
Plant-Growth-Promoting Rhizobacteria Improve Germination and Bioactive Compounds in Cucumber Seedlings
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Vegetal Material
2.2. Bacterial Strains
2.3. Treatments
2.4. Germination and Growth Conditions
2.5. Parameters Evaluated during the Bioassay
2.6. Indole Acetic Acid Production
2.7. Preparation of Extracts for Phytochemical Compounds
2.8. Phytochemical Compounds
2.9. Statistical Analysis
3. Results
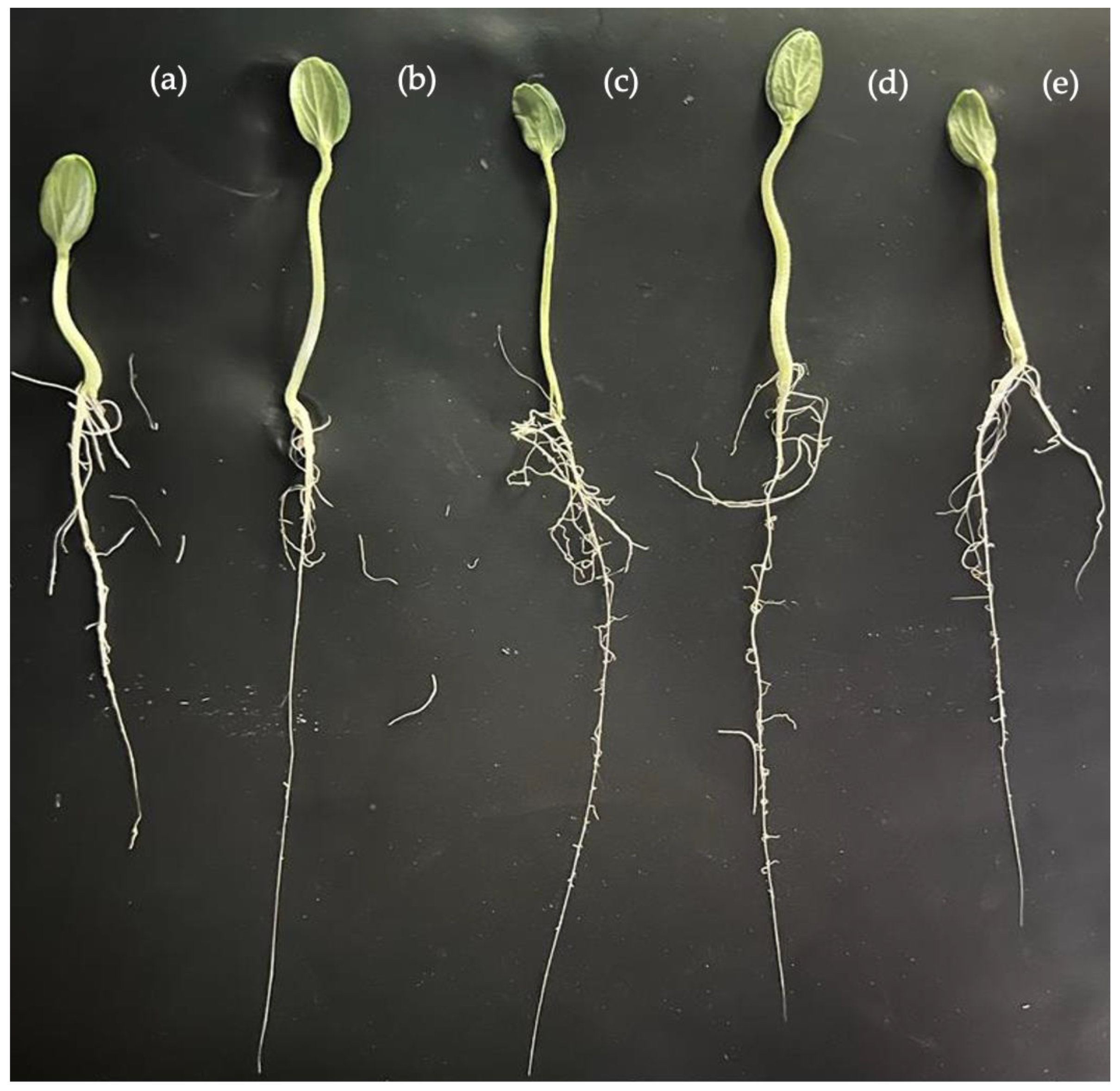
3.1. Seed Germination
3.2. Production of Indole-3-Acetic Acid
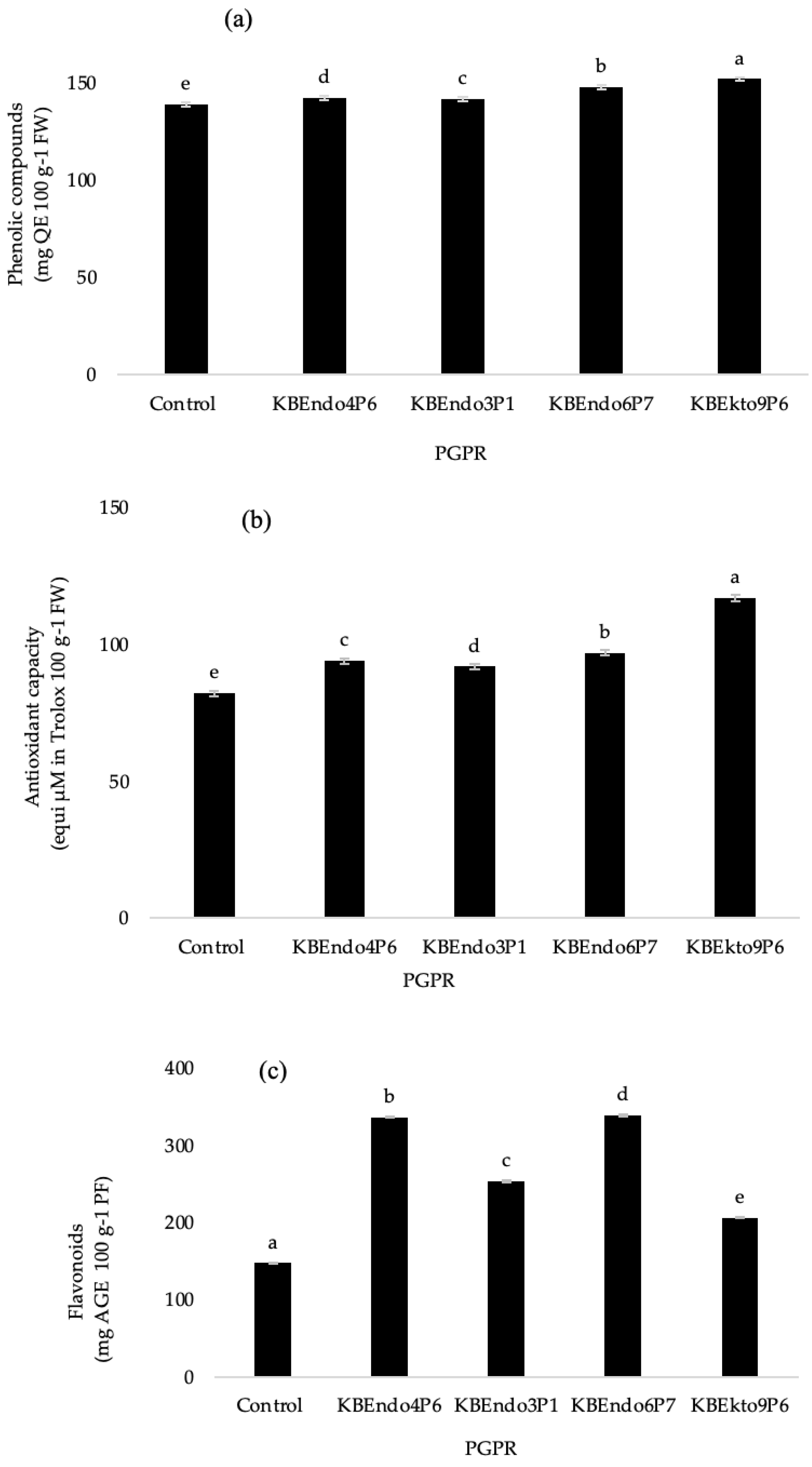
3.3. Phytochemical Compounds
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Behera, T.K.; Boopalakrishnan, G.; Jat, G.S.; Das Munshi, A.; Choudhary, H.; Ravindran, A.; Kumari, S.; Kumari, R. Deriving stable tropical gynoecious inbred lines of slicing cucumber from American pickling cucumber using MABB. Hortic. Environ. Biotechnol. 2022, 63, 263–274. [Google Scholar] [CrossRef]
- Dumitru, E.A.; Șurcă, E.D. The Importance of Cucumber Culture in the Vegetable Sector in 2012–2017. Sci. Pap. 2019, 19, 169. [Google Scholar]
- Álvarez-García, J.-A.; Santoyo, G.; Rocha-Granados, M.d.C. Pseudomonas fluorescens: Mechanisms and applications in sustainable agriculture. Lat. Am. J. Nat. Resour. 2020, 16, 1–10. Available online: https://revista.itson.edu.mx/index.php/rlrn/article/view/286 (accessed on 22 November 2022).
- Zörb, C.; Geilfus, C.M.; Dietz, K.J. Salinity and crop yield. Plant Biol. 2019, 21, 31–38. [Google Scholar] [CrossRef]
- Lu, L.; Huang, M.; Huang, Y.; Corvini, P.F.X.; Ji, R.; Zhao, L. Mn3O4 nanozymes boost endogenous antioxidant metabolites in cucumber (Cucumis sativus) plant and enhance resistance to salinity stress. Environ. Sci. Nano 2020, 7, 1692–1703. [Google Scholar] [CrossRef]
- Kazaz, S.; Erba, S.; Baydar, H. Breaking seed dormancy in oil rose (Rosa damascena Mill.) by microbial inoculation. Afr. J. Biotechnol. 2010, 9, 6503–6508. [Google Scholar]
- Perez-Garcia, L.A.; Castañeda-Ramírez, J.C.; Lafuente-Rincón, D. Successful and unexpected potential of entomopathogen Bacillus thuringiensis as plant growth promoting rhizobacterium Exitoso e inesperado potencial del entomopatógeno Bacillus thuringiensis como rizobacteria promotora del crecimiento de las plantas. J. Nat. Agric. Sci. 2019, 6, 40–46. [Google Scholar] [CrossRef]
- Swarnalakshmi, K.; Yadav, V.; Tyagi, D.; Dhar, D.W.; Kannepalli, A.; Kumar, S. Significance of plant growth promoting rhizobacteria in grain legumes: Growth promotion and crop production. Plants 2020, 9, 1596. [Google Scholar] [CrossRef]
- Nazir, U.; Zargar, M.; Baba, Z.; Mir, S.; Mohiddin, F.; Bhat, N. Isolation and characterization of plant growth promoting rhizobacteria associated with pea rhizosphere in North Himalayan region. Int. J. Chem. Stud. 2020, 8, 1131–1135. [Google Scholar] [CrossRef]
- Amador-Alférez, K.A.; Díaz-González, J.; Loza-Cornejo, S.; Bivián-Castro, E.Y. Effect of different plant growth regulators on seed germination and seedling development of two species of Ferocactus (Cactaceae). Polybotany 2013, 35, 109–131. Available online: http://www.scielo.org.mx/scielo.php?script=sci_arttext&pid=S1405-27682013000100007&lng=es&tlng= (accessed on 3 December 2022).
- Saberi-Riseh, R.; Fathi, F.; Moradzadeh-Eskandari, M. Effect of some Pseudomonas fluorescens and Bacillus subtilis strains on osmolytes and antioxidants of cucumber under salinity stress. J. Crop Prot. 2020, 9, 1–16. [Google Scholar]
- Basu, A.; Prasad, P.; Das, S.N.; Kalam, S.; Sayyed, R.Z.; Reddy, M.S.; El Enshasy, H. Plant Growth Promoting Rhizobacteria (PGPR) as Green Bioinoculants: Recent Developments, Constraints, and Prospects. Sustainability 2021, 13, 1140. [Google Scholar] [CrossRef]
- Mohanty, P.; Singh, P.K.; Chakraborty, D.; Mishra, S.; Pattnaik, R. Insight into the role of PGPR in sustainable agriculture and environment. Front. Sustain. Food Syst. 2021, 5, 667150. [Google Scholar] [CrossRef]
- Mondal, S.; Pramanik, K.; Ghosh, S.K.; Pal, P.; Mondal, T.; Soren, T.; Maiti, T.K. Unraveling the role of plant growth-promoting rhizobacteria in the alleviation of arsenic phytotoxicity: A review. Microbiol. Res. 2021, 250, 126809. [Google Scholar] [CrossRef] [PubMed]
- Meena, M.; Swapnil, P.; Divyanshu, K.; Kumar, S.; Tripathi, Y.N.; Zehra, A.; Marwal, A.; Upadhyay, R.S. PGPR-mediated induction of systemic resistance and physiochemical alterations in plants against the pathogens: Current perspectives. J. Basic Microbiol. 2020, 60, 828–861. [Google Scholar] [CrossRef] [PubMed]
- Suryadi, Y.; Susilowati, D.N.; Fauziah, F. Management of Plant Diseases by PGPR-Mediated Induced Resistance with Special Reference to Tea and Rice Crops. In Plant Growth Promoting Rhizobacteria for Sustainable Stress Management; Sayyed, R., Ed.; Microorganisms for Sustainability; Springer: Singapore, 2019; Volume 13. [Google Scholar] [CrossRef]
- Jabborova, D.; Wirth, S.; Kannepalli, A.; Narimanov, A.; Desouky, S.; Davranov, K.; Sayyed, R.; El Enshasy, H.; Malek, R.; Syed, A. Co-inoculation of rhizobacteria and biochar application improves growth and nutrientsin soybean and enriches soil nutrients and enzymes. Agronomy 2020, 10, 1142. [Google Scholar] [CrossRef]
- Rahma, H.; Kristina, N. Plant Growth Promoting Rhizobacteria (PGPR): As a Potential Biocontrol for Curvularia lunata in vitro. J. Phys. Conf. Ser. 2021, 1940, 012097. [Google Scholar] [CrossRef]
- Hamedalla, A.M.; Ali, M.M.; Ali, W.M.; Ahmed, M.A.; Kaseb, M.O.; Kalaji, H.M.; Gajc-Wolska, J.; Yousef, A.F. Increasing the performance of cucumber (Cucumis sativus L.) seedlings by LED illumination. Sci. Rep. 2022, 12, 852. [Google Scholar] [CrossRef]
- Bric, J.M.; Bostock, R.M.; Silverstone, S.E. Rapid In Situ assay for indoleacetic acid production by bacteria immobilized on a nitrocellulose membrane. Appl. Environ. Microbiol. 1991, 57, 535–538. [Google Scholar] [CrossRef] [Green Version]
- Glickmann, E.; Dessaux, Y. A critical examination of the specificity of the Salkowski reagent for indolic compounds produced by phytopathogenic bacteria. Appl. Environ. Microbiol. 1995, 61, 793–796. [Google Scholar] [CrossRef] [Green Version]
- Ortega-Ortiz, H.; Gaucin-Delgado, J.M.; Preciado-Rangel, P.; Hernandez, M.F.; Hernandez-Montiel, L.G.; DE La Cruz-Lazaro, E.; Lara-Capistrán, L. Copper oxide nanoparticles biosynthetized improve germination and bioactive compounds in wheat sprouts. Not. Bot. Horti Agrobot. Cluj-Napoca 2022, 50, 12657. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of folin-ciocalteu reagent. Methods Enzymol. 1999, 299, 152–178. [Google Scholar] [CrossRef]
- Zhishen, J.; Mengcheng, T.; Jianming, W. The determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Li, T.; Wang, L.; Sun, D.; Li, Y.; Chen, Z. Effect of enzymolysis-assisted electron beam irradiation on structural characteristics and antioxidant activity of rice protein. J. Cereal Sci. 2019, 89, 102789. [Google Scholar] [CrossRef]
- Panigrahi, S.; Mohanty, S.; Rath, C.C. Characterization of endophytic bacterium Enterobacter cloacae MG00145 isolated from Ocimum sanctum with Indole Acetic Acid (IAA) production and plant growth promoting capabilities against selected crops. South Afr. J. Bot. 2020, 134, 17–26. [Google Scholar] [CrossRef]
- Fouda, A.; Eid, A.M.; Elsaied, A.; El-Belely, E.F.; Barghoth, M.G.; Azab, E.; Gobouri, A.A.; Hassan, S.E.D. Plant growth-promoting endophytic bacterial community inhabiting the leaves of Pulicaria incisa (Lam.) DC inherent to arid regions. Plants 2021, 10, 76. [Google Scholar] [CrossRef] [PubMed]
- Hagaggi, N.S.; Mohamed, A.A. Enhancement of Zea mays (L.) growth performance using indole acetic acid producing endophyte Mixta theicola isolated from Solenostemma argel (Hayne). S. Afr. J. Bot. 2020, 134, 64–71. [Google Scholar] [CrossRef]
- Nawaz, A.; Shahbaz, M.; Imran, A.; Marghoob, M.U.; Imtiaz, M.; Mubeen, F. Potential of salt tolerant PGPR in growth and yield augmentation of wheat (Triticum aestivum L.) under saline conditions. Front. Microbiol. 2020, 11, 2019. [Google Scholar] [CrossRef]
- Ureche, M.A.L.; Pérez-Rodriguez, M.M.; Ortiz, R.; Monasterio, R.P.; Cohen, A.C. Rhizobacteria improve the germination and modify the phenolic compound profile of pepper (Capsicum annum L.). Rhizosphere 2021, 18, 100334. [Google Scholar] [CrossRef]
- Monalisa, S.P.; Roy, S. Chapter-5 Effect of Biopriming with Plant Growth Promoting Rhizobacteria (PGPR). In Research Trends in Agriculture Sciences; Naresh, R.K., Ed.; AkiNik Publications: New Delhi, India, 2022; p. 73. Available online: https://www.researchgate.net/profile/Rk-Naresh-2/publication/362684642_Research_Trends_in_Agriculture_Vol_30/links/62f86e96b8dc8b4403df7b23/Research-Trends-in-Agriculture-Vol-30.pdf#page=77 (accessed on 30 December 2022).
- Miljaković, D.; Marinković, J.; Tamindžić, G.; Đorđević, V.; Tintor, B.; Milošević, D.; Ignjatov, M.; Nikolić, Z. Bio-priming of soybean with Bradyrhizobium japonicum and Bacillus megaterium: Strategy to improve seed germination and the initial seedling growth. Plants 2022, 11, 1927. [Google Scholar] [CrossRef]
- Khatoon, Z.; Huang, S.; Rafique, M.; Fakhar, A.; Kamran, M.A.; Santoyo, G. Unlocking the potential of plant growth-promoting rhizobacteria on soil health and the sustainability of agricultural systems. J. Environ. Manag. 2020, 273, 111118. [Google Scholar] [CrossRef] [PubMed]
- Vega-Celedón, P.; Canchignia Martínez, H.; González, M.; Seeger, M. Biosynthesis of indole-3-acetic acid and promotion of plant growth by bacteria. Trop. Crops 2016, 37, 33–39. Available online: http://scielo.sld.cu/scielo.php?script=sci_arttext&pid=S0258-59362016000500005&lng=es&tlng=en (accessed on 9 December 2022).
- Yuzikhin, O.S.; Gogoleva, N.E.; Shaposhnikov, A.I.; Konnova, T.A.; Osipova, E.V.; Syrova, D.S.; Ermakova, E.; Shevchenko, V.; Nagaev, I.; Shevchenko, K.; et al. Rhizosphere bacterium rhodococcus sp. P1Y metabolizes abscisic acid to form dehydrovomifoliol. Biomolecules 2021, 11, 345. [Google Scholar] [CrossRef]
- Chiappero, J.; Cappellari, L.D.R.; Alderete, L.G.S.; Palermo, T.B.; Banchio, E. Plant growth promoting rhizobacteria improve the antioxidant status in Mentha piperita grown under drought stress leading to an enhancement of plant growth and total phenolic content. Ind. Crops Prod. 2019, 139, 111553. [Google Scholar] [CrossRef]
- Khanna, K.; Jamwal, V.L.; Sharma, A.; Gandhi, S.G.; Ohri, P.; Bhardwaj, R.; Al-Huqail, A.A.; Siddiqui, M.H.; Ali, H.M.; Ahmad, P. Supplementation with plant growth promoting rhizobacteria (PGPR) alleviates cadmium toxicity in Solanum lycopersicum by modulating the expression of secondary metabolites. Chemosphere 2019, 230, 628–639. [Google Scholar] [CrossRef]
- Batool, T.; Ali, S.; Seleiman, M.F.; Naveed, N.H.; Ali, A.; Ahmed, K.; Abid, M.; Rizwan, M.; Shahid, M.R.; Alotaibi, M.; et al. Plant growth promoting rhizobacteria alleviates drought stress in potato in response to suppressive oxidative stress and antioxidant enzymes activities. Sci. Rep. 2020, 10, 16975. [Google Scholar] [CrossRef]
- Asghari, B.; Khademian, R.; Sedaghati, B. Plant growth promoting rhizobacteria (PGPR) confer drought resistance and stimulate biosynthesis of secondary metabolites in pennyroyal (Mentha pulegium L.) under water shortage condition. Sci. Hortic. 2020, 263, 109132. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| PGPR | G | GI | V | PL | RL | FW | DW |
|---|---|---|---|---|---|---|---|
| % | cm | mg | |||||
| Control | 76.6 e ± 0.51 * | 39.6 e ± 0.22 | 23.3 d ± 11.54 | 2.2 c ± 0.44 | 6.2 d ± 0.40 | 231.9 a ± 0.08 | 15.0 a ± 0.008 |
| KBEndo4P6 | 93.3 b ± 0.51 | 78.7 b ± 0.46 | 80.0 b ± 0.001 | 3.5 a ± 0.54 | 9.5 b ± 0.77 | 328.7 a ± 0.04 | 18.1 a ± 0.001 |
| KBEndo3P1 | 96.6 a ± 0.40 | 89.7 a ± 0.21 | 86.6 a ± 5.74 | 3.4 a ± 1.10 | 10.9 a ± 0.37 | 298.7 a ± 0.07 | 17.4 a ± 0.001 |
| KBEndo6P7 | 86.6 d ± 0.54 | 69.5 d ± 0.16 | 60.6 c ± 5.74 | 2.8 b ± 0.43 | 9.5 b ± 0.67 | 269.2 a ± 0.05 | 19.6 a ± 0.003 |
| KBekto9p6 | 90.0 c ± 0.54 | 72.3 c ± 0.26 | 86.6 to ± 5.74 | 2.8 b ± 0.001 | 8.7 c ± 0.78 | 311.4 a ± 0.05 | 18.1 a ± 0.006 |
| PGPR | IAA mg mL−1 |
|---|---|
| KBEndo4P6 | 0.076 ± 0.020 |
| KBEndo3P1 | 0.392 ± 0.048 |
| KBEndo6P7 | 0.086 ± 0.014 |
| KBEkto9P6 | 0.085 ± 0.011 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pérez-García, L.-A.; Sáenz-Mata, J.; Fortis-Hernández, M.; Navarro-Muñoz, C.E.; Palacio-Rodríguez, R.; Preciado-Rangel, P. Plant-Growth-Promoting Rhizobacteria Improve Germination and Bioactive Compounds in Cucumber Seedlings. Agronomy 2023, 13, 315. https://doi.org/10.3390/agronomy13020315
Pérez-García L-A, Sáenz-Mata J, Fortis-Hernández M, Navarro-Muñoz CE, Palacio-Rodríguez R, Preciado-Rangel P. Plant-Growth-Promoting Rhizobacteria Improve Germination and Bioactive Compounds in Cucumber Seedlings. Agronomy. 2023; 13(2):315. https://doi.org/10.3390/agronomy13020315
Chicago/Turabian StylePérez-García, Laura-Andrea, Jorge Sáenz-Mata, Manuel Fortis-Hernández, Claudia Estefanía Navarro-Muñoz, Rubén Palacio-Rodríguez, and Pablo Preciado-Rangel. 2023. "Plant-Growth-Promoting Rhizobacteria Improve Germination and Bioactive Compounds in Cucumber Seedlings" Agronomy 13, no. 2: 315. https://doi.org/10.3390/agronomy13020315