
Responses of Soil Bacterial Communities and Chemical Properties to Grazing Regulation in Desert Steppe
Abstract
:1. Introduction
2. Material and Methods
2.1. Study Site
2.2. Sampling
2.3. Soil Chemical Properties Analysis
2.4. Molecular Characterization of the Soil Bacteria
2.5. Bioinformation Analysis and Statistical Analysis
3. Results
3.1. Soil Properties under Different Stocking Density Groups
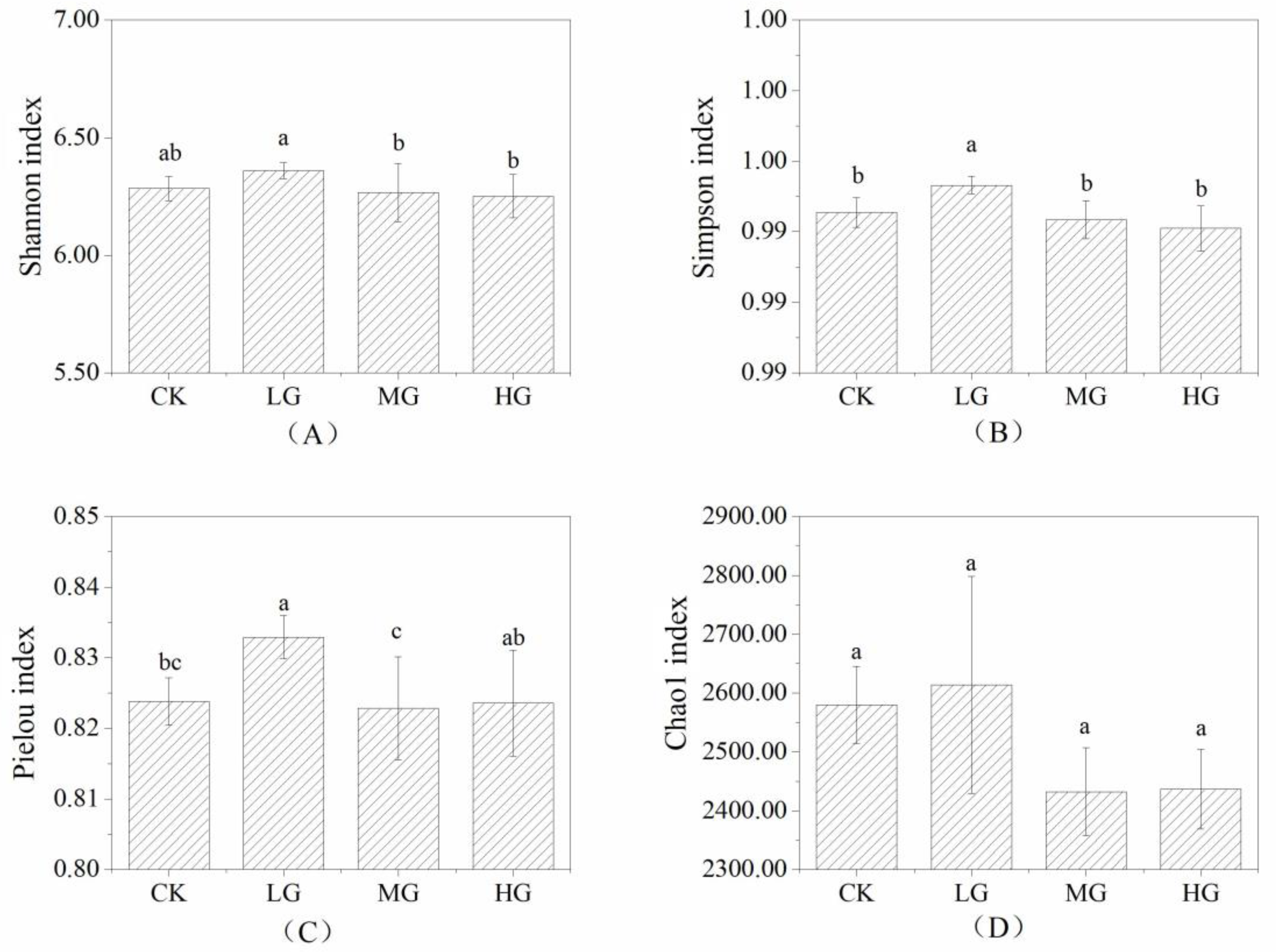
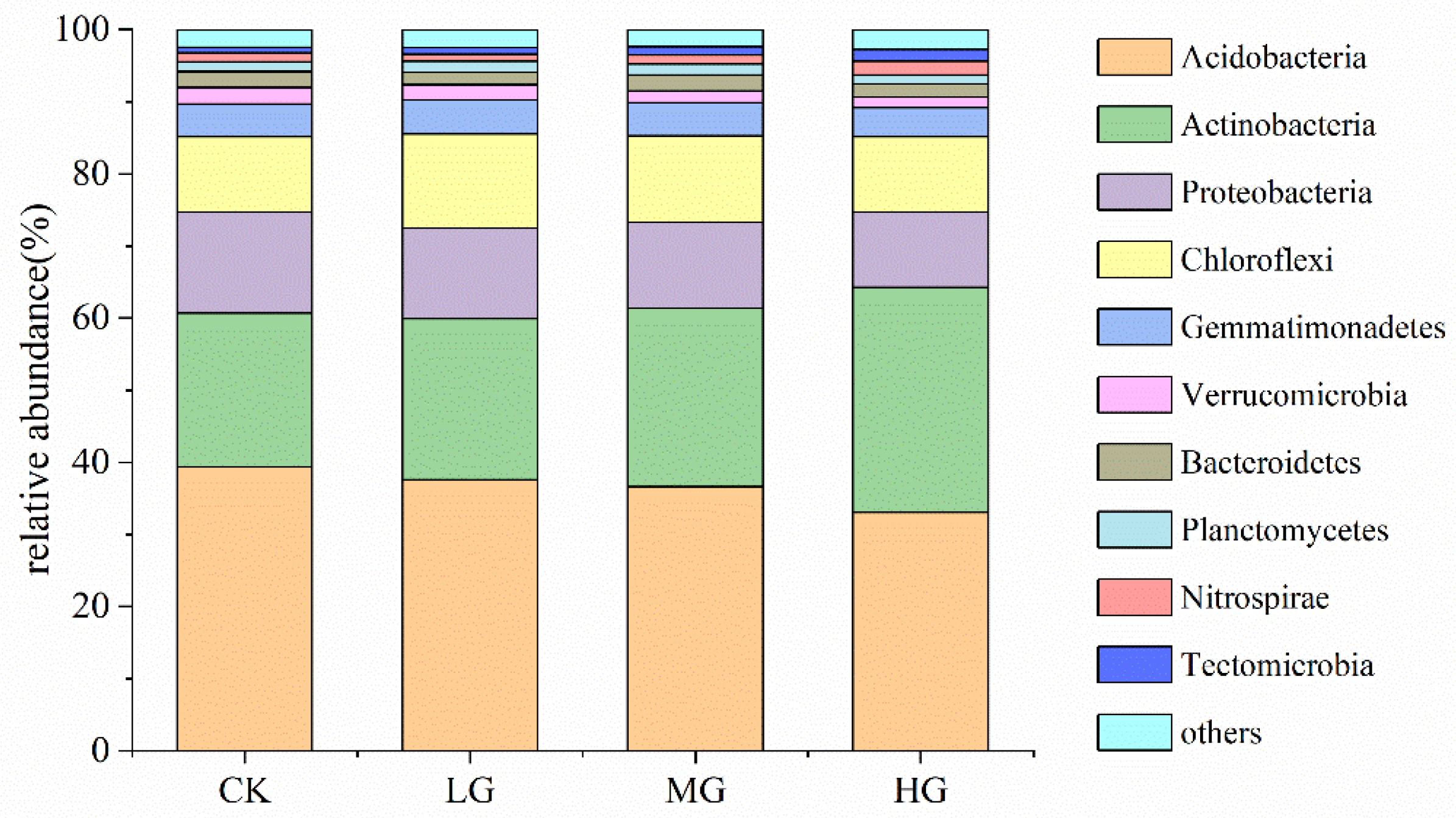
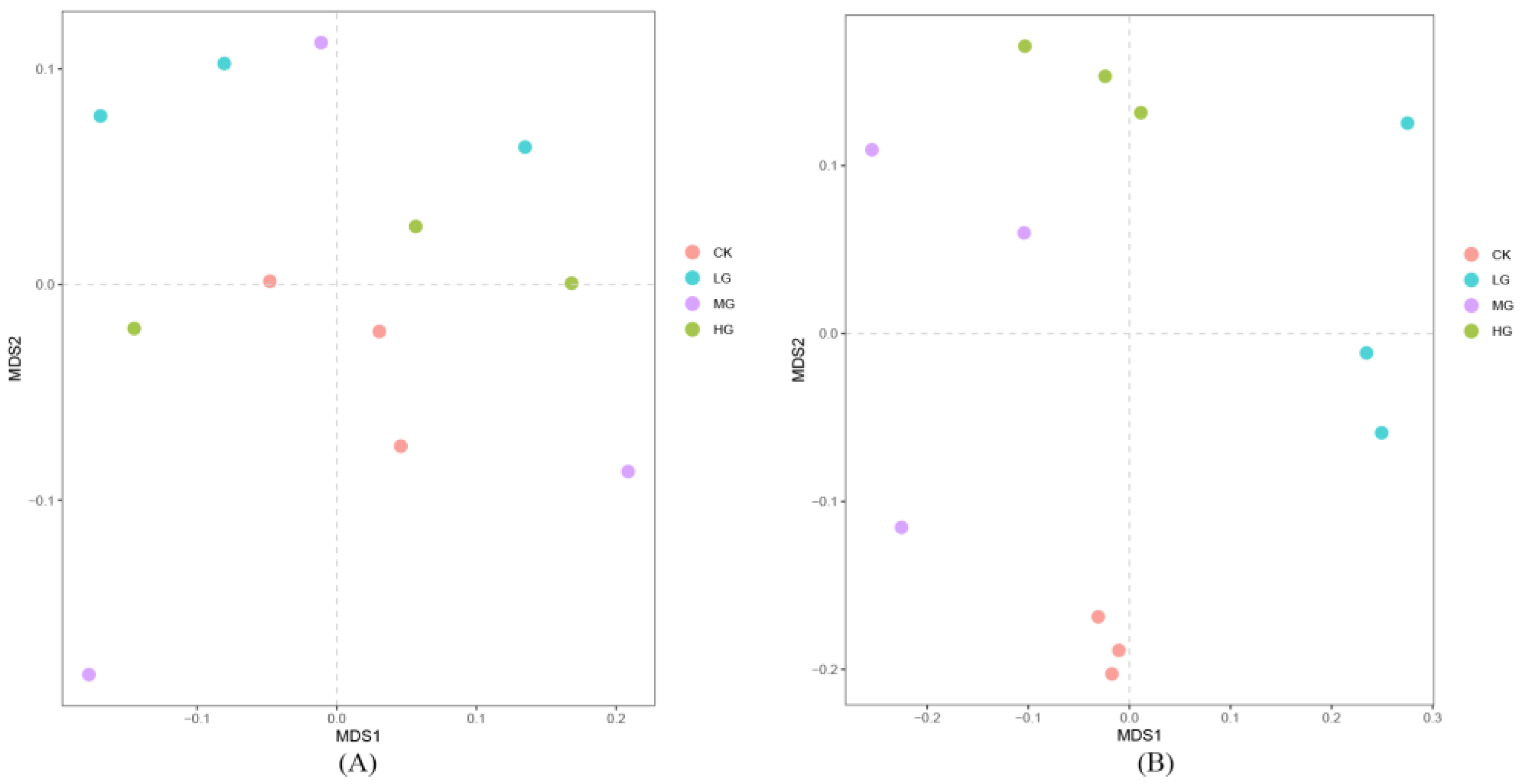
3.2. The Composition and Structure of the Soil Bacterial Community under Different Grazing Intensities
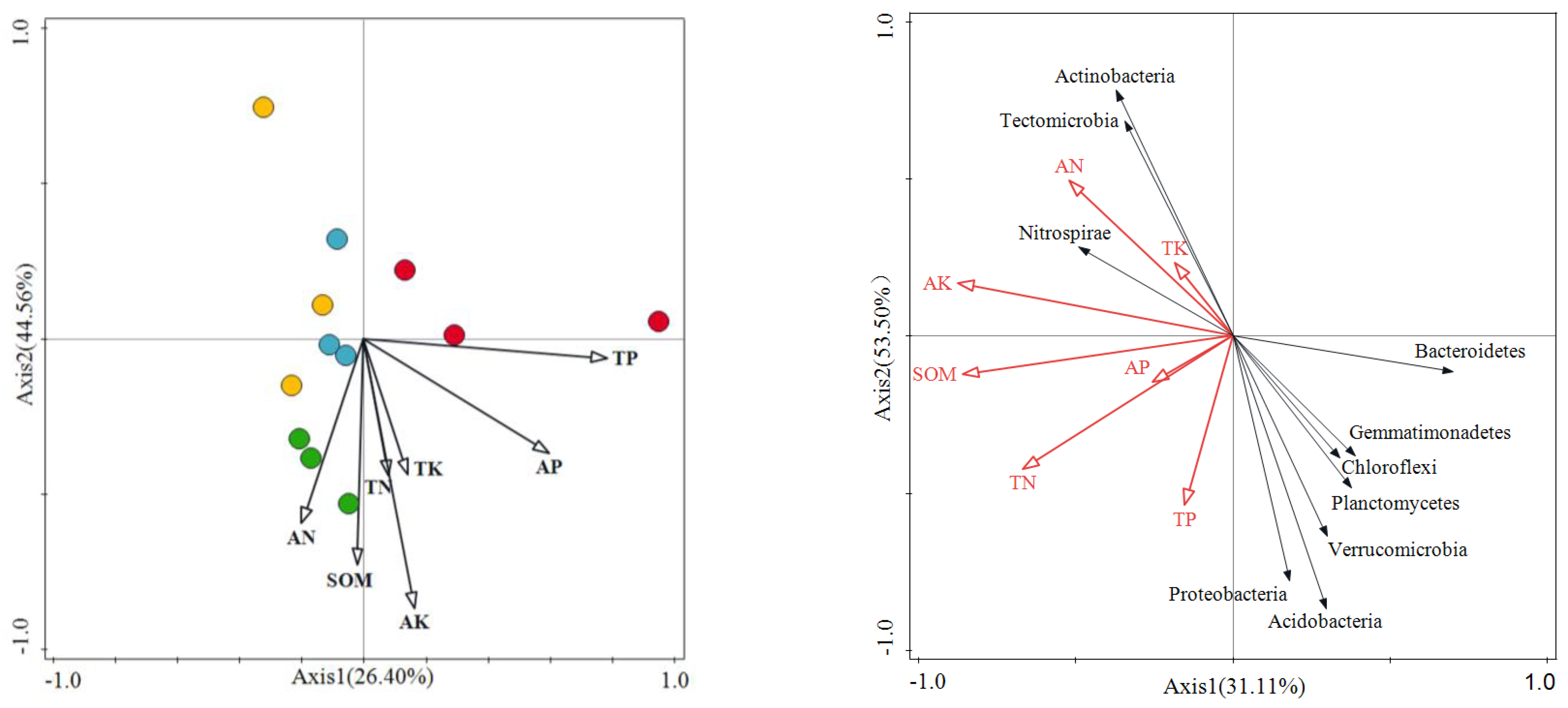
3.3. Correlations between Soil Bacteria and Soil Properties under Different Grazing Intensities
4. Discussion
4.1. Effects of Stocking Density on Soil Nutrient Content
4.2. Responses of Soil Bacterial Diversity and Community Structure to Stocking Density
4.3. The Correlation between Stocking Density, Soil Nutrient Content, and Soil Bacteria
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Delgado-Baquerizo, M.; Maestre, F.T.; Reich, P.B.; Jeffries, T.C.; Gaitan, J.J.; Encinar, D.; Berdugo, M.; Campbell, C.D.; Singh, B.K. Microbial diversity drives multifunctionality in terrestrial ecosystems. Nat. Commun. 2016, 7, 10541. [Google Scholar] [CrossRef]
- Geisen, S.; Wall, D.H.; van der Putten, W.H. Challenges and opportunities for soil biodiversity in the anthropocene. Curr. Biol. 2019, 29, 1036–1044. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Zhao, Z.; Yin, X.; Zhu, Y. Impacts of biochars on bacterial community shifts and biodegradation of antibiotics in an agricultural soil during short-term incubation. Sci. Total Environ. 2021, 771, 144751. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.C.; Shi, Z.Q.; Yang, J.Y.; Hao, B.H.; Hao, L.J.; Diao, F.W.; Wang, L.X.; Bao, Z.H.; Guo, W. A new strategy for evaluating the improvement effectiveness of degraded soil based on the synergy and diversity of microbial ecological function. Ecol. Indic. 2021, 120, 106917. [Google Scholar] [CrossRef]
- Xue, Y.; Bai, X.; Zhao, C.; Tan, Q.; Li, Y.; Luo, G.; Long, M. Spring photosynthetic phenology of Chinese vegetation in response to climate change and its impact on net primary productivity. Agric. For. Meteorol. 2023, 342, 109734. [Google Scholar] [CrossRef]
- Qiu, D.; Zhu, G.; Lin, X.; Jiao, Y.; Lu, S.; Liu, J.; Chen, L. Dissipation and movement of soil water in artificial forest in arid oasis areas: Cognition based on stable isotopes. CATENA 2023, 228, 107178. [Google Scholar] [CrossRef]
- Pan, D.R.; Yan, H.W.; Li, Q.; Liu, D.Y.; Liu, X.N.; Zhang, D.G.; Han, T.H.; Sun, B.; Jiang, J.C. Loss of grassland ecosystem service values based on potential vegetation in China. Rangel. J. 2022, 43, 363–375. [Google Scholar] [CrossRef]
- Li, M.Y.; Li, X.B.; Liu, S.Y.; Li, X.; Lyu, X.; Dang, D.L.; Dou, H. Ecosystem services under different grazing intensities in typical grasslands in Inner Mongolia and their relationships. Glob. Ecol. Conserv. 2021, 26, e01526. [Google Scholar] [CrossRef]
- Akiyama, T.; Kawamura, K. Grassland degradation in China: Methods of monitoring, management and restoration. Grassl. Sci. 2007, 53, 1–17. [Google Scholar] [CrossRef]
- Zhang, H.Y.; Fan, J.W.; Shao, Q.Q.; Zhang, Y.X. Ecosystem dynamics in the ‘Returning Rangeland to Grassland’ programs, China. Acta Prataculturae Sin. 2016, 25, 1–15. [Google Scholar]
- Coban, O.; Deyn, G.B.; van der Ploeg, M. Soil microbiota as game-changers in restoration of degraded lands. Science 2022, 375, abe0725. [Google Scholar] [CrossRef] [PubMed]
- Walkley, A.; Black, I.A. An examination of the degtjareff method for determining soil organic matter, and a proposed modification of the chromic acid Titration method. Soil Sci. 1934, 37, 29–38. [Google Scholar] [CrossRef]
- Mason, C.J.; Edwards, M.; Riby, P.G.; Coe, G. The use of microwaves in the acceleration of digestion and colour development in the determination of total Kjeldahl nitrogen in soil. Analyst 1999, 124, 1719–1726. [Google Scholar] [CrossRef]
- Bao, S. Soil and Agricultural Chemistry Analysis, 3rd ed.; Agriculture Press of China: Beijing, China, 2000; p. 11. [Google Scholar]
- Liu, J.S.; Ma, Q.; Hui, X.L.; Ran, J.Y.; Ma, Q.X.; Wang, X.; Wang, Z.H. Long-term high-P fertilizer input decreased the total bacterial diversity but not phoD-harboring bacteria in wheat rhizosphere soil with available-P deficiency. Soil Biol. Biochem. 2020, 149, 107918. [Google Scholar] [CrossRef]
- Edgar, R. Search and Clustering Orders of Magnitude Faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Huntley, J.; Fierer, N.; Gormley, J.A.; Smith, G.; Knight, R. Ultra-high-throughput microbial community analysis on the Illumina HiSeq and MiSeq platforms. ISME J. 2012, 6, 1621–1624. [Google Scholar] [CrossRef]
- Shanon, C.E. A mathematical theory of communications. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef]
- Simpson, E.H. Measurement of diversity. Nature 1949, 168, 668. [Google Scholar] [CrossRef]
- Chao, A. Non-parametric estimation of the number of classes in a population. Scand. J. Stat. 1984, 11, 265–270. [Google Scholar]
- Pielou, E.C. Species-diversity and pattern-diversity in the study of ecological succession. J. Theor. Biol. 1966, 10, 370–383. [Google Scholar] [CrossRef]
- Haynes, R.; Williams, P.H. Nutrient Cycling and Soil Fertility in the Grazed Pasture Ecosystem. Adv. Agron. 1993, 49, 119–199. [Google Scholar]
- Ma, W.M.; Ding, K.Y.; Zhong, W.L. Comparison of soil carbon and nitrogen stocks at grazing-excluded and yak grazed alpine meadow sites in Qinghai-Tibetan Plateau, China. Ecol. Eng. 2016, 87, 203–211. [Google Scholar] [CrossRef]
- Rakkar, M.K.; Blanco-Canqui, H. Grazing of crop residues: Impacts on soils and crop production. Agric. Ecosyst. Environ. 2018, 258, 71–90. [Google Scholar] [CrossRef]
- Zhu, G.Y.; Deng, L.; Zhang, X.B.; Shangguan, Z.P. Effects of grazing exclusion on plant community and soil physicochemical properties in a desert steppe on the Loess Plateau, China. Ecol. Eng. 2016, 90, 372–381. [Google Scholar] [CrossRef]
- Stewart Ibarra, A.; Frank, D. Short sampling intervals reveal very rapid root turnover in temperate grassland. Oecologia 2008, 157, 453–458. [Google Scholar] [CrossRef]
- Zhou, X.; Wang, J.; Hao, Y.; Wang, Y. Intermediate grazing intensities by sheep increase soil bacterial diversities in an Inner Mongolian steppe. Biol. Fert. Soils 2010, 46, 817–824. [Google Scholar] [CrossRef]
- Zhang, Y.T.; Gao, X.L.; Hao, X.Y.; Alexander, T.; Shi, X.J.; Jin, L.; Thomas, B.W. Heavy grazing over 64 years reduced soil bacterial diversity in the foothills of the Rocky Mountains, Canada. Appl. Soil Ecol. 2020, 147, 103361. [Google Scholar] [CrossRef]
- Wang, Z.; Jiang, S.Y.; Struik, P.C.; Wang, H.; Jin, K.; Wu, R.; Na, R.; Mu, H.B.; Ta, N. Plant and soil responses to grazing intensity drive changes in the soil microbiome in a desert steppe. Plant Soil 2022, 491, 219–237. [Google Scholar] [CrossRef]
- Li, Y.M.; Wang, S.P.; Jiang, L.L.; Zhang, L.R.; Cui, S.J.; Meng, F.D.; Wang, Q.; Li, X.; Zhou, Y. Changes of soil microbial community under different degraded gradients of alpine meadow. Agr. Ecosyst. Environ. 2016, 222, 213–222. [Google Scholar] [CrossRef]
- Xu, S.; Silveira, M.; Inglett, K.; Sollenberger, L.; Gerber, S. Soil microbial community responses to long-term land use intensification in subtropical grazing lands. Geoderma 2017, 293, 73–81. [Google Scholar] [CrossRef]
- Wang, M.M.; Wang, S.P.; Wu, L.W.; Xu, D.P.; Lin, Q.Y.; Yi, G.H.; Li, X.Z.; Zhou, J.Z.; Yang, Y.F. Evaluating the lingering effect of livestock grazing on functional potentials of microbial communities in Tibetan grassland soils. Plant Soil 2016, 407, 385–399. [Google Scholar] [CrossRef]
- Baker, K.L.; Langenheder, S.; Nicol, G.W.; Ricketts, D.; Killham, K.; Campbell, C.D.; Prosser, J.I. Environmental and spatial characterisation of bacterial community composition in soil to inform sampling strategies. Soil Biol. Biochem. 2009, 41, 2292–2298. [Google Scholar] [CrossRef]
- Yin, Y.L.; Wang, Y.Q.; Li, S.X.; Liu, Y.; Zhao, W.; Ma, Y.S.; Bao, G.S. Effects of enclosing on soil microbial community diversity and soil stoichiometric characteristics in a degraded alpine meadow. J. Appl. Ecol. 2019, 30, 127–136. [Google Scholar]
- Le Roux, X.; Poly, F.; Currey, P.; Commeaux, C.; Hai, B.; Nicol, G.W.; Prosser, J.I.; Schloter, M.; Attard, E.; Klumpp, K. Effects of aboveground grazing on coupling among nitrifier activity, abundance and community structure. ISME J. 2008, 2, 221–232. [Google Scholar] [CrossRef] [PubMed]
- Qu, T.B.; Du, W.C.; Yuan, X.; Yang, Z.M.; Liu, D.B.; Wang, D.L.; Yu, L.J. Impacts of grazing intensity and plant community composition on soil bacterial community diversity in a steppe grassland. PLoS ONE 2016, 11, e0159680. [Google Scholar] [CrossRef]
- Xun, W.B.; Yan, R.R.; Ren, Y.; Jin, D.Y.; Xiong, W.; Zhang, G.S.; Cui, Z.L.; Xin, X.P.; Zhang, R.F. Grazing-induced microbiome alterations drive soil organic carbon turnover and productivity in meadow steppe. Microbiome 2018, 6, 170. [Google Scholar] [CrossRef]
- Gao, F.; Wang, B.; Shi, Y.X.; Zhang, G.X.; Wang, J.; Si, G.C.; Han, C.H.; Yuan, Y.L.; Hu, Z. The response of alpine grasslands ecosystem in the north Tibet to short-term enclosure. Acta Ecol. Sin. 2017, 37, 4366–4374. [Google Scholar]
- Wang, Z.; Li, X.L.; Ji, B.M.; Struik, P.C.; Jin, K.; Tang, S.M. Coupling between the responses of plants, soil, and microorganisms following grazing exclusion in an cvergrazed grassland. Front. Plant Sci. 2021, 12, 640789. [Google Scholar] [CrossRef]
- Xun, W.B.; Zhao, J.; Xue, C.; Zhang, G.S.; Ran, W.; Wang, B.R.; Shen, Q.R.; Zhang, R.F. Significant alteration of soil bacterial communities and organic carbon decomposition by different long-term fertilization management conditions of extremely low-productivity arable soil in South China. Environ. Microbiol. 2016, 18, 1907–1917. [Google Scholar] [CrossRef]
- Thomson, B.C.; Ostle, N.; McNamara, N.; Bailey, M.J.; Whiteley, A.S.; Griffiths, R.I. Vegetation affects the relative abundances of dominant soil bacterial taxa and soil respiration rates in an upland grassland soil. Microb. Ecol. 2010, 59, 335–343. [Google Scholar] [CrossRef]
- He, Z.L.; Piceno, Y.; Deng, Y.; Xu, M.Y.; Lu, Z.M.; DeSantis, T.; Andersen, G.; Hobbie, S.E.; Reich, P.B.; Zhou, J.Z. The phylogenetic composition and structure of soil microbial communities shifts in response to elevated carbon dioxide. ISME J. 2011, 6, 259–272. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Li, L.; Ji, L.; Li, Y.; Liu, J.; Li, F. Divergent effects of grazing versus mowing on plant nutrients in typical steppe grasslands of Inner Mongolia. J. Plant Ecol. 2022, 16, rtac032. [Google Scholar] [CrossRef]
- Melo, V.; Barros, L.; Silva, M.; Veloso, T.; Senwo, Z.; Matos, K.; Nunes, T.K.O. Soil bacterial diversities and response to deforestation, land use and burning in North Amazon, Brazil. Appl. Soil Ecol. 2021, 158, 103775. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, J.X.; Yang, X.L.; Huang, X.; Chen, S.L.; Qiao, Y.M. Characteristics of soil microbial diversity in different patches of alpine meadow. Acta Agrestia Sin. 2021, 29, 1916–1926. [Google Scholar]
- Adair, K.L.; Wratten, S.; Lear, G. Soil phosphorus depletion and shifts in plant communities change bacterial community structure in a long-term grassland management trial. Env. Microbiol. Rep. 2013, 5, 404–413. [Google Scholar] [CrossRef] [PubMed]
- John, M.K. Colorimetric determination in soil and plant material with ascorbic acid. Soil Sci. 1970, 109, 214–220. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SOM (g/kg) | TN (g/kg) | TP (g/kg) | TK (g/kg) | AN (mg/kg) | AP (mg/kg) | AK (mg/kg) | |
|---|---|---|---|---|---|---|---|
| CK | 24.62 ± 1.43a | 1.61 ± 0.12a | 0.46 ± 0.02a | 28.93 ± 2.11a | 70.98 ± 6.28a | 4.82 ± 0.59a | 169.22 ± 16.66a |
| LG | 24.70 ± 0.91a | 1.60 ± 0.06a | 0.44 ± 0.01a | 29.21 ± 3.86a | 77.31 ± 19.54a | 3.88 ± 0.93a | 157.33 ± 12.88a |
| MG | 24.49 ± 4.35a | 1.52 ± 0.18a | 0.44 ± 0.02a | 27.46 ± 0.76a | 70.84 ± 14.37a | 3.63 ± 0.58a | 151.97 ± 45.02a |
| HG | 25.70 ± 0.43a | 1.61 ± 0.06a | 0.43 ± 0.00a | 30.09 ± 1.04a | 95.42 ± 13.79a | 4.44 ± 1.28a | 196.72 ± 8.44a |
| SOM | TN | TP | TK | AN | AP | AK | |
|---|---|---|---|---|---|---|---|
| Shannon | −0.652 * | −0.263 | −0.053 | 0.231 | −0.151 | −0.308 | −0.604 * |
| Simpson | −0.433 | −0.012 | 0.117 | 0.162 | −0.222 | −0.328 | −0.495 |
| Pielou | −0.453 | −0.120 | −0.031 | 0.314 | −0.031 | −0.469 | −0.467 |
| Chao1 | −0.141 | 0.010 | 0.297 | −0.245 | −0.186 | 0.454 | −0.068 |
| SOM | TN | TP | TK | AN | AP | AK | |
|---|---|---|---|---|---|---|---|
| r | 0.4353 | 0.2983 | −0.1221 | 0.2819 | −0.0122 | −0.0840 | 0.2790 |
| p | 0.019 | 0.085 | 0.675 | 0.052 | 0.514 | 0.671 | 0.076 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Guo, M.; Li, Y.; Yin, X.; Guo, J.; Wang, J. Responses of Soil Bacterial Communities and Chemical Properties to Grazing Regulation in Desert Steppe. Agronomy 2023, 13, 2817. https://doi.org/10.3390/agronomy13112817
Wang Y, Guo M, Li Y, Yin X, Guo J, Wang J. Responses of Soil Bacterial Communities and Chemical Properties to Grazing Regulation in Desert Steppe. Agronomy. 2023; 13(11):2817. https://doi.org/10.3390/agronomy13112817
Chicago/Turabian StyleWang, Yue, Mishan Guo, Yongfu Li, Xiaolin Yin, Jianying Guo, and Jing Wang. 2023. "Responses of Soil Bacterial Communities and Chemical Properties to Grazing Regulation in Desert Steppe" Agronomy 13, no. 11: 2817. https://doi.org/10.3390/agronomy13112817