
Potential of Pseudomonas and Trichoderma from the Brazilian Amazon as Biocontrol Agents against the Wheat Blast Disease

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. In Vitro Antagonism of Fluorescent Pseudomonas against the Wheat Blast Pathogen
2.2. In Vitro Antagonism of Trichoderma against the Wheat Blast Pathogen
2.3. Scanning Electron Microscopy Analyses of In Vitro Pathogen–Biocontrol Agents Interactions
2.4. Potential of Pseudomonas and Trichoderma as Biocontrol Agents Controlling Wheat Blast
3. Results
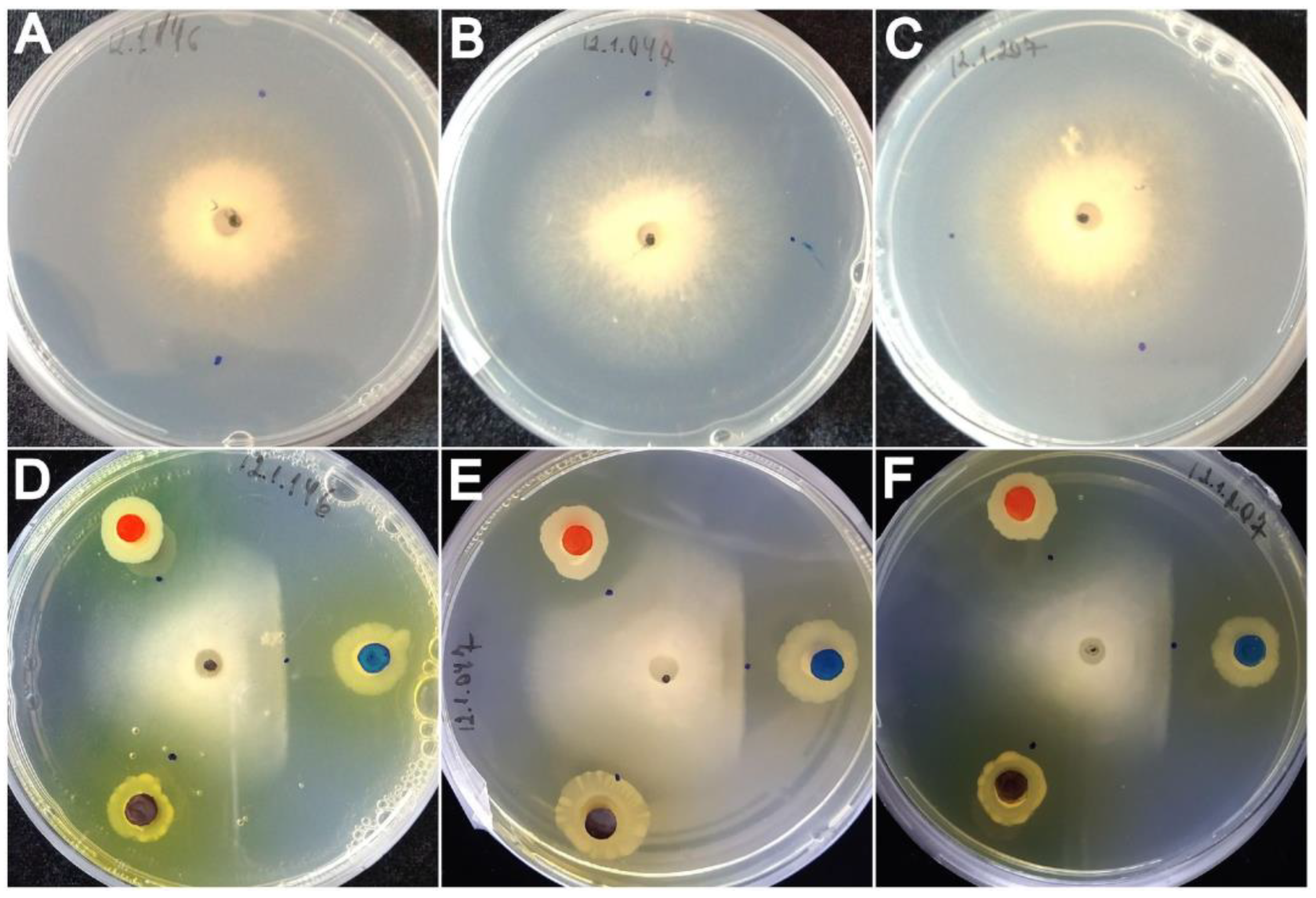
3.1. In Vitro Antagonism of Fluorescent Pseudomonas against the Wheat Blast Pathogen
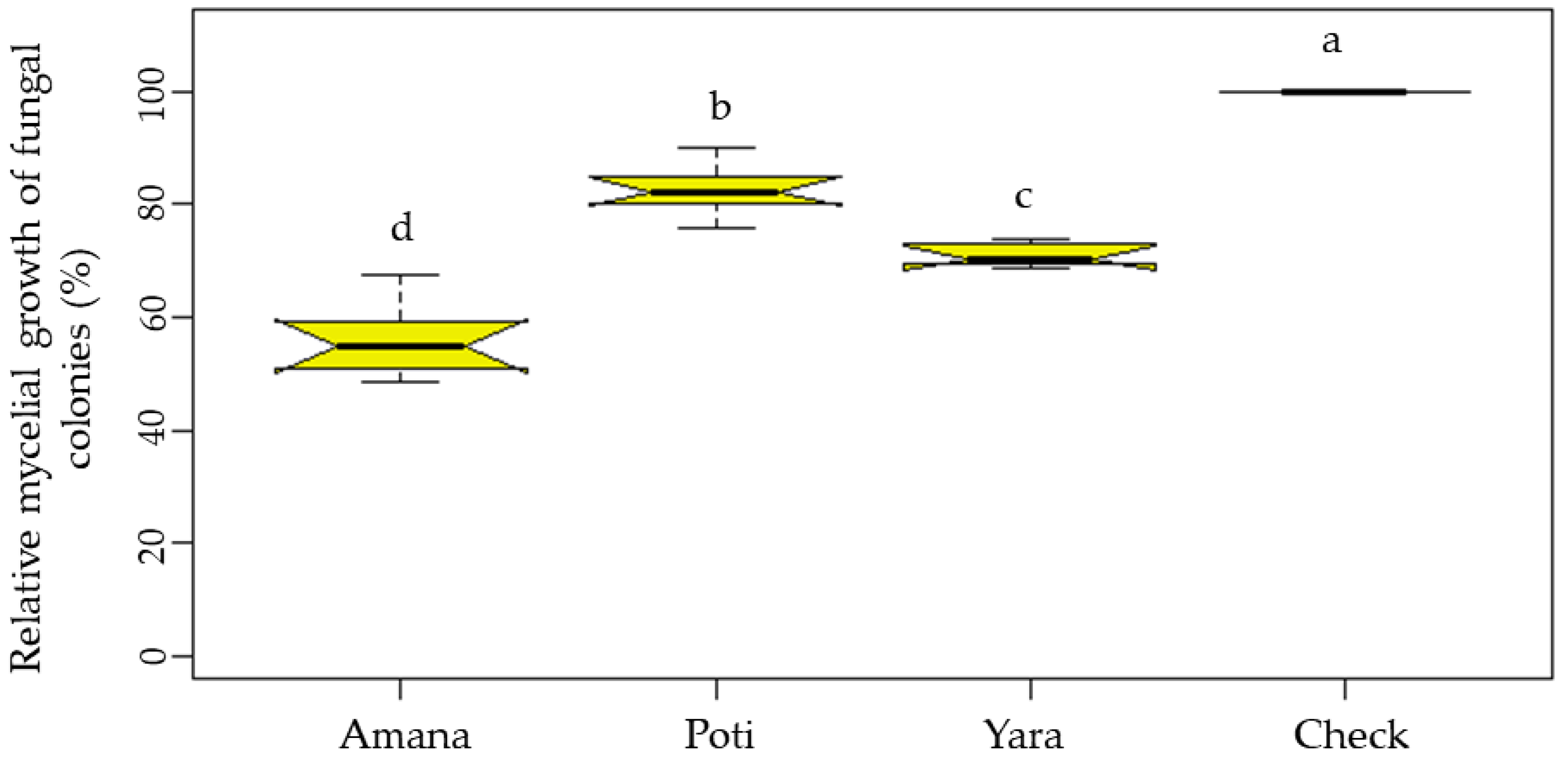
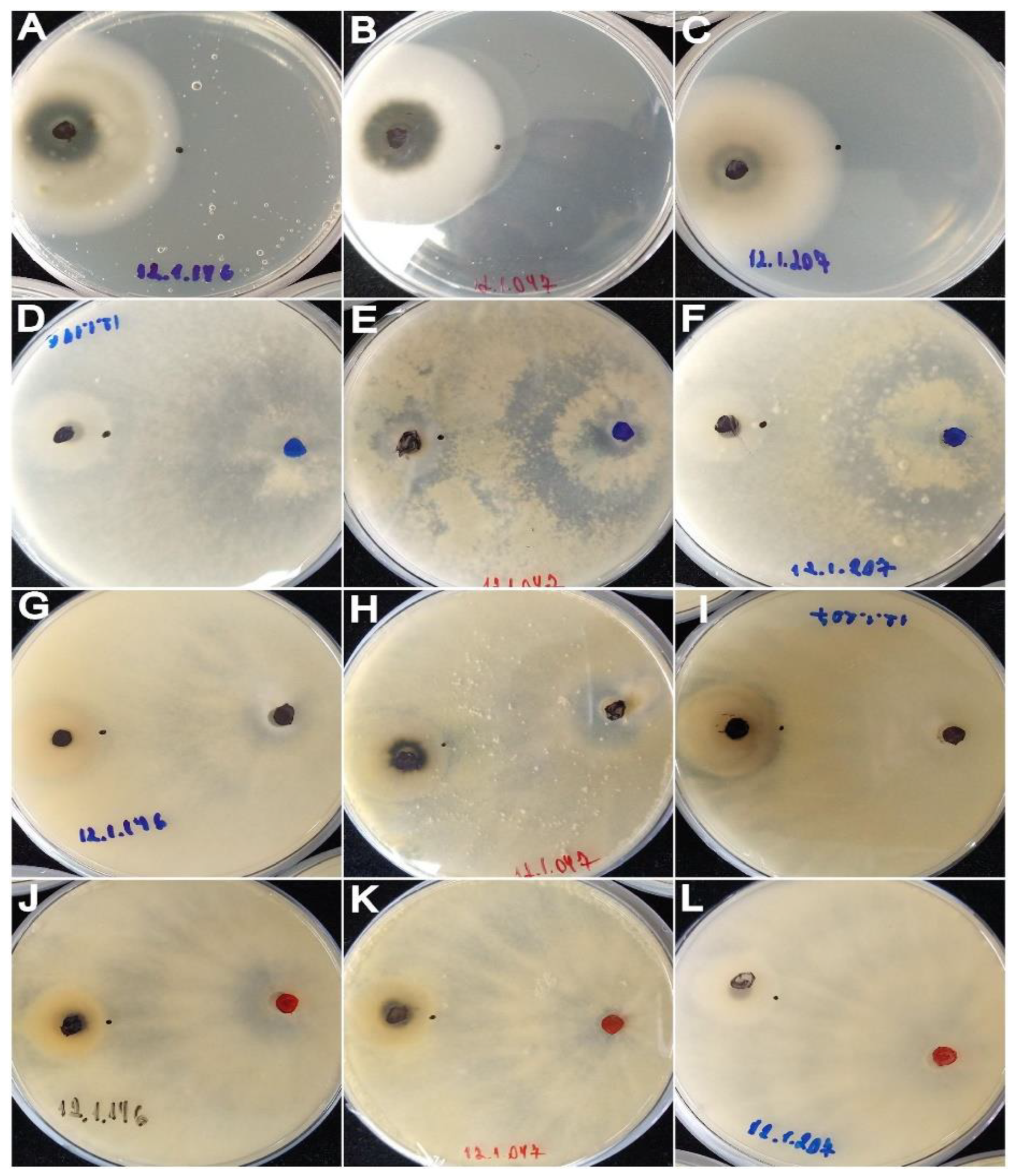
3.2. In Vitro Antagonism of Trichoderma against the Wheat Blast Pathogen
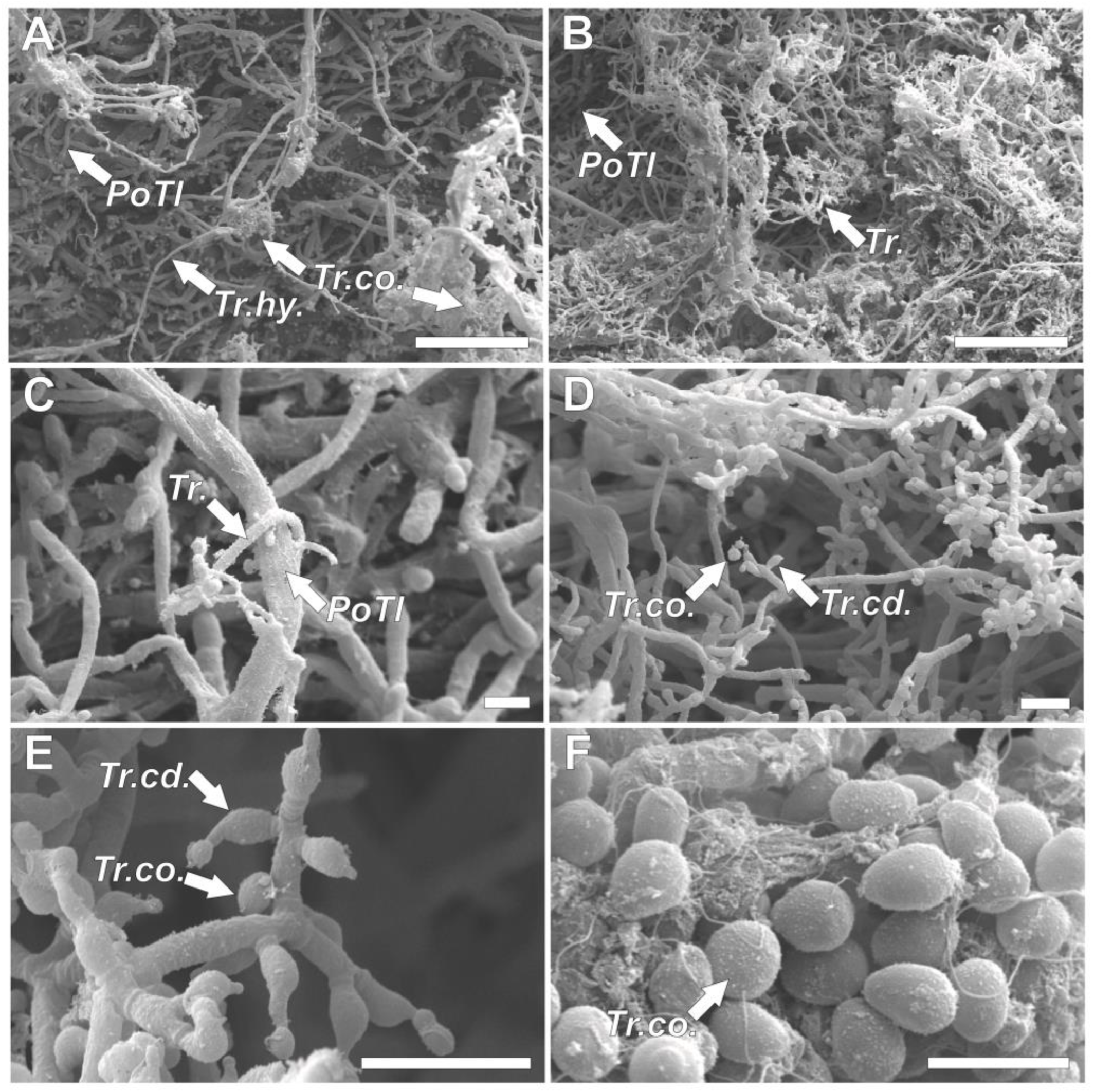
3.3. Scanning Electron Microscopy Analyses of In Vitro Pathogen–Biocontrol Agents Interactions
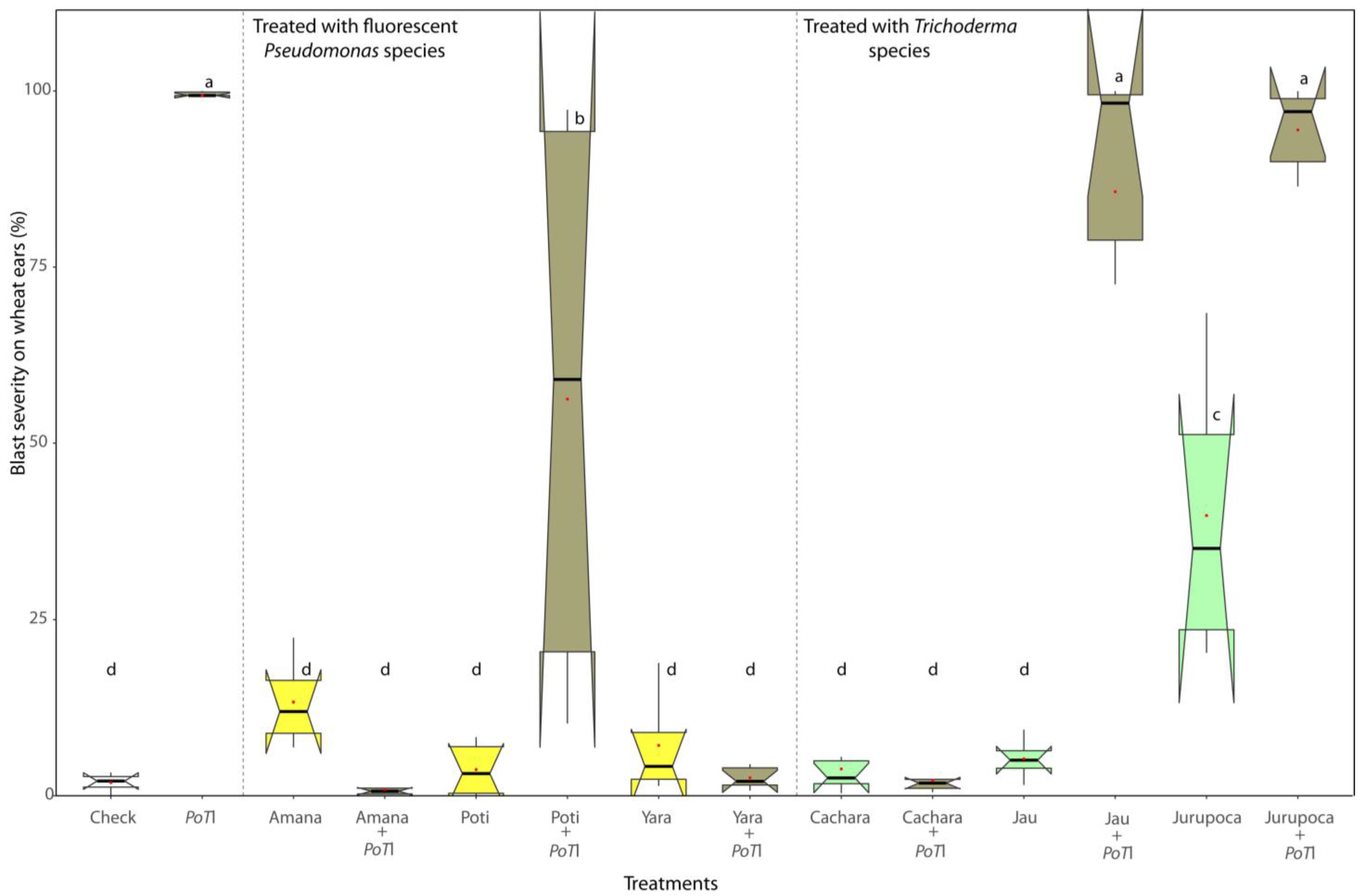
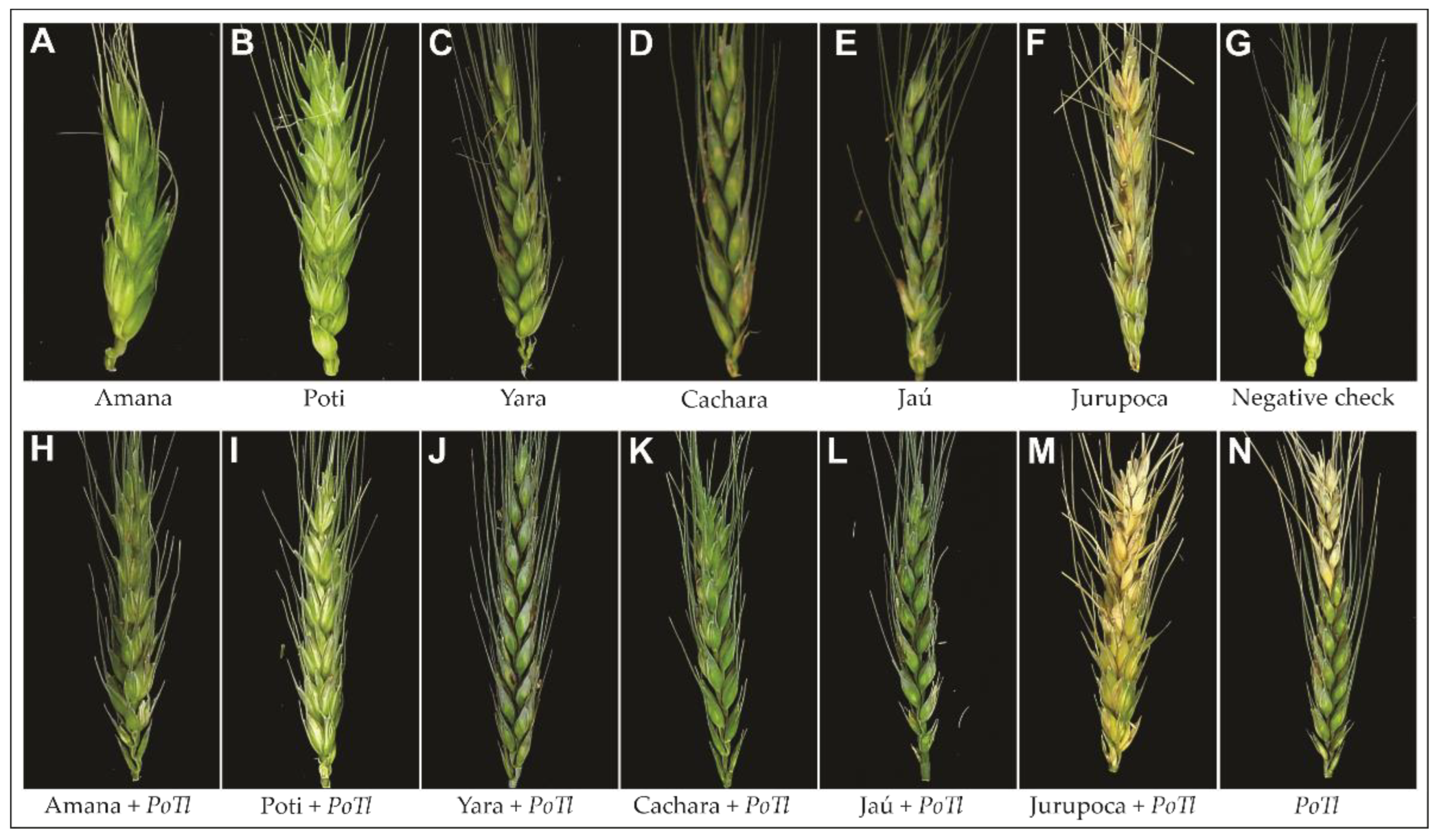
3.4. Potential of Pseudomonas and Trichoderma as Biocontrol Agents Controlling Wheat Blast In Vivo
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Duveiller, E.A.; Singh, P.K.; He, X.Y. Wheat Blast: An Emerging Disease in South America Potentially Threatening Wheat Production. In Wheat World Book; A History of Wheat; Bonjean, A., van Ginkel, M., Eds.; Lavoisier: Paris, France, 2016; Volume 3, pp. 1107–1122. [Google Scholar]
- Urashima, A.S.; Kato, H. Varietal Resistance and Chemical Control of Wheat Blast Fungus. Summa Phytopathol. 1994, 20, 107–112. [Google Scholar]
- Ceresini, P.C.; Castroagudín, V.L.; Rodrigues, F.Á.; Rios, J.A.; Eduardo Aucique-Pérez, C.; Moreira, S.I.; Alves, E.; Croll, D.; Maciel, J.L.N. Wheat Blast: Past, Present, and Future. Annu. Rev. Phytopathol. 2018, 56, 427–456. [Google Scholar] [CrossRef] [PubMed]
- Gladieux, P.; Condon, B.; Ravel, S.; Soanes, D.; Maciel, J.L.N.; Nhani, A.; Chen, L.; Terauchi, R.; Lebrun, M.-H.; Tharreau, D.; et al. Gene Flow between Divergent Cereal- and Grass-Specific Lineages of the Rice Blast Fungus Magnaporthe oryzae. mBio 2018, 9, e01219-17. [Google Scholar] [CrossRef]
- Castroagudín, V.L.; Moreira, S.I.; Pereira, D.A.; Moreira, S.S.; Brunner, P.C.; Maciel, J.L.; Crous, P.W.; McDonald, B.A.; Alves, E.; Ceresini, P.C. Pyricularia graminis-tritici, a New Pyricularia Species Causing Wheat Blast. Pers.-Mol. Phylogeny Evol. Fungi 2016, 37, 199–216. [Google Scholar] [CrossRef]
- Goulart, A.C.P.; Sousa, P.G.; Urashima, A.S. Danos Em Trigo Causados Pela Infecção de Pyricularia grisea. Summa Phytopathol. 2007, 33, 358–363. [Google Scholar] [CrossRef]
- Ceresini, P.C.; Castroagudín, V.L.; Rodrigues, F.Á.; Rios, J.A.; Aucique-Pérez, C.E.; Moreira, S.I.; Croll, D.; Alves, E.; de Carvalho, G.; Maciel, J.L.N.; et al. Wheat Blast: From Its Origins in South America to Its Emergence as a Global Threat. Mol. Plant Pathol. 2019, 20, 155–172. [Google Scholar] [CrossRef]
- Castroagudín, V.L.; Ceresini, P.C.; de Oliveira, S.C.; Reges, J.T.; Maciel, J.L.; Bonato, A.L.; Dorigan, A.F.; McDonald, B.A. Resistance to QoI Fungicides Is Widespread in Brazilian Populations of the Wheat Blast Pathogen Magnaporthe oryzae. Phytopathology 2015, 105, 284–294. [Google Scholar] [CrossRef] [PubMed]
- Poloni, N.M.; Carvalho, G.; Nunes Campos Vicentini, S.; Francis Dorigan, A.; Nunes Maciel, J.L.; McDonald, B.A.; Intra Moreira, S.; Hawkins, N.; Fraaije, B.A.; Kelly, D.E. Widespread Distribution of Resistance to Triazole Fungicides in Brazilian Populations of the Wheat Blast Pathogen. Plant Pathol. 2021, 70, 436–448. [Google Scholar] [CrossRef]
- Dorigan, A.F.; de Carvalho, G.; Poloni, N.M.; Negrisoli, M.M.; Maciel, J.L.N.; Ceresini, P.C. Resistance to Triazole Fungicides in Pyricularia Species Is Associated with Invasive Plants from Wheat Fields in Brazil. Acta Sci. Agron. 2019, 41, e39332. [Google Scholar] [CrossRef]
- Vicentini, S.N.; Casado, P.S.; de Carvalho, G.; Moreira, S.I.; Dorigan, A.F.; Silva, T.C.; Silva, A.G.; Custódio, A.A.; Gomes, A.C.S.; Nunes Maciel, J.L. Monitoring of Brazilian Wheat Blast Field Populations Reveals Resistance to QoI, DMI, and SDHI Fungicides. Plant Pathol. 2022, 71, 304–321. [Google Scholar] [CrossRef]
- Ministry of Agriculture. Livestock and Supply (MAPA) Agrofit System 2022. Available online: https://agrofit.agricultura.gov.br/agrofit_cons/principal_agrofit_cons (accessed on 29 June 2022).
- Köhl, J.; Kolnaar, R.; Ravensberg, W.J. Mode of Action of Microbial Biological Control Agents Against Plant Diseases: Relevance Beyond Efficacy. Front. Plant Sci. 2019, 10, 845. [Google Scholar] [CrossRef] [Green Version]
- Thambugala, K.M.; Daranagama, D.A.; Phillips, A.J.L.; Kannangara, S.D.; Promputtha, I. Fungi vs. Fungi in Biocontrol: An Overview of Fungal Antagonists Applied Against Fungal Plant Pathogens. Front. Cell. Infect. Microbiol. 2020, 10, 604923. [Google Scholar] [CrossRef]
- Müller, T.; Ruppel, S.; Behrendt, U.; Lentzsch, P.; Müller, M.E.H. Antagonistic Potential of Fluorescent Pseudomonads Colonizing Wheat Heads Against Mycotoxin Producing Alternaria and Fusaria. Front. Microbiol. 2018, 9, 2124. [Google Scholar] [CrossRef]
- El-Katatny, M.H.; Somitsch, W.; Robra, K.H.; El-Katatny, M.S.; Gübitz, G.M. Production of Chitinase and β-1,3-Glucanase by Trichoderma Harzianum for Control of the Phytopathogenic Fungus Sclerotium Rolfsii. Food Technol. Biotechnol. 2000, 38, 173–180. [Google Scholar]
- Morandi, M.A.B.; Bettiol, W. Controle Biológico de Doenças de Plantas No Brasil. In Biocontrole de Doenças de Plantas: Uso e Perspectivas; Bettiol, W., Morandi, M.A., Eds.; Embrapa Meio Ambiente: Jaguariúna, Brazil, 2009; pp. 7–14. [Google Scholar]
- Haddad, F.; Maffia, L.A.; Mizubuti, E.S.G.; Teixeira, H. Biological Control of Coffee Rust by Antagonistic Bacteria under Field Conditions in Brazil. Biol. Control 2009, 49, 114–119. [Google Scholar] [CrossRef]
- Halfeld-Vieira, B.A.; Romeiro, R.S.; Mizubuti, E.S.G. Métodos de Isolamento de Bactérias Do Filoplano de Tomateiro Visando Populações Específicas e Implicações Como Agentes de Biocontrole. Fitopatol. Bras. 2004, 29, 638–643. [Google Scholar] [CrossRef]
- Mizubuti, E.; Maffia, L.; Muchovej, J.; Romeiro, R.; Batista, U. Selection of Isolates of Bacillus Subtilis with Potential for the Control of Dry Bean Rust. Fitopatol. Bras. 1995, 20, 540–544. [Google Scholar]
- Romeiro, R.S.; Neves, D.M.S.; Halfed-Vieira, B.A.; Mizubuti, E.S.G.; Deuner, C.C. Inadequação de Apenas Um Patógeno Desafiante Na Seleção Massal de Residentes de Filoplano Para Fins de Controle Biológico—Um Caso. Summa Phytopathol. 2000, 26, 142. [Google Scholar]
- Bettiol, W.; Morandi, M.A. Biocontrole de Doenças de Plantas: Uso e Perspectivas; Embrapa Meio Ambiente: Jaguariúna, Brazil, 2009; ISBN 85-85771-48-8. [Google Scholar]
- Silva, G.B.P.; Heckler, L.I.; Santos, R.F.; Durigon, M.R.; Blume, E. Identificação e Utilização de Trichoderma Spp. Armazenados e Nativos No Biocontrole de Sclerotinia Sclerotiorum. Rev. Caatinga 2015, 28, 33–42. [Google Scholar] [CrossRef]
- Kong, P.; Hong, C. Biocontrol of Boxwood Blight by Trichoderma Koningiopsis Mb2. Crop Prot. 2017, 98, 124–127. [Google Scholar] [CrossRef]
- Yu, C.; Luo, X. Trichoderma Koningiopsis Controls Fusarium Oxysporum Causing Damping-off in Pinus Massoniana Seedlings by Regulating Active Oxygen Metabolism, Osmotic Potential, and the Rhizosphere Microbiome. Biol. Control 2020, 150, 104352. [Google Scholar] [CrossRef]
- Ruangwong, O.-U.; Pornsuriya, C.; Pitija, K.; Sunpapao, A. Biocontrol Mechanisms of Trichoderma Koningiopsis PSU3-2 against Postharvest Anthracnose of Chili Pepper. J. Fungi 2021, 7, 276. [Google Scholar] [CrossRef]
- Kloepper, J.W.; Leong, J.; Teintze, M.; Schroth, M.N. Enhanced Plant Growth by Siderophores Produced by Plant Growth-Promoting Rhizobacteria. Nature 1980, 286, 885–886. [Google Scholar] [CrossRef]
- Ferreira, E.P.B.; Voss, M.; Santos, H.P.; De-Polli, H.; Neves, M.C.P.; Rumjanek, N.G. Diversidade de Pseudomonas Fluorescentes Em Diferentes Sistemas de Manejo Do Solo e Rotação de Culturas. Rev. Bras. Ciênc. Agrar. 2009, 4, 140–148. [Google Scholar] [CrossRef] [Green Version]
- Subashri, R.; Gurusamy, R.; Sakthivel, N.; Raman, B.; Sakthivel, B.; Maheshwari, D. Biological Control of Pathogens and Plant Growth Promotion Potential of Fluorescent Pseudomonads. In Bacteria in Agrobiology: Disease Management; Maheshwari, D.K., Ed.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 77–110. [Google Scholar]
- Vicentini, S.N.C.; de Carvalho, G.; Krug, L.D.; Nunes, T.C.; da Silva, A.G.; Moreira, S.I.; Gonçalves, L.M.d.D.P.; Silva, T.C.; Ceresini, P.C. Bioprospecting Fluorescent Pseudomonas from the Brazilian Amazon for the Biocontrol of Signal Grass Foliar Blight. Agronomy 2022, 12, 1395. [Google Scholar] [CrossRef]
- Ujváry, I. Chapter 3—Pest Control Agents from Natural Products. In Handbook of Pesticide Toxicology, 2nd ed.; Krieger, R.I., Krieger, W.C., Eds.; Academic Press: San Diego, CA, USA, 2001; pp. 109–179. ISBN 978-0-12-426260-7. [Google Scholar]
- Nunes, T.C. Controle Biológico da Queima-Das-Folhas e Morte de Pastagens de Braquiária (Urochloa Brizantha) Causadas Por Rhizoctonia Solani AG-1 IA. Ph.D. Dissertation, Universidade Estadual Paulista “Júlio de Mesquita Filho” Faculdade de Engenharia (Campus de Ilha Solteira), Ilha Solteira, Brazil, 2019. [Google Scholar]
- Chavarro-Mesa, E.; Ceresini, P.; Pereira, D.; Vicentini, S.; Silva, T.; Ramos-Molina, L.; Negrisoli, M.; Schurt, D.; Vieira Júnior, J.R. A Broad Diversity Survey of Rhizoctonia Species from the Brazilian Amazon Reveals the Prevalence of R. Solani AG-1 IA on Signal Grass and the New Record of AG-1 IF on Cowpea and Soybeans. Plant Pathol. 2020, 69, 455–466. [Google Scholar] [CrossRef]
- Camporota, P. Antagonisme in Vitro de Trichoderma Spp. Vis-a-Vis de Rhizoctonia Solani Kuhn. Agronomie 1985, 7, 613–620. [Google Scholar] [CrossRef]
- Johansen, D.A. Plant Microtechnique; McGraw-Hill Book Company, Inc.: London, UK, 1940. [Google Scholar]
- Zadoks, J.C.; Chang, T.T.; Konzak, C.F. A Decimal Code for the Growth Stages of Cereals. Weed Res. 1974, 14, 415–421. [Google Scholar] [CrossRef]
- Lamari, L. Assess 2.0: Image Analysis Software for Plant Disease Quantification; American Phytopathological Society: Saint Paul, MN, USA, 2008. [Google Scholar]
- Masák, J.; Čejková, A.; Schreiberová, O.; Řezanka, T. Pseudomonas Biofilms: Possibilities of Their Control. FEMS Microbiol. Ecol. 2014, 89, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Mulcahy, L.R.; Isabella, V.M.; Lewis, K. Pseudomonas aeruginosa Biofilms in Disease. Microb. Ecol. 2014, 68, 1–12. [Google Scholar] [CrossRef]
- Rasamiravaka, T.; Labtani, Q.; Duez, P.; El Jaziri, M. The Formation of Biofilms by Pseudomonas Aeruginosa: A Review of the Natural and Synthetic Compounds Interfering with Control Mechanisms. BioMed Res. Int. 2015, 2015, 759348. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, S.M.; Felício, M.R.; Boas, E.V.; Gonçalves, S.; Costa, F.F.; Samy, R.P.; Santos, N.C.; Franco, O.L. New Frontiers for Anti-Biofilm Drug Development. Pharmacol. Ther. 2016, 160, 133–144. [Google Scholar] [CrossRef] [PubMed]
- Flemming, H.-C.; Wingender, J. The Biofilm Matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef] [PubMed]
- O’Toole, G.; Kaplan, H.B.; Kolter, R. Biofilm Formation as Microbial Development. Annu. Rev. Microbiol. 2000, 54, 49–79. [Google Scholar] [CrossRef]
- Beveridge, T.J.; Makin, S.A.; Kadurugamuwa, J.L.; Li, Z. Interactions between Biofilms and the Environment. FEMS Microbiol. Rev. 1997, 20, 291–303. [Google Scholar] [CrossRef]
- Carvalho, C.C.C.R.; Wick, L.Y.; Heipieper, H.J. Cell Wall Adaptations of Planktonic and Biofilm Rhodococcus Erythropolis Cells to Growth on C5 to C16 N-Alkane Hydrocarbons. Appl. Microbiol. Biotechnol. 2009, 82, 311–320. [Google Scholar] [CrossRef]
- Heipieper, H.J.; Keweloh, H.; Rehm, H.J. Influence of Phenols on Growth and Membrane Permeability of Free and Immobilized Escherichia coli. Appl. Environ. Microbiol. 1991, 57, 1213–1217. [Google Scholar] [CrossRef]
- Velusamy, P.; Gnanamanickam, S.S. The Effect of Bacterial Secondary Metabolites on Bacterial and Fungal Pathogens of Rice. In Secondary Metabolites in Soil Ecology; Karlovsky, P., Ed.; Soil Biology; Springer: Berlin/Heidelberg, Germany, 2008; Volume 14, pp. 93–106. ISBN 978-3-540-74542-6. [Google Scholar]
- Reddy, K.R.N.; Farhana, N.I.; Wardah, A.R.; Salleh, B. Morphological Identification of Foodborne Pathogens Colonizing Rice Grains in South Asia. Pak. J. Biol. Sci. 2010, 13, 794–801. [Google Scholar] [CrossRef]
- Omoboye, O.O.; Oni, F.E.; Batool, H.; Yimer, H.Z.; De Mot, R.; Höfte, M. Pseudomonas Cyclic Lipopeptides Suppress the Rice Blast Fungus Magnaporthe Oryzae by Induced Resistance and Direct Antagonism. Front. Plant Sci. 2019, 10, 901. [Google Scholar] [CrossRef] [PubMed]
- Sousa, T.P.; Chaibub, A.A.; Carvalho Barros Cortes, M.V.; Batista, T.F.C.; Andrade Bezerra, G.; Silva, G.B.; de Filippi, M.C.C. Molecular Identification of Trichoderma Sp. Isolates and Biochemical Characterization of Antagonistic Interaction against Rice Blast. Arch. Microbiol. 2021, 203, 3257–3268. [Google Scholar] [CrossRef] [PubMed]
- Reyes Rondón, T.; Rodríguez Gutiérrez, G.; Alarcón Pérez, L.; Pupo Zayas, A.D. Efectividad in vitro de Trichoderma harzianum (Rifai) en el biocontrol de Rhizoctonia solani Kunh y Pyricularia grisea (Sacc.) en el cultivo del arroz (Oryza sativa L.). Fitosanidad 2005, 9, 57–60. [Google Scholar]
- Victoria Arellano, A.D.; da Silva, G.M.; Guatimosim, E.; da Rosa Dorneles, K.; Moreira, L.G.; Dallagnol, L.J. Seeds Coated with Trichoderma Atroviride and Soil Amended with Silicon Improve the Resistance of Lolium Multiflorum against Pyricularia oryzae. Biol. Control 2021, 154, 104499. [Google Scholar] [CrossRef]
- Adnan, M.; Islam, W.; Shabbir, A.; Khan, K.A.; Ghramh, H.A.; Huang, Z.; Chen, H.Y.H.; Lu, G. Plant Defense against Fungal Pathogens by Antagonistic Fungi with Trichoderma in Focus. Microb. Pathog. 2019, 129, 7–18. [Google Scholar] [CrossRef] [PubMed]
- Troian, R.F.; Steindorff, A.S.; Ramada, M.H.S.; Arruda, W.; Ulhoa, C.J. Mycoparasitism Studies of Trichoderma Harzianum against Sclerotinia Sclerotiorum: Evaluation of Antagonism and Expression of Cell Wall-Degrading Enzymes Genes. Biotechnol. Lett. 2014, 36, 2095–2101. [Google Scholar] [CrossRef] [PubMed]
- Chaverri, P.; Samuels, G.J. Evolution of Habitat Preference and Nutrition Mode in a Cosmopolitan Fungal Genus with Evidence of Interkingdom Host Jumps and Major Shifts in Ecology: Trichoderma Evolution. Evolution 2013, 67, 2823–2837. [Google Scholar] [CrossRef] [PubMed]
- Abdullah, M.T.; Ali, N.Y.; Suleman, P. Biological Control of Sclerotinia Sclerotiorum (Lib.) de Bary with Trichoderma Harzianum and Bacillus amyloliquefaciens. Crop Prot. 2008, 27, 1354–1359. [Google Scholar] [CrossRef]
- Zhang, F.; Ge, H.; Zhang, F.; Guo, N.; Wang, Y.; Chen, L.; Ji, X.; Li, C. Biocontrol Potential of Trichoderma Harzianum Isolate T-Aloe against Sclerotinia Sclerotiorum in Soybean. Plant Physiol. Biochem. 2016, 100, 64–74. [Google Scholar] [CrossRef]
- Juliatti, F.C.; Rezende, A.A.; Juliatti, B.C.M.; Morais, T.P. Trichoderma as a Biocontrol Agent against Sclerotinia Stem Rot or White Mold on Soybeans in Brazil: Usage and Technology. In Trichoderma—The Most Widely Used Fungicide; Shah, M.M., Sharif, U., Buhari, T.R., Eds.; IntechOpen: London, UK, 2019; ISBN 978-1-78923-917-1. [Google Scholar]
- Hermosa, R.; Viterbo, A.; Chet, I.; Monte, E. Plant-Beneficial Effects of Trichoderma and of Its Genes. Microbiology 2012, 158, 17–25. [Google Scholar] [CrossRef]
- Illescas, M.; Morán-Diez, M.E.; Martínez de Alba, Á.E.; Hermosa, R.; Monte, E. Effect of Trichoderma asperellum on Wheat Plants’ Biochemical and Molecular Responses, and Yield under Different Water Stress Conditions. IJMS 2022, 23, 6782. [Google Scholar] [CrossRef]
- Benítez, T.; Rincón, A.M.; Limón, M.C.; Codón, A.C. Biocontrol Mechanisms of Trichoderma Strains. Int. Microbiol. 2004, 7, 249–260. [Google Scholar] [PubMed]
- Monteiro, V.N.; do Nascimento Silva, R.; Steindorff, A.S.; Costa, F.T.; Noronha, E.F.; Ricart, C.A.O.; de Sousa, M.V.; Vainstein, M.H.; Ulhoa, C.J. New Insights in Trichoderma harzianum Antagonism of Fungal Plant Pathogens by Secreted Protein Analysis. Curr. Microbiol. 2010, 61, 298–305. [Google Scholar] [CrossRef]
- Morán-Diez, E.; Hermosa, R.; Ambrosino, P.; Cardoza, R.E.; Gutiérrez, S.; Lorito, M.; Monte, E. The ThPG1 Endopolygalacturonase Is Required for the Trichoderma harzianum—Plant Beneficial Interaction. Mol. Plant Microbe Interact. 2009, 22, 1021–1031. [Google Scholar] [CrossRef] [PubMed]
- Lesueur, D.; Deaker, R.; Herrmann, L.; Bräu, L.; Jansa, J. The Production and Potential of Biofertilizers to Improve Crop Yields. In Bioformulations: For Sustainable Agriculture; Arora, N.K., Mehnaz, S., Balestrini, R., Eds.; Springer: New Delhi, India, 2016; pp. 71–92. ISBN 978-81-322-2777-9. [Google Scholar]
- van Lenteren, J.C.; Bolckmans, K.; Köhl, J.; Ravensberg, W.J.; Urbaneja, A. Biological Control Using Invertebrates and Microorganisms: Plenty of New Opportunities. BioControl 2018, 63, 39–59. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolates | Species | References |
|---|---|---|
| Amana | Pseudomonas wayambapalatensis (P. putida group) | Vicentini et al. [30] |
| Poti | Pseudomonas sp. nov. (P. asplenii group) | Vicentini et al. [30] |
| Yara | Pseudomonas sp. nov. (P. putida group) | Vicentini et al. [30] |
| Cachara | Trichoderma koningiopsis | Nunes [32] |
| Jaú | Trichoderma virens | Nunes [32] |
| Jurupoca | Trichoderma lentiforme | Nunes [32] |
| Source of Variation | df | SS | MS | F | p |
|---|---|---|---|---|---|
| Treatments | 3 | 8381.53 | 2793.84 | 180.60 | 0.0000 *** |
| Error | 28 | 433.16 | 15.47 | ||
| Total CV(%): 5.09 | 31 | 8814.69 |
| Source of Variation | df | SS | MS | F | p |
|---|---|---|---|---|---|
| Treatments | 3 | 33,858.42 | 11,286.14 | 1287.57 | 0.0000 *** |
| Error | 28 | 245.43 | 8.77 | ||
| Total | 31 | ||||
| CV(%): 6.20 |
| Source of Variation | df | MS | F | p |
|---|---|---|---|---|
| Treatments | 13 | 7087.59 | 29.01 | 0.0000 *** |
| Experiments (1 and 2) | 1 | 0.62 | 0.003 | 0.9606 NS |
| Blocks | 2 | 145.24 | 0.59 | 0.5636 NS |
| Treatments*experiments | 13 | 562.36 | 2.30 | 0.0582 NS |
| Treatments*blocks | 25 | 70.30 | 0.29 | 0.9974 NS |
| Error | 16 | 244.34 | ||
| Total | 70 | 105.407.43 | ||
| CV(%): 52.51 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pereira, M.R.d.A.; Moreira, S.I.; Silva, A.G.d.; Nunes, T.C.; Vicentini, S.N.C.; Silva, D.P.d.; Silveira, P.R.d.; Oliveira, T.Y.K.d.; Silva, T.C.; Botelho, D.M.d.S.; et al. Potential of Pseudomonas and Trichoderma from the Brazilian Amazon as Biocontrol Agents against the Wheat Blast Disease. Agronomy 2022, 12, 2003. https://doi.org/10.3390/agronomy12092003
Pereira MRdA, Moreira SI, Silva AGd, Nunes TC, Vicentini SNC, Silva DPd, Silveira PRd, Oliveira TYKd, Silva TC, Botelho DMdS, et al. Potential of Pseudomonas and Trichoderma from the Brazilian Amazon as Biocontrol Agents against the Wheat Blast Disease. Agronomy. 2022; 12(9):2003. https://doi.org/10.3390/agronomy12092003
Chicago/Turabian StylePereira, Maikon Richer de Azambuja, Silvino Intra Moreira, Abimael Gomes da Silva, Tiago Calves Nunes, Samara Nunes Campos Vicentini, Davi Prata da Silva, Patrícia Ricardino da Silveira, Tamiris Yoshie Kiyama de Oliveira, Tatiane Carla Silva, Deila Magna dos Santos Botelho, and et al. 2022. "Potential of Pseudomonas and Trichoderma from the Brazilian Amazon as Biocontrol Agents against the Wheat Blast Disease" Agronomy 12, no. 9: 2003. https://doi.org/10.3390/agronomy12092003