
Effects of Long-Term Exclosure on Main Plant Functional Groups and Their Biochemical Properties in a Patchily Degraded Alpine Meadow in the Source Zone of the Yellow River, West China
Abstract
:1. Introduction
2. Materials and Methods
2.1. Overview of the Study Area
2.2. Experimental Design
2.3. Sample Collection and Analysis
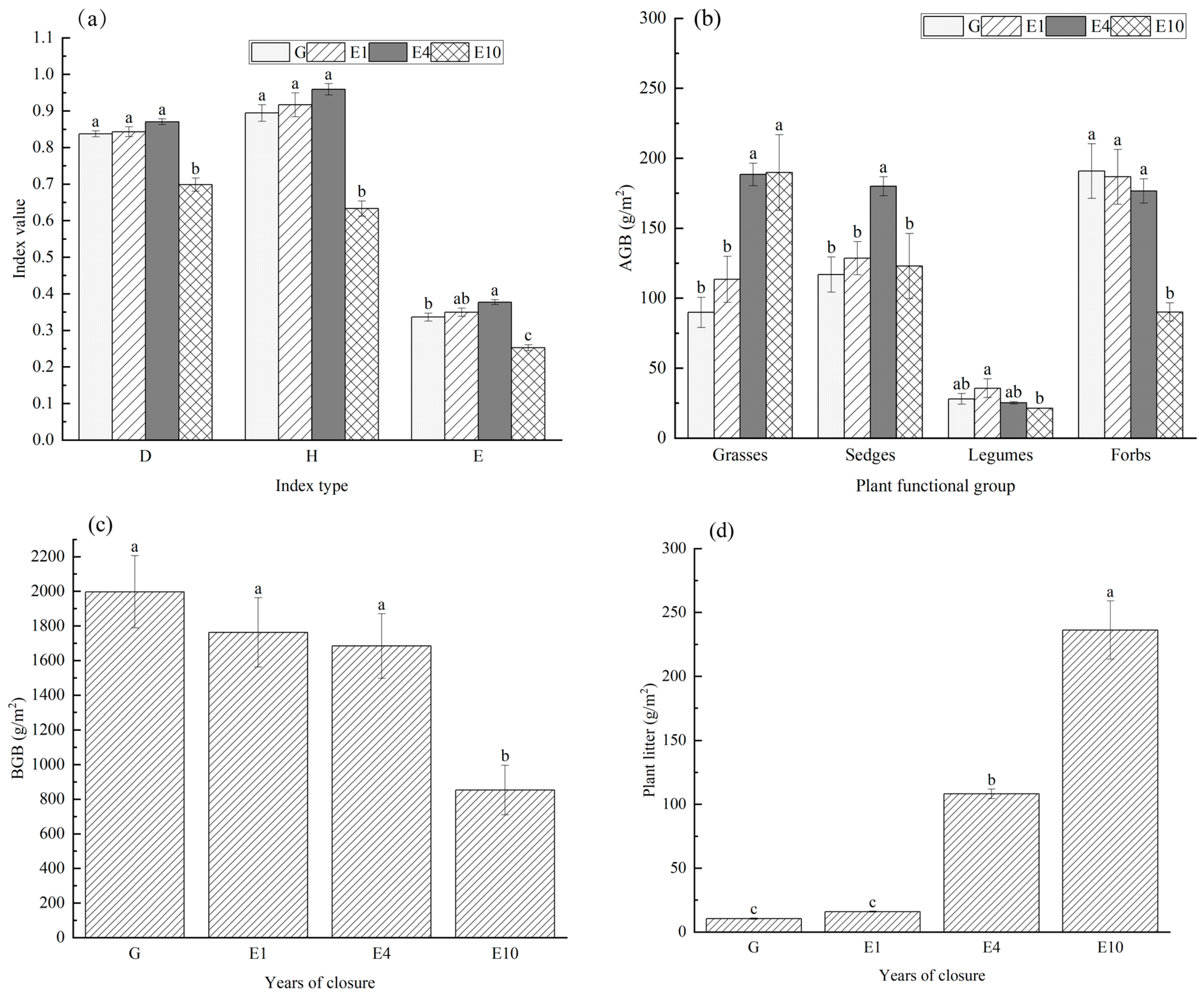
2.3.1. Determination of Plant Community Dynamics, Population Characteristics, and Aboveground and Belowground Biomass
2.3.2. Plant Community Structure Indices
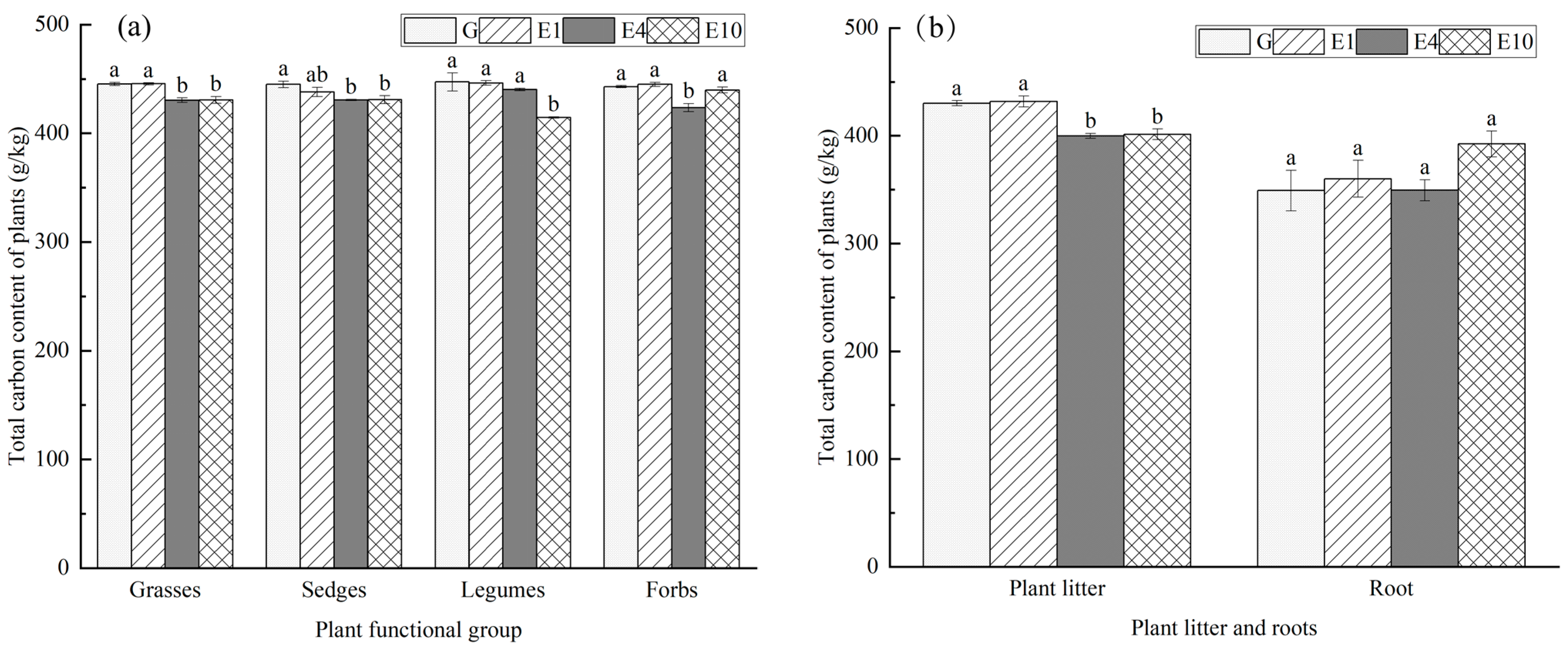
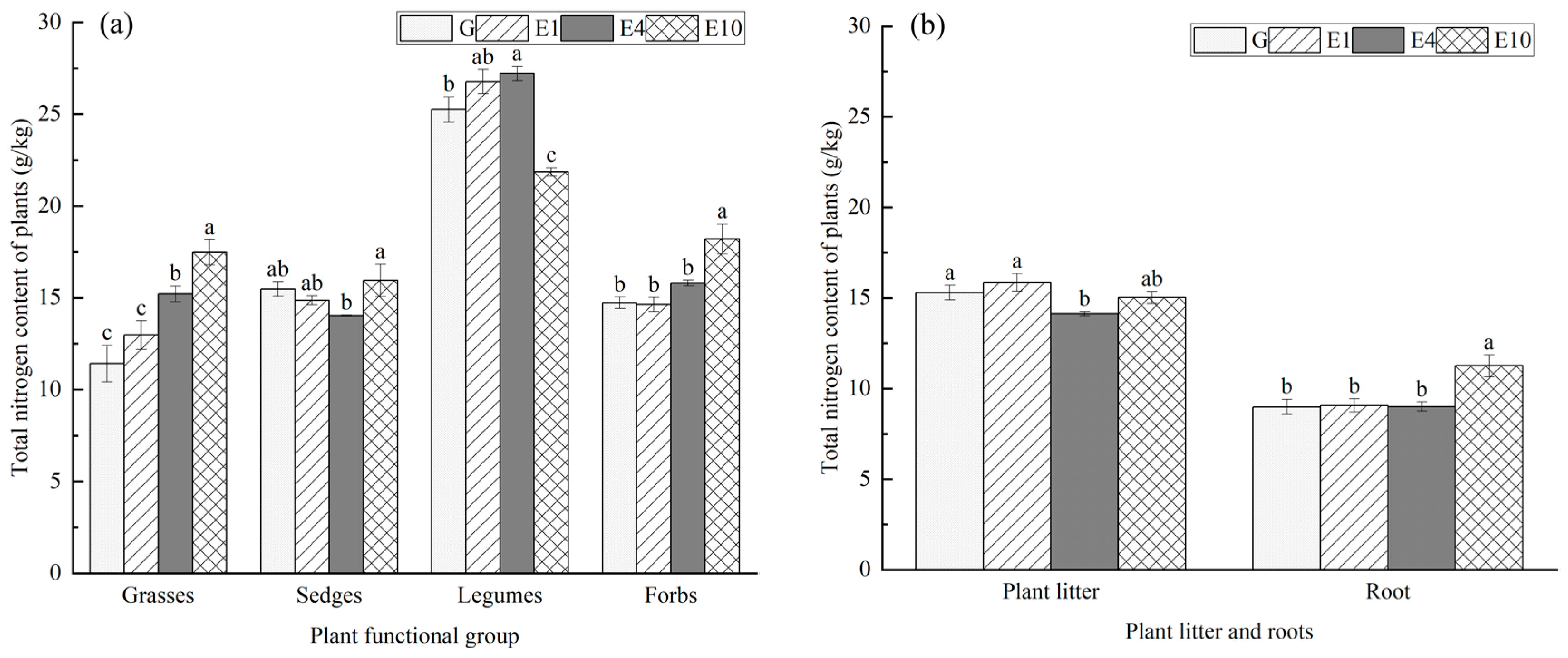
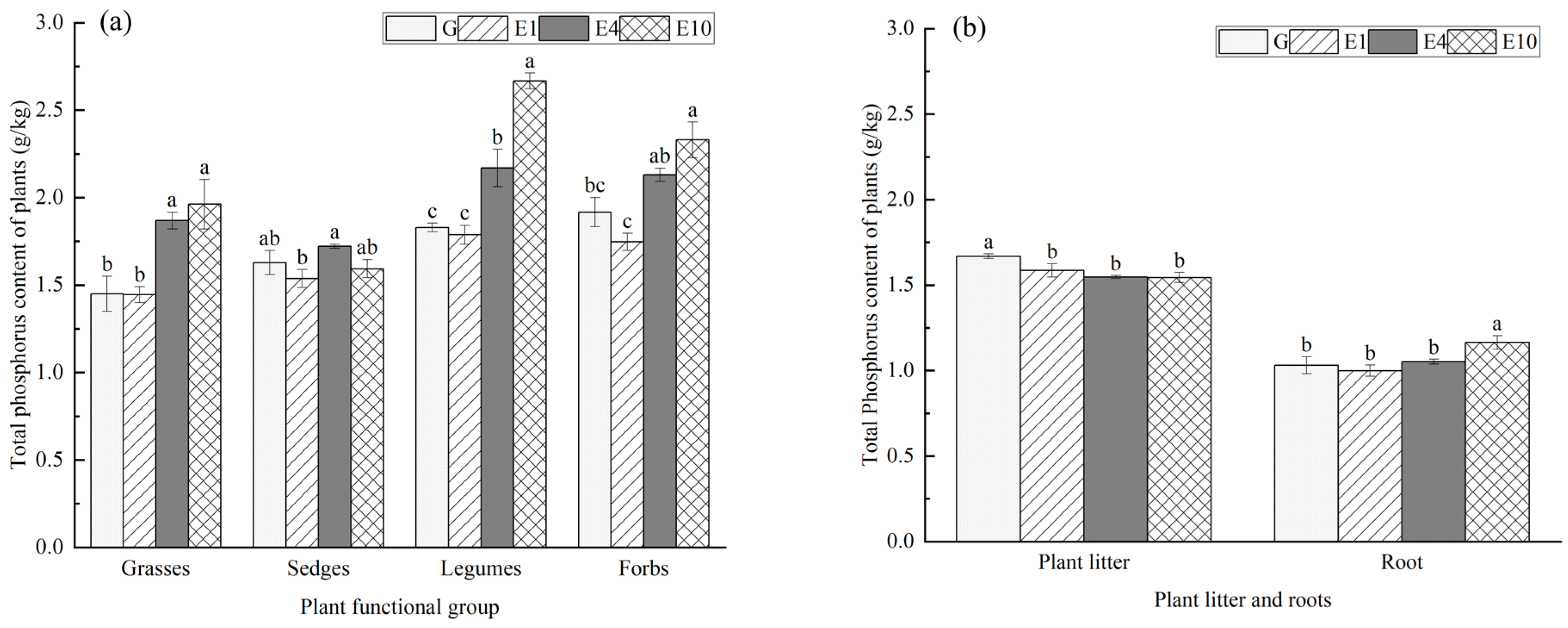
2.3.3. Determination of Plant Carbon, Nitrogen, and Phosphorus
2.4. Data Analysis and Representation
3. Results
3.1. Plant Characteristics by Functional Group
3.2. Main Plant Characteristics by Functional Group
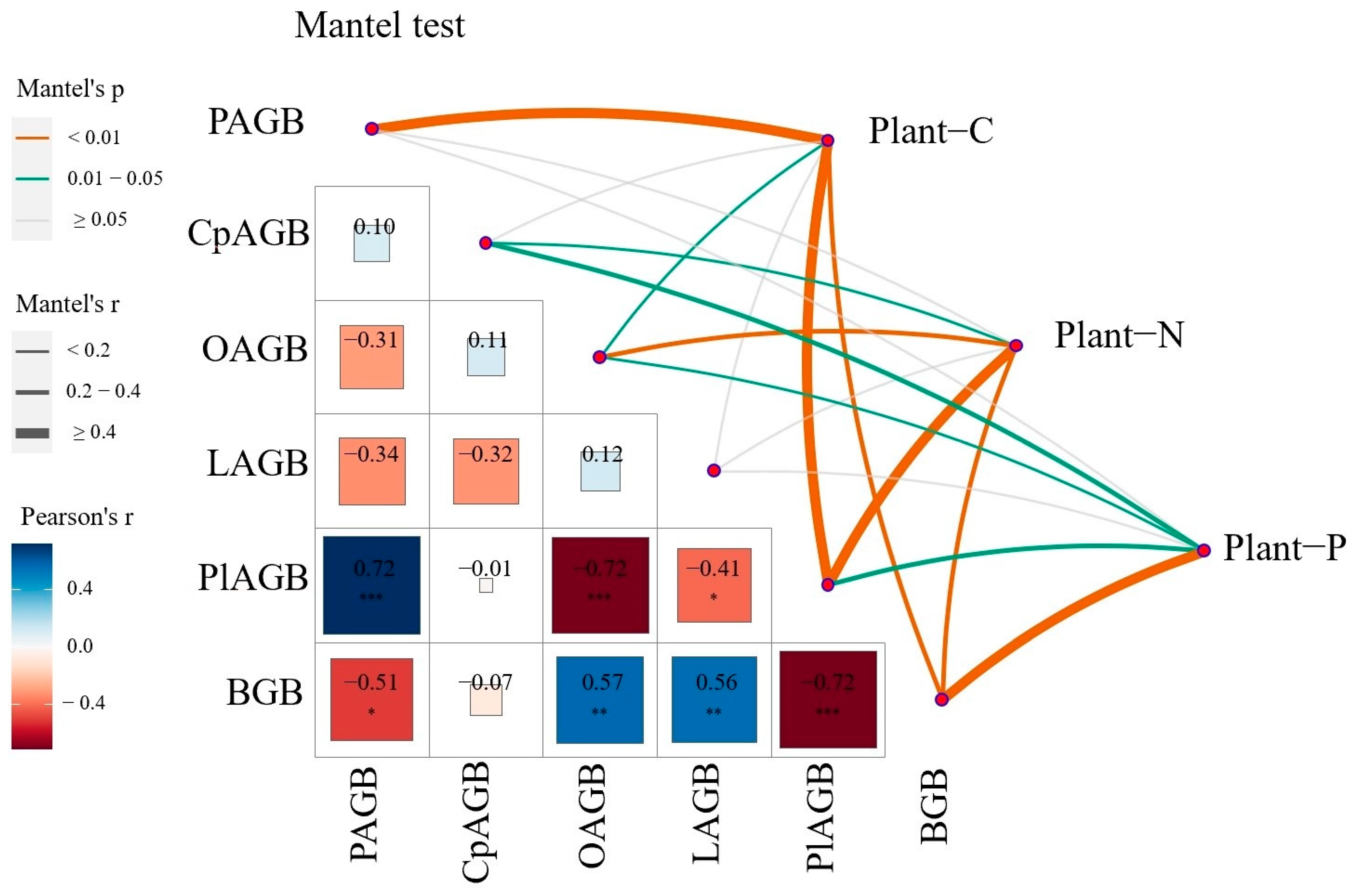
3.3. Correlation between Plant Carbon, Nitrogen, and Phosphorus Content and Biomass
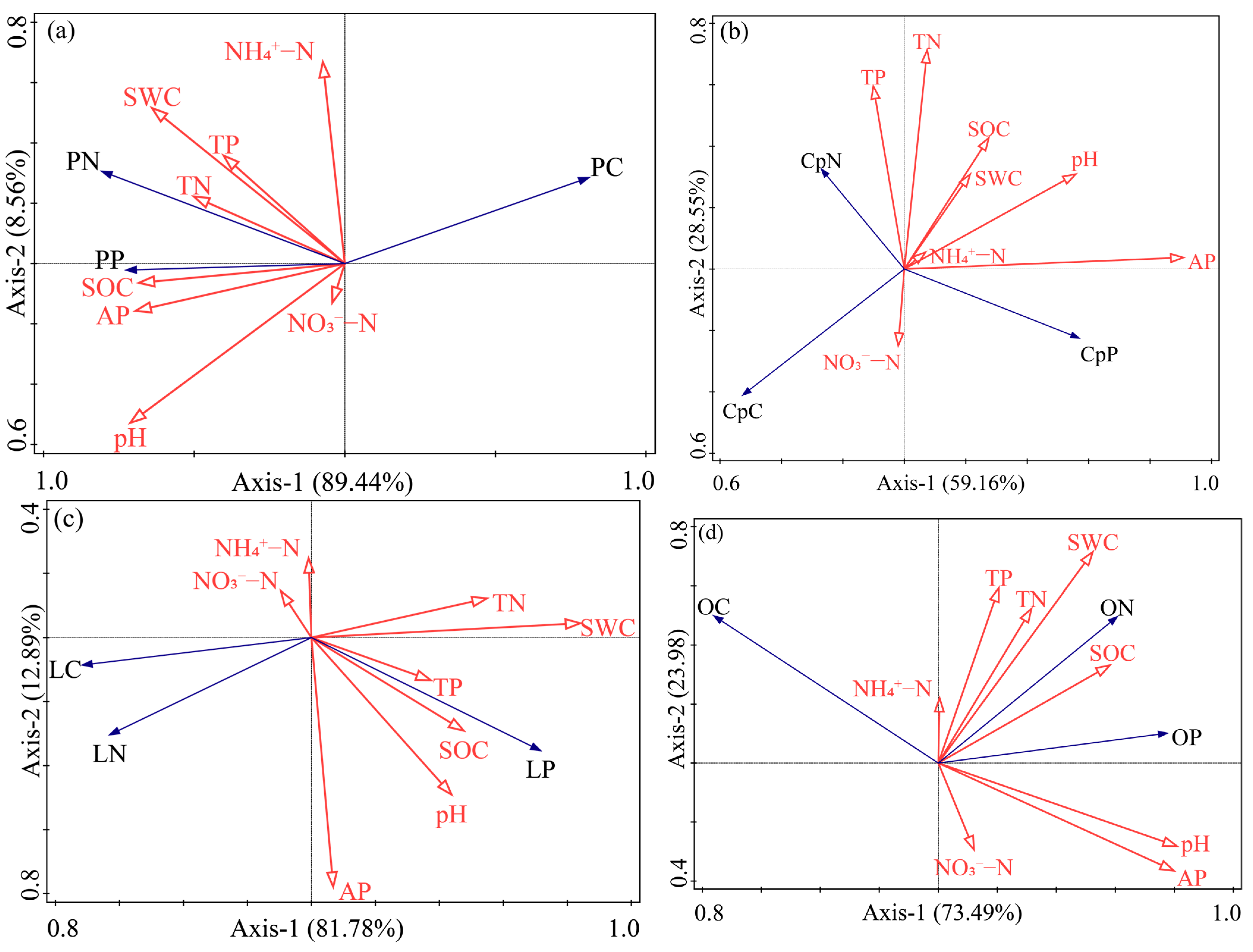
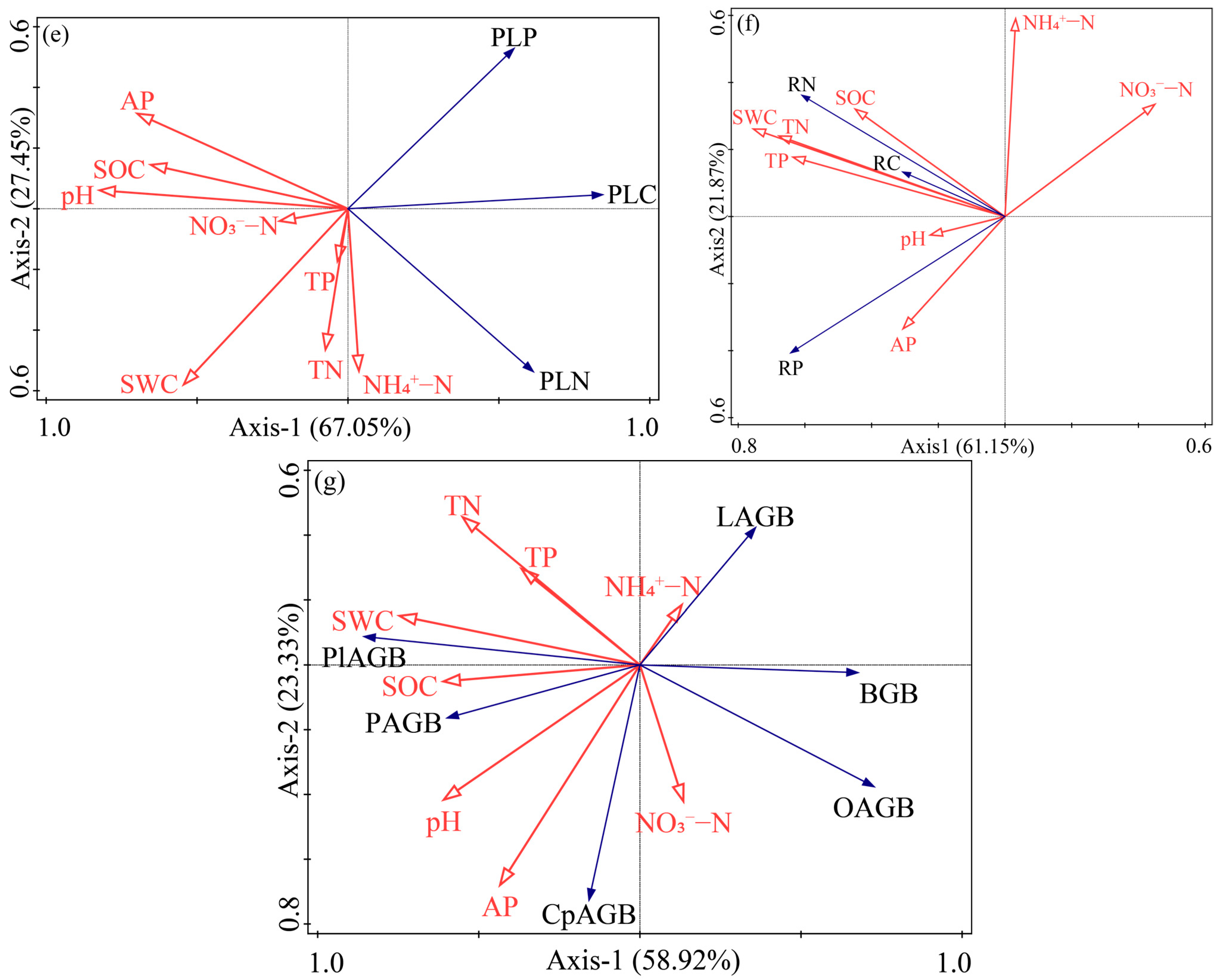
3.4. Relationship between Plant Carbon, Nitrogen, Phosphorus Content and the Environment
4. Discussion
4.1. Effects of Long-Term Exclosure on the Diversity and Biomass of Main Plant Functional Groups
4.2. Effects of Long-Term Exclosure on Carbon, Nitrogen, and Phosphorus Contents of Plants
4.3. Effects of Long-Term Exclosure on Plant Stoichiometric Characteristics
4.4. Relationship between Enclosed Grassland and Environmental Factors
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Exclosure Duration | pH | SWC/% | ω (SOC)/g·kg−1 | ω (TN)/g·kg−1 | ω (TP)/g·kg−1 | ω (NH4+−N)/ mg·kg−1 | ω (NO3−−N)/ mg·kg−1 | ω (AP)/mg·kg−1 |
|---|---|---|---|---|---|---|---|---|
| G | 6.46 ± 0.06 c | 18.91 ± 0.01 b | 93.38 ± 8.63 ab | 3.85 ± 0.07 c | 0.67 ± 0.01 b | 9.00 ± 0.79 a | 17.14 ± 1.14 ab | 7.12 ± 0.38 b |
| E1 | 6.30 ± 0.06 d | 21.65 ± 0.02 b | 83.56 ± 9.52 b | 4.37 ± 0.10 ab | 0.75 ± 0.03 a | 9.65 ± 0.93 a | 14.78 ± 0.76 b | 7.05 ± 0.18 b |
| E4 | 6.91 ± 0.06 a | 21.88 ± 0.01 b | 115.01 ± 0.60 a | 4.00 ± 0.24 bc | 0.72 ± 0.01 ab | 9.17 ± 0.49 a | 18.65 ± 0.05 a | 9.51 ± 0.47 a |
| E10 | 6.68 ± 0.03 b | 26.15 ± 0.01 a | 116.75 ± 10.09 a | 4.66 ± 0.15 a | 0.78 ± 0.03 a | 8.86 ± 0.26 a | 14.95 ± 0.90 b | 7.84 ± 0.43 b |
References
- Fu, B.; Li, S.; Yu, X.; Yang, P.; Yu, G.; Feng, R.; Zhuang, X. Chinese ecosystem research network: Progress and perspectives. Ecol. Complex. 2010, 7, 225–233. [Google Scholar] [CrossRef]
- Bardgett, R.D.; Bullock, J.M.; Lavorel, S.; Manning, P.; Schaffner, U.; Ostle, N.; Chomel, M.; Durigan, G.; Fry, E.L.; Johnson, D.; et al. Combatting global grassland degradation. Nat. Rev. Earth Environ. 2021, 2, 720–735. [Google Scholar] [CrossRef]
- Shao, Q.; Cao, W.; Fan, J.; Huang, L.; Xu, X. Effects of an ecological conservation and restoration project in the Three-River Source Region, China. J. Geogr. 2017, 27, 183–204. [Google Scholar] [CrossRef]
- Zhang, L.; Xiao, P.; Yu, H.; Zhao, T.; Liu, S.; Yang, L.; He, Y.; Luo, Y.L.; Wang, X.; Dong, W.; et al. Effects of Climate Changes on the Pasture Productivity From 1961 to 2016 in Sichuan Yellow River Source, Qinghai-Tibet Plateau, China. Front. Ecol. Evol. 2022, 10, 908924. [Google Scholar] [CrossRef]
- Liu, L.; Zhang, Y.; Bai, W.; Yan, J.; Ding, M.; Shen, Z.; Li, S.; Zheng, D. Characteristics of grassland degradation and driving forces in the source region of the Yellow River from 1985 to 2000. J. Geogr. 2006, 16, 131–142. [Google Scholar] [CrossRef]
- Dong, S.C.; Zhou, C.J.; Wang, H.Y. Ecological crisis and countermeasures of the Three Rivers’ Headstream Regions. J. Nat. Resour. 2002, 17, 713–720. [Google Scholar]
- Martin, D.M. Ecological restoration should be redefined for the twenty-first century. Restor. Ecol. 2017, 25, 668–673. [Google Scholar] [CrossRef]
- Ren, Y.; Lü, Y.; Fu, B. Quantifying the impacts of grassland restoration on biodiversity and ecosystem services in China: A meta-analysis. Ecol. Eng. 2016, 95, 542–550. [Google Scholar] [CrossRef]
- Zhang, Z.H.; Zhou, H.K.; Zhao, X.Q.; Yao, B.Q.; Ma, Z.; Dong, Q.M.; Dong, Q.; Zhang, Z.H.; Wang, W.Y.; Yang, Y.W. Relationship between biodiversity and ecosystem functioning in alpine meadows of the Qinghai-Tibet Plateau. Biodivers. Sci. 2018, 26, 111. [Google Scholar] [CrossRef]
- Xue, Y.F.; Zong, N.; He, N.P.; Tian, J.; Zhang, Y.Q. Influence of long-term enclosure and free grazing on soil microbial community structure and carbon metabolic diversity of alpine meadow. J. Appl. Ecol. 2018, 29, 2705–2712. [Google Scholar]
- Jachowski, D.S.; Slotow, R.; Millspaugh, J.J. Good virtual fences make good neighbors: Opportunities for conservation. Anim. Conserv. 2014, 17, 187–196. [Google Scholar] [CrossRef]
- Chen, Q.; Shang, Y.T.; Zhu, R.; Bao, Q.L.; Lin, S. Long-term enclosure at heavy grazing grassland affects soil nitrification via ammonia-oxidizing bacteria in Inner Mongolia. Sci. Rep. 2022, 12, 21464. [Google Scholar] [CrossRef]
- Zhang, Z.W.; Han, J.H.; Yin, H.Y.; Xue, J.; Jia, L.Z.; Zhen, X.; Chang, J.J.; Wang, S.K.; Yu, B. Assessing the effects of different long-term ecological engineering enclosures on soil quality in an alpine desert grassland area. Ecol. Indic. 2022, 143, 109426. [Google Scholar] [CrossRef]
- Shang, Z.H.; Cao, J.J.; Guo, R.Y.; Henkin, Z.; Ding, L.M.; Long, R.J.; Deng, B. Effect of enclosure on soil carbon, nitrogen and phosphorus of alpine desert rangeland. Land Degrad. Dev. 2017, 28, 1166–1177. [Google Scholar] [CrossRef]
- Asadian, G.; Javadi, S.A.; Jafary, M.; Arzani, H.; Akbarzade, M. Relationships between environmental factors and plant communities in enclosure rangelands (case study: Gonbad, Hamadan). J. Rangel. Sci. 2016, 7, 20–34. [Google Scholar]
- Bode, M.; Wintle, B. How to build an efficient conservation fence. Conserv. Biol. 2010, 24, 182–188. [Google Scholar] [CrossRef]
- Zuo, W.Q.; Wang, Y.H.; Wang, F.Y.; Shi, G.X. Effects of enclosure on the community characteristics of Leymus chinensis in degenerated steppe. Acta Prataculturae Sin. 2009, 18, 12. [Google Scholar]
- Yao, X.; Wu, J.; Gong, X.; Lang, X.; Wang, C.; Song, S.; Ahmad, A.A. Effects of long term fencing on biomass, coverage, density, biodiversity and nutritional values of vegetation community in an alpine meadow of the Qinghai-Tibet Plateau. Ecol. Eng. 2019, 130, 80–93. [Google Scholar] [CrossRef]
- Babur, E.; Kara, O.; Fathi, R.A.; Susam, Y.E.; Riaz, M.; Arif, M.; Akhtar, K. Wattle fencing improved soil aggregate stability, organic carbon stocks and biochemical quality by restoring highly eroded mountain region soil. J. Environ. Manag. 2021, 288, 112489. [Google Scholar] [CrossRef]
- Zhang, C.; Yan, R.R.; Liang, Q.W.; Na, R.S.; Li, T.; Yang, X.F.; Bao, Y.H.; Xin, X.P. Study on soil physical and chemical properties and carbon and nitrogen sequestration of grassland under different utilization modes. Acta Prataculturae Sin. 2021, 30, 90–98. [Google Scholar]
- Wu, G.L.; Du, G.Z.; Liu, Z.H.; Thirgood, S. Effect of fencing and grazing on a Kobresia-dominated meadow in the Qinghai-Tibetan Plateau. Plant Soil 2009, 319, 115–126. [Google Scholar] [CrossRef]
- Xu, L.; Nie, Y.; Chen, B.; Xin, X.; Yang, G.; Xu, D.; Ye, L. Effects of fence enclosure on vegetation community characteristics and productivity of a degraded temperate meadow steppe in northern China. Appl. Sci. 2020, 10, 2952. [Google Scholar] [CrossRef]
- Astaiken, G.; Dong, Y.Q.; Zhou, S.J.; Nie, T.T.; Jiang, A.J.; An, S.Z. Effects of enclosure on plant community diversity and niche characteristics of different grassland types-Taking different types of grassland in Xinjiang as an example. Pratacultural Sci. 2023, 40, 1168–1185. [Google Scholar]
- Smith, D.; King, R.; Allen, B.L. Impacts of exclusion fencing on target and non-target fauna: A global review. Biol. Rev. 2020, 95, 1590–1606. [Google Scholar] [CrossRef] [PubMed]
- Boone, R.B.; Hobbs, N.T. Lines around fragments: Effects of fencing on large herbivores. Afr. J. Range Forage Sci. 2004, 21, 147–158. [Google Scholar] [CrossRef]
- Lu, R.K. Soil Agrochemical Analysis Method; China Agricultural Science and Technology Press: Beijing, China, 2000; pp. 266–292. [Google Scholar]
- Nie, Y.Y.; Xu, L.J.; Xin, X.P.; Chen, B.R.; Zhang, B.H. Effects of fence enclosure on the plant community composition and niche characteristics in a temperate meadow steppe. Acta Prataculturae Sin. 2020, 29, 11–22. [Google Scholar]
- Zhu, N.; Sun, J.; Shi, N.; Wang, J.N.; Zhang, L.; Luo, D.L.; Shen, C.; Gai, A.H. Effects of Short-term Fence Enclosing on Plant Community and the Physical and Chemica Properties of Alpine Meadow Soils. Acta Agrestia Sin. 2023, 31, 834–843. [Google Scholar]
- Fenetahun, Y.; Yuan, Y.; Xinwen, X.; Yongdong, W. Effects of grazing enclosures on species diversity, phenology, biomass, and carrying capacity in Borana Rangeland, Southern Ethiopia. Front. Ecol. Evol. 2021, 8, 623627. [Google Scholar] [CrossRef]
- Li, R.F.; Niu, H.S.; Kong, Q.; Liu, Q. Effects of exclosure on plant and soil nutrients in an alpine grassland. Pratacultural Sci. 2021, 38, 399–409. [Google Scholar]
- Loeser, M.R.R.; Sisk, T.D.; Crews, T.E. Impact of grazing intensity during drought in an Arizona grassland. Conserv. Biol. 2007, 21, 87–97. [Google Scholar] [CrossRef]
- Ebrahimi, M.; Khosravi, H.; Rigi, M. Short-term grazing exclusion from heavy livestock rangelands affects vegetation cover and soil properties in natural ecosystems of southeastern Iran. Ecol. Eng. 2016, 95, 10–18. [Google Scholar] [CrossRef]
- Inderjit. Plant invasions: Habitat invasibility and dominance of invasive plant species. Plant Soil 2005, 277, 1–5. [Google Scholar] [CrossRef]
- Liu, M.; Liu, G.; Wu, X.; Wang, H.; Chen, L. Vegetation traits and soil properties in response to utilization patterns of grassland in Hulun Buir City, Inner Mongolia, China. Chin. Geogr. Sci. 2014, 24, 471–478. [Google Scholar] [CrossRef]
- Yuan, J.; Li, H.; Yang, Y. The compensatory tillering in the forage grass Hordeum brevisubulatum after simulated grazing of different severity. Front. Plant Sci. 2020, 11, 792. [Google Scholar] [CrossRef] [PubMed]
- Qin, R.; Wang, X. Effects of crown height on the compensatory growth of Italian ryegrass based on combined effects of stored organic matter and cytokinin. Grassl. Sci. 2020, 66, 29–39. [Google Scholar] [CrossRef]
- Wang, C.T.; Long, R.J.; Wang, Q.L.; Jing, Z.C.; Shi, J.J. Changes in plant diversity, biomass and soil C, in alpine meadows at different degradation stages in the headwater region of three rivers, China. Land Degrad. Dev. 2009, 20, 187–198. [Google Scholar] [CrossRef]
- Zechmeister-Boltenstern, S.; Keiblinger, K.M.; Mooshammer, M.; Peñuelas, J.; Richter, A.; Sardans, J.; Wanek, W. The application of ecological stoichiometry to plant–microbial–soil organic matter transformations. Ecol. Monogr. 2015, 85, 133–155. [Google Scholar] [CrossRef]
- Malhotra, H.; Vandana; Sharma, S.; Pandey, R. Phosphorus nutrition: Plant growth in response to deficiency and excess. In Plant Nutrients and Abiotic Stress Tolerance; Hasanuzzaman, M., Fujita, M., Oku, H., Nahar, K., Hawrylak-Nowak, B., Eds.; Springer: Singapore, 2018; pp. 171–190. [Google Scholar]
- Elser, J.J.; Fagan, W.F.; Kerkhoff, A.J.; Swenson, N.G.; Enquist, B.J. Biological stoichiometry of plant production: Metabolism, scaling and ecological response to global change. New Phytol. 2010, 186, 593–608. [Google Scholar] [CrossRef]
- Song, S.; Wang, X.; He, C.; Chi, Y. Effects of Utilization Methods on C, N, P Rate and Enzyme Activity of Artificial Grassland in Karst Desertification Area. Agronomy 2023, 13, 1368. [Google Scholar] [CrossRef]
- Yu, Q.; Chen, Q.S.; Elser, J.J.; He, N.P.; Wu, H.H.; Zhang, G.M.; Wu, J.G.; Bai, Y.F.; Han, X.G. Linking stoichiometric homoeostasis with ecosystem structure, functioning and stability. Ecol. Lett. 2010, 13, 1390–1399. [Google Scholar] [CrossRef]
- Cheng, J.; Jing, G.; Wei, L.; Jing, Z. Long-term grazing exclusion effects on vegetation characteristics, soil properties and bacterial communities in the semi-arid grasslands of China. Ecol. Eng. 2016, 97, 170–178. [Google Scholar] [CrossRef]
- Ayres, E.; Steltzer, H.; Simmons, B.L.; Simpson, R.T.; Steinweg, J.M.; Wallenstein, M.D.; Mellor, N.; Parton, W.J.; Moor, J.C.; Wall, D.H. Home-field advantage accelerates leaf litter decomposition in forests. Soil Biol. Biochem. 2009, 41, 606–610. [Google Scholar] [CrossRef]
- Wardle, D.A.; Bardgett, R.D.; Klironomos, J.N.; Setala, H.; Van Der Putten, W.H.; Wall, D.H. Ecological linkages between aboveground and belowground biota. Science 2004, 304, 1629–1633. [Google Scholar] [CrossRef] [PubMed]
- Van der Putten, W.H.; Bardgett, R.D.; Bever, J.D.; Bezemer, T.M.; Casper, B.B.; Fukami, T.; Wardle, D.A. Plant–soil feedbacks: The past, the present and future challenges. J. Ecol. 2013, 101, 265–276. [Google Scholar] [CrossRef]
- McGroddy, M.E.; Daufresne, T.; Hedin, L.O. Scaling of C:N:P stoichiometry in forests worldwide: Implications of terrestrial redfield-type ratios. Ecology 2004, 85, 2390–2401. [Google Scholar] [CrossRef]
- Yu, Y.C.; Yang, J.Y.; Zeng, S.C.; Wu, D.M.; Jacobs, D.F.; Sloan, J.L. Soil pH, organic matter, and nutrient content change with the continuous cropping of Cunninghamia lanceolata plantations in South China. J. Soils Sediments 2017, 17, 2230–2238. [Google Scholar] [CrossRef]
- Filser, J.; Faber, J.H.; Tiunov, A.V.; Brussaard, L.; Frouz, J.; De Deyn, G.; Uvarov, A.V.; Berg, M.P.; Lavelle, P.; Loreau, M.; et al. Soil fauna: Key to new carbon models. Soil 2016, 2, 565–582. [Google Scholar] [CrossRef]
- Liu, T.Y.; Zhou, T.C.; Sun, J.; Wang, Y.; Ye, C.C. Distribution and coupling characteristics of plant nitrogen and phosphorus along desertification gradients in alpine meadows, eastern Tibet Plateau. Pratacultural. Sci. 2021, 38, 209–220. [Google Scholar]
- Hati, K.M.; Swarup, A.; Mishra, B.; Manna, M.C.; Wanjari, R.H.; Mandal, K.G.; Misra, A.K. Impact of long-term application of fertilizer, manure and lime under intensive cropping on physical properties and organic carbon content of an Alfisol. Geoderma 2008, 148, 173–179. [Google Scholar] [CrossRef]
- Yang, P.N.; Li, X.L.; Li, C.Y.; Duan, C.W. Response of Soil Microbial Diversity to Long-term Enclosure in Degraded Patches of Alpine Meadow in the Source Zone of the Yellow River. Huan Jing Ke Xue = Huanjing Kexue 2023, 44, 2293–2303. [Google Scholar]








| Exclosure Duration | Plot Location | Elevation/m | Patch Numbers | Patch Density (Area Ratio)/% | The Main Composition of Plant Community |
|---|---|---|---|---|---|
| G | E 101°46′06″, N 34°41′02″ | 3610 | 1086.45 | 41.64 | Elymus nutans Griseb., Koeleria cristata L., Kobresia pygmaea Willd., Ligularia virgaurea, Gentiana scabra Bunge, Pedicularis spicata, Oxytropis ochrocephala, Carun buriaticum, Gentiana macrophylla Pall., and Taraxacum officinalis |
| E1 | E 101°46′08″, N 34°41′04″ | 3610 | 989.52 | 36.41 | Poa annua L., Elymus nutans Griseb., Kobresia pygmaea Willd., Koeleria cristata L., Potentilla anserina, Pedicularis spicata, Saussurea pulchra, Gentiana scabra Bunge, and Ligularia virgaurea |
| E4 | E 101°46′10″, N 34°41′06″ | 3610 | 536.71 | 18.63 | Elymus nutans Griseb., Poa annua L., Koeleria cristata L., Potentilla anserina, Ajania tenuifolia, Astragalus sp., and Gentiana scabra Bunge |
| E10 | E 101°26′34″, N 34°18′36″ | 3720 | 331.81 | 6.67 | Elymus nutans Griseb., Poa annua L., Koeleria cristata L., Carex L., Potentilla anserina, and Gentiana scabra Bunge |
| Group/Type | df | F (C) | F (N) | F (P) | F (AGB) | F (Index) |
|---|---|---|---|---|---|---|
| Grasses | 3 | 17.52 ** | 12.45 ** | 8.64 ** | 8.92 ** | |
| Sedges | 3 | 4.60 * | 2.75 ns | 2.34 ns | 3.80 * | |
| Legumes | 3 | 12.64 ** | 21.26 ** | 39.20 ** | 2.47 ns | |
| Forbs | 3 | 14.50 ** | 11.92 ** | 12.15 ** | 10.3 ** | |
| Plant litter | 3 | 20.01 ** | 3.95 * | 5.11 ** | 82.94 ** | |
| Root | 3 | 1.87 ns | 6.79 ** | 3.96 * | 7.16 ** | |
| D | 3 | 37.63 ** | ||||
| H | 3 | 32.70 ** | ||||
| E | 3 | 38.80 ** | ||||
| Group/Type | df | F (C/N) | F (C/P) | F (N/P) |
|---|---|---|---|---|
| Grasses | 3 | 9.61 ** | 11.05 ** | 1.61 ns |
| Sedges | 3 | 2.69 ns | 2.96 ns | 3.85 * |
| Legumes | 3 | 10.32 ** | 43.60 ** | 65.73 ** |
| Forbs | 3 | 14.07 ** | 13.57 ** | 1.24 ns |
| Plant litter | 3 | 1.64 ns | 1.65 ns | 1.80 ns |
| Root | 3 | 0.56 ns | 0.40 ns | 1.06 ns |
| Exclosure Duration | Grasses | Sedges | Legumes | Forbs | Plant Litter | Root |
|---|---|---|---|---|---|---|
| G | 40.50 ± 3.43 a | 28.85 ± 0.77 a | 17.78 ± 0.59 b | 30.14 ± 0.64 a | 28.21 ± 0.79 a | 39.22 ± 2.88 a |
| E1 | 35.11 ± 2.59 a | 29.50 ± 0.40 a | 16.72 ± 0.40 bc | 30.52 ± 0.83 a | 27.32 ± 0.66 a | 40.24 ± 3.19 a |
| E4 | 28.41 ± 0.79 b | 30.71 ± 0.10 a | 16.19 ± 0.20 c | 26.80 ± 0.16 b | 28.30 ± 0.42 a | 38.96 ± 1.59 a |
| E10 | 24.84 ± 1.07 b | 27.42 ± 1.43 a | 18.98 ± 0.21 a | 24.40 ± 1.13 c | 26.74 ± 0.35 a | 35.61 ± 2.85 a |
| Exclosure Duration | Grasses | Sedges | Legumes | Forbs | Plant Litter | Root |
|---|---|---|---|---|---|---|
| G | 313.89 ± 19.97 a | 275.68 ± 12.33 a | 244.64 ± 5.12 a | 233.21 ± 10.25 a | 257.71 ± 2.53 a | 344.54 ± 30.73 a |
| E1 | 309.91 ± 9.49 a | 286.21 ± 7.78 a | 250.81 ± 7.72 a | 255.60 ± 6.68 a | 272.99 ± 6.95 a | 363.32 ± 24.97 a |
| E4 | 231.19 ± 6.17 b | 250.16 ± 2.01 a | 205.20 ± 9.16 b | 199.17 ± 4.90 b | 258.39 ± 0.42 a | 331.79 ± 8.11 a |
| E10 | 225.76 ± 17.76 b | 271.89 ± 9.65 a | 155.60 ± 2.69 c | 190.83 ± 9.70 b | 260.59 ± 8.33 a | 338.33 ± 14.65 a |
| Exclosure Duration | Grasses | Sedges | Legumes | Forbs | Grasses | Root |
|---|---|---|---|---|---|---|
| G | 7.87 ± 0.46 a | 9.62 ± 0.59 a | 13.80 ± 0.25 b | 7.74 ± 0.32 a | 9.17 ± 0.30 a | 8.82 ± 0.63 a |
| E1 | 9.00 ± 0.52 a | 9.71 ± 0.30 a | 15.02 ± 0.48 a | 8.39 ± 0.22 a | 10.04 ± 0.43 a | 9.09 ± 0.33 a |
| E4 | 8.15 ± 0.16 a | 8.14 ± 0.05 b | 12.66 ± 0.48 c | 7.43 ± 0.18 a | 9.14 ± 0.13 a | 8.55 ± 0.21 a |
| E10 | 9.12 ± 0.66 a | 10.02 ± 0.54 a | 8.20 ± 0.13 d | 7.91 ± 0.58 a | 9.76 ± 0.39 a | 9.72 ± 0.63 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, P.; Li, X.; Li, C.; Zhang, J. Effects of Long-Term Exclosure on Main Plant Functional Groups and Their Biochemical Properties in a Patchily Degraded Alpine Meadow in the Source Zone of the Yellow River, West China. Agronomy 2023, 13, 2781. https://doi.org/10.3390/agronomy13112781
Yang P, Li X, Li C, Zhang J. Effects of Long-Term Exclosure on Main Plant Functional Groups and Their Biochemical Properties in a Patchily Degraded Alpine Meadow in the Source Zone of the Yellow River, West China. Agronomy. 2023; 13(11):2781. https://doi.org/10.3390/agronomy13112781
Chicago/Turabian StyleYang, Pengnian, Xilai Li, Chenyi Li, and Jing Zhang. 2023. "Effects of Long-Term Exclosure on Main Plant Functional Groups and Their Biochemical Properties in a Patchily Degraded Alpine Meadow in the Source Zone of the Yellow River, West China" Agronomy 13, no. 11: 2781. https://doi.org/10.3390/agronomy13112781