
Effects of Plant Growth-Promoting Rhizobacteria on the Physioecological Characteristics and Growth of Walnut Seedlings under Drought Stress
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Screening and Identification
2.2. Basic Soil and Plant Materials
2.3. Experimental Design
2.4. Data Collection and Determinations
2.5. Statistical Analyses
3. Results
3.1. Physioecological Characteristics Measured in Leaves
3.2. Endogenous Hormone Levels in Leaves
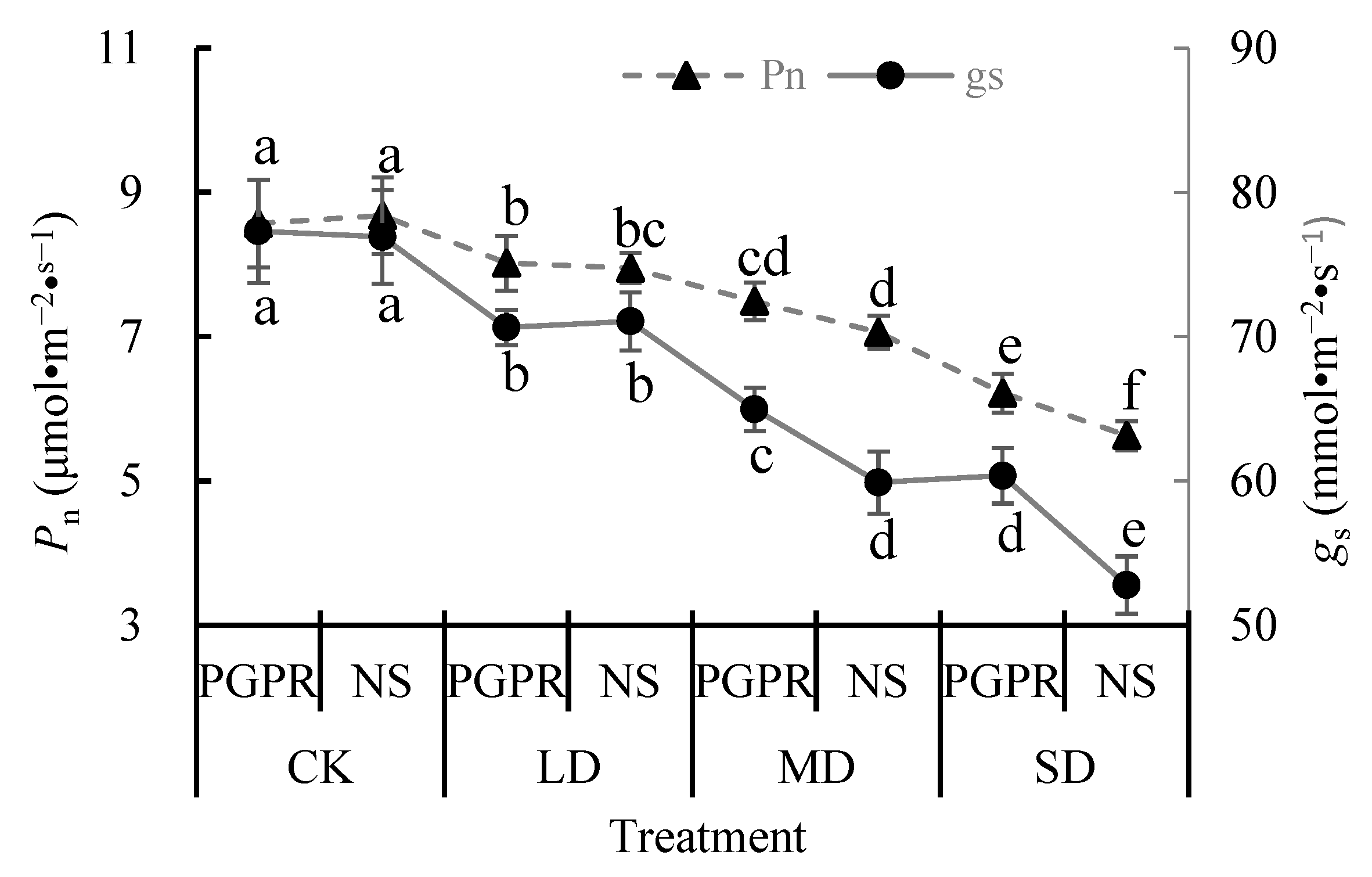
3.3. Photosynthetic Characteristics
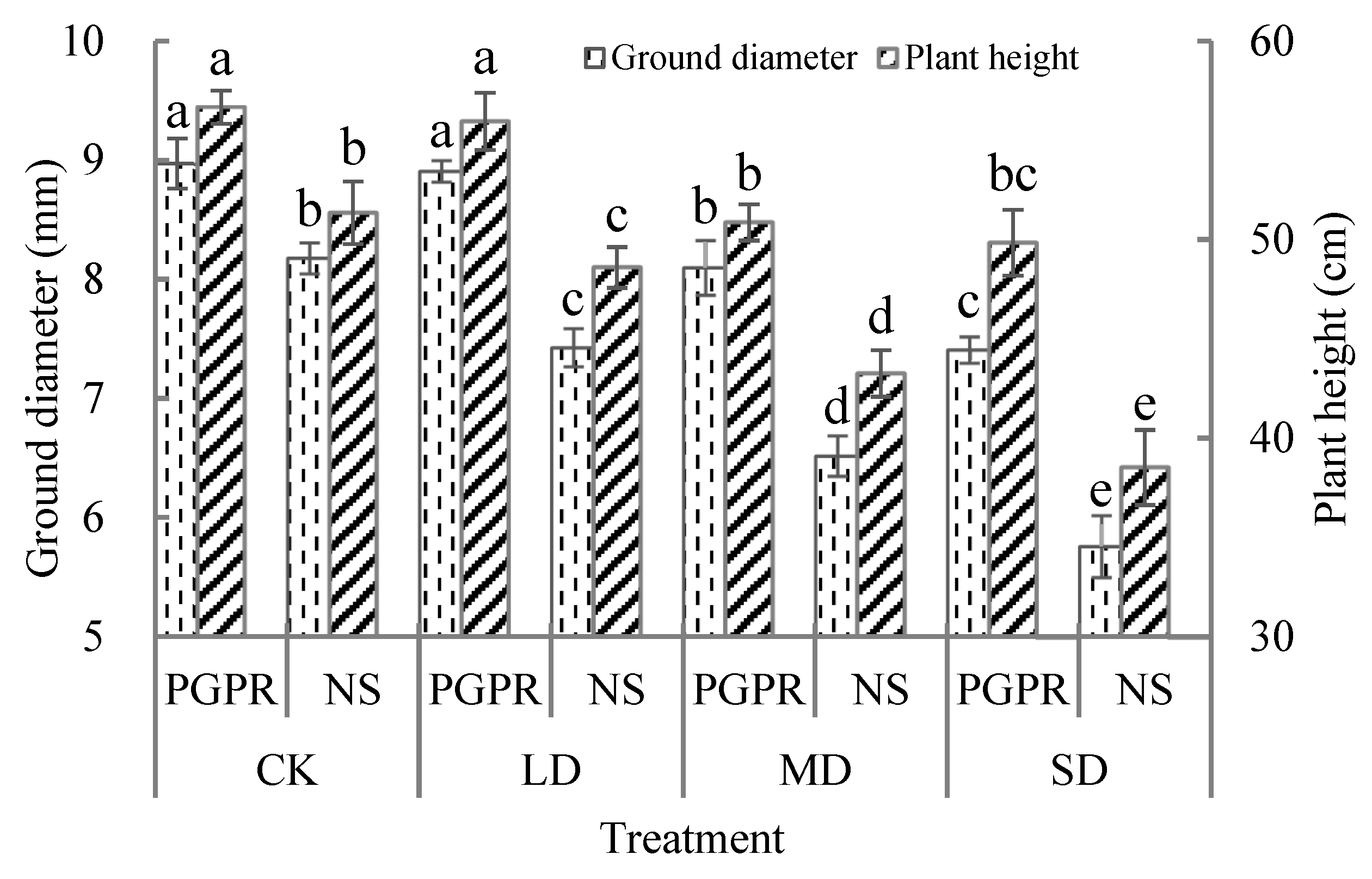
3.4. Ground Diameter and Plant Height
4. Discussion
4.1. Regulatory Enzymes in Leaves
4.2. Endogenous Hormones in Leaves
4.3. Photosynthetic Characteristics and Walnut Seedling Growth
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vadez, V.; Berger, J.D.; Warkentin, T.; Asseng, S.; Ratnakumar, P.; Rao, K.P.C.; Gaur, P.M.; Munier-Jolain, N.; Larmure, A.; Voisin, A.-S.; et al. Adaptation of grain legumes to climate change: A review. Agron. Sustain. Dev. 2011, 32, 31–44. [Google Scholar] [CrossRef]
- Wallace, J. Increasing agricultural water use efficiency to meet future food production. Agric. Ecosyst. Environ. 2000, 82, 105–119. [Google Scholar] [CrossRef]
- Hong, W.J.; Luo, J.; Chen, Y.L.; Mai, Z.T.; Chen, W.Y.; Cai, K.L. Effects of mycorrhizal fungi on the drought tolerance of Aquilaria sinensis seedlings. J. Anhui Agric. Univ. 2018, 45, 455–461. [Google Scholar]
- Zhang, Y.M.; Ma, K.M.; Qu, L.Y. Inoculation with arbuscular mycorrhizal fungi enhances the root system of Bauhinia faberi var. microphylla seedlings under drought stress conditions. Acta Ecol. Sin. 2017, 37, 2611–2619. [Google Scholar]
- Qiu, L.; Bi, Y.L.; Zhang, Y.X.; Cai, Y.; Yu, H.Y. Effect of film mulching and microbial inoculation on maize growth and water use efficiency under drought stress. Chin. J. Appl. Ecol. 2018, 29, 2915–2924. [Google Scholar]
- Bi, Y.L.; Zhang, Y.X.; Jiang, B.; Qiu, L. Effects of AM fungi and phosphate-solubilizing bacteria inoculation on maize growth and soil fertility under water stress. J. China Coal Soc. 2019, 44, 3655–3661. [Google Scholar]
- Etesami, H.; Maheshwari, D.K. Use of plant growth promoting rhizobacteria (PGPRs) with multiple plant growth promoting traits in stress agriculture: Action mechanisms and future prospects. Ecotoxicol. Environ. Saf. 2018, 156, 225–246. [Google Scholar] [CrossRef]
- Liu, F.C.; Xing, S.J.; Ma, H.L.; Du, Z.Y.; Ma, B.Y. Cytokinin-producing, plant growth-promoting rhizobacteria that confer resistance to drought stress in Platycladus orientalis container seedlings. Appl. Microbiol. Biotechnol. 2013, 97, 9155–9164. [Google Scholar] [CrossRef]
- Niu, X.G.; Song, L.C.; Yinong Xiao, Y.N.; Weide Ge, W.D. Drought-tolerant plant growth-promoting rhizobacteria associated with foxtail millet in a semi-arid agroecosystem and their potential in alleviating drought stress. Front. Microbiol. 2018, 8, 2580. [Google Scholar] [CrossRef] [Green Version]
- Prudent, M.; Salon, C.; Souleimanov, A.; Emery, R.J.N.; Smith, D.L. Soybean is less impacted by water stress using Bradyrhizobium japonicum and thuricin-17 from Bacillus thuringiensis. Agron. Sustain. Dev. 2015, 35, 749–757. [Google Scholar]
- Calvo-Polanco, M.; Sánchez-Romera, B.; Aroca, R.; Asins, M.J.; Declerck, S.; Dodd, I.C.; Martínez-Andújar, C.; Albacete, A.; Ruiz-Lozano, J.M. Exploring the use of recombinant inbred lines in combination with beneficial microbial inoculants (AM fungus and PGPR) to improve drought stress tolerance in tomato. Environ. Exp. Bot. 2016, 131, 47–57. [Google Scholar] [CrossRef] [Green Version]
- Kanwal, S.; Ilyas, N.; Batool, N.; Arshad, M.; Ahmad, B. Amelioration of drought stress in wheat by combined application of PGPR, compost, and mineral fertilizer. J. Plant Nutr. 2016, 40, 1250–1260. [Google Scholar] [CrossRef]
- Ngumbi, E.; Kloepper, J. Bacterial-mediated drought tolerance: Current and future prospects. Appl. Soil Ecol. 2016, 105, 109–125. [Google Scholar] [CrossRef]
- Gontia-Mishra, I.; Sapre, S.; Kachare, S.; Tiwari, S. Molecular diversity of 1-aminocyclopropane-1-carboxylate (ACC) deaminase producing PGPR from wheat (Triticum aestivum L.) rhizosphere. Plant Soil 2017, 414, 213–227. [Google Scholar] [CrossRef]
- Timmusk, S.; Wagner, E.G.H.; Gruau, C.; Trotel-Aziz, P.; Villaume, S.; Rabenoelina, F.; Clément, C.; Baillieul, F.; Aziz, A.; Stearns, J.C.; et al. The Plant-Growth-Promoting Rhizobacterium Paenibacillus polymyxa Induces Changes in Arabidopsis thaliana Gene Expression: A Possible Connection Between Biotic and Abiotic Stress Responses. Mol. Plant-Microbe Interact. 1999, 12, 951–959. [Google Scholar] [CrossRef] [PubMed]
- Kasim, W.; Osman, M.; Omar, M.; Abd El-Daim, I.; Bejai, S.; Meijer, J. Control of drought stress in wheat using plant-growth promoting rhizobacteria. J. Plant Growth Regul. 2013, 32, 122–130. [Google Scholar] [CrossRef]
- Yang, J.; Kloepper, J.W.; Ryu, C. Rhizosphere bacteria help plants tolerate abiotic stress. Trends Plant Sci. 2009, 14, 1–4. [Google Scholar] [CrossRef]
- Shah, U.N.; Mir, J.I.; Ahmed, N.; Jan, S.; Fazili, K.M. Bioefficacy potential of different genotypes of walnut Juglans regia L. J. Food Sci. Technol. 2018, 55, 605–618. [Google Scholar] [CrossRef]
- Li, J.W.; Yan, S.L.; Huang, Y.C.; Xia, X.X.; Chu, L.F.; Li, C.Y.; Wang, Z.J. Physiological and biochemical responses of pecan seedlings to drought stress. J. Nucl. Agric. Sci. 2020, 34, 2326–2334. [Google Scholar]
- Li, D.P.; Li, S.P.; Gao, X.Q.; Zhao, T.T.; Zhang, F.F.; Yang, G.Y. Physiological evaluation of response of ′Ziren′ walnut from Ankang under drought stress. J. Southwest For. Univ. 2018, 38, 202–206. [Google Scholar]
- Han, L.; Xu, M.; Zhang, P. Changes of osmoregulation in Xinjiang wild walnut seedlings under drought stress. Tianjin Agric. Sci. 2018, 24, 1–3. [Google Scholar]
- Hussain, A.; Hasnain, S. Phytostimulation and biofertilization in wheat by cyanobacteria. J. Ind. Microbiol. Biotechnol. 2010, 38, 85–92. [Google Scholar] [CrossRef]
- Holt, J.G.; Krieg, N.R.; Sneath, P.H.A.; Staley, J.T.; Williams, S.T. Bergey’s Manual of Determinative Bacteriology, 9th ed.; William & Wilkins: Baltimore, MD, USA, 1994. [Google Scholar]
- Bollmark, M.; Kubát, B.; Eliasson, L. Variation in Endogenous Cytokinin Content during Adventitious Root Formation in Pea Cuttings. J. Plant Physiol. 1988, 132, 262–265. [Google Scholar] [CrossRef]
- He, Z. Guidance to Experiment on Chemical Control in Crop Plants. In Guidance to Experiment on Chemical Control in Crop Plants; He, Z.P., Ed.; Beijing Agricultural University Publishers: Beijing, China, 1993; pp. 60–68. [Google Scholar]
- Barrs, H.D.; Weatherley, P.E. A re-examination of the relative turgidity technique for estimating water deficits in leaves. Aust. J. Biol. Sci. 1962, 15, 413–428. [Google Scholar] [CrossRef] [Green Version]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Naidu, B.P. Separation of sugars, polyols, proline analogues, and betaines in stressed plant extracts by high-performance liquid chromatography and quantification by ultra violet detection. Aust. J. Plant Physiol. 1998, 25, 793–800. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric methods for determination of sugars of related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Beyer, W.F.; Fridovich, I. Assaying for superoxide dismutase activity: Some large consequences of minor changes in conditions. Anal. Biochem. 1987, 161, 559–566. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar]
- Dong, B.D.; Zheng, X.; Liu, H.P.; Able, J.A.; Yang, H.; Zhao, H.; Zhang, M.M.; Qiao, Y.Z.; Wang, Y.K.; Liu, M.Y. Effects of drought stress on pollen sterility, grain yield, abscisic acid and protective enzymes in two winter wheat cultivars. Front. Plant Sci. 2017, 8, 1008. [Google Scholar] [CrossRef]
- Sandhya, V.; Ali, S.Z.; Grover, M.; Reddy, G.; Venkateswarlu, B. Effect of plant growth promoting Pseudomonas spp. on compatible solutes, antioxidant status and plant growth of maize under drought stress. Plant Growth Regul. 2010, 62, 21–30. [Google Scholar] [CrossRef]
- Vurukonda, S.S.K.P.; Vardharajula, S.; Shrivastava, M.; SkZ, A. Enhancement of drought stress tolerance in crops by plant growth promoting rhizobacteria. Microbiol. Res. 2016, 184, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.N.; Zhou, H.H.; Chen, Y.P. Adaptation strategies of desert riparian forest vegetation in response to drought stress. Ecohydrology 2013, 6, 956–973. [Google Scholar] [CrossRef]
- Guo, Y.Y.; Yu, H.Y.; Yang, M.M.; Kong, D.S.; Zhang, Y.J. Effect of drought stress on lipid peroxidation, osmotic adjustment and antioxidant enzyme activity of leaves and roots of Lycium ruthenicum Murr. seedling. Russ. J. Plant Physiol. 2018, 65, 244–250. [Google Scholar] [CrossRef]
- Martínez, J.P.; Silva, H.; Ledent, J.F.; Pinto, M. Effect of drought stress on the osmotic adjustment, cell wall elasticity and cell volume of six cultivars of common beans (Phaseolus vulgaris L.). Eur. J. Agron. 2007, 26, 30–38. [Google Scholar] [CrossRef]
- Meise, P.; Seddig, S.; Uptmoor, R.; Ordon, F.; Schum, A. Impact of nitrogen supply on leaf water relations and physiological traits in a set of potato (Solanum tuberosum L.) cultivars under drought stress. J. Agron. Crop Sci. 2018, 204, 359–374. [Google Scholar] [CrossRef]
- Feng, H.F.; Xue, L.; Ren, X.R.; Fu, J.D.; Zheng, W.G.; Shi, X.L. Physiological responses of four broad leaved seedlings to drought stress simulated by PEG. Acta Ecol. Sin. 2011, 31, 371–382. [Google Scholar]
- Ma, Z.R.; Liu, R.T. Physiological Principles of Drought Resistance in Herbage; Lanzhou University Press: Lanzhou, China, 1993. [Google Scholar]
- Izanloo, A.; Condon, A.G.; Langridge, P.; Tester, M.; Schnurbusch, T. Different mechanisms of adaptation to cyclic water stress in two South Australian bread wheat cultivars. J. Exp. Bot. 2008, 59, 3327–3346. [Google Scholar] [CrossRef] [Green Version]
- Kohler, J.; Hernández, J.A.; Caravaca, F.; Roldán, A. Plant-growth-promoting rhizobacteria and arbuscular mycorrhizal fungi modify alleviation biochemical mechanisms in water-stressed plants. Funct. Plant Biol. 2008, 35, 141–151. [Google Scholar] [CrossRef]
- Esmaeilpour, A.; Van Labeke, M.C.; Samson, R.; Boeckx, P.; Van Damme, P. Variation in biochemical characteristics, water status, stomata features, leaf carbon isotope composition and its relationship to water use efficiency in pistachio (Pistacia vera L.) cultivars under drought stress condition. Sci. Hortic. 2016, 211, 158–166. [Google Scholar] [CrossRef]
- Li, L.J.; Gu, W.R.; Li, J.; Li, C.F.; Xie, T.L.; Qu, D.Y.; Meng, Y.; Li, C.F.; Wei, S. Exogenously applied spermidine alleviates photosynthetic inhibition under drought stress in maize (Zea mays L.) seedlings associated with changes in endogenous polyamines and phytohormones. Plant Physiol. Biochem. 2018, 129, 35–55. [Google Scholar] [CrossRef]
- Kurepin, L.V.; Ivanov, A.G.; Zaman, M.; Pharis, R.P.; Allakhverdiev, S.I.; Hurry, V.; Hüner, N.P. Stress-related hormones and glycinebetaine interplay in protection of photosynthesis under abiotic stress conditions. Photosynth. Res. 2015, 126, 221–235. [Google Scholar] [CrossRef]
- Arkhipova, T.N.; Prinsen, E.; Veselov, S.U.; Martinenko, E.V.; Melentiev, A.I.; Kudoyarova, G.R. Cytokinin producing bacteria enhance plant growth in drying soil. Plant Soil 2007, 292, 305–315. [Google Scholar] [CrossRef]
- Zhang, S.H.; Xu, X.F.; Sun, Y.M.; Zhang, J.L.; Li, C.Z. Influence of drought hardening on the resistance physiology of potato seedlings under drought stress. J. Integr. Agric. 2018, 17, 336–347. [Google Scholar] [CrossRef] [Green Version]
- Liang, D.; Ni, Z.Y.; Xia, H.; Xie, Y.; Lv, X.L.; Wang, J.; Lin, L.J.; Deng, Q.X.; Luo, X. Exogenous melatonin promotes biomass accumulation and photosynthesis of kiwifruit seedlings under drought stress. Sci. Hortic. 2019, 246, 34–43. [Google Scholar] [CrossRef]
- Farooq, M.; Hussain, M.; Siddique, K.H.M. Drought stress in wheat during flowering and grain-filling periods. Crit. Rev. Plant Sci. 2014, 33, 331–349. [Google Scholar] [CrossRef]
- Thapa, S.; Reddy, S.K.; Fuentealba, M.P.; Xue, Q.; Rudd, J.C.; Jessup, K.E.; Devkota, R.N.; Liu, S. Physiological responses to water stress and yield of winter wheat cultivars differing in drought tolerance. J. Agron. Crop Sci. 2018, 204, 347–358. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Gluten Hydrolysis | Starch Hydrolysis | Indole Test | V-P Test | Nitrate Reduction | Mannitol Test | Peroxidase Test | Lecithinase Test | Gas Production of Glucose | Acid Production of Glucose | Citrate Utilization |
|---|---|---|---|---|---|---|---|---|---|---|
| + | + | + | + | + | − | + | + | − | + | + |
| Drought Treatment | Inoculation Treatment | Antioxidant Enzyme Activity (U·g−1 FW) | Osmotic Adjustment Substance Content (mg·g−1 FW) | Relative Water Content (%) | ||
|---|---|---|---|---|---|---|
| SOD | CAT | Proline | Soluble Sugars | |||
| CK | PGPR | 347.63 ± 20.09 f | 73.16 ± 2.57 d | 16.49 ± 5.33 d | 6.37 ± 0.68 d | 87.11 ± 0.96 a |
| NS | 353.02 ± 12.96 f | 69.87 ± 5.93 d | 18.25 ± 2.78 d | 6.21 ± 1.80 d | 87.64 ± 1.05 a | |
| LD | PGPR | 449.95 ± 25.87 d | 82.90 ± 3.71 c | 18.68 ± 3.09 d | 6.55 ± 1.22 d | 84.03 ± 1.37 b |
| NS | 392.26 ± 17.23 e | 73.39 ± 4.06 d | 20.13 ± 2.28 d | 6.86 ± 0.98 d | 83.56 ± 0.65 b | |
| MD | PGPR | 687.54 ± 21.63 b | 95.76 ± 5.32 b | 26.38 ± 1.95 c | 9.32 ± 0.79 c | 78.45 ± 1.53 c |
| NS | 571.60 ± 38.05 c | 82.03 ± 2.81 c | 27.59 ± 2.62 c | 12.96 ± 0.51 b | 75.31 ± 1.16 d | |
| SD | PGPR | 865.31 ± 41.54 a | 116.90 ± 8.65 a | 36.62 ± 5.10 b | 13.55 ± 0.92 b | 71.93 ± 0.72 e |
| NS | 702.98 ± 49.16 b | 91.85 ± 3.16 b | 47.50 ± 3.25 a | 16.71 ± 1.54 a | 69.89 ± 0.97 f | |
| Drought Treatment | Inoculation Treatment | IAA | GA | ZT | ABA |
|---|---|---|---|---|---|
| CK | PGPR | 30.65 ± 0.81 a | 180.34 ± 5.26 a | 3.16 ± 0.05 a | 0.72 ± 0.02 c |
| NS | 26.71 ± 1.03 b | 168.05 ± 3.53 b | 2.87 ± 0.05 b | 0.54 ± 0.03 e | |
| LD | PGPR | 25.98 ± 1.45 b | 177.52 ± 3.91 a | 3.09 ± 0.07 a | 0.73 ± 0.02 c |
| NS | 23.87 ± 0.92 bc | 156.81 ± 6.05 c | 2.65 ± 0.09 c | 0.57 ± 0.05 e | |
| MD | PGPR | 22.36 ± 0.79 c | 141.96 ± 5.70 d | 2.68 ± 0.10 c | 0.80 ± 0.03 b |
| NS | 19.22 ± 1.63 d | 126.57 ± 8.13 e | 2.39 ± 0.07 d | 0.83 ± 0.06 b | |
| SD | PGPR | 18.90 ± 0.57 d | 128.62 ± 6.94 e | 2.41 ± 0.16 d | 0.66 ± 0.02 d |
| NS | 14.19 ± 1.28 e | 103.79 ± 8.68 f | 1.93 ± 0.22 e | 0.95 ± 0.03 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, F.; Ma, H.; Liu, B.; Du, Z.; Ma, B.; Jing, D. Effects of Plant Growth-Promoting Rhizobacteria on the Physioecological Characteristics and Growth of Walnut Seedlings under Drought Stress. Agronomy 2023, 13, 290. https://doi.org/10.3390/agronomy13020290
Liu F, Ma H, Liu B, Du Z, Ma B, Jing D. Effects of Plant Growth-Promoting Rhizobacteria on the Physioecological Characteristics and Growth of Walnut Seedlings under Drought Stress. Agronomy. 2023; 13(2):290. https://doi.org/10.3390/agronomy13020290
Chicago/Turabian StyleLiu, Fangchun, Hailin Ma, Binghua Liu, Zhenyu Du, Bingyao Ma, and Dawei Jing. 2023. "Effects of Plant Growth-Promoting Rhizobacteria on the Physioecological Characteristics and Growth of Walnut Seedlings under Drought Stress" Agronomy 13, no. 2: 290. https://doi.org/10.3390/agronomy13020290