
Influence of Microalgae Planktochlorella nurekis Clones on Seed Germination

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
- Brassica oleracea L.—cultivar: Cezar (W. Legutko, Przedsiębiorstwo Handlowo-Nasienne Sp. z o.o.)
- Lactuca sativa L.—cultivar: May King—(W. Legutko, Przedsiębiorstwo Handlowo-Nasienne Sp. z o.o.)
- Triticum aestivum L.—spring wheat, cultivar: Serenada, (Hodowla Roślin Strzelce)
- Raphanis sativus L.—cultivar: Mino Early (W. Legutko, Przdsiębiorstwo Handlowo-Nasienne Sp. z o.o.)
2.2. Preparation of Seaweed Extract
2.3. Germination Test
- Germination energy (%GE) [28] after 3 days: %GE = (number of germinated seeds/total number of seeds) × 100
- Growth parameters—length of root and leaves (cm)
- Fresh plant weight (g).
2.4. Statistical Analysis
3. Results
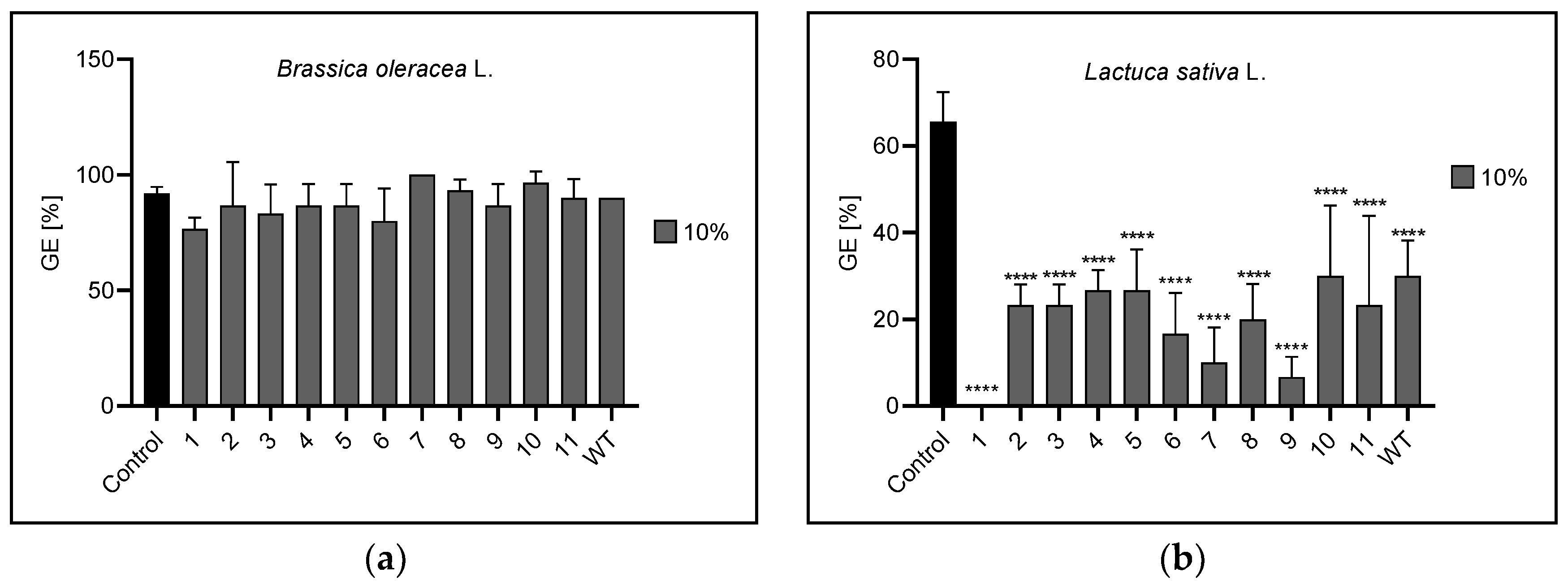
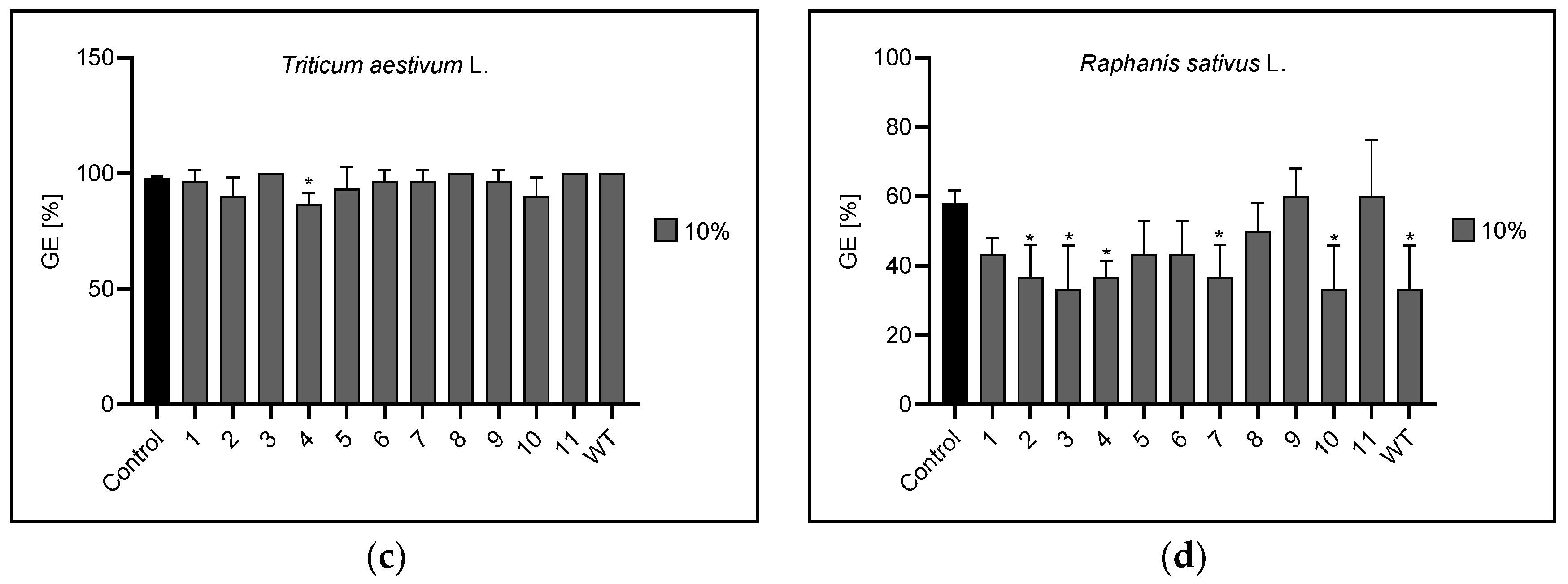
3.1. Germination Energy
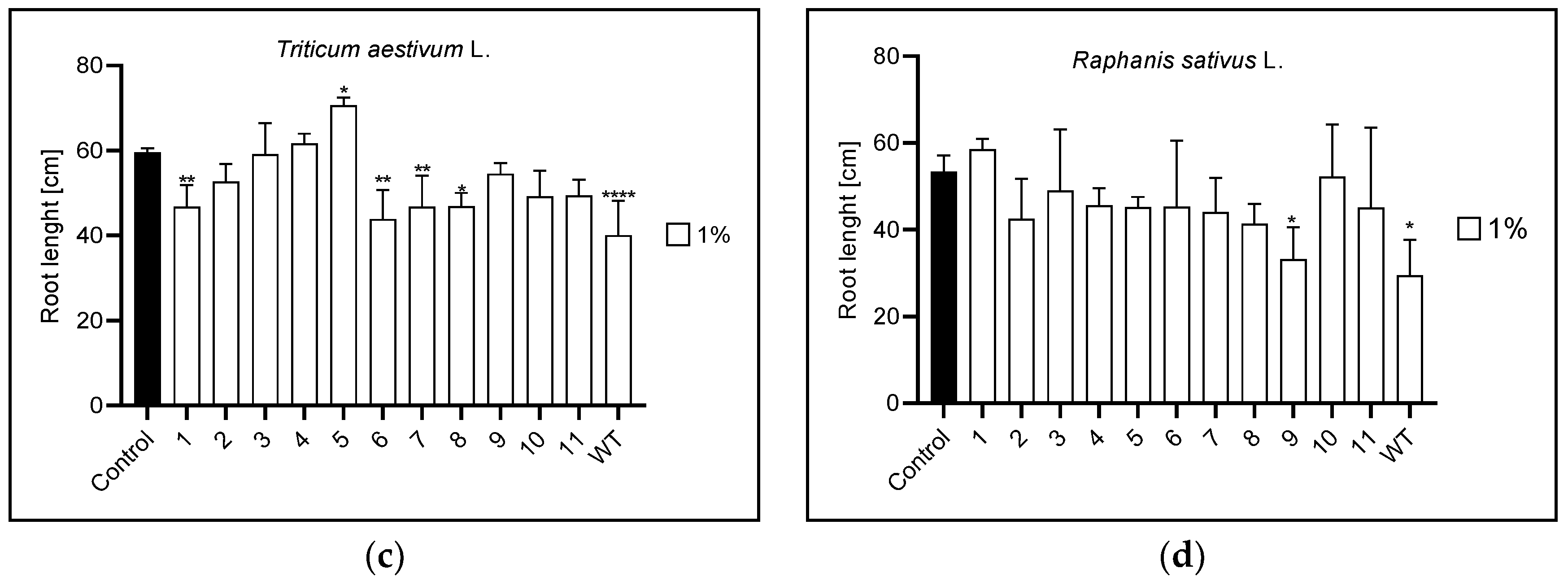
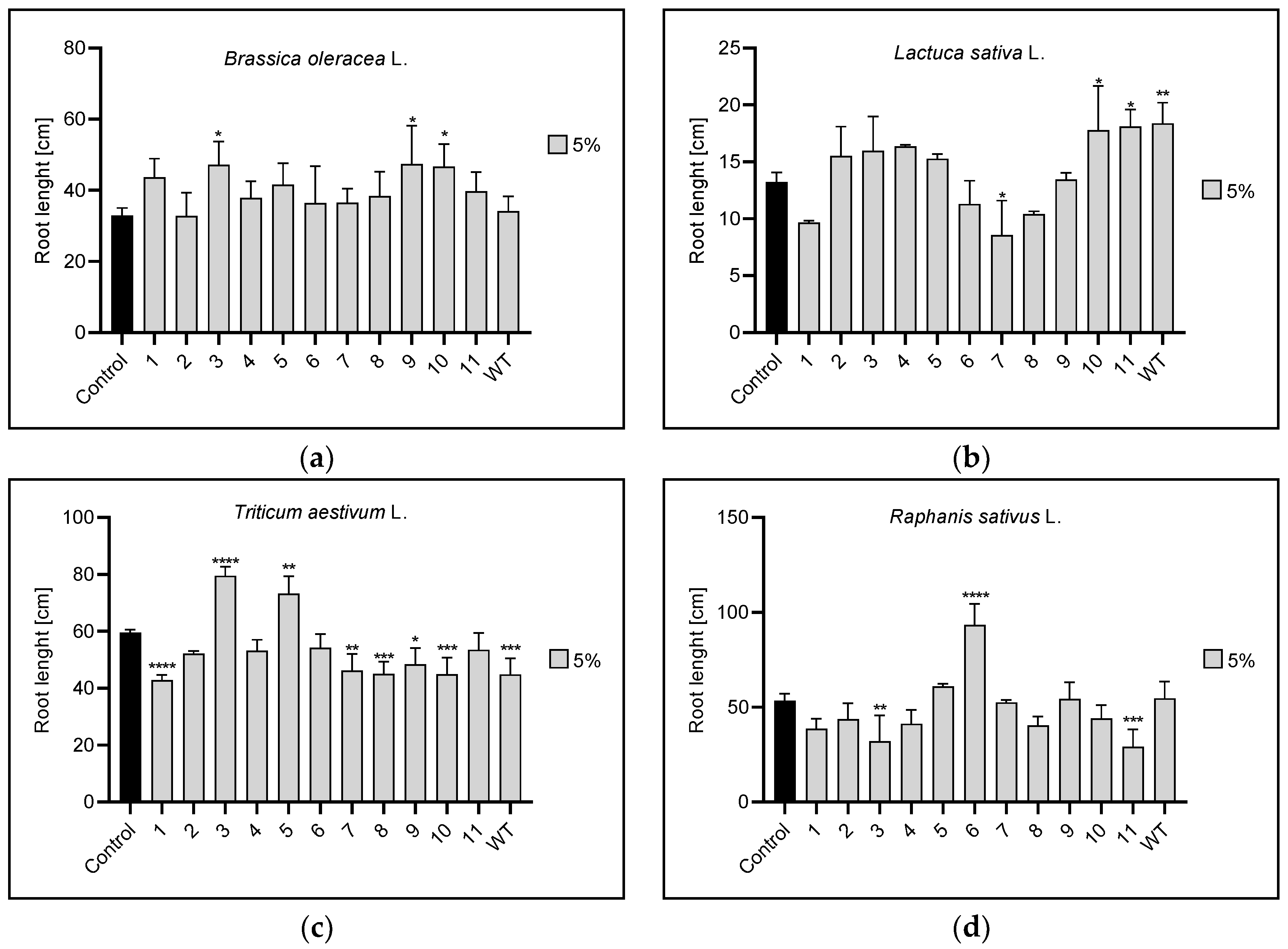
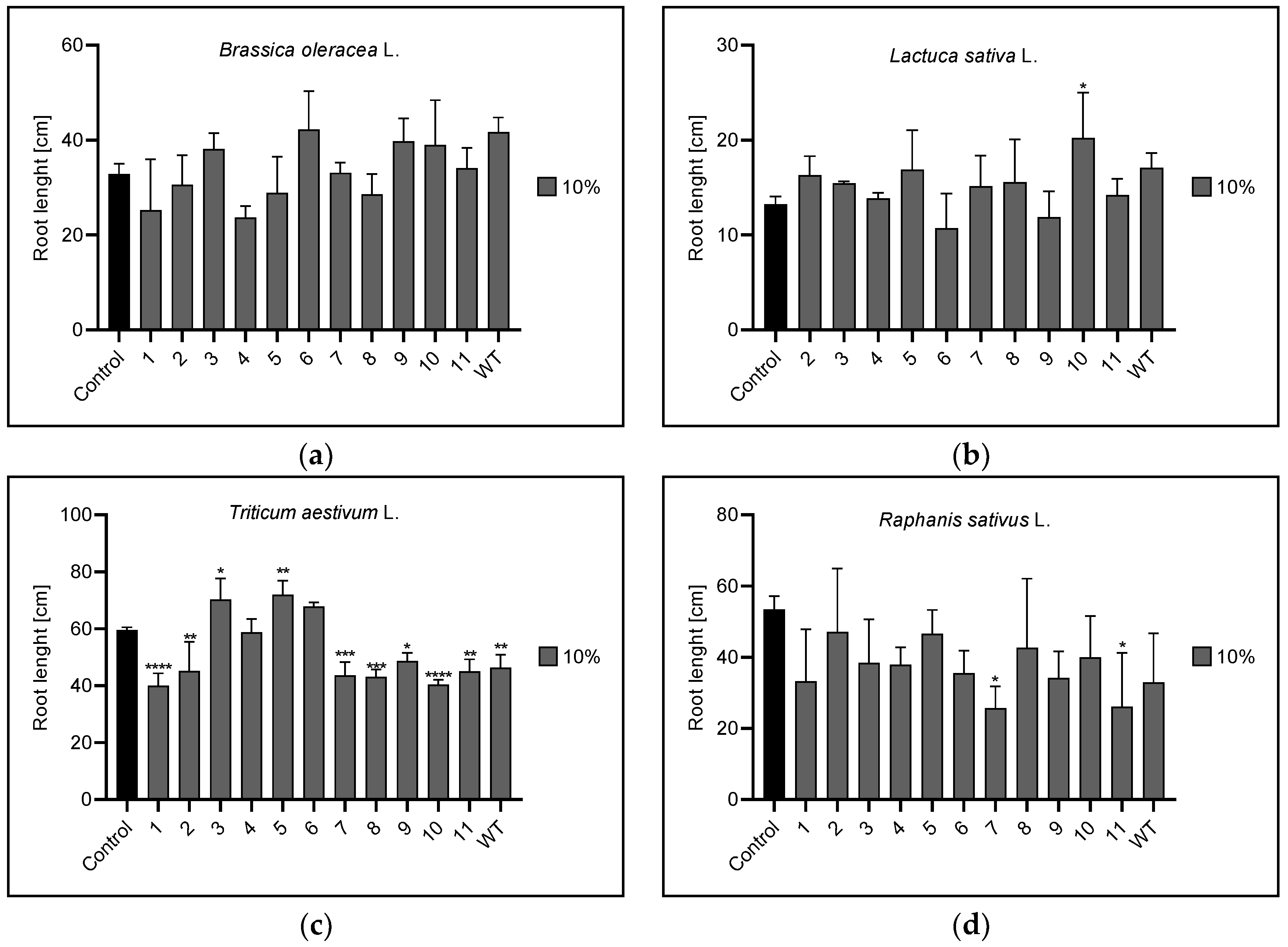
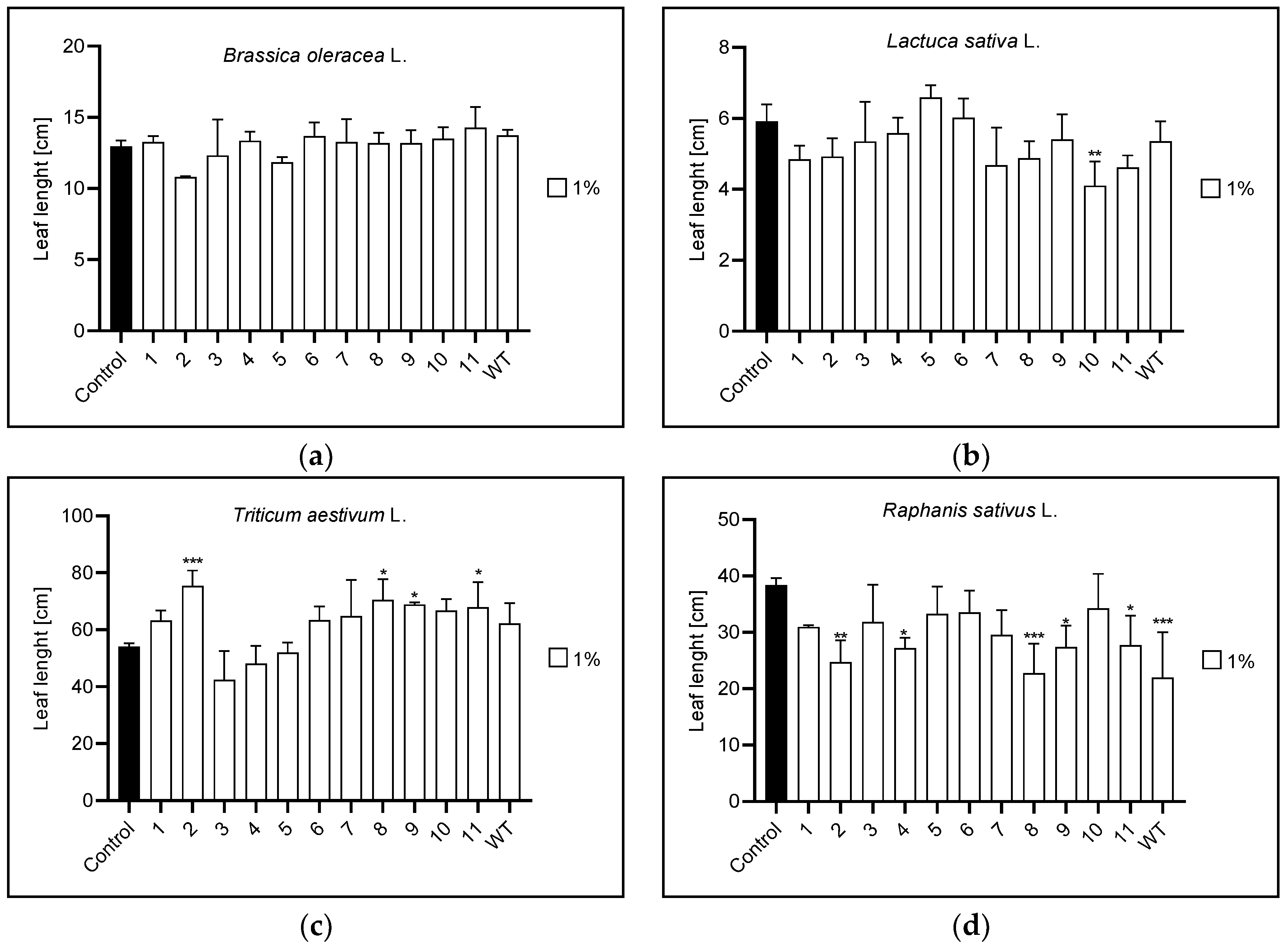
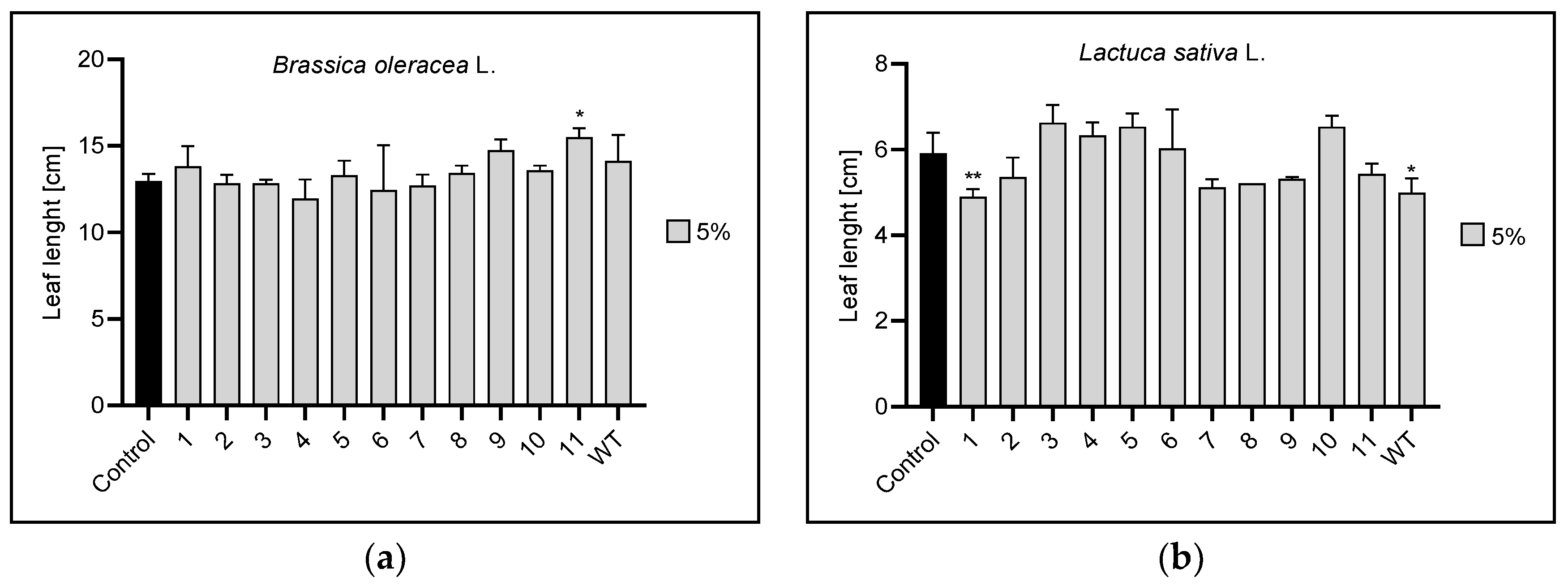
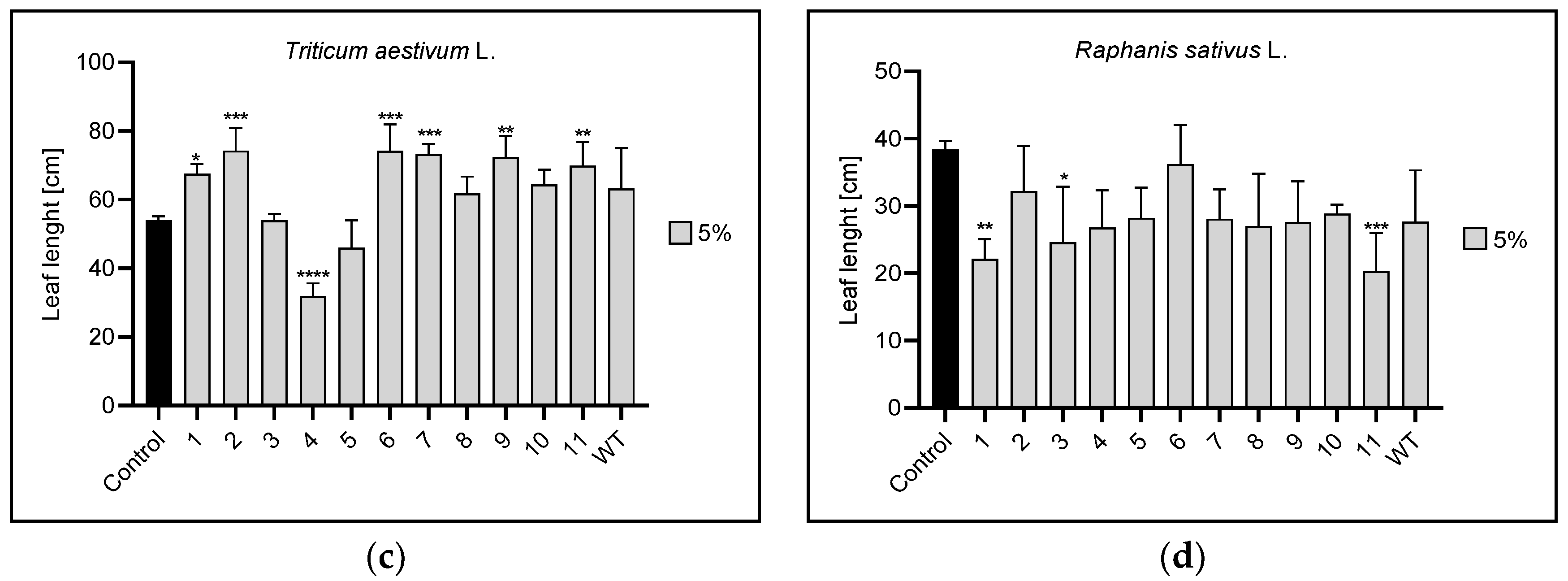
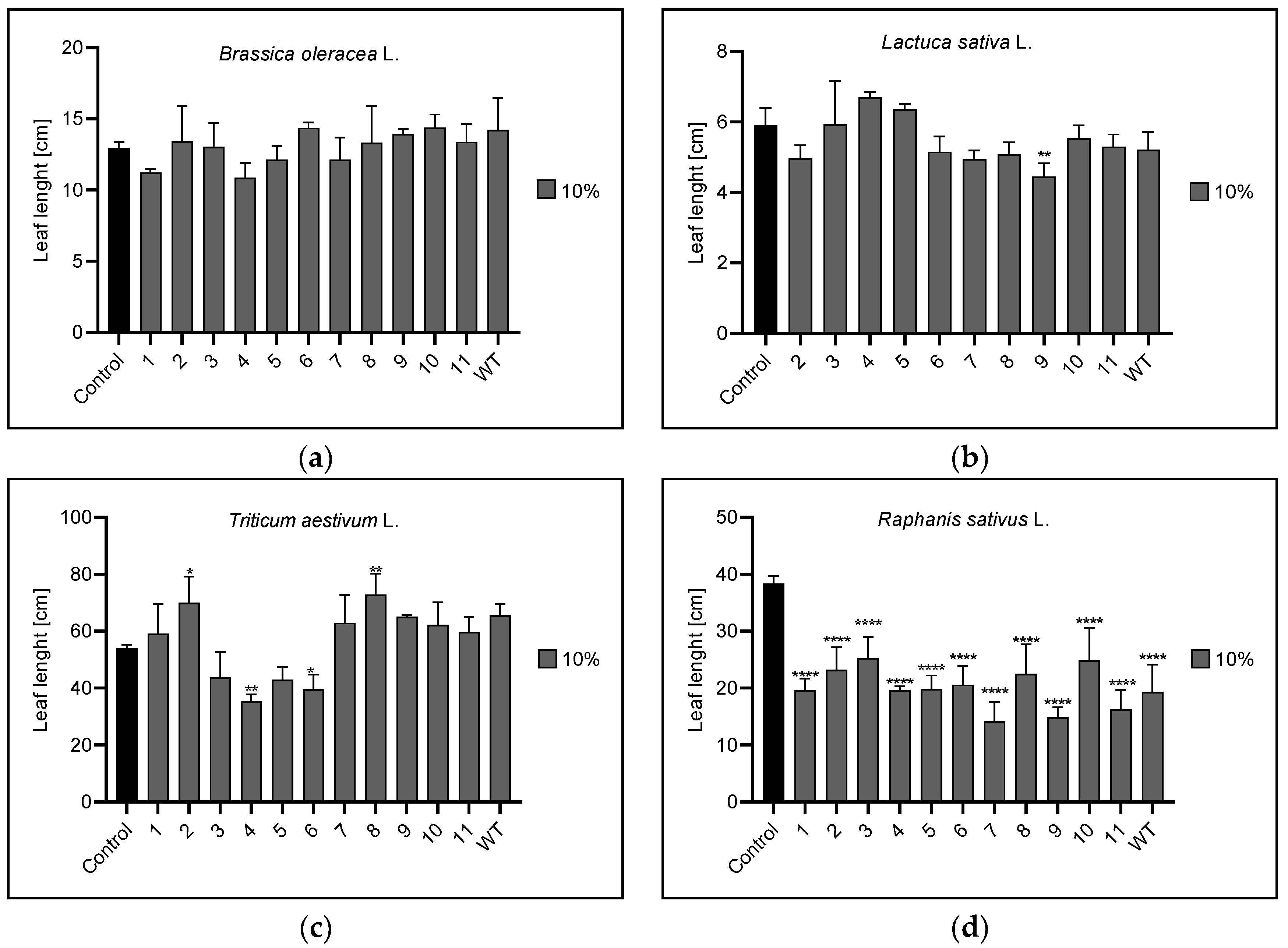
3.2. Root and Leaf Length
3.2.1. Root Length
3.2.2. Leaf Length
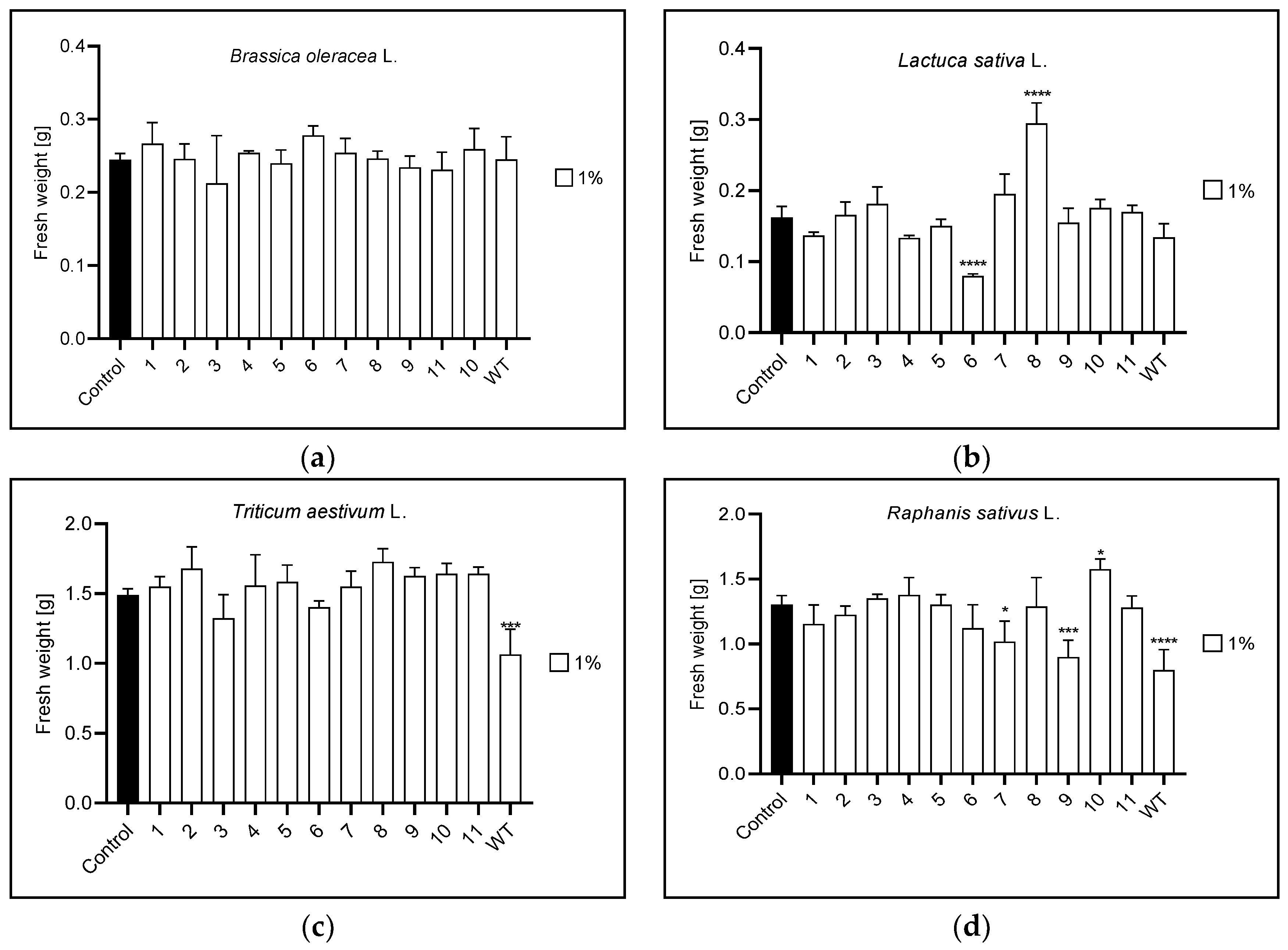
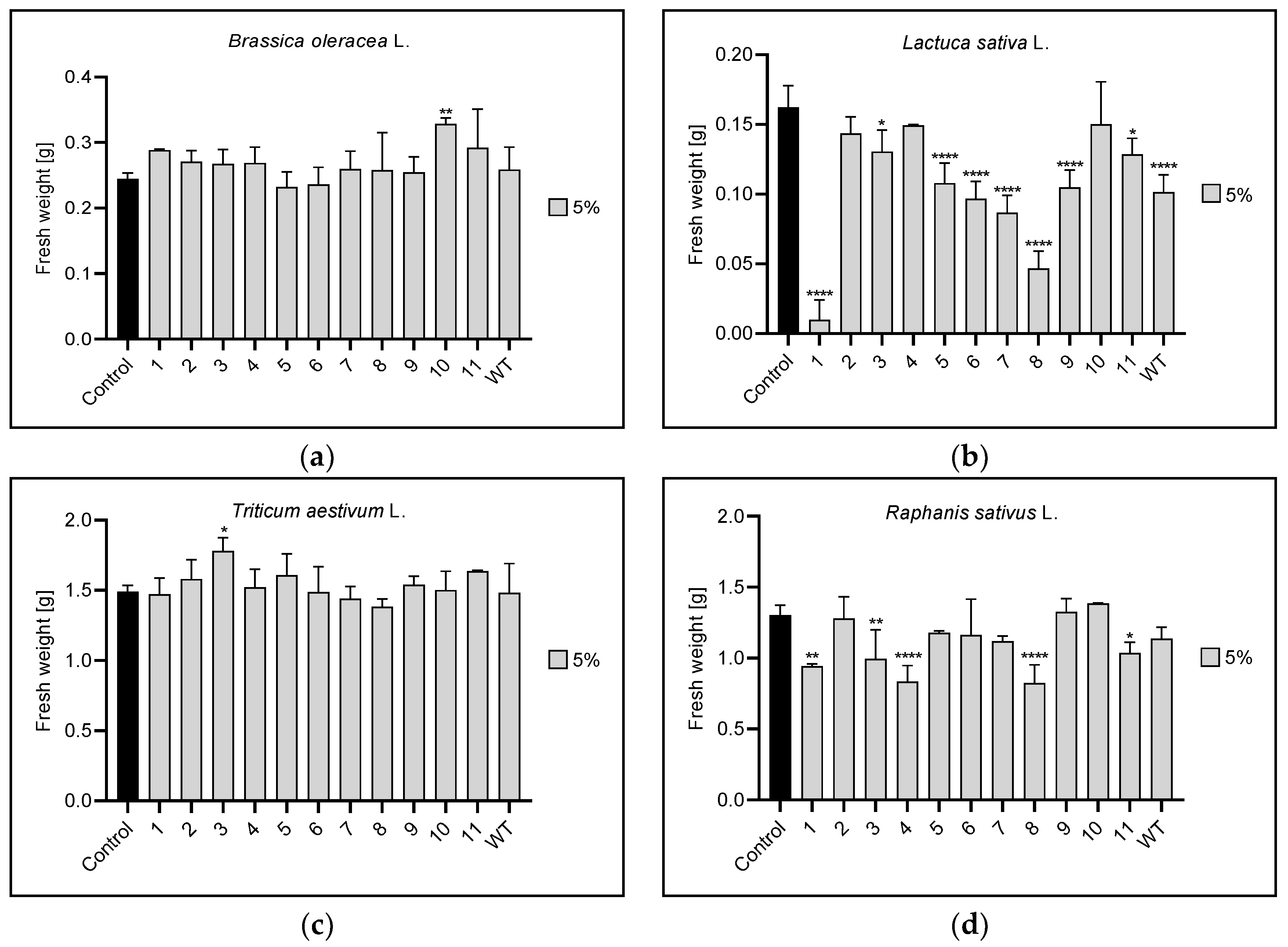
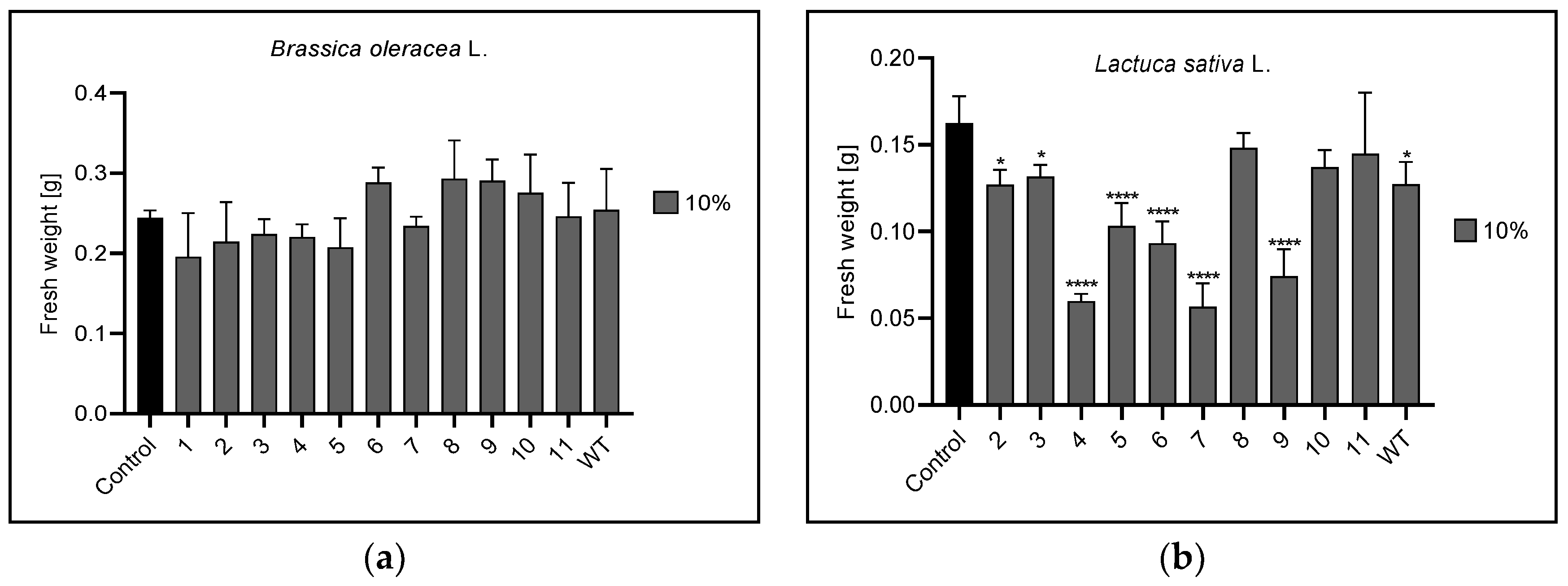
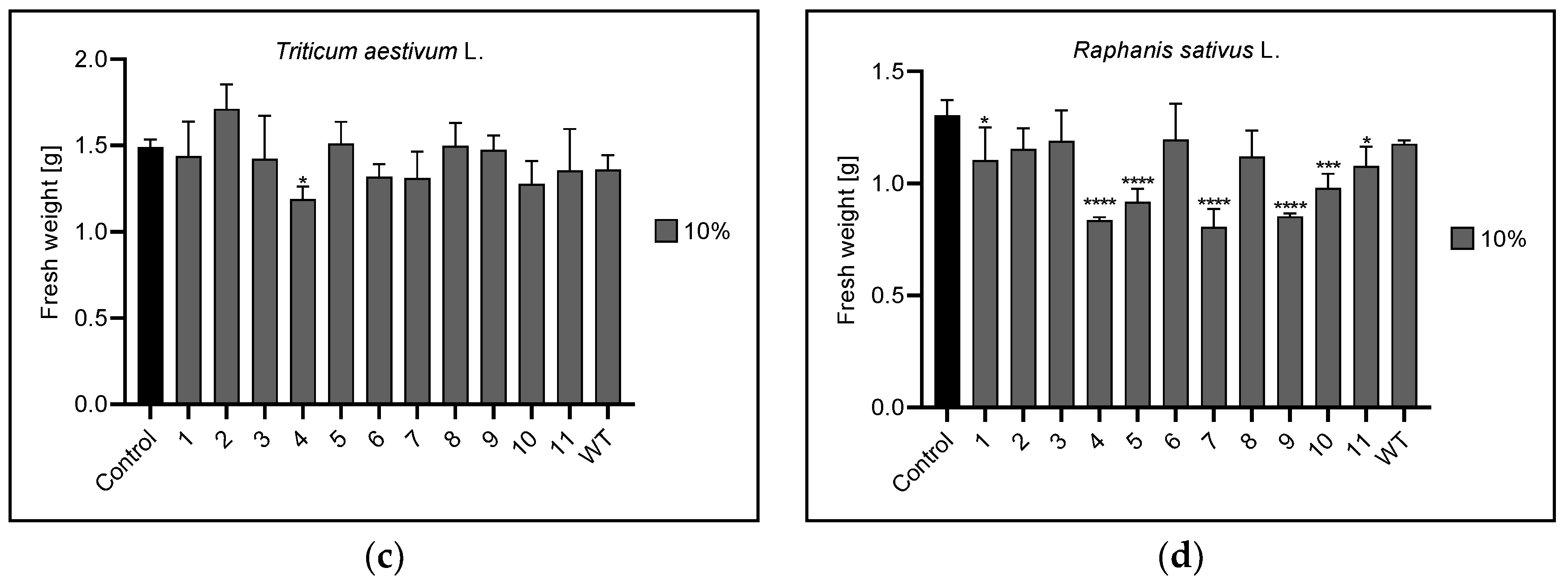
3.3. Fresh Plant Weight
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- FAO. Global Report on Food Crises. Available online: https://www.fao.org/3/cb9997en/cb9997en.pdf (accessed on 11 December 2022).
- FAO. The International Code of Conduct for the Sustainable Use and Management of Fertilizers. Available online: https://www.fao.org/3/ca5253en/ca5253en.pdf (accessed on 7 December 2022).
- Khan, W.; Rayirath, U.P.; Subramanian, S.; Jithesh, M.N.; Rayorath, P.; Hodges, D.M.; Critchley, A.T.; Craigie, J.S.; Norrie, J.; Prithiviraj, B. Seaweed Extracts as Biostimulants of Plant Growth and Development. J. Plant Growth Regul. 2009, 28, 386–399. [Google Scholar] [CrossRef]
- Tang, D.Y.Y.; Khoo, K.S.; Chew, K.W.; Tao, Y.; Ho, S.H.; Show, P.L. Potential utilization of bioproducts from microalgae for the quality enhancement of natural products. Bioresour. Technol. 2020, 304, 122997. [Google Scholar] [CrossRef] [PubMed]
- Górka, B.; Korzeniowska, K.; Lipok, J.; Wieczorek, P.P. The Biomass of Algae and Algal Extracts in Agricultural Production | SpringerLink. In Developments in Applied Phycology; SpringerLink: Berlin/Heidelberg, Germany, 2018; pp. 103–114. [Google Scholar]
- Mobin, S.; Alam, F. Some Promising Microalgal Species for Commercial Applications: A review. Energy Procedia 2017, 110, 510–517. [Google Scholar] [CrossRef]
- Škaloud, P.; Němcová, Y.; Pytela, J.; Bogdanov, I.N.; Bock, C.; Pickinpaugh, H.S. Planktochlorella nurekis gen. et sp. nov. (Trebouxiophyceae, Chlorophyta), a novel coccoid green alga carrying significant biotechnological potential. Fottea 2014, 14, 53–62. [Google Scholar] [CrossRef] [Green Version]
- Moore, T.C. Biochemistry and Physiology of Plant Hormones; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- vander Knaap, E.; Kim, J.H.; Kende, H. A Novel Gibberellin-Induced Gene from Rice and Its Potential Regulatory Role in Stem Growth. Plant Physiol. 2000, 122, 695–704. [Google Scholar] [CrossRef] [Green Version]
- Popko, J.; Hänsch, R.; Mendel, R.R.; Polle, A.; Teichmann, T. The role of abscisic acid and auxin in the response of poplar to abiotic stress-Popko-2010-Plant Biology-Wiley Online Library. Plant Biol. 2010, 12, 242–258. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, H.; Zhao, Y.; Feng, Z.; Li, Q.; Yang, H.-Q.; Luan, S.; Li, J.; He, Z.-H. Auxin controls seed dormancy through stimulation of abscisic acid signaling by inducing ARF-mediated ABI3 activation in Arabidopsis. Proc. Natl. Acad. Sci. USA 2013, 110, 15485–15490. [Google Scholar] [CrossRef] [Green Version]
- Hussain, S.; Nanda, S.; Zhang, J.; Rehmani, M.I.A.; Suleman, M.; Li, G.; Hou, H. Auxin and Cytokinin Interplay during Leaf Morphogenesis and Phyllotaxy. Plants 2021, 10, 1732. [Google Scholar] [CrossRef]
- Rupawalla, Z.; Shaw, L.; Ross, I.L.; Schmidt, S.; Hankamer, B.; Wolf, J. Germination screen for microalgae-generated plant growth biostimulants. Algal Res. 2022, 66, 102784. [Google Scholar] [CrossRef]
- Rouphael, Y.; Colla, G.; Pascale, S.D. Sprouts, Microgreens and Edible Flowers as Novel Functional Foods. Agronomy 2021, 11, 2568. [Google Scholar] [CrossRef]
- Liu, H.; Kang, Y.; Zhao, X.; Liu, Y.; Zhang, X.; Zhang, S. Effects of elicitation on bioactive compounds and biological activities of sprouts. J. Funct. Foods 2019, 53, 136–145. [Google Scholar] [CrossRef]
- Osuna-Ruíz, I.; Ledezma, A.K.D.; Martínez-Montaño, E.; Salazar-Leyva, J.A.; Tirado, V.A.R.; García, I.B. Enhancement of in-vitro antioxidant properties and growth of amaranth seed sprouts treated with seaweed extracts. J. Appl. Phycol. 2022, 1–11. [Google Scholar] [CrossRef]
- Zhang, X.; Schmidt, R.E. The impact of growth regulators on the α-tocopherol status in water-stressed Poa pratensis. Int. Turfgrass Soc. Res. J. 1997, 8, 1364–1371. [Google Scholar]
- González-Pérez, B.K.; Rivas-Castillo, A.M.; Valdez-Calderón, A.; Gayosso-Morales, M.A. Microalgae as biostimulants: A new approach in agriculture. World J. Microbiol. Biotechnol. 2021, 38, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Qi, M.; Guo, J.; Zhou, C.; Yan, X.; Ruan, R.; Cheng, P. The Active Phytohormone in Microalgae: The Characteristics, Efficient Detection, and Their Adversity Resistance Applications. Molecules 2021, 27, 46. [Google Scholar] [CrossRef]
- Michalak, I.; Chojnacka, K.; Saeid, A. Plant Growth Biostimulants, Dietary Feed Supplements and Cosmetics Formulated with Supercritical CO2 Algal Extracts. Molecules 2017, 22, 66. [Google Scholar] [CrossRef]
- Pan, S.; Jeevanandam, J.; Danquah, M.K. Benefits of Algal Extracts in Sustainable Agriculture. In Grand Challenges in Algae Biotechnology; Hallmann, A., Rampelotto, P., Eds.; SpringerLink: Berlin/Heidelberg, Germany, 2020; pp. 501–534. [Google Scholar]
- Das, P.; Khan, S.; Chaudhary, A.K.; AbdulQuadir, M.; Thaher, M.I.; Al-Jabri, H. Potential Applications of Algae-Based Bio-fertilizer. In Biofertilizers for Sustainable Agriculture and Environment. Soil Biology; Giri, B., Prasad, R., Wu, Q.S., Varma, A., Eds.; SpringerLink: Berlin/Heidelberg, Germany, 2019. [Google Scholar]
- Fornes, F.; Sánchez-Perales, M.; Guardiola, J.L. Effect of a Seaweed Extract on the Productivity of ’de Nules’ Clementine Mandarin and Navelina Orange. Botanica Marina 2002, 45, 486–489. [Google Scholar] [CrossRef]
- Adamczyk-Grochala, J.; Wnuk, M.; Duda, M.; Zuczek, J.; Lewinska, A. Treatment with Modified Extracts of the Microalga Planktochlorella nurekis Attenuates the Development of Stress-Induced Senescence in Human Skin Cells. Nutrients 2020, 12, 1005. [Google Scholar] [CrossRef] [Green Version]
- Szpyrka, E.; Broda, D.; Oklejewicz, B.; Podbielska, M.; Slowik-Borowiec, M.; Jagusztyn, B.; Chrzanowski, G.; Kus-Liskiewicz, M.; Duda, M.; Zuczek, J.; et al. A Non-Vector Approach to Increase Lipid Levels in the Microalga Planktochlorella nurekis. Molecules 2020, 25, 270. [Google Scholar] [CrossRef] [Green Version]
- Potocki, L.; Oklejewicz, B.; Kuna, E.; Szpyrka, E.; Duda, M.; Zuczek, J. Application of Green Algal Planktochlorella nurekis Biomasses to Modulate Growth of Selected Microbial Species. Molecules 2021, 26, 4038. [Google Scholar] [CrossRef]
- Czyczyło-Mysza, I.; Marcińska, I.; Skrzypek, E.; Cyganek, K.; Juzoń, K.; Karbarz, M. QTL mapping for germination of seeds obtained from previous wheat generation under drought. Cent. Eur. J. Biol. 2014, 9, 374–382. [Google Scholar] [CrossRef]
- Hernández-Herrera, R.M.; Santacruz-Ruvalcaba, F.; Ruiz-López, M.A.; Norrie, J.; Hernández-Carmona, G. Effect of liquid seaweed extracts on growth of tomato seedlings (Solanum lycopersicum L.). J. Appl. Phycol. 2013, 26, 619–628. [Google Scholar] [CrossRef]
- Lu, Q.; Yu, X. From manure to high-value fertilizer: The employment of microalgae as a nutrient carrier for sustainable agriculture. Algal Res. 2022, 67, 102855. [Google Scholar] [CrossRef]
- Kumar, N.A.; Vanlalzarzova, B.; Sridhar, S.; Baluswami, M. Effect of liquid seaweed fertilizer of Sargassum wightii grev. on the growth and biochemical content of green gram (Vigna radiata (L.) R. wilczek). Recent Res. Sci. Technol. 2015, 4, 40–45. [Google Scholar]
- Hamouda, M.M.; Saad-Allah, K.M.; Gad, D. Potential of Seaweed Extract on Growth, Physiological, Cytological and Biochemical Parameters of Wheat (Triticum aestivum L.) Seedlings. J. Soil Sci. Plant Nutr. 2022, 22, 1818–1831. [Google Scholar] [CrossRef]
- Mohy El-Din, S.M. Utilization of seaweed extracts as bio-fertilizers to stimulate the growth of wheat seedlings. Egypt. J. Exp. Biol. 2015, 11, 31–39. [Google Scholar]
- Kasim, W.A.E.A.; Saad-Allah, K.M.; Hamouda, M. Seed priming with extracts of two seaweeds alleviates the physiological and molecular impacts of salinity stress on radish (Raphanus sativus). Int. J. Agric. Biol. 2016, 18, 653–660. [Google Scholar] [CrossRef]
- Kusvuran, S. Microalgae (Chlorella vulgaris Beijerinck) alleviates drought stress of broccoli plants by improving nutrient uptake, secondary metabolites, and antioxidative defense system. Hortic. Plant J. 2021, 7, 221–231. [Google Scholar] [CrossRef]
- Faheed, F.A.; Fattah, Z.A. Effect of Chlorella vulgaris as Bio-fertilizer on Growth Parameters and Metabolic Aspects of Lettuce Plant. Plant J. Agric. Soc. Sci. 2008, 4, 165–169. [Google Scholar] [CrossRef] [Green Version]
- Finkelstein, R.R.; Gampala, S.S.L.; Rock, C.D. Abscisic Acid Signaling in Seeds and Seedlings. Plant Cell 2002, 14. [Google Scholar] [CrossRef] [Green Version]
- Asif, R.; Yasmin, R.; Mustafa, M.; Ambreen, A.; Mazhar, M.; Rehman, A.; Umbreen, S.; Ahmad, M.; Asif, R.; Yasmin, R.; et al. Chapter 7: Phytohormones as Plant Growth Regulators and Safe Protectors against Biotic and Abiotic Stress. In Plant Hormones: Recent Advances, New Perspectives and Applications; Hano, C., Ed.; IntechOpen: London, UK, 2022. [Google Scholar] [CrossRef]
- Asimakis, E.; Shehata, A.A.; Eisenreich, W.; Acheuk, F.; Lasram, S.; Basiouni, S.; Emekci, M.; Ntougias, S.; Taner, G.; May-Simera, H.; et al. Algae and Their Metabolites as Potential Bio-Pesticides. Microorganisms 2022, 10, 307. [Google Scholar] [CrossRef]
- Han, X.; Zeng, H.; Bartocci, P.; Fantozzi, F.; Yan, Y. Phytohormones and Effects on Growth and Metabolites of Microalgae: A Review. Fermentation 2018, 4, 25. [Google Scholar] [CrossRef]
- Kapoore, R.V.; Wood, E.E.; Llewellyn, C.A. Algae biostimulants: A critical look at microalgal biostimulants for sustainable agricultural practices. Biotechnol. Adv. 2021, 49, 107754. [Google Scholar] [CrossRef]
- Du, K.; Tao, H.; Wen, X.; Geng, Y.; Li, Y. Enhanced growth and lipid production of Chlorella pyrenoidosa by plant growth regulator GA3. Fresenius Environ. Bull. 2015, 24, 3414–3419. [Google Scholar]
- Tarakhovskaya, E.R.; Maslov, Y.I.; Shishova, M.F. Phytohormones in algae. Russ. J. Plant Physiol. 2007, 54, 163–170. [Google Scholar] [CrossRef]
- Rover, S.; de Freitas, M.B.; Barcelos-Oliveira, J.L.; Stadnik, M.J. An algal extract enriched with amino acids increases the content of leaf pigments but also the susceptibility to the powdery mildew of lettuce. Phytoparasitica 2022, 1–12. [Google Scholar] [CrossRef]
- Ebert, A.W. Sprouts and Microgreens—Novel Food Sources for Healthy Diets. Plants 2022, 11, 571. [Google Scholar] [CrossRef]
- Michell, K.A.; Isweiri, H.; Newman, S.E.; Bunning, M.; Bellows, L.L.; Dinges, M.M.; Grabos, L.E.; Rao, S.; Foster, M.T.; Heuberger, A.L.; et al. Microgreens: Consumer sensory perception and acceptance of an emerging functional food crop. J. Food Sci. 2020, 85, 926–935. [Google Scholar] [CrossRef] [PubMed]
- Microgreens Market by Type (Broccoli, Cabbage, Cauliflower, Arugula, Peas, Basil, Radish, Cress, Others), by Farming (Indoor vertical farming, Commercial greenhouse, Others), by End User (Retail, Food Service, Others): Global Opportunity Analysis and Industry Forecast, 2019–2028. Available online: https://www.alliedmarketresearch.com/microgreens-market-A08733 (accessed on 10 December 2022).
- Caracciolo, F.; El-Nakhel, C.; Raimondo, M.; Kyriacou, M.C.; Cembalo, L.; Pascale, S.D.; Rouphael, Y. Sensory Attributes and Consumer Acceptability of 12 Microgreens Species. Agronomy 2020, 10, 1043. [Google Scholar] [CrossRef]


















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Observation | Method | Unit | Day of Measurement |
|---|---|---|---|
| Germination energy | %GE formula | % | at day 3 |
| Length of root and leaves | Ruler | cm | at day 5 |
| Fresh plant weight | Scale (Radwag PS 210.R2, precision 1 mg) | g | at day 5 |
| Parameter | Conc. | Broccoli | Lettuce | Wheat | Radish |
|---|---|---|---|---|---|
| Germination energy | 1% | 5, 11 | 1,6,7,8,9,10 | 8 | 7 |
| 5% | n | 1,2,3,6,7,8,9,11,WT | 10 | 2,3 | |
| 10% | n | 1,2,3,4,5,6,7,8,9,10,11,WT | 4 | 2,3,4,7,10,WT | |
| Root length | 1% | 1,3 | 1,3,4,5,6,7, | 1,5,6,7,8,WT | 9,WT |
| 5% | 3,9,10 | 7,10,11,WT | 1,3,5,7,8,9,10,WT | 3,6,11 | |
| 10% | n | 10 | 1,2,3,5,7,8,9,10,11,WT | 7,11 | |
| Leaf length | 1% | n | 10 | 2,8,9,11 | 2,4,8,9,11,WT |
| 5% | 11 | 1,WT | 1,2,4,6,7,9,11 | 1,3,11 | |
| 10% | n | 9 | 4,6,8 | 1,2,3,4,5,6,7,8,9,10,11,WT | |
| Fresh weight | 1% | n | 6,8 | WT | 7,9,10,WT |
| 5% | 10 | 1,3,5,6,7,8,9,11,WT | 3 | 1,3,4,8,11 | |
| 10% | n | 2,3,4,5,6,7,9,WT | 4 | 1,4,5,7,9,10,11 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karbarz, M.; Piziak, M.; Żuczek, J.; Duda, M. Influence of Microalgae Planktochlorella nurekis Clones on Seed Germination. Agronomy 2023, 13, 9. https://doi.org/10.3390/agronomy13010009
Karbarz M, Piziak M, Żuczek J, Duda M. Influence of Microalgae Planktochlorella nurekis Clones on Seed Germination. Agronomy. 2023; 13(1):9. https://doi.org/10.3390/agronomy13010009
Chicago/Turabian StyleKarbarz, Małgorzata, Magdalena Piziak, Janusz Żuczek, and Magdalena Duda. 2023. "Influence of Microalgae Planktochlorella nurekis Clones on Seed Germination" Agronomy 13, no. 1: 9. https://doi.org/10.3390/agronomy13010009