
The Microbiomes of Various Types of Abandoned Fallow Soils of South Taiga (Novgorod Region, Russian North-West)
 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
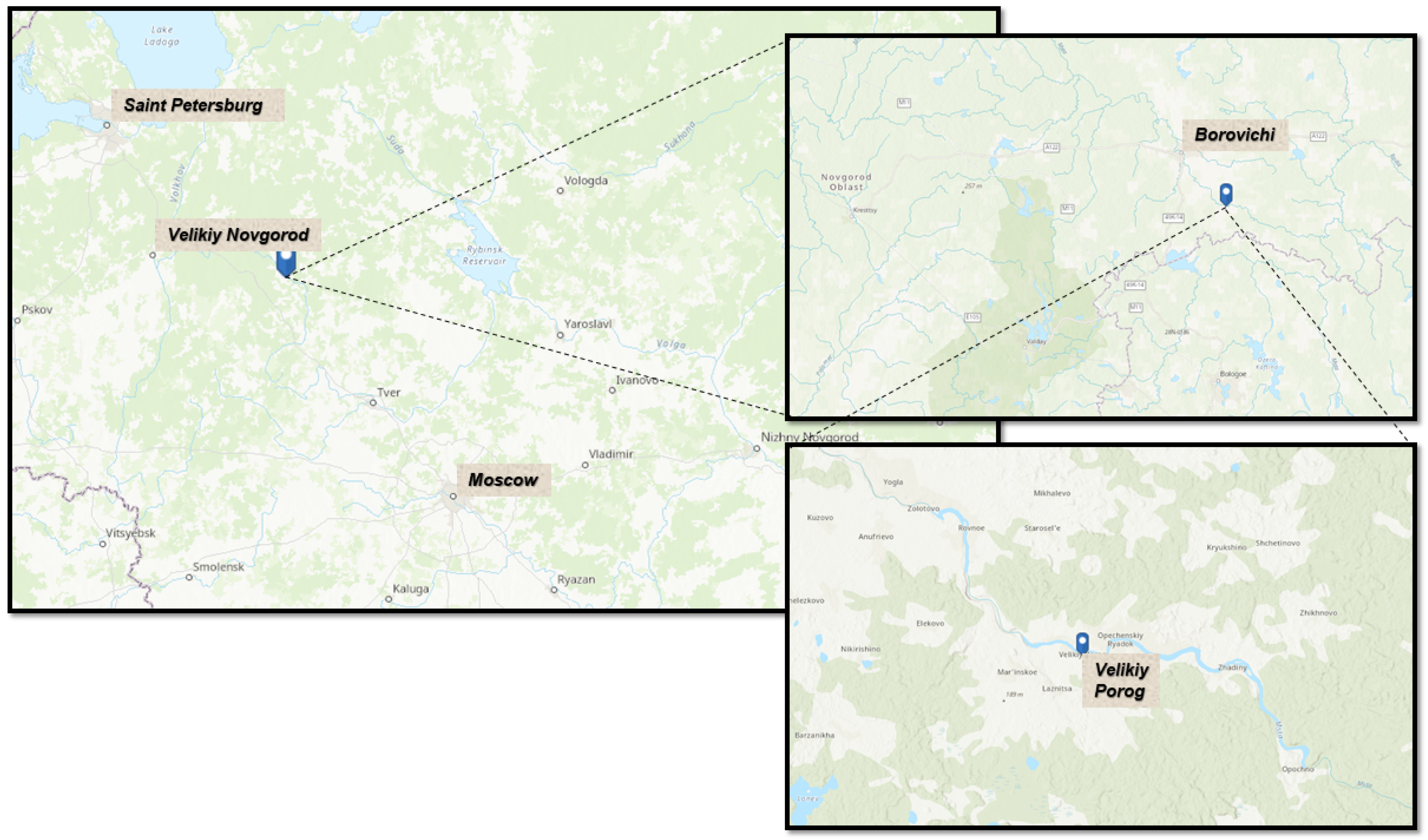
2.1. The Study Sites
2.2. Laboratory Analysis
2.3. Data Processing and Statistical Analysis
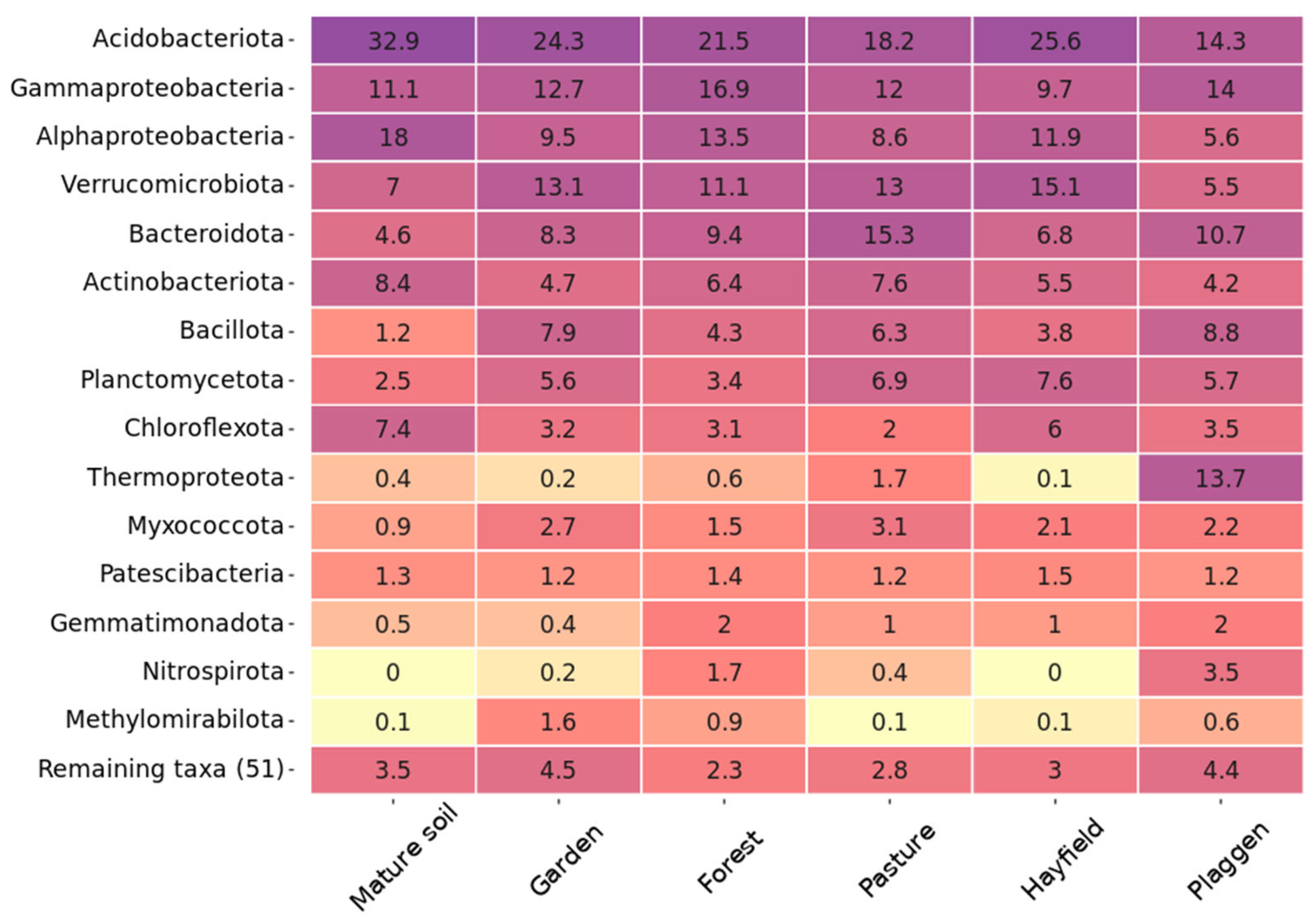
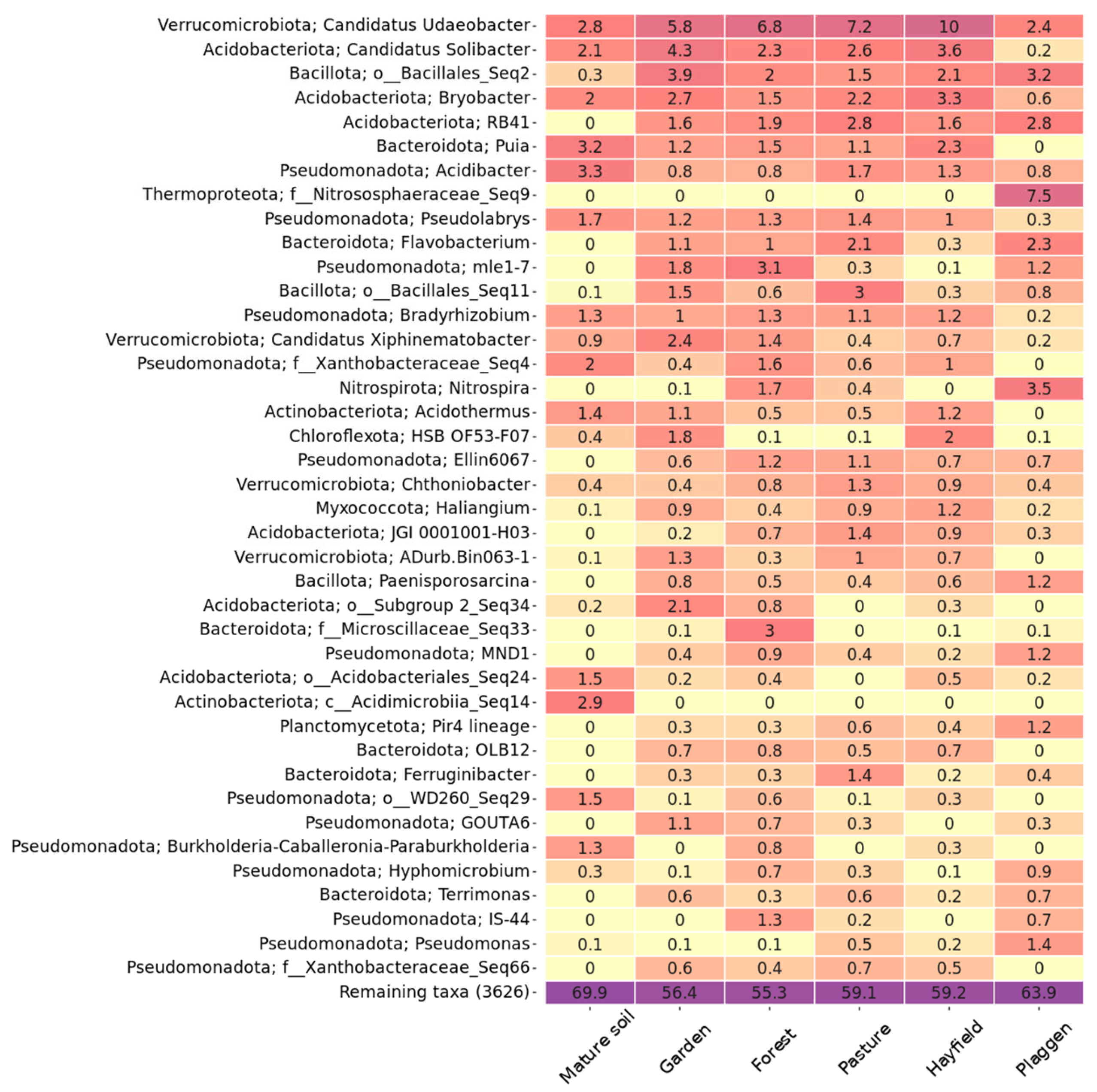
3. Results and Discussion
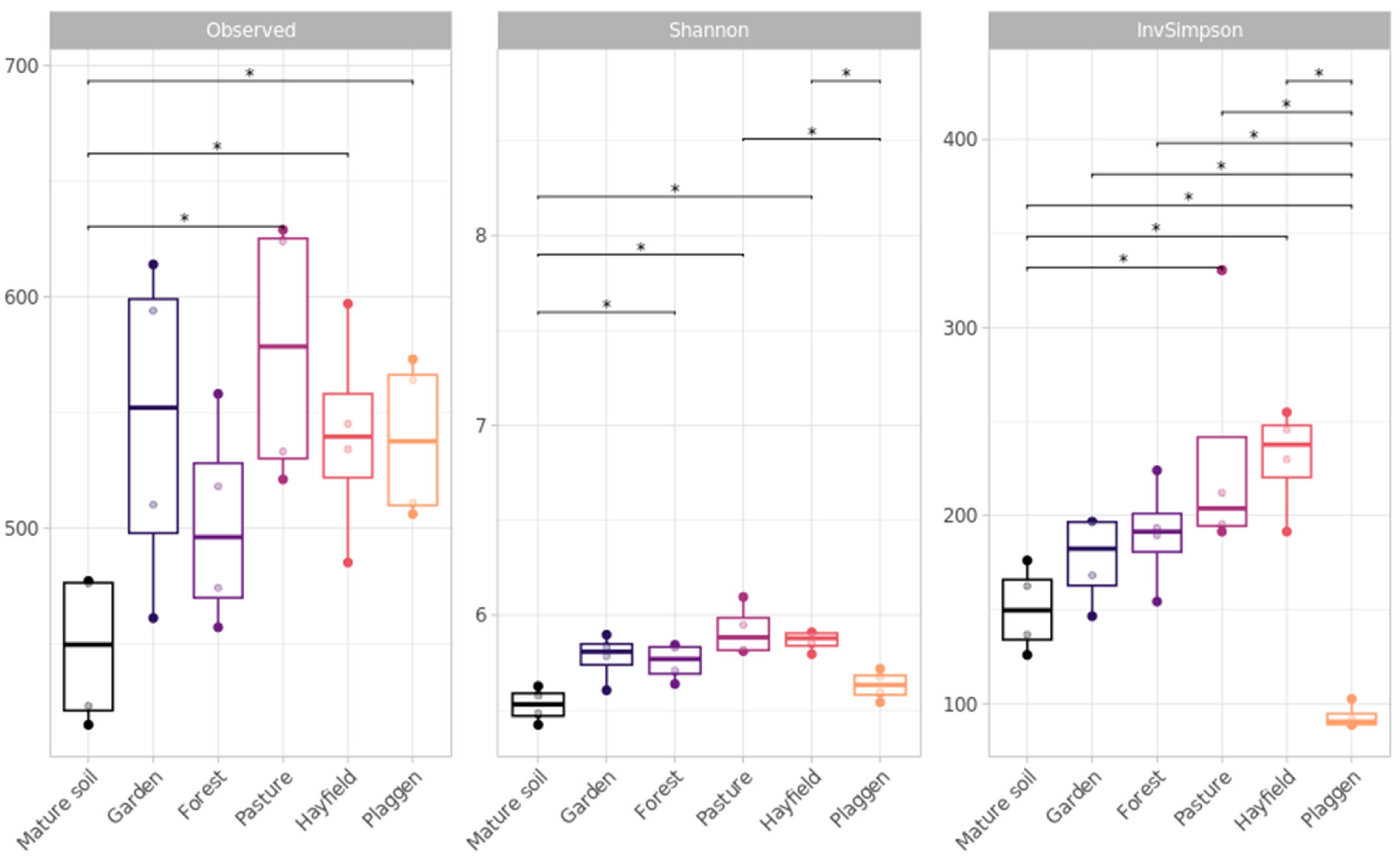
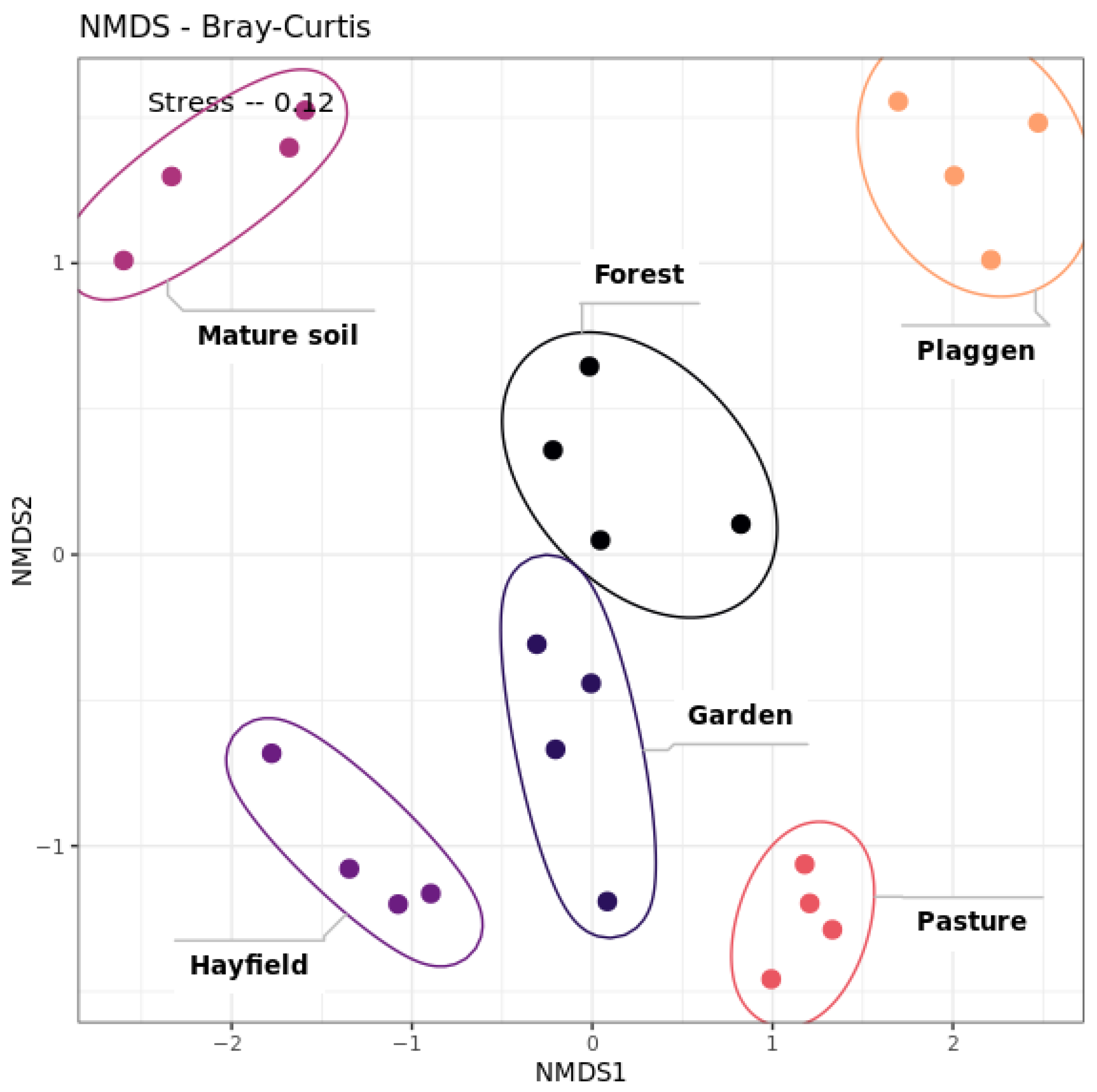
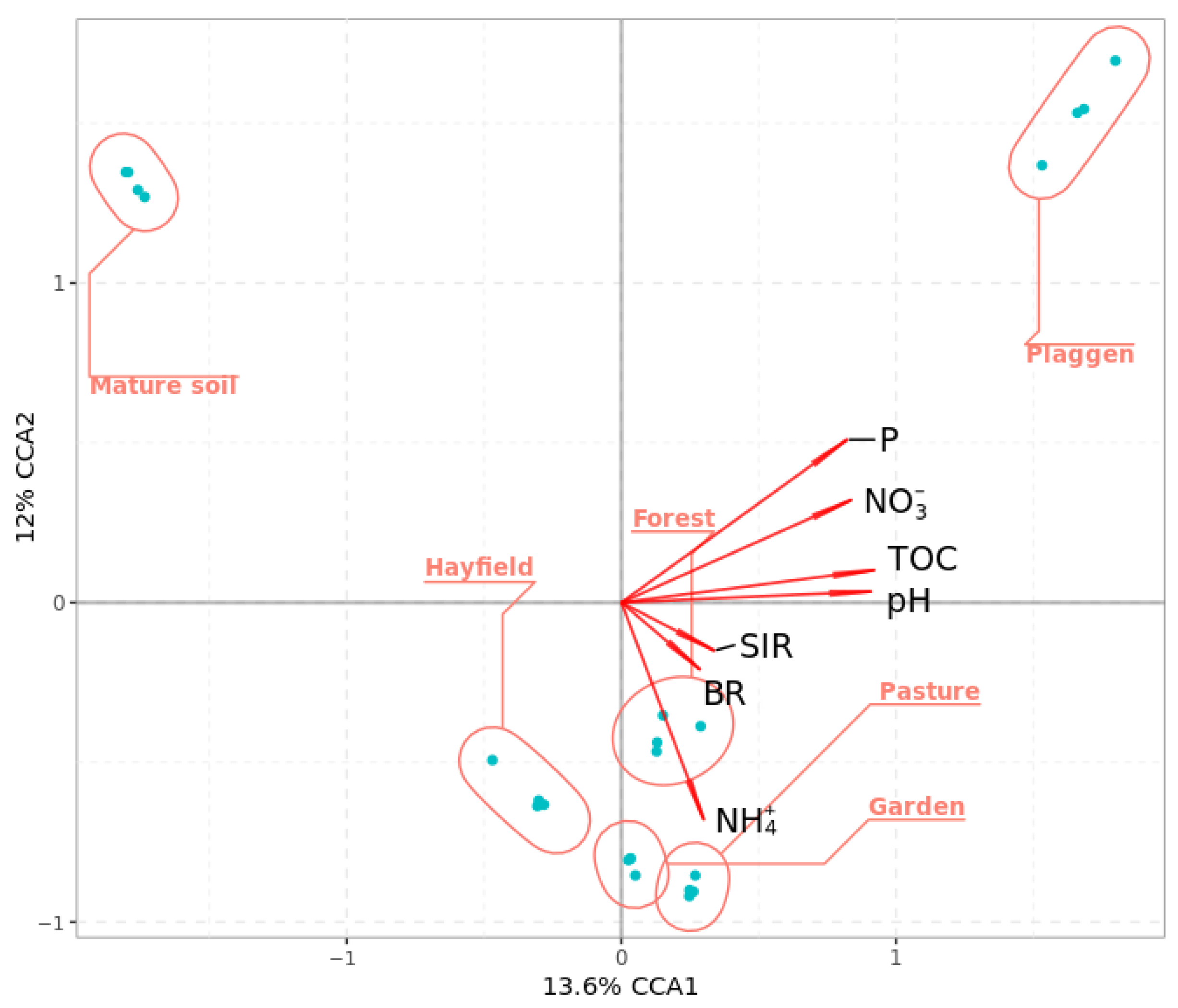
Soil Microbiome
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lyuri, D.I.; Nekrich, A.S.; Karelin, D.V. Cropland dynamics in Russia in 1990–2015 and soil emission of carbon dioxide. Vestn. Mosc. Univ. 2018, 3, 70–76. [Google Scholar]
- Dymov, A.; Dubrovskiy, Y.; Startsev, V. Postagrogenic development of Retisols in the middle taiga subzone of European Russia (Komi Republic). Land Degrad. Dev. 2018, 29, 495–505. [Google Scholar] [CrossRef]
- Orlova, O.I. Struggle for the ground: Restoration of laylands. Karelian Sci. J. 2015, 2, 130–133. [Google Scholar]
- Kechaikina, I.O.; Ryumin, A.G.; Chukov, S.N. Postagrogenic transformation of organic matter in soddy-Podzolic soils. Eurasian Soil Sci. 2011, 44, 1077–1089. [Google Scholar] [CrossRef]
- Iurmanov, A.; Romanov, M.; Marika, G.; Volodina, A.; Baikova, I.; Popov, I.; Markovets, M. Seagrass Zostera in the Russian Section of the Baltic Sea. Geogr. Environ. Sustain. 2022, 15, 111–115. [Google Scholar] [CrossRef]
- Ipatov, V.S.; Lebedeva, V.K. Some aspects of the study of the functional structure of meadow phytocenosys. Izv. Samara Sci. Cent. Russ. Acad. Sci. 2012, 36, 1252–1256. [Google Scholar]
- Frouz, J.; Nováková, A. Development of soil microbial properties in topsoil layer during spontaneous succession in heaps after brown coal mining in relation to humus microstructure development. Geoderma 2005, 129, 54–64. [Google Scholar] [CrossRef]
- Pershina, E.; Ivanova, E.; Korvigo, I.; Chirak, E.; Sergaliev, N.; Abakumov, E.; Provorov, N.; Andronov, E. Investigation of the core microbiome in main soil types from the East European plain. Sci. Total Environ. 2018, 631–632, 1421–1430. [Google Scholar] [CrossRef]
- Kaiser, K.; Wemheuer, B.; Korolkow, V.; Wemheuer, F.; Nacke, H.; Schöning, I. Driving forces of soil bacterial community structure, diversity, and function in temperate grasslands and forests. Sci. Rep. 2016, 6, 33696. [Google Scholar] [CrossRef]
- Zverev, A.O.; Kichko, A.A.; Pinaev, A.G.; Provorov, N.A.; Andronov, E.E. Diversity Indices of Plant Communities and Their Rhizosphere Microbiomes: An Attempt to Find the Connection. Microorganisms 2021, 9, 2339. [Google Scholar] [CrossRef]
- Semenov, M.V.; Ksenofontova, N.A.; Nikitin, D.A.; Tkhakakhova, A.K.; Lukin, S.M. Microbiological Parameters of Soddy-Podzolic Soil and Its Rhizosphere in a Half-Century Field Experiment with Different Fertilizer Systems. Eurasian Soil Sci. 2023, 56, 756–768. [Google Scholar] [CrossRef]
- Malakhovsky, D.B.; Markov, K.K. Geomorphology and Quaternary Deposits of the North-West of the European Part of the USSR; Nauka: Leningrad, Russia, 1969; p. 256. [Google Scholar]
- FAO. Standard Operating Procedure for Soil pH Determination; FAO: Rome, Italy, 2021. [Google Scholar]
- FAO. Standard Operating Procedure for Soil Organic Carbon: Tyurin Spectrophotometric Method; FAO: Rome, Italy, 2021. [Google Scholar]
- FAO. Standard Operating Procedure for Soil Available Phosphorus—Olsen Method; FAO: Rome, Italy, 2021. [Google Scholar]
- GOST R 54650–2011; Soils: Determination of Mobile Phosphorus and Potassium Compounds by Kirsanov Method Modified by ClNAO. Standartinform: Moscow, Russia, 2019; p. 8.
- Caporaso, J.; Lauber, C.; Walters, W.; Berg-Lyons, D.; Lozupone, C.; Turnbaugh, P.; Fierer, N.; Knight, R. Global Patterns of 16S RRNA Diversity at a Depth of Millions of Sequences per Sample. Proc. Natl. Acad. Sci. USA 2011, 108 (Suppl. S1), 4516–4522. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Callahan, B.; Mcmurdie, P.; Rosen, M.; Han, A.; Johnson, A.J.; Holmes, S. DADA2: High-Resolution Sample Inference from Illumina Amplicon Data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed]
- Mcmurdie, P.; Holmes, S. Phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [PubMed]
- Gladkov, G.; Kimeklis, A.; Zverev, A.; Pershina, E.; Ivanova, E.; Kichko, A.; Andronov, E.; Abakumov, E. Soil Microbiome of the Postmining Areas in Polar Ecosystems in Surroundings of Nadym, Western Siberia, Russia. Open Agric. 2019, 4, 684–696. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F. The SILVA Ribosomal RNA Gene Database Project: Improved Data Processing and Web-Based Tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Parte, A.C.; Sardà Carbasse, J.; Meier-Kolthoff, J.P.; Reimer, L.C.; Göker, M. List of Prokaryotic Names with Standing in Nomenclature (LPSN) Moves to the DSMZ. Int. J. Syst. Evol. Microbiol. 2020, 70, 5607–5612. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Vegan: Community Ecology Package. R Package Version 2.4-3. 2017. Available online: https://cran.r-project.org/package=vegan (accessed on 9 December 2022).
- Anderson, M.J. Permutational Multivariate Analysis of Variance (PERMANOVA). In Wiley StatsRef: Statistics Reference Online; Balakrishnan, N., Colton, T., Everitt, B., Piegorsch, W., Ruggeri, F., Teugels, J.L., Eds.; Wiley: New York, NY, USA, 2017; pp. 1–15. [Google Scholar]
- Litvinovich, A.V. Postagrogenic Evolution of Well Cultivated Soddy-Podzolic Soils in the Northwestern Nonchernozemic Zone. Agrochemistry 2009, 7, 85–93. [Google Scholar]
- Bujalský, L.; Kaneda, S.; Dvorščík, P.; Frouz, J. In situ soil respiration at reclaimed and unreclaimed post-mining sites: Responses to temperature and reclamation treatment. Ecol. Eng. 2014, 68, 53–59. [Google Scholar] [CrossRef]
- Bogdevitch, I.M.; Putyatin, Y.V.; Stanilevitch, I.S.; Lomonos, O.L. Dynamic of potassium supply level of arable and meadow soils of Belarus. Soils Sci. Agrochem. 2020, 62, 104–116. [Google Scholar]
- Suleymanov, A.; Nizamutdinov, T.; Morgun, E. Assessing and geostatistical mapping of metal contamination in the polar arable plot (Yamal-Nenets Autonomous District, Russia). Int. J. Environ. Sci. Technol. 2023, 1–10. [Google Scholar] [CrossRef]
- Gamzikov, G.P.; Suleimenov, S.Z. Nitrogen Mineralization Capacity of Gray Forest Soil of the Novosibirsk Ob River Region during Composting and Fallowing of Plant Residues. Eurasian Soil Sci. 2021, 54, 729–737. [Google Scholar] [CrossRef]
- Ponomareva, V. Theory of Podzol Formation; Nauka: Moscow, Russia, 1964. [Google Scholar]
- Zheng, M.; Zhu, P.; Zheng, J.; Xue, L.; Zhu, Q.; Cai, X.; Cheng, S.; Zhang, Z.; Kong, F.; Zhang, J. Effects of soil texture and nitrogen fertilisation on soil bacterial community structure and nitrogen uptake in flue-cured tobacco. Sci. Rep. 2021, 11, 22643. [Google Scholar] [CrossRef] [PubMed]
- Seaton, F.; George, P.; Lebron, I.; Jones, L.; Creer, S.; Robinson, D. Soil textural heterogeneity impacts bacterial but not fungal diversity. Soil Biol. Biochem. 2020, 144, 107766. [Google Scholar] [CrossRef]
- Korneykova, M.V.; Vasenev, V.I.; Nikitin, D.A.; Soshina, A.S.; Dolgikh, A.V.; Sotnikova, Y.L. Urbanization Affects Soil Microbiome Profile Distribution in the Russian Arctic Region. Int. J. Environ. Res. Public Health 2021, 18, 11665. [Google Scholar] [CrossRef]
- Gelashvili, D.B.; Iudin, D.I.; Rozenberg, G.S.; Snegireva, M.S.; Solntsev, L.A.; Fedjunin, V.A.; Yakimov, V.N. Fractal characteristics of the species structure of ichneumon wasp communities in the middle urals. Dokl. Biol. Sci. 2010, 434, 351–354. [Google Scholar] [CrossRef]
- Lupwayi, N.; Clayton, G.; O’Donovan, J.; Harker, K.; Turkington, T.; Rice, W. Soil microbiological properties during decomposition of crop residues under conventional and zero tillage. Can. J. Soil Sci. 2004, 84, 411–419. [Google Scholar] [CrossRef]
- Choma, M.; Šamonil, P.; Kaštovská, E.; Bárta, J.; Tahovská, K.; Valtera, M.; Šantrůčková, H. Soil Microbiome Composition along the Natural Norway Spruce Forest Life Cycle. Forests 2021, 12, 410. [Google Scholar] [CrossRef]
- Naliukhin, A.N.; Khamitova, S.M.; Glinushkin, A.P.; Avdeev, Y.M.; Snetilova, V.S.; Laktionov, Y.V.; Surov, V.V.; Siluyanova, O.V.; Belozerov, D.A. Changes in the Metagenome of Prokaryotic Community as an Indicator of Fertility of Arable Soddy-Podzolic Soils upon Fertilizer Application. Eurasian Soil Sci. 2018, 51, 321–326. [Google Scholar] [CrossRef]
- Wang, L.; Ye, X.; Hu, H.; Du, J.; Xi, Y.; Shen, Z.; Lin, J.; Chen, D. Soil bacterial communities triggered by organic matter inputs associates with a high-yielding pear production. Soil 2022, 8, 337–348. [Google Scholar] [CrossRef]
- Saubenova, M.; Oleinikova, Y.; Sadanov, A.; Yermekbay, A.; Bokenov, D.; Shorabaev, Y. The input of microorganisms to the cultivation of mushrooms on lignocellulosic waste. AIMS Agric. Food 2023, 8, 239–277. [Google Scholar] [CrossRef]
- Nelkner, J.; Huang, L.; Lin, T.W.; Schulz, A.; Osterholz, B.; Henke, C.; Blom, J.; Pühler, A.; Sczyrba, A.; Schlüter, A. Abundance, classification and genetic potential of Thaumarchaeota in metagenomes of European agricultural soils: A meta-analysis. Environ. Microbiome 2023, 18, 26. [Google Scholar] [CrossRef] [PubMed]
- Colette, M.; Guentas, L.; Patrona, L.D.; Ansquer, D.; Callac, N. Dynamic of active microbial diversity in rhizosphere sediments of halophytes used for bioremediation of earthen shrimp ponds. Environ. Microbiome 2023, 18, 58. [Google Scholar] [CrossRef]
- Tian, Z.; Li, G.; Tang, W.; Zhu, Q.; Li, X.; Du, C.; Li, C.; Li, J.; Zhao, C.; Zhang, L. Role of Sedum alfredii and soil microbes in the remediation of ultra-high content heavy metals contaminated soil. Agric. Ecosyst. Environ. 2022, 339, 108090. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample ID | pH | TOC, % | BR | SIR | N-NH4+ | N-NO3− | p | K |
|---|---|---|---|---|---|---|---|---|
| µg CO2−C/g × hour | Exchangeable forms, mg kg−1 | |||||||
| Mature Soil | 5.73 | 0.83 ± 0.06 | 0.82 ± 0.05 | 2.88 ± 0.08 | 4.08 ± 0.24 | 0.27 ± 0.09 | 25 ± 5 | 10 ± 2 |
| Garden | 6.11 | 1.40 ± 0.06 | 0.50 ± 0.09 | 0.87 ± 0.03 | 6.49 ± 0.13 | 0.49 ± 0.05 | 59 ± 4 | 13 ± 3 |
| Forest | 6.67 | 2.55 ± 0.05 | 2.03 ± 0.05 | 7.31 ± 0.22 | 11.78 ± 0.22 | 1.06 ± 0.08 | 70 ± 3 | 69 ± 4 |
| Pasture | 6.37 | 2.42 ± 0.09 | 1.60 ± 0.15 | 8.17 ± 0.72 | 11.52 ± 0.24 | 2.31 ± 0.09 | 161 ± 7 | 195 ± 6 |
| Hayfield | 6.23 | 2.21 ± 0.07 | 1.45 ± 0.12 | 3.48 ± 0.09 | 7.91 ± 0.31 | 0.70 ± 0.06 | 93 ± 6 | 93 ± 8 |
| Plaggen | 6.87 | 3.77 ± 0.03 | 1.27 ± 0.02 | 4.93 ± 0.25 | 5.92 ± 0.16 | 3.61 ± 0.14 | 499 ± 12 | 505 ± 19 |
| Sample ID | 1–0.25 | 0.25–0.05 | 0.05–0.01 | 0.01–0.005 | 0.005–0.001 | <0.001 | <0.01 |
|---|---|---|---|---|---|---|---|
| Mature soil | 50 | 38 | 3 | 1 | 1 | 7 | 9 |
| Garden | 45 | 30 | 16 | 1 | 1 | 7 | 9 |
| Forest | 29 | 48 | 11 | 1 | 4 | 7 | 12 |
| Pasture | 40 | 46 | 4 | 3 | 4 | 3 | 10 |
| Hayfield | 43 | 42 | 6 | 1 | 3 | 5 | 9 |
| Plaggen | 55 | 28 | 5 | 2 | 4 | 6 | 12 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abakumov, E.V.; Gladkov, G.V.; Kimeklis, A.K.; Andronov, E.E. The Microbiomes of Various Types of Abandoned Fallow Soils of South Taiga (Novgorod Region, Russian North-West). Agronomy 2023, 13, 2592. https://doi.org/10.3390/agronomy13102592
Abakumov EV, Gladkov GV, Kimeklis AK, Andronov EE. The Microbiomes of Various Types of Abandoned Fallow Soils of South Taiga (Novgorod Region, Russian North-West). Agronomy. 2023; 13(10):2592. https://doi.org/10.3390/agronomy13102592
Chicago/Turabian StyleAbakumov, Evgeny V., Grigory V. Gladkov, Anastasiia K. Kimeklis, and Evgeny E. Andronov. 2023. "The Microbiomes of Various Types of Abandoned Fallow Soils of South Taiga (Novgorod Region, Russian North-West)" Agronomy 13, no. 10: 2592. https://doi.org/10.3390/agronomy13102592