
Transcriptome and Re-Sequencing Analyses Reveal Photosynthesis-Related Genes Involvement in Lutein Accumulation in Yellow Taproot Mutants of Carrot



Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Carotenoid Evaluation in the Taproot and Callus
2.3. Microscopic Identification of Carotenoid Crystals
2.4. Transmission Electron Microscopy (TEM)
2.5. Transcriptome Sequencing of Taproots of wt and ym
2.6. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR) and Gene Expression Analysis
2.7. Resequencing of wt and ym
3. Results
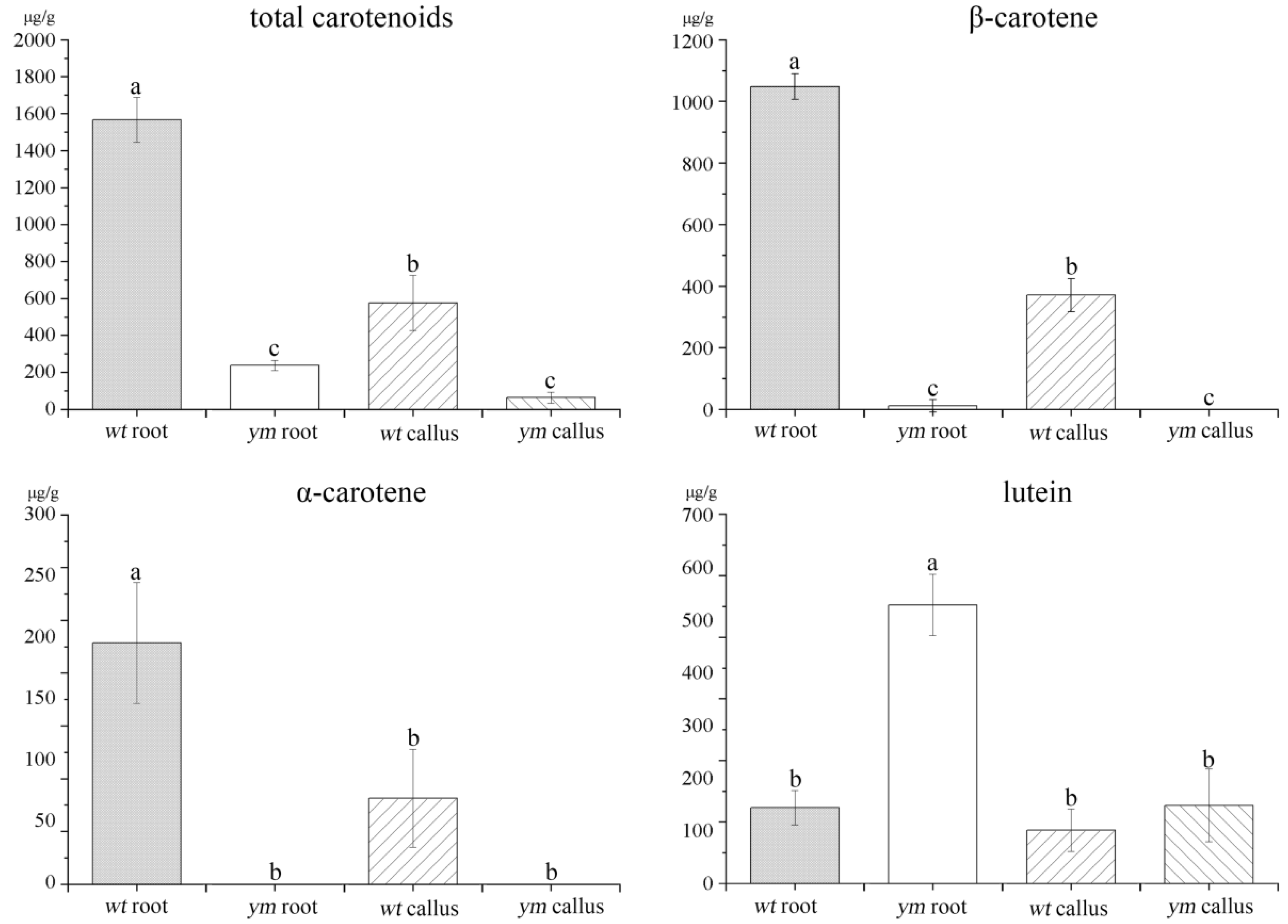
3.1. Carotenoid Content in the Roots and Calli of wt and ym
3.2. Obvious Differences in the Callus of wt and ym at the Histological and Ultrastructural Levels
3.3. Analysis Results of DEGs between wt and ym
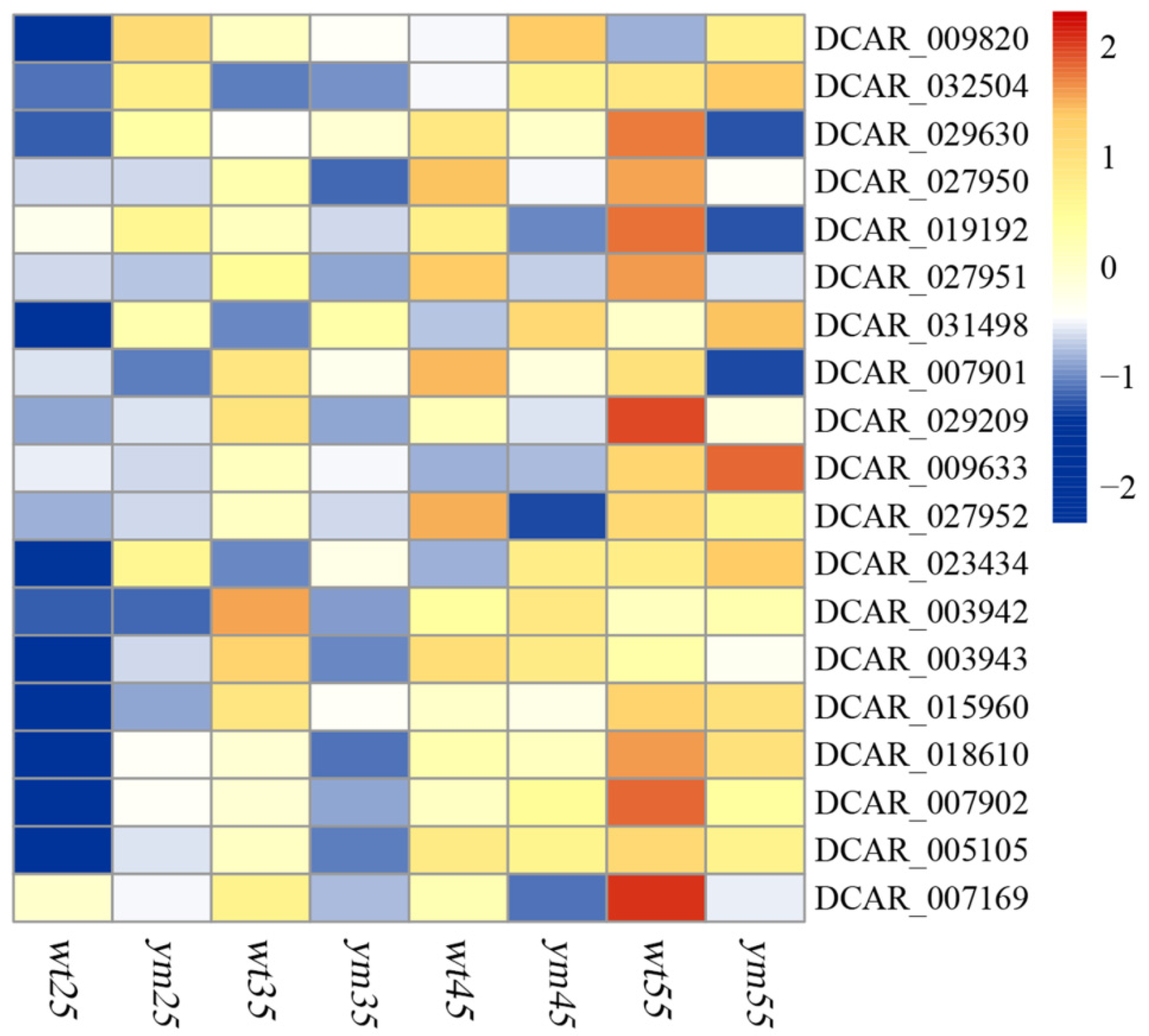
3.4. Identification of Variations of 19 Photosynthesis-Related Genes and 4 Mapped Genes Controlling Carotenoids in the Root of wt and ym
4. Discussion
4.1. The Origin of Globular Chromoplasts in ym Callus Might Be Different from That in wt Callus
4.2. Lut 5 Was the Only Gene of All Mapped Genes Controlling Carotenoids in the Carrot Root That Contained Nonsynonymous Mutations in ym
4.3. Photosynthesis-Related Genes Might Be Involved in Lutein Accumulation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Surles, R.L.; Weng, N.; Simon, P.W.; Tanumihardjo, S.A. Carotenoid profiles and consumer sensory evaluation of specialty carrots (Daucus carota L.) of various colors. J. Agric. Food Chem. 2004, 52, 3417–3421. [Google Scholar] [CrossRef] [PubMed]
- Schweiggert, R.M.; Carle, R. Carotenoid deposition in plant and animal foods and its impact on bioavailability. Crit. Rev. Food Sci. Nutr. 2017, 57, 1807–1830. [Google Scholar] [CrossRef]
- Solymosi, K.; Keresztes, A. Plastid structure, diversification and interconversions II. land plants. Curr. Chem. Biol. 2013, 6, 187–204. [Google Scholar] [CrossRef]
- Schweiggert, R.M.; Mezger, D.; Schimpf, F.; Steingass, C.B.; Carle, R. Influence of chromoplast morphology on carotenoid bioaccessibility of carrot, mango, papaya, and tomato. Food Chem. 2012, 135, 2736–2742. [Google Scholar] [CrossRef] [PubMed]
- Oleszkiewicz, T.; Klimek-Chodacka, M.; Milewska-Hendel, A.; Zubko, M.; Stroz, D.; Kurczynska, E.; Boba, A.; Szopa, J.; Baranski, R. Unique chromoplast organisation and carotenoid gene expression in carotenoid-rich carrot callus. Planta 2018, 248, 1455–1471. [Google Scholar] [CrossRef]
- Iorizzo, M.; Ellison, S.; Senalik, D.; Zeng, P.; Satapoomin, P.; Huang, J.; Bowman, M.; Iovene, M.; Sanseverino, W.; Cavagnaro, P.; et al. A high-quality carrot genome assembly provides new insights into carotenoid accumulation and asterid genome evolution. Nat. Genet. 2016, 48, 657–666. [Google Scholar] [CrossRef] [Green Version]
- Just, B.J.; Santos, C.A.; Fonseca, M.E.; Boiteux, L.S.; Oloizia, B.B.; Simon, P.W. Carotenoid biosynthesis structural genes in carrot (Daucus carota): Isolation, sequence-characterization, single nucleotide polymorphism (SNP) markers and genome mapping. Theor. Appl. Genet. 2007, 114, 693–704. [Google Scholar] [CrossRef]
- Bowman, M.J.; Willis, D.K.; Simon, P.W. Transcript Abundance of Phytoene Synthase 1 and Phytoene Synthase 2 is associated with natural variation of storage root carotenoid pigmentation in carrot. J. Am. Soc. Hortic. Sci. 2014, 139, 63–68. [Google Scholar] [CrossRef] [Green Version]
- Clotault, J.; Peltier, D.; Berruyer, R.; Thomas, M.; Briard, M.; Geoffriau, E. Expression of carotenoid biosynthesis genes during carrot root development. J. Exp. Bot. 2008, 59, 3563–3573. [Google Scholar] [CrossRef] [Green Version]
- Fuentes, P.; Pizarro, L.; Moreno, J.C.; Handford, M.; Rodriguez-Concepcion, M.; Stange, C. Light-dependent changes in plastid differentiation influence carotenoid gene expression and accumulation in carrot roots. Plant Mol. Biol. 2012, 79, 47–59. [Google Scholar] [CrossRef]
- Ma, J.; Li, J.; Xu, Z.; Wang, F.; Xiong, A. Transcriptome profiling of genes involving in carotenoid biosynthesis and accumulation between leaf and root of carrot (Daucus carota L.). Acta Biochim. Biophys. Sin. (Shanghai) 2018, 50, 481–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Ou, C.G.; Zhuang, F.Y.; Ma, Z.G. The dual role of phytoene synthase genes in carotenogenesis in carrot roots and leaves. Mol. Breed. 2014, 34, 2065–2079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreno, J.C.; Pizarro, L.; Fuentes, P.; Handford, M.; Cifuentes, V.; Stange, C. Levels of lycopene beta-cyclase 1 modulate carotenoid gene expression and accumulation in Daucus carota. PLoS ONE 2013, 8, e58144. [Google Scholar] [CrossRef] [Green Version]
- Arango, J.; Jourdan, M.; Geoffriau, E.; Beyer, P.; Welsch, R. Carotene hydroxylase activity determines the levels of both alpha-carotene and total carotenoids in orange carrots. Plant Cell 2014, 26, 2223–2233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arias, D.; Maldonado, J.; Silva, H.; Stange, C. A de novo transcriptome analysis revealed that photomorphogenic genes are required for carotenoid synthesis in the dark-grown carrot taproot. Mol. Genet. Genom. 2020, 295, 1379–1392. [Google Scholar] [CrossRef]
- Buishand, J.G.; Gabelman, W.H. Investigations on the inheritance of color and carotenoid content in phloem and xylem of carrot roots (Daucus carota L.). Euphytica 1979, 28, 611–632. [Google Scholar] [CrossRef]
- Ellison, S.; Senalik, D.; Bostan, H.; Iorizzo, M.; Simon, P. Fine mapping, transcriptome analysis, and marker development for Y2, the gene that conditions beta-carotene accumulation in carrot (Daucus carota L.). G3 2017, 7, 2665–2675. [Google Scholar] [CrossRef] [Green Version]
- Egea, I.; Barsan, C.; Bian, W.; Purgatto, E.; Latche, A.; Chervin, C.; Bouzayen, M.; Pech, J.C. Chromoplast differentiation: Current status and perspectives. Plant Cell Physiol. 2010, 51, 1600–1611. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Yuan, H. Chromoplast biogenesis and carotenoid accumulation. Arch. Biochem. Biophys. 2013, 539, 102–109. [Google Scholar] [CrossRef]
- Ellison, S.L.; Luby, C.H.; Corak, K.E.; Coe, K.M.; Senalik, D.; Iorizzo, M.; Goldman, I.L.; Simon, P.W.; Dawson, J.C. Carotenoid presence is associated with the Or gene in domesticated carrot. Genetics 2018, 210, 1497–1508. [Google Scholar] [CrossRef] [Green Version]
- Coe, K.M.; Ellison, S.; Senalik, D.; Dawson, J.; Simon, P. The influence of the Or and Carotene Hydroxylase genes on carotenoid accumulation in orange carrots [Daucus carota (L.)]. Theor. Appl. Genet. 2021, 134, 3351–3362. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.; Wang, L.G.; Han, Y.; He, Q.Y. ClusterProfiler: An R package for comparing biological themes among gene clusters. Omics 2012, 16, 284–287. [Google Scholar] [CrossRef] [PubMed]
- Alexa, A.; Rahnenführer, J. Gene set enrichment analysis with topGO. Encycl. Syst. Biol. 2007, 9, 589. Available online: https://mirrors.nju.edu.cn/bioconductor/3.2/bioc/vignettes/topGO/inst/doc/topGO.pdf (accessed on 3 May 2016).
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [Green Version]
- Zhu, P.; He, L.; Li, Y.; Huang, W.; Xi, F.; Lin, L.; Zhi, Q.; Zhang, W.; Tang, Y.T.; Geng, C.; et al. OTG-snpcaller: An optimized pipeline based on TMAP and GATK for SNP calling from ion torrent data. PLoS ONE 2014, 9, e97507. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Liu, Z.; Chang, S.; Zhao, Y. An EMS mutant library for carrot and genetic analysis of some mutants. Breed. Sci. 2020, 70, 540–546. [Google Scholar] [CrossRef]
- Hempel, J.; Schadle, C.N.; Sprenger, J.; Heller, A.; Carle, R.; Schweiggert, R.M. Ultrastructural deposition forms and bioaccessibility of carotenoids and carotenoid esters from goji berries (Lycium barbarum L.). Food Chem. 2017, 218, 525–533. [Google Scholar] [CrossRef]
- Paolillo, D.J., Jr.; Garvin, D.F.; Parthasarathy, M.V. The chromoplasts of Or mutants of cauliflower (Brassica oleracea L. var botrytis). Protoplasma 2004, 224, 245–253. [Google Scholar] [CrossRef]
- Roman, M.; Marzec, K.M.; Grzebelus, E.; Simon, P.W.; Baranska, M.; Baranski, R. Composition and (in)homogeneity of carotenoid crystals in carrot cells revealed by high resolution Raman imaging. Spectrochim. Acta. A Mol. Biomol. Spectrosc. 2015, 136, 1395–1400. [Google Scholar] [CrossRef]
- Vásquez-Caicedo, A.L.; Heller, A.; Neidhart, S.; Carle, R. Chromoplast morphology and β-carotene accumulation during postharvest ripening of mango Cv. ‘Tommy Atkins’. J. Agric. Food Chem. 2006, 54, 5769–5776. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.E.; Rensing, K.H.; Douglas, C.J.; Cheng, K.M. Chromoplasts ultrastructure and estimated carotene content in root secondary phloem of different carrot varieties. Planta 2010, 231, 549–558. [Google Scholar] [CrossRef] [PubMed]
- Arscott, S.A.; Tanumihardjo, S.A. Carrots of many colors provide basic nutrition and bioavailable phytochemicals acting as a functional food. Compr. Rev. Food Sci. Food Saf. 2010, 9, 223–239. [Google Scholar] [CrossRef]
- Santos, C.A.; Simon, P.W. QTL analyses reveal clustered loci for accumulation of major provitamin A carotenes and lycopene in carrot roots. Mol. Genet. Genom. 2002, 268, 122–129. [Google Scholar] [CrossRef]
- Just, B.J.; Santos, C.A.; Yandell, B.S.; Simon, P.W. Major QTL for carrot color are positionally associated with carotenoid biosynthetic genes and interact epistatically in a domesticated x wild carrot cross. Theor. Appl. Genet. 2009, 119, 1155–1169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavagnaro, P.F.; Chung, S.-M.; Manin, S.; Yildiz, M.; Ali, A.; Alessandro, M.S.; Iorizzo, M.; Senalik, D.A.; Simon, P.W. Microsatellite isolation and marker development in carrot—Genomic distribution, linkage mapping, genetic diversity analysis and marker transferability across Apiaceae. BMC Genom. 2011, 12, 386. [Google Scholar] [CrossRef] [Green Version]
- Lu, S.; Van Eck, J.; Zhou, X.; Lopez, A.B.; O’Halloran, D.M.; Cosman, K.M.; Conlin, B.J.; Paolillo, D.J.; Garvin, D.F.; Vrebalov, J.; et al. The cauliflower Or gene encodes a DnaJ cysteine-rich domain-containing protein that mediates high levels of beta-carotene accumulation. Plant Cell 2006, 18, 3594–3605. [Google Scholar] [CrossRef] [Green Version]
- Karlin-Neumann, G.A.; Sun, L.; Tobin, E.M. Expression of light-harvesting chlorophyll a/b-protein Genes is phytochrome-regulated in etiolated Arabidopsis thaliana seedlings. Plant Physiol. 1988, 88, 1323–1331. [Google Scholar] [CrossRef] [Green Version]
- Qin, X.; Wang, W.; Wang, K.; Xin, Y.; Kuang, T. Isolation and characteristics of the PSI-LHCI-LHCII supercomplex under high light. Photochem. Photobiol. 2011, 87, 143–150. [Google Scholar] [CrossRef]
- Zhang, S.; Scheller, H.V. Light-harvesting complex II binds to several small subunits of photosystem I. J. Biol. Chem. 2004, 279, 3180–3187. [Google Scholar] [CrossRef] [Green Version]
- Hou, X.; Fu, A.; Garcia, V.J.; Buchanan, B.B.; Luan, S. PSB27: A thylakoid protein enabling Arabidopsis to adapt to changing light intensity. Proc. Natl. Acad. Sci. USA 2015, 112, 1613–1618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vermaas, W.F.; Williams, J.G.; Arntzen, C.J. Sequencing and modification of psbB, the gene encoding the CP-47 protein of Photosystem II, in the cyanobacterium Synechocystis 6803. Plant Mol. Biol. 1987, 8, 317–326. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | α-Carotene Content | β-Carotene Content | Lutein Content |
|---|---|---|---|
| DCAR_003942 | −0.204 | −0.355 | 0.732 ** |
| DCAR_003943 | 0.231 | 0.511 * | 0.469 |
| DCAR_005105 | 0.932 ** | 0.841 ** | 0.883 ** |
| DCAR_007169 | 0.681 * | 0.735 ** | −0.214 |
| DCAR_007901 | 0.696 * | 0.557 * | 0.015 |
| DCAR_007902 | 0.793 ** | 0.727 ** | 0.677 * |
| DCAR_009633 | 0.553 | 0.232 | 0.687 * |
| DCAR_009820 | 0.054 | 0.700 ** | 0.103 |
| DCAR_015960 | 0.526 | 0.685 ** | 0.762 ** |
| DCAR_018610 | 0.795 ** | 0.755 ** | 0.787 ** |
| DCAR_019192 | 0.850 ** | 0.691 ** | −0.885 ** |
| DCAR_023434 | 0.734 ** | 0.804 ** | 0.685 ** |
| DCAR_027950 | 0.952 ** | 0.783 ** | 0.448 |
| DCAR_027951 | 0.927 ** | 0.053 | 0.479 |
| DCAR_027952 | 0.798 ** | 0.769 ** | 0.573 |
| DCAR_029209 | 0.642 * | 0.720 ** | 0.369 |
| DCAR_029630 | 0.892 ** | 0.764 ** | −0.778 ** |
| DCAR_031498 | 0.720 ** | 0.742 ** | 0.901 ** |
| DCAR_032504 | 0.777 ** | 0.101 | 0.590 * |
| Gene Name | Gene ID | Chromosome | Mutation (wt→ym/Location) | Protein (wt→ym) | Annotation |
|---|---|---|---|---|---|
| LHCB1.3 | DCAR_005105 | 2 | T→\:5310926 | Ser→Gln | Chlorophyll a-b binding protein 1 |
| LHCB1.3 | DCAR_007902 | 2 | \→A:38079097; \→G:38079148 | Ser→Phe; His→Pro | Chlorophyll a-b binding protein 1 |
| PSAL | DCAR_018610 | 5 | \→T:31527204; \→A:31527207; T→\:31527877 | Leu→Thr; Val→Cys; Gln→Arg | Photosystem I reaction center subunit XI |
| PSB27-1 | DCAR_019192 | 5 | \→T:36996224; \→T:36996230; \→C:36996242; \→C:36996245 | Leu→Thr; Leu→Thr; Gln→Ala; *→Val | Photosystem II repair protein PSB27-H1 |
| psbB | DCAR_023434 | 7 | \→G:1473188; \→C:1473283 | Ser→Thr; Gln→Thr | Photosystem II CP47 protein |
| LHCB1.3 | DCAR_029630 | 9 | A→\: 9939156 | Ser→Leu | Chlorophyll a-b binding protein 1 |
| Lut5 | DCAR_023843 | 7 | \→G:6068378; \→C:6071826; \→C:6071862 | Cys→Met; Met→Ser; Ile→Ser | Protein Lutein deficient 5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Z.; Xu, H.; Yang, X.; Li, L.; Luo, D.; Liu, Z.; Jia, L. Transcriptome and Re-Sequencing Analyses Reveal Photosynthesis-Related Genes Involvement in Lutein Accumulation in Yellow Taproot Mutants of Carrot. Agronomy 2022, 12, 1866. https://doi.org/10.3390/agronomy12081866
Wu Z, Xu H, Yang X, Li L, Luo D, Liu Z, Jia L. Transcriptome and Re-Sequencing Analyses Reveal Photosynthesis-Related Genes Involvement in Lutein Accumulation in Yellow Taproot Mutants of Carrot. Agronomy. 2022; 12(8):1866. https://doi.org/10.3390/agronomy12081866
Chicago/Turabian StyleWu, Zhe, Hui Xu, Xuan Yang, Lixia Li, Dan Luo, Zhenzhen Liu, and Li Jia. 2022. "Transcriptome and Re-Sequencing Analyses Reveal Photosynthesis-Related Genes Involvement in Lutein Accumulation in Yellow Taproot Mutants of Carrot" Agronomy 12, no. 8: 1866. https://doi.org/10.3390/agronomy12081866