared, which are performed in plant phenotyping and screening, such as in normalized difference vegetation index (NDVI), photochemical reflectance indices, and simple ratio index [
19,
30]. Based on SRIs, calculate several related traits, such as leaf and canopy water status, abundance of pigments, and photosynthesis products in many plant species [
27,
28,
31]. For example, Jackson and Ezra [
32] reported that reflectance indices were related to water stress in wheat [
19,
33,
34] and quinoa [
27]. Additionally, selection efficiency increases through the integration of grain yield and spectral reflectance indices into phenotyping potential [
19,
26,
35]. Breeding efficiency can be increased when using spectral reflectance in these SRI indices and grain yield, which simultaneously improves performance and understanding of the genetic architecture of plants exposed to normal and saline conditions. Thus, breeding efficiency is increasing in the near-infrared region and vice versa [
34,
36]. Based on spectral reflectance of canopy, several vegetation indices can be estimated, particularly in plants exposed to stress, which are indirect indicators for agro-morphological traits [
34]. For example, SRIs correlated with the growth and yield of cultivar Sakha 93 were greater than water indices compared to the Sakha 61 variety [
34]. It was reported that reflectance ratio and NDVI were significantly correlated with above-ground biomass fresh weight of contrasting salt tolerance wheat genotypes [
34,
37]. Furthermore, canopy spectral reflectance represents obvious responses to changes in plant water stats [
28]. The results of researchers [
27] showed that genotypes Baer, Pison, and QQ 74 had the highest NDVI across environments. However, the genotype Japanese Strin had the lowest value. Canopy spectral signature can be exploited in the calculation of plant biomass, dry weight in the different growth stages, and biologically of 64 genotypes treated by 150 mM NaCl to calculate 13 SRI; genotypic differences were found between three traits and spectral reflectance indices and their interaction with growth stages and years [
9].
Chlorophyll fluorescence parameters, such as maximum quantum photosystem II (PSII) photochemical efficiency estimated as Fv/Fm ratio and chlorophyll content, have been confirmed as a physiological trait of salt tolerance, expressly, under saline conditions as maintenance of photosynthetic activities [
38,
39], for example, the relationship between chlorophyll content and genotype salinity tolerance reported by Wu et al. [
40] in barley and wheat crops [
39]. Additionally, these measurements can be used in the early determination of chlorophyll fluorescence to prevent reduction of plant biomass under salinity conditions [
41]. Salinity tolerance is a complex phenomenon controlled by physiological and genetic factors and influenced by growth stages [
39,
42,
43], as well as drought and heat stresses in wheat [
44].
The genotype by trait (GT) and genotype by yield*trait (GYT) was proposed by [
45,
46]. It is considered a graphical selection tool in breeders’ hands for screening and ranking genotypes, which subjected not only grain yield across environments but also the associated traits such as agronomic traits, physiological traits, and end-use characteristics, for example, biotic stress of rusts disease characteristics [
47,
48] and abiotic stress such as barley drought tolerance [
49], and rice [
50], and durum wheat [
51]. Moreover, Sardouie–Nasab et al. [
13] performed salinity tolerance indices using principal component analysis (PCA) to identify salinity tolerance of wheat genotypes. The genotype by trait (GT) and GYT biplot techniques are practical tools, which make genotype selection beneficial and appropriate, specifically in cases evaluated under stress conditions [
49]. The recent GYT model calculates by multiplication of genotype grain yield mean by other traits, such as grain yield (GY) by plant height (PH), (GY*PH) in case the desired value is the highest value. However, grain yield is used when the lowest value of the combination is desired, such as for grain yield and lodging (GY/LO) [
46].
Therefore, the aims of this study are (i) to characterize and screen 40 elite genotypes selected from the Egyptian national breeding program (local and exotic materials multi-location trails) under artificial saline conditions, and (ii) to identify the appropriate genotypes using genotype by yield*trait (GYT) and genotypes by stress tolerance indices (GSTI) biplots of agronomic, physiological traits, and spectral reflectance indices (SRI) to select the salinity tolerance genotypes and involve them in the breeding program.
3. Results
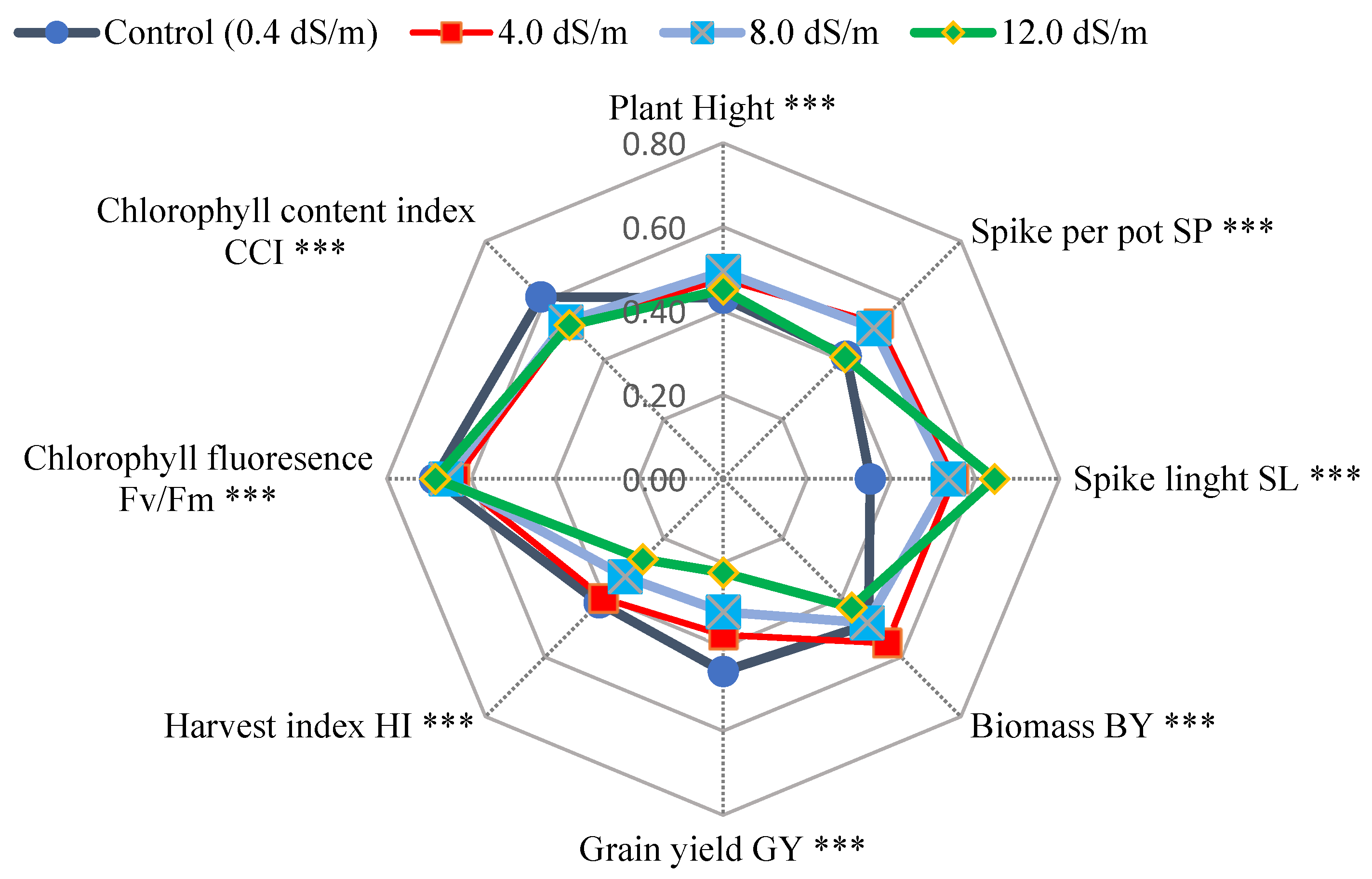
Averaged across forty wheat genotypes, the means of agronomic traits, e.g., grain yield gm per pot (GY), biomass gm per pot (BY), spikes per pot (SP), spike length in cm (SL), and harvest index % (HI), and measured physiological traits, chlorophyll fluoresce (Fv/Fm), and chlorophyll content index (CCI) under four salinity treatments (0.4, 4.0, 8.0 and 12.0 dS/m of seawater) are illustrated in
Figure 1. GY and HI means were affected by the treatments gradually from 0.4 dS/m followed by 4.0, 8.0, and 12.0 dS/m as well as BY, with the exception of the control treatment, recorded the same mean of 8.0 dS/m. However, PH and Fv/Fm had almost the same performance. The control (0.4 dS/m) mean recorded was higher than other salinity treatments of CCI. In contrast, 4.0 and 8.0 dS/m means possessed similar values of SP and SL traits. Control and 12.0 dS/m behaved equally for SP, but they varied in SL. Additionally, the mean square of the genotype varied significantly (
p < 0.001) for all studied traits as shown in
Figure 1 and
Table 4.
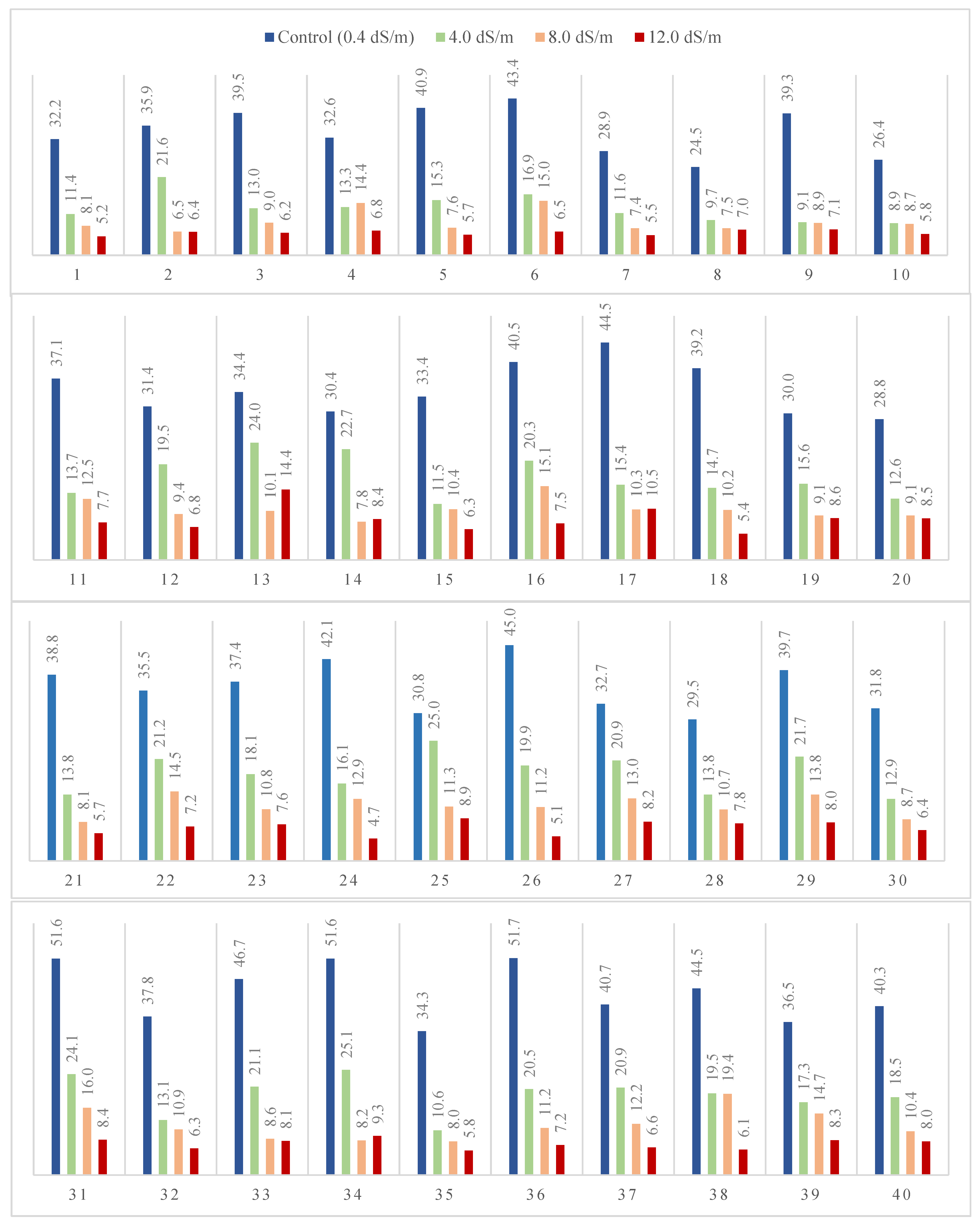
Figure 2 and
Figure 3 and
Table 5 and
Table 6 show the grain yield mean performance of forty genotypes evaluated under four salinity treatments as well as combined data. In general, results indicate that forty genotypes decreased dramatically under salinity treatments in an orderly manner, from control followed by 4.0, 8.0, and 12.0 dS/m. Genotypes 31 and 34 had the highest values (25.0 and 22.6, respectively) in combined data over treatments
Table 5 and
Table 6. Moreover, 8.0 dS/m genotypes, 38 and 31 recorded 19.4 gm and 16.0 gm in comparison with the control. Additionally, genotypes 13 and 17 possess the highest values of 10.46 and 14.4 in 12.0 dS/m treatment.
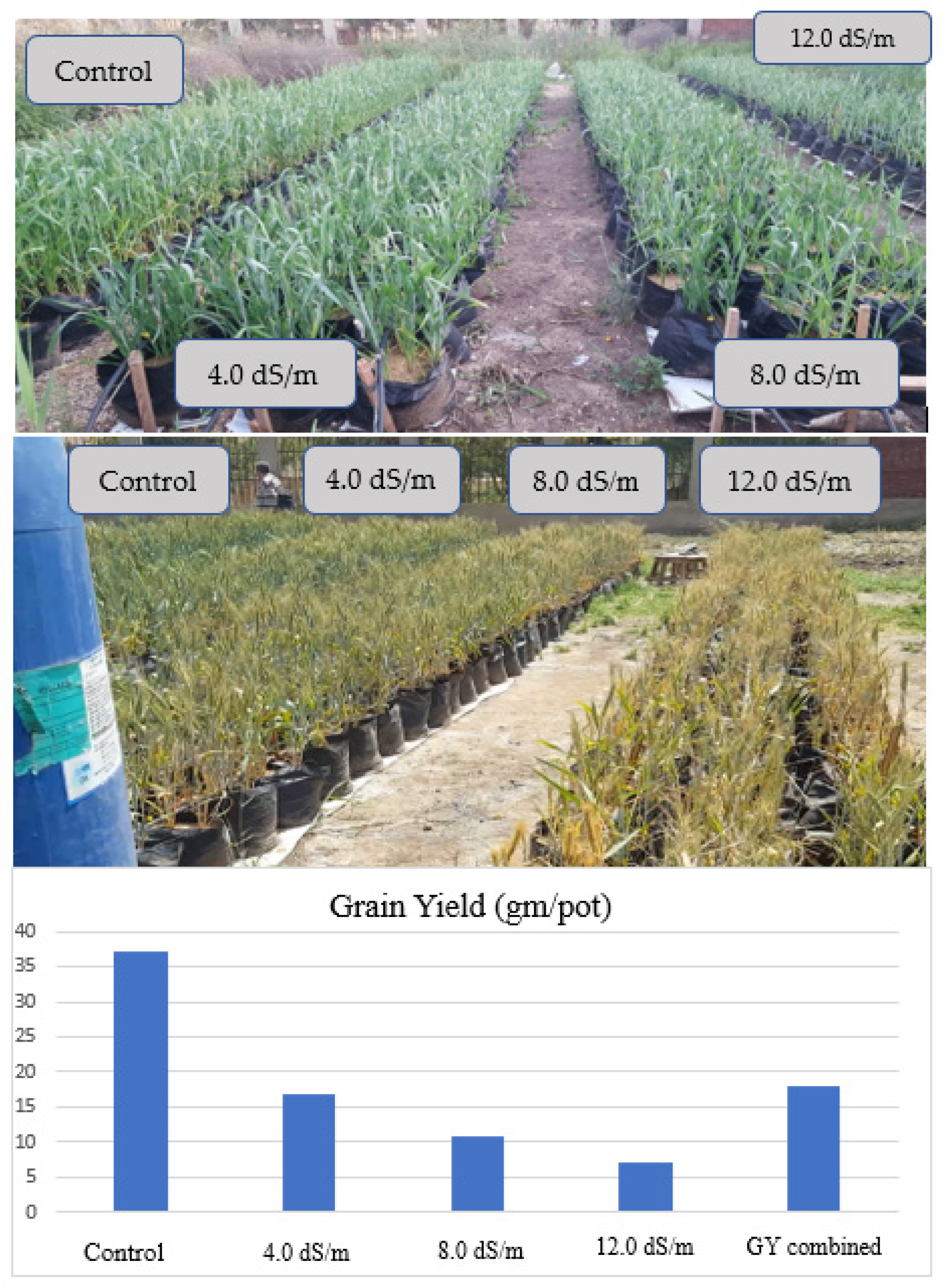
Figure 7 reveals the mean of grain yield with the degradation from control followed by 4.0, 8.0, and 12.0 dS/m of forty genotypes evaluated in the pots experiment in bar plot and pictures, and the combined data analysis across all salinity treatments.
Table 5.
Grain yield of control (GYN), grain yield at 8.0 dS/m (GY8), and salinity tolerance/susceptible indices of forty genotypes in 2019/20 season.
Table 5.
Grain yield of control (GYN), grain yield at 8.0 dS/m (GY8), and salinity tolerance/susceptible indices of forty genotypes in 2019/20 season.
| Genotype | GYN | GY8 | TOL | GMP | STI | MP | HM | SSI |
|---|
| 1 | 32.2† a–d | 8.1 cd | 24.1 | 16.1 | 0.19 | 20.1 | 12.9 | 1.05 |
| 2 | 35.9 a–d | 6.5 d | 29.4 | 15.3 | 0.17 | 21.2 | 11.1 | 1.15 |
| 3 | 39.5 a–d | 9.0 bcd | 30.5 | 18.9 | 0.26 | 24.2 | 14.7 | 1.09 |
| 4 | 32.6 a–d | 14.4 abc | 18.2 | 21.7 | 0.34 | 23.5 | 20.0 | 0.78 |
| 5 | 40.9 a–d | 7.6 cd | 33.3 | 17.6 | 0.22 | 24.2 | 12.8 | 1.15 |
| 6 | 43.4 a–d | 15.0 abc | 28.4 | 25.6 | 0.47 | 29.2 | 22.3 | 0.92 |
| 7 | 28.9 bcd | 7.4 cd | 21.4 | 14.7 | 0.15 | 18.2 | 11.8 | 1.05 |
| 8 | 24.5 d | 7.5 cd | 17.0 | 13.5 | 0.13 | 16.0 | 11.5 | 0.98 |
| 9 | 39.3 a–d | 8.9 bcd | 30.4 | 18.7 | 0.25 | 24.1 | 14.5 | 1.09 |
| 10 | 26.4 cd | 8.7 bcd | 17.8 | 15.1 | 0.16 | 17.6 | 13.1 | 0.95 |
| 11 | 37.1 a–d | 12.5 a–d | 24.6 | 21.6 | 0.33 | 24.8 | 18.7 | 0.93 |
| 12 | 31.4 bcd | 9.4 bcd | 22.0 | 17.2 | 0.21 | 20.4 | 14.5 | 0.99 |
| 13 | 34.4 a–d | 10.1 bcd | 24.3 | 18.6 | 0.25 | 22.2 | 15.6 | 1.00 |
| 14 | 30.4 bcd | 7.8 cd | 22.5 | 15.4 | 0.17 | 19.1 | 12.5 | 1.04 |
| 15 | 33.4 a–d | 10.4 bcd | 23.0 | 18.7 | 0.25 | 21.9 | 15.9 | 0.97 |
| 16 | 40.5 a–d | 15.1 abc | 25.5 | 24.7 | 0.44 | 27.8 | 22.0 | 0.88 |
| 17 | 44.5 abc | 10.3 bcd | 34.2 | 21.4 | 0.33 | 27.4 | 16.8 | 1.08 |
| 18 | 39.2 a–d | 10.2 bcd | 29.0 | 20.0 | 0.29 | 24.7 | 16.2 | 1.04 |
| 19 | 30.0 bcd | 9.1 bcd | 20.9 | 16.5 | 0.20 | 19.6 | 14.0 | 0.98 |
| 20 | 28.8 bcd | 9.1 bcd | 19.7 | 16.2 | 0.19 | 19.0 | 13.8 | 0.96 |
| 21 | 38.8 a–d | 8.1 cd | 30.7 | 17.8 | 0.23 | 23.5 | 13.4 | 1.11 |
| 22 | 35.5 a–d | 14.5 abc | 21.0 | 22.7 | 0.37 | 25.0 | 20.6 | 0.83 |
| 23 | 37.4 a–d | 10.8 bcd | 26.6 | 20.1 | 0.29 | 24.1 | 16.7 | 1.00 |
| 24 | 42.1 a–d | 12.9 a–d | 29.1 | 23.3 | 0.39 | 27.5 | 19.8 | 0.97 |
| 25 | 30.8 bcd | 11.3 bcd | 19.4 | 18.7 | 0.25 | 21.1 | 16.6 | 0.89 |
| 26 | 45.0 abc | 11.2 bcd | 33.8 | 22.4 | 0.36 | 28.1 | 17.9 | 1.06 |
| 27 | 32.7 a–d | 13.0 a–d | 19.7 | 20.7 | 0.31 | 22.9 | 18.6 | 0.85 |
| 28 | 29.5 bcd | 10.7 bcd | 18.7 | 17.8 | 0.23 | 20.1 | 15.7 | 0.89 |
| 29 | 39.7 a–d | 13.8 a–d | 25.9 | 23.4 | 0.39 | 26.8 | 20.5 | 0.92 |
| 30 | 31.8 bcd | 8.7 bcd | 23.1 | 16.6 | 0.20 | 20.2 | 13.6 | 1.02 |
| 31 | 51.6 a | 16.0 ab | 35.5 | 28.8 | 0.59 | 33.8 | 24.5 | 0.97 |
| 32 | 37.8 a–d | 10.9 bcd | 27.0 | 20.3 | 0.30 | 24.4 | 16.9 | 1.00 |
| 33 | 46.7 ab | 8.6 bcd | 38.1 | 20.1 | 0.29 | 27.7 | 14.6 | 1.15 |
| 34 | 51.6 a | 8.2 bcd | 43.4 | 20.6 | 0.31 | 29.9 | 14.2 | 1.18 |
| 35 | 34.3 a–d | 8.0 cd | 26.3 | 16.6 | 0.20 | 21.2 | 13.0 | 1.08 |
| 36 | 51.7 a | 11.2 bcd | 40.5 | 24.1 | 0.42 | 31.5 | 18.4 | 1.10 |
| 37 | 40.7 a–d | 12.2 a–d | 28.5 | 22.3 | 0.36 | 26.4 | 18.8 | 0.99 |
| 38 | 44.5 abc | 19.4 a | 25.2 | 29.4 | 0.62 | 31.9 | 27.0 | 0.80 |
| 39 | 36.5 a–d | 14.7 abc | 21.8 | 23.1 | 0.38 | 25.6 | 20.9 | 0.84 |
| 40 | 40.3 a–d | 10.4 bcd | 29.8 | 20.5 | 0.30 | 25.4 | 16.6 | 1.04 |
Table 6.
Grain yield of control (GYN), grain yield at 12.0 dS/m (GY12) treatment, and salinity tolerance/susceptible indices along with grain yield combined data (GYC) of forty genotypes in 2019/20 season.
Table 6.
Grain yield of control (GYN), grain yield at 12.0 dS/m (GY12) treatment, and salinity tolerance/susceptible indices along with grain yield combined data (GYC) of forty genotypes in 2019/20 season.
| Genotype | GYN | GY12 | TOL | GMP | STI | MP | HM | SSI | GYC |
|---|
| 1 | 32.2† a–d | 5.2 c | 27.0 | 12.9 | 0.12 | 18.7 | 9.0 | 1.04 | 14.2 f–i |
| 2 | 35.9 a–d | 6.4 bc | 29.5 | 15.2 | 0.17 | 21.2 | 10.9 | 1.02 | 17.6 b–i |
| 3 | 39.5 a–d | 6.2 bc | 33.3 | 15.6 | 0.18 | 22.8 | 10.7 | 1.05 | 16.9 c–i |
| 4 | 32.6 a–d | 6.8 bc | 25.8 | 14.9 | 0.16 | 19.7 | 11.2 | 0.98 | 16.8 c–i |
| 5 | 40.9 a–d | 5.7 bc | 35.2 | 15.2 | 0.17 | 23.3 | 10.0 | 1.07 | 17.4 c–i |
| 6 | 43.4 a–d | 6.5 bc | 36.9 | 16.8 | 0.20 | 25.0 | 11.4 | 1.05 | 20.5 a–e |
| 7 | 28.9 bcd | 5.5 bc | 23.3 | 12.6 | 0.11 | 17.2 | 9.3 | 1.00 | 13.4 ghi |
| 8 | 24.5 d | 7.0 bc | 17.4 | 13.1 | 0.12 | 15.8 | 10.9 | 0.88 | 12.2 i |
| 9 | 39.3 a–d | 7.1 bc | 32.2 | 16.7 | 0.20 | 23.2 | 12.0 | 1.02 | 16.1 d–i |
| 10 | 26.4 cd | 5.8 bc | 20.6 | 12.4 | 0.11 | 16.1 | 9.6 | 0.97 | 12.5 hi |
| 11 | 37.1 a–d | 7.7 bc | 29.4 | 16.9 | 0.21 | 22.4 | 12.8 | 0.98 | 17.8 b–i |
| 12 | 31.4 bcd | 6.8 bc | 24.6 | 14.6 | 0.15 | 19.1 | 11.1 | 0.97 | 16.8 c–i |
| 13 | 34.4 a–d | 14.4 a | 20.0 | 22.5 | 0.36 | 24.4 | 20.3 | 0.72 | 20.7 a–e |
| 14 | 30.4 bcd | 8.4 bc | 22.0 | 15.9 | 0.18 | 19.4 | 13.1 | 0.90 | 17.3 c–i |
| 15 | 33.4 a–d | 6.3 bc | 27.1 | 14.5 | 0.15 | 19.9 | 10.6 | 1.01 | 15.4 d–i |
| 16 | 40.5 a–d | 7.5 bc | 33.1 | 17.4 | 0.22 | 24.0 | 12.6 | 1.01 | 20.8 a–e |
| 17 | 44.5 abc | 10.5 ab | 34.0 | 21.6 | 0.33 | 27.5 | 17.0 | 0.95 | 20.2 a–f |
| 18 | 39.2 a–d | 5.4 bc | 33.9 | 14.5 | 0.15 | 22.3 | 9.4 | 1.07 | 17.4 b–i |
| 19 | 30.0 bcd | 8.6 bc | 21.4 | 16.0 | 0.18 | 19.3 | 13.3 | 0.89 | 15.8 d–i |
| 20 | 28.8 bcd | 8.5 bc | 20.3 | 15.7 | 0.18 | 18.7 | 13.2 | 0.87 | 14.8 e–i |
| 21 | 38.8 a–d | 5.7 bc | 33.1 | 14.9 | 0.16 | 22.3 | 10.0 | 1.06 | 16.6 c–i |
| 22 | 35.5 a–d | 7.2 bc | 28.3 | 15.9 | 0.18 | 21.3 | 11.9 | 0.99 | 19.6 a–f |
| 23 | 37.4 a–d | 7.6 bc | 29.8 | 16.9 | 0.20 | 22.5 | 12.6 | 0.99 | 18.5 b–h |
| 24 | 42.1 a–d | 4.7 c | 37.4 | 14.0 | 0.14 | 23.4 | 8.4 | 1.10 | 19.0 a–g |
| 25 | 30.8 bcd | 8.9 bc | 21.9 | 16.5 | 0.20 | 19.8 | 13.8 | 0.88 | 19.0 a–g |
| 26 | 45.0 abc | 5.1 c | 39.9 | 15.1 | 0.16 | 25.1 | 9.2 | 1.10 | 20.3 a–f |
| 27 | 32.7 a–d | 8.2 bc | 24.6 | 16.4 | 0.19 | 20.5 | 13.1 | 0.93 | 18.7 b–g |
| 28 | 29.5 bcd | 7.8 bc | 21.7 | 15.2 | 0.17 | 18.6 | 12.3 | 0.91 | 15.5 d–i |
| 29 | 39.7 a–d | 8.0 bc | 31.7 | 17.8 | 0.23 | 23.9 | 13.3 | 0.99 | 20.8 a–e |
| 30 | 31.8 bcd | 6.4 bc | 25.4 | 14.3 | 0.15 | 19.1 | 10.7 | 0.99 | 14.9 d–i |
| 31 | 51.6 a | 8.4 bc | 43.2 | 20.8 | 0.31 | 30.0 | 14.5 | 1.04 | 25.0 a |
| 32 | 37.8 a–d | 6.3 bc | 31.5 | 15.5 | 0.17 | 22.1 | 10.9 | 1.03 | 17.0 c–i |
| 33 | 46.7 ab | 8.1 bc | 38.6 | 19.5 | 0.27 | 27.4 | 13.8 | 1.03 | 21.1 a–d |
| 34 | 51.6 a | 9.3 abc | 42.3 | 21.9 | 0.35 | 30.5 | 15.8 | 1.02 | 23.6 ab |
| 35 | 34.3 a–d | 5.8 bc | 28.5 | 14.2 | 0.14 | 20.1 | 10.0 | 1.03 | 14.7 e–i |
| 36 | 51.7 a | 7.2 bc | 44.5 | 19.3 | 0.27 | 29.4 | 12.6 | 1.07 | 22.6 abc |
| 37 | 40.7 a–d | 6.6 bc | 34.1 | 16.4 | 0.19 | 23.6 | 11.4 | 1.04 | 20.1 a–f |
| 38 | 44.5 abc | 6.1 bc | 38.5 | 16.4 | 0.19 | 25.3 | 10.7 | 1.07 | 22.4 abc |
| 39 | 36.5 a–d | 8.3 bc | 28.2 | 17.4 | 0.22 | 22.4 | 13.5 | 0.96 | 19.2 a–g |
| 40 | 40.3 a–d | 8.0 bc | 32.2 | 18.0 | 0.23 | 24.2 | 13.4 | 0.99 | 19.3 a–g |
Figure 2.
Wheat grain yield gm per pot (GY) of forty genotypes evaluated in pot and grown under 0.4, 4.0, 8.0, or 12.0 dS/m of seawater in the 2019/20 season.
Figure 2.
Wheat grain yield gm per pot (GY) of forty genotypes evaluated in pot and grown under 0.4, 4.0, 8.0, or 12.0 dS/m of seawater in the 2019/20 season.
Figure 3.
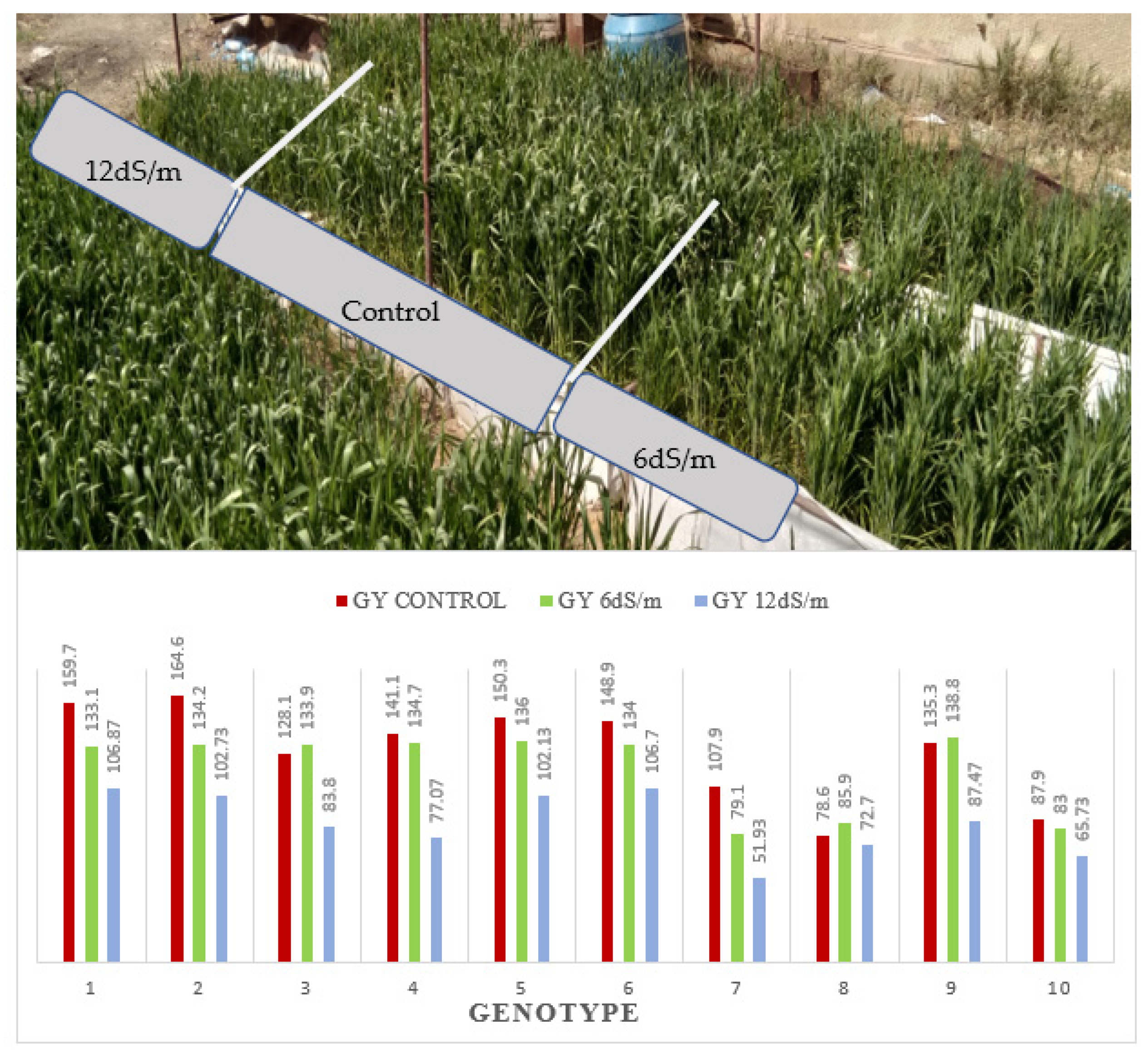
Bar chart of wheat genotypes grain yield gm per pot (GY) grown under 0.4, 4.0, 8.0, and 12.0 dS/m diluted seawater, and combined data over forty genotypes and photos representing the visual differences among the four saline treatments in 2019/20 season.
Figure 3.
Bar chart of wheat genotypes grain yield gm per pot (GY) grown under 0.4, 4.0, 8.0, and 12.0 dS/m diluted seawater, and combined data over forty genotypes and photos representing the visual differences among the four saline treatments in 2019/20 season.
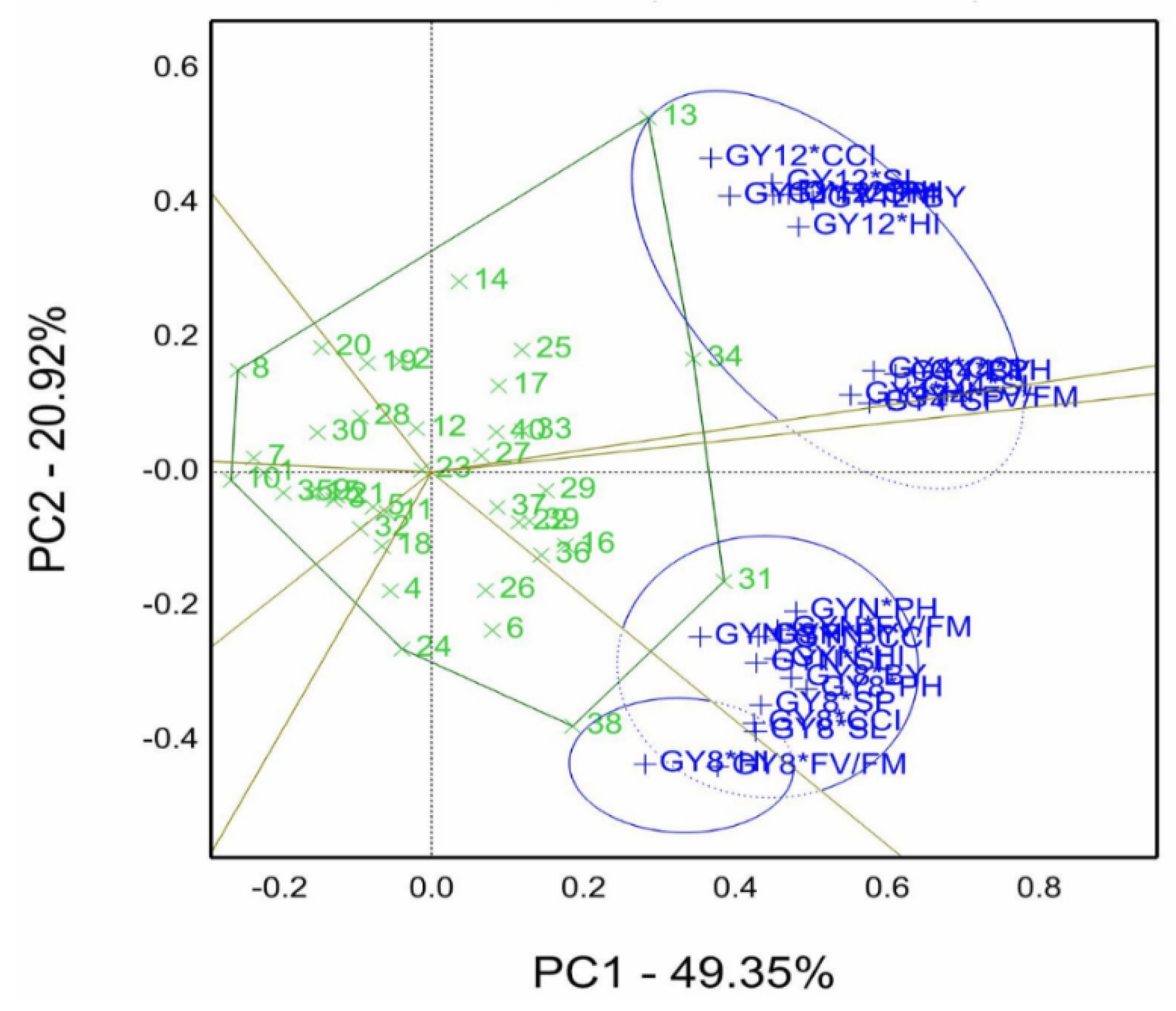
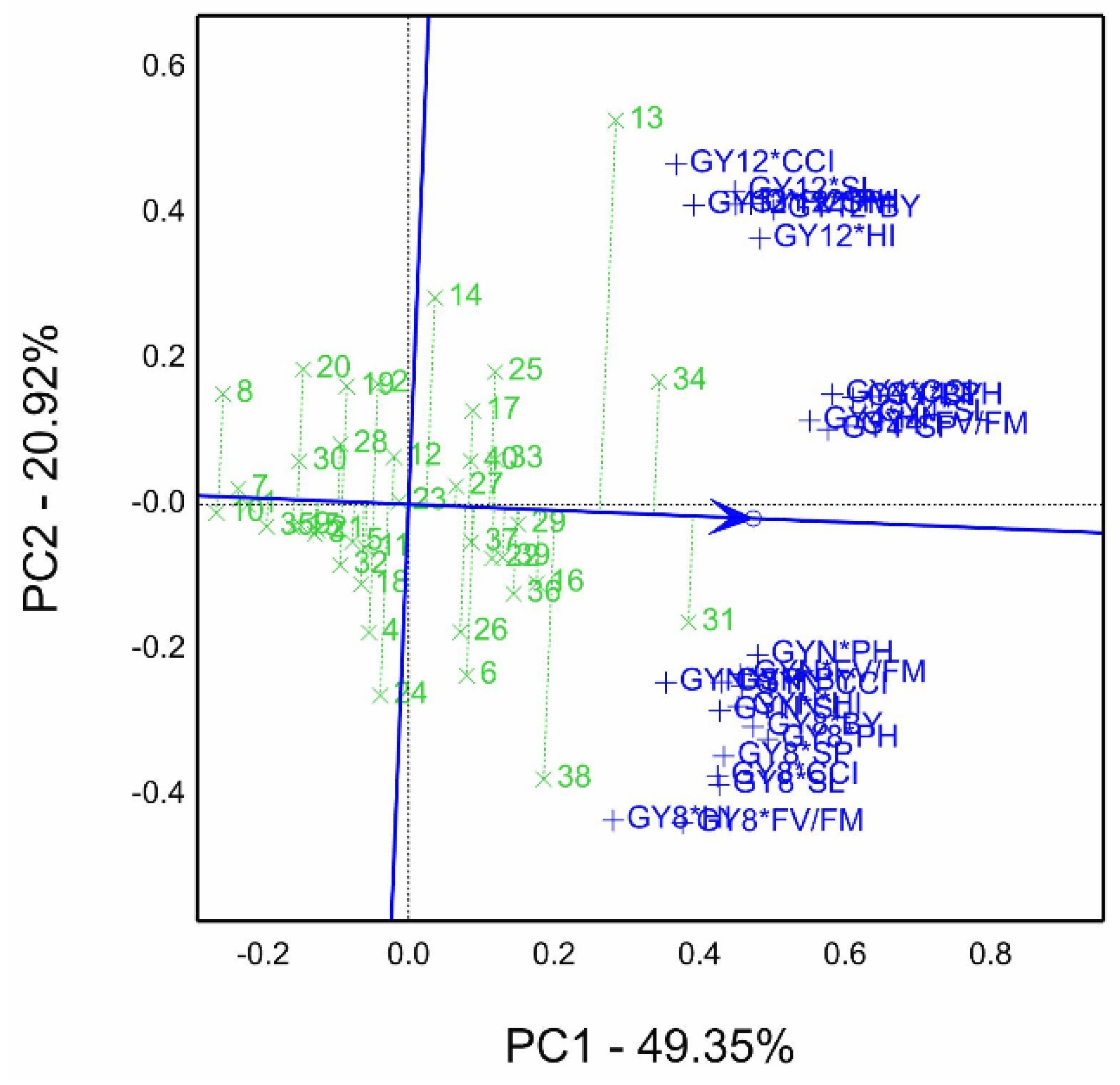
The combined means of genotypes by grain yield by all trait combinations (GY*T) evaluated under four salinity conditions (pot experiments) in the 2019/20 season is illustrated in
Figure 4. Data were normalized to perform the GYT biplot, which accounted for 70.3% of total variation under four salinity levels. In the GYT biplot, the figure is divided into seven sectors, and the sector contains genotypes 13 and 34 on the polygon vertexes winner for GY12*CCI, HI, SL, BY, and GY4*BY, PH, SP, and Fv/Fm combinations. While genotype 31 is situated on the vertex of another sector with GYN*PH, CCI, Fv/Fm, BY, SL, and HI, and GY8*BY, PH, SP, SL, CCI. In the sector of genotypes 38 and 24 with combinations of GY8*HI and Fv/Fm.
Genotypes ranking based on GYT data is revealed in
Figure 5. The order of genotype 31 > 34 > 13 > 38 > 16 > 36 > 29 joint with projection on average tester coordination (ATC blue line with arrow) and over grand mean (vertical line on ATC). However, genotypes 10 and 8 had the lowest performance of GYT combinations. Additionally, genotypes 31 and 34 had the best mean performance and stability; thus, they are close to the ATC line and combine the yield with all trait combinations (GY*T) evaluated under four salinity levels (pot experiments) in 2019/20 season. However, genotypes 13 and 38 are far away from ATC. Thus, they are unstable genotypes that rely on the GYT pattern of biplots.
Salinity tolerance/susceptible indices (STI) calculated using grain yield gm per pot (GY) of control at 0.4 dS/m (GYN), grain yield at 8.0 dS/m (GY 8.0 dS/m), and grain yield at 12.0 dS/m (GY 12.0 dS/m) are shown in
Table 3. In this study, data in
Table 3 were used to estimate the graphical genotypes by traits model (GT) in
Figure 6.
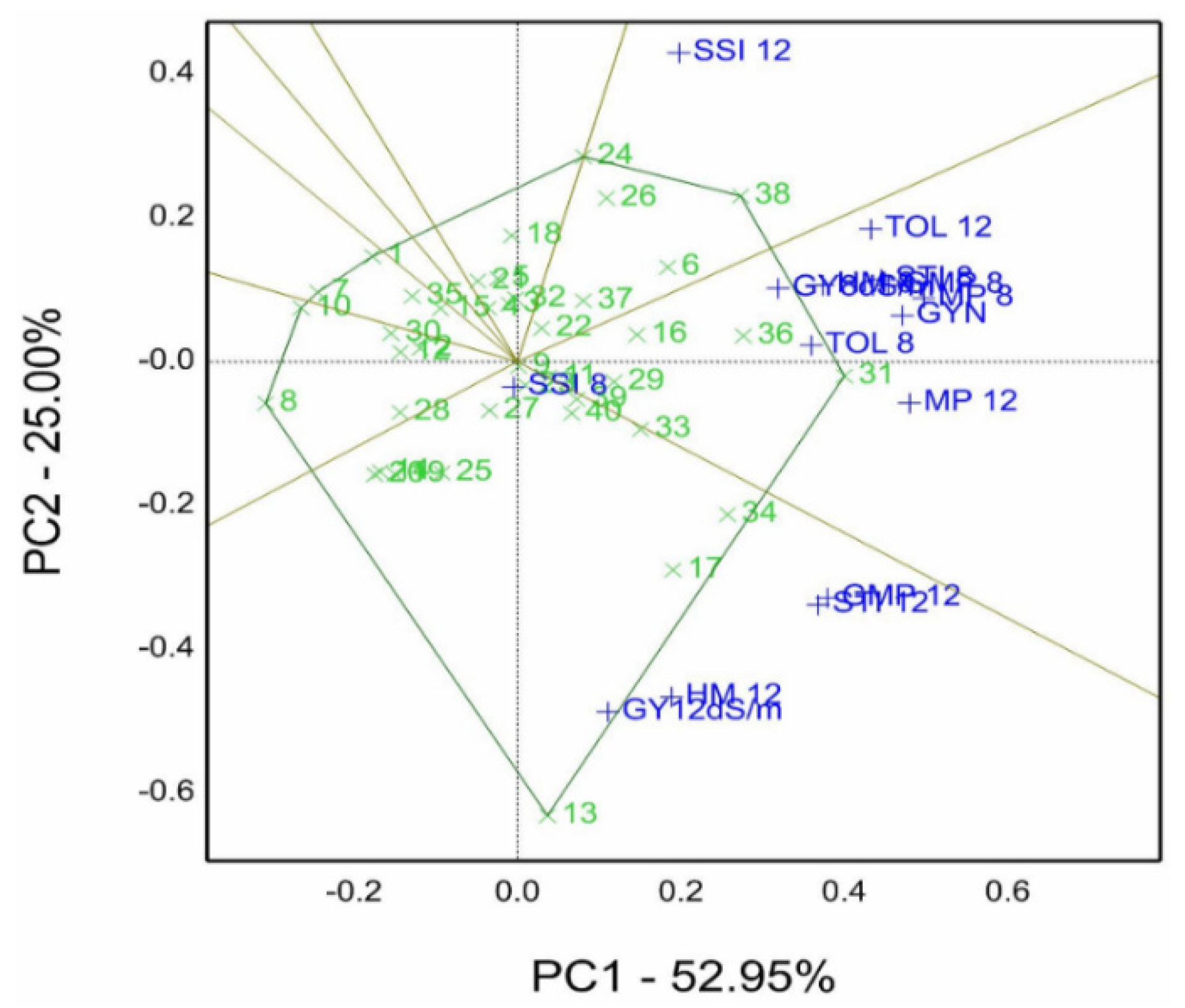
The sum of principal components PC1 and PC2 is 79.41% of the total variation of forty genotypes combined with STI, which is represented in figures in the titled genotype by salinity tolerance indices (GSTI) biplot. The findings helped to select genotypes 31 and 36, which are pointed on the vertex of the polygon in the sector containing GSTI, such as grain yield gm per pot of normal treatment (GYN), TOL8 (for 8.0 dS/m), TOL12 (for 12.0 dS/m), MP8, STI8, GMP8, MP12, STI12, HM8, and GY8. Additionally, genotype 13 is the winner of GY12, HM12, and GMP12 in its sector, but genotypes 24 and 26 situated in the sector had SSI8 and SSI12 as sensitive indies/genotypes.
Means vs stability based on GSTI analysis divided from values in
Table 5 and
Table 6 and 6 of forty genotypes treated by four diluted seawaters in the 2019/20 season in pot trials, illustrated visually in
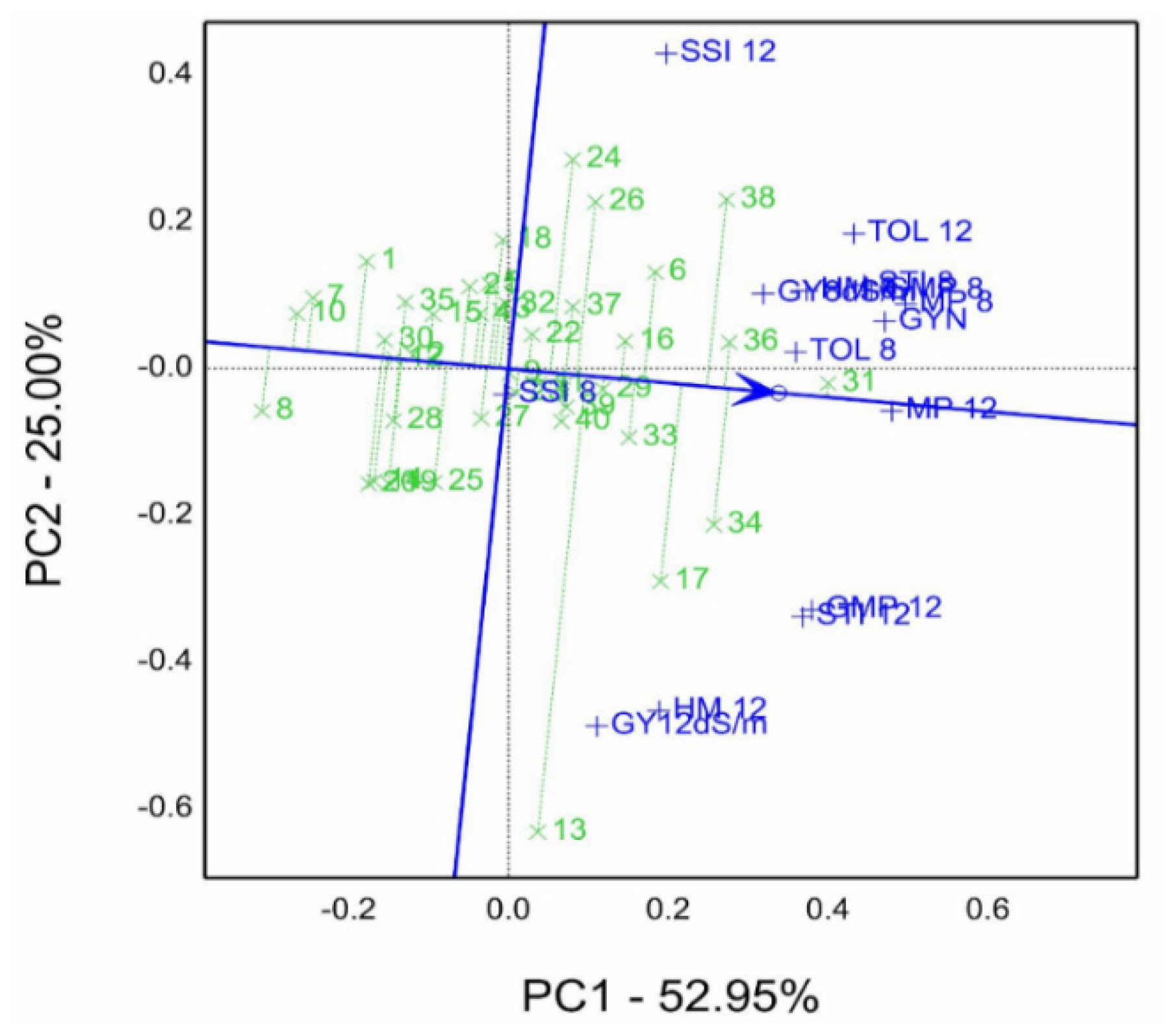
Figure 7. Genotype 31, followed by 36, 34, 38, 17, 33, 6, and 16, recorded a high rank and stability for salinity tolerance indices. In contrast, genotype 8 had the lowest mean, and genotype 13 possessed the lowest stability referring to its long projection on ATC.
Figure 7.
Mean vs. stability (GSTI) biplot of ranking genotypes based on grain yield gm per pot (GYN), grain yield gm per pot (GY 8.0, 12.0 dS/m), and salinity tolerance/susceptible indices (calculated at control with 0.4 dS/m, 8.0 dS/m, and 12.0 dS/m) of forty genotypes in the 2019/20 season. TOL, tolerance index; MP, mean productivity; STI, stress tolerance index; GMP geometric mean productivity; HM, harmonic mean; SSI, stress susceptibility index.
Figure 7.
Mean vs. stability (GSTI) biplot of ranking genotypes based on grain yield gm per pot (GYN), grain yield gm per pot (GY 8.0, 12.0 dS/m), and salinity tolerance/susceptible indices (calculated at control with 0.4 dS/m, 8.0 dS/m, and 12.0 dS/m) of forty genotypes in the 2019/20 season. TOL, tolerance index; MP, mean productivity; STI, stress tolerance index; GMP geometric mean productivity; HM, harmonic mean; SSI, stress susceptibility index.
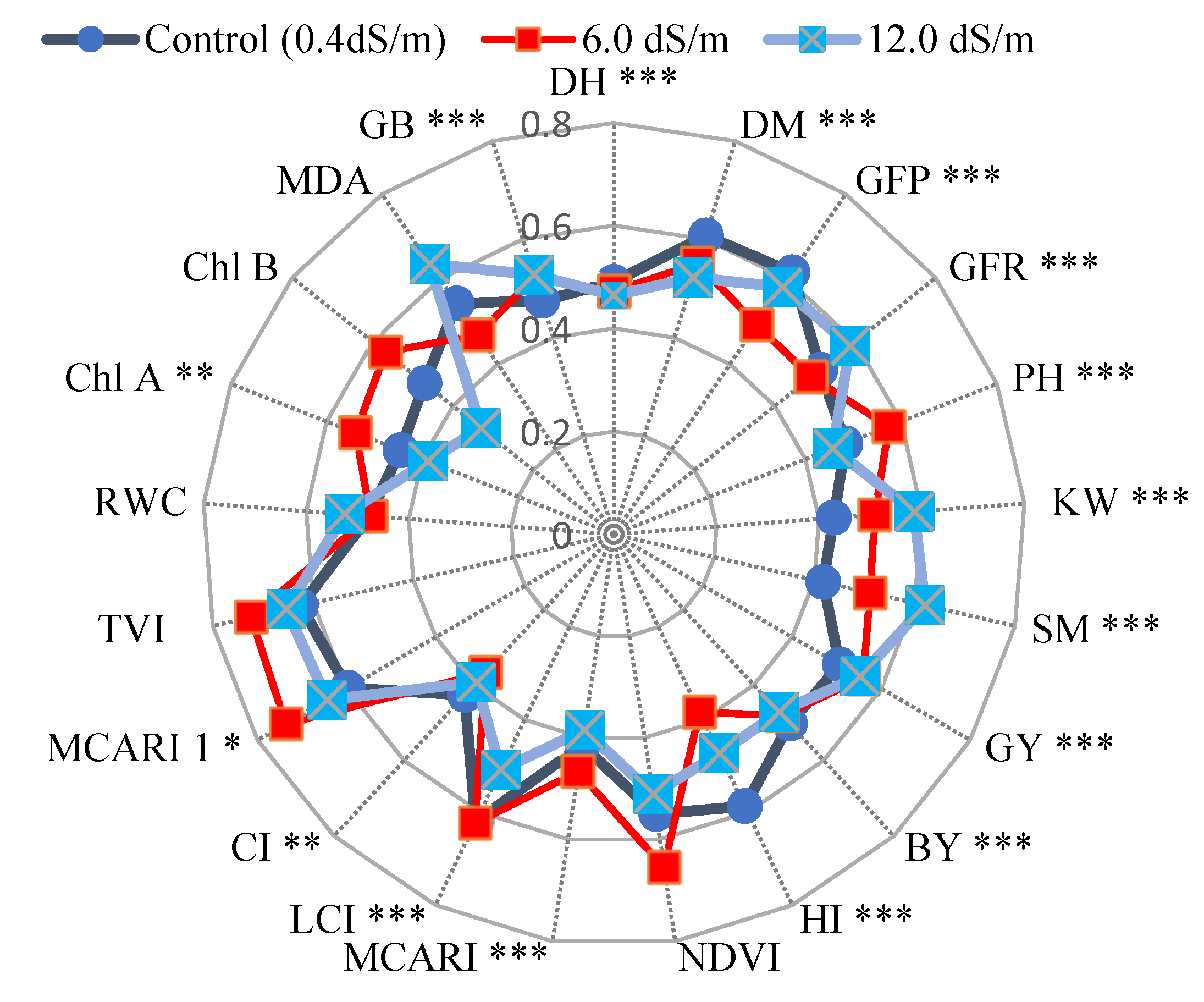
Agronomic and physiological traits, along with spectral reflectance indices (SRI), data were normalized to remove the effect of different units, in order to compare an averaged ten genotypes across three levels of salinity (seawater mitigation), e.g., control, 6.0 dS/m, and 12.0 dS/m.
Figure 8 shows that traits such as GY, BY, DH, GB, CI, and RWC recorded almost the same mean performance under three salinity levels. However, traits like KW, SM, GFR, TVI, and MCARI possess high mean values of 6.0 dS/m and 12.0 dS/m in comparison with the control. Additionally, control of traits DM, GFP, and HI tend to be higher than 6.0 dS/mor 12 dS/m. In contrast, control of traits Chl A, B, PH, and MDA is situated in between other treatments. The genotype component of the source of variation for all traits varied differences significantly (
p < 0.01), with the exception of TVI1, RWC, Chl B, NDVI, shown in
Table 7 and
Figure 8.
Figure 9 and
Table 8 show the grain yield gm per row (GY) mean performance of ten genotypes evaluated in a simulated lysimeter and irrigated with 0.4, 6.0, and 12.0 dS/m diluted seawater. The data represent genotypes classified into two categories, i.e., genotypes 1, 2, 3, 4, 5, 6, and 9 (Misr 4) recorded as high performance, while the old varieties, i.e., genotypes 7 (Kharchia 65), 8 (Oasis F86), and 10 (Sakha 8) under 3 saline irrigation were not. In addition, in the combined analysis, genotypes, 1 and 2 recorded the highest values of 133.3 and 133.8 g, respectively. However, varieties 7, 8, and 10 had 79.6, 79.1, and 78.9, respectively, shown in
Table 8.
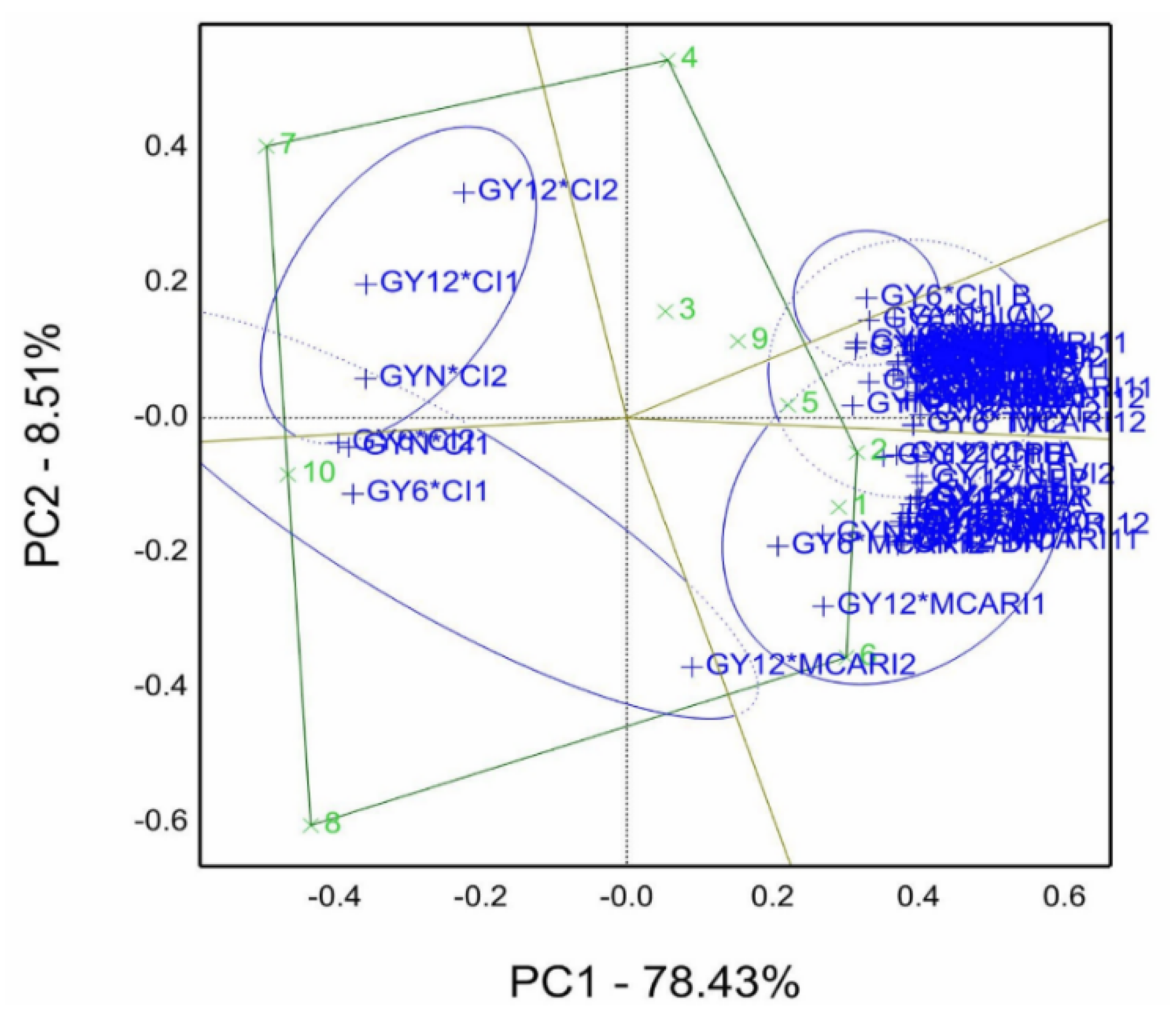
Genotype by grain yield*traits (GYT) pattern was used to select the best one from ten genotypes evaluated in a simulated lysimeter in the 2020/21 season illustrated in
Figure 10. The GYT biplot accounted for 86.94% of the total variation. Moreover, the sector of genotypes 2 and 6 had the most combinations of agronomic and physiological traits and spectral reflectance indices. The sector of genotypes 8 and 10 had combinations of GY6*CI1, CI2, and GYN*CI1, while the sector of genotype 7 contains GYN*CI2, GY12*CI1, and CI2. Thus, the best genotypes that rely on this view are genotypes 2 and 6.
Based on the GYT model graphical analysis, ten entries were evaluated under three salinity treatments by simulated lysimeter in the 2020/21 season, as shown in
Figure 11. The genotypes rank is 2 > 6>1 > 5>9 > 4>8 > 10 > 7 joint with projection on average tester coordination (ATC blue line with arrow). Additionally, genotypes 2, 1, and 5 would be the best mean performance and stability; thus, they are close to the ATC line and combine the yield with all trait combinations (GY*T). However, genotypes 6 and 4 are far away from the ATC, thus they are unstable genotypes.
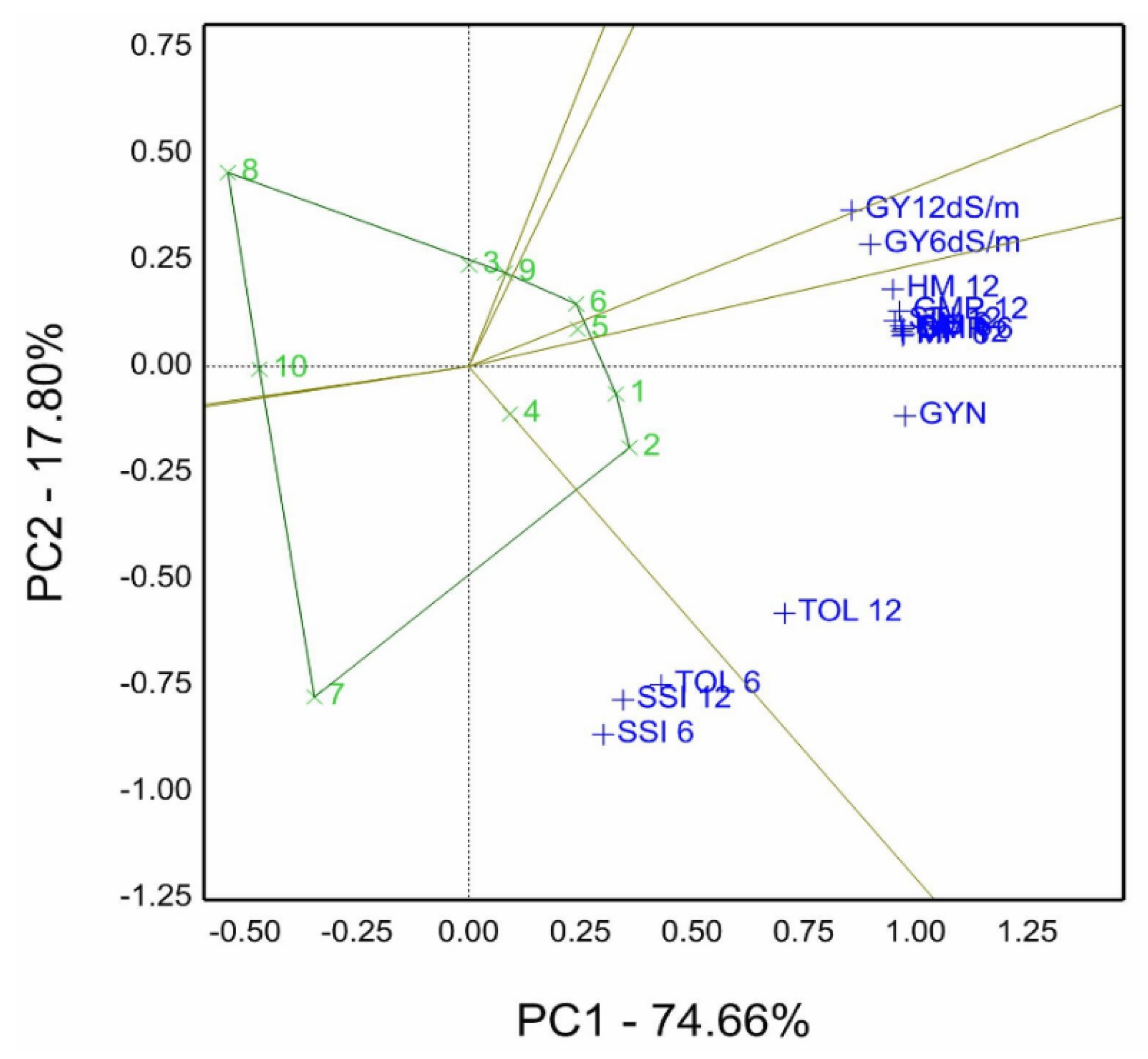
Relying on the genotype by trait (GT) biplot pattern, we compute salinity tolerance/sensitive indices (STI) using grain yield of control gm per row (GYN), grain yield of 6.0 dS/m (GY6 dS/m), grain yield of 12.0 dS/m gm per row (GY12 dS/m), and another STI of ten genotypes in
Table 4. In this study, we performed data in
Table 4 to depict graphical genotypes by traits model (GT) in
Figure 12. The sum of principal components was 93.11% of the total variation for ten genotypes combined with STI, represented in figures termed genotype by salinity tolerance indices (GSTI) biplot. The findings indicate that genotypes 1 and 2 are located on the polygon’s vertex in its sector containing GSTI combinations such as grain yield normal gm per row (GYN), TOL12 (for 12.0 dS/m), HM12, MP6, STI6, GMP6, MP12, STI12, HM6, GMP12, and GYC combined. Additionally, genotype 7 is the winner of TOL6 (for 6.0 dS/m), SSI6, and SSI12. However, genotype 6 is the winner in its sector for GY12 dS/m.
Based on the means vs stability GSTI biplot, ten entries evaluated under three salinity treatments by simulated lysimeter in the 2020/21 season are illustrated in
Figure 13. The rank commences from genotype 2 followed by 1, 5, 6, and 4 placed on the average tester coordination line (ATC blue line with an arrow). Additionally, genotypes 2 and 1 would show the best mean performance and stability. However, genotypes 7 and 8 are far away from the ATC; thus, they are unstable genotypes.
4. Discussion
The genotype by yield*trait (GYT) biplots approach, as proposed by Yan and Frégeau–Reid [
46], graphically identified the relationship between genotypes and studied traits, specifically, in the case of large genotype numbers and characters. For example, in salinity and drought stress conditions with contrasting regimes. Additionally, these methodologies can be used by breeders to select the elite genotypes not only had grain yield performance but also combined with other trait combinations. For instance, the selection for barley drought tolerance reported by [
47,
49], wheat rusts resistance selected using genotype*traits (GT) technique by the researchers [
48], the authors [
65] selected common bean genotypes using the GYT model, and the researchers identified durum wheat drought tolerance evaluated in multi-years under rainfed conditions using GYT [
51]. In our study, we evaluated forty bread wheat genotypes irrigated by four diluted seawater levels in the pot experiments to select six genotypes (i.e., 6, 16, 31, 33, 34, 36) in the first season based on results shown in
Figure 2 and
Figure 6. To evaluate wheat genotypes grown under three saline levels, lysimeter experiments were conducted in the second season to compare the performance of 10 wheat genotypes and varieties, i.e., three old varieties i.e., 7, 8, 10 (Kharchia 65, Oasis F86, and Sakha 8, respectively) along with new cultivar genotype 9 (Misr 4) for comparison with 6 other wheat genotypes. Genotypes 2 and 1 were selected, which might be salinity tolerance genotypes, as the outcome of these experiments are shown in
Figure 11 and
Figure 13.
The wheat grain-filling stage provides the most informative measurements using spectral reflectance indices (SRIs) [
35]. Thus, more genotypic variation was detected than for early growth vegetation and heading [
35]. On the contrary, ref. [
9] reported significant differences in genotypes, salinity treatments, growth stages, years, and their interactions for all vegetation SRIs and water SRIs. The SRI scores detected significant differences for quinoa genotypes and contrasting water regimes [
27]. Nonetheless, the SRIs in one of the locations studied (irrigated and non-irrigated) did not differ among genotypes, which may refer to the closeness of the plants ripening. In our study, reflectance indices such as MACRI 1, LCI1, and CI 1 in the first measurement, as well as all SRIs in the second reading, recorded that the genotypes significantly varied (
Table 7). The results of the combination of GYT with SRIs in
Figure 10 and
Figure 11 confirmed that genotypes 2 and 1 are the best salinity tolerance genotypes from this view and GSTI view of salinity tolerance indices in
Figure 12 and
Figure 13. In this study, we used GT and GYT approaches to select the superior genotypes not only based on grain yield but also other physiological traits such as amino acid glycine betaine and chlorophyll fluorescence (Fv/Fm) and chlorophyll content index (CCI), as well as agronomic traits such as PH, HI, and BY. These findings are in agreement with the results described by other reports [
9,
27,
66,
67].
In the 2019/20 season, forty genotypes were evaluated together under four salinity treatments. This large number might have created significant variations by interaction with treatments, interpreted by the coefficient of variation (CV) of GY (32.4%), as shown in
Table 4. Thus, in the second season we reduced the number to ten genotypes but added the old varieties, i.e., 7, 8, 10 (Kharchia65, Oasis F86, and Sakha 8, respectively), whose low achieved GY might refer to their yield potential, even with rusts fungicide applied (
Figure 9;
Table 7). The approach of GYT was used with agronomic and physiological traits and spectral reflectance indices to select the best genotypes.
Figure 3 and
Figure 10 identified more than thirteen combinations for genotype 31 and the other combinations situated in the sector of genotypes 34 and 13 (however, 13 was unstable, as shown in
Figure 5). Moreover, genotypes 2 and 6 in the second season had several combinations of spectral reflectance indices (SRI) (
Figure 10). These results were similar to those found by others [
66,
68,
69].
Additionally, GT models suggested by [
45] were used to identify and rank genotypes based on salinity tolerance/susceptible indices (GSTI), genotypes 6, 16, 31, 33, 34, and 36 were selected in the first season in pot experiments, and genotypes 2 and 1 were selected in lysimeter experiments as superior genotypes in the second season (
Figure 7 and
Figure 13). However, it is hard to select genotypes 13 and 8, as shown in
Figure 7, as well as genotypes 7 and 8, shown in
Figure 13, which are unstable and had low performance. These findings are consistent with others [
50] who performed the GT technique to select the elite rice entry [
13] using principal component analyses (PCA). Moreover, other reports [
49] used genotype by yield stress index (GYSI) to select the best genotypes that had drought tolerance.
Both experiments used a radar chart to examine the studied characters’ relationship and genotype means. For example, GY and HI were gradually affected by salinity treatment (
Figure 1 and
Figure 8). Additionally, genotypes varied significantly for all traits in the first season (
Figure 1 and
Table 7). All agronomic, physiological traits, and spectral reflectance indices, are shown in
Figure 8 and
Table 7. Similarly, findings of nitrogen starvation treatments did not impact on Fv/Fm ratio [
70]. While the same trend of GY is affected positively by sowing depth [
71]. Other reports [
72] stated that genotype clusters had varied responses to salinity levels in several traits, such as SL. Additionally, water stress treatment benefited winter wheat growth using subsurface drip irrigation of GY, HI and BY in the second season [
72,
73].





 ,
, 


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}












