

Silencing CaTPS1 Increases the Sensitivity to Low Temperature and Salt Stresses in Pepper
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
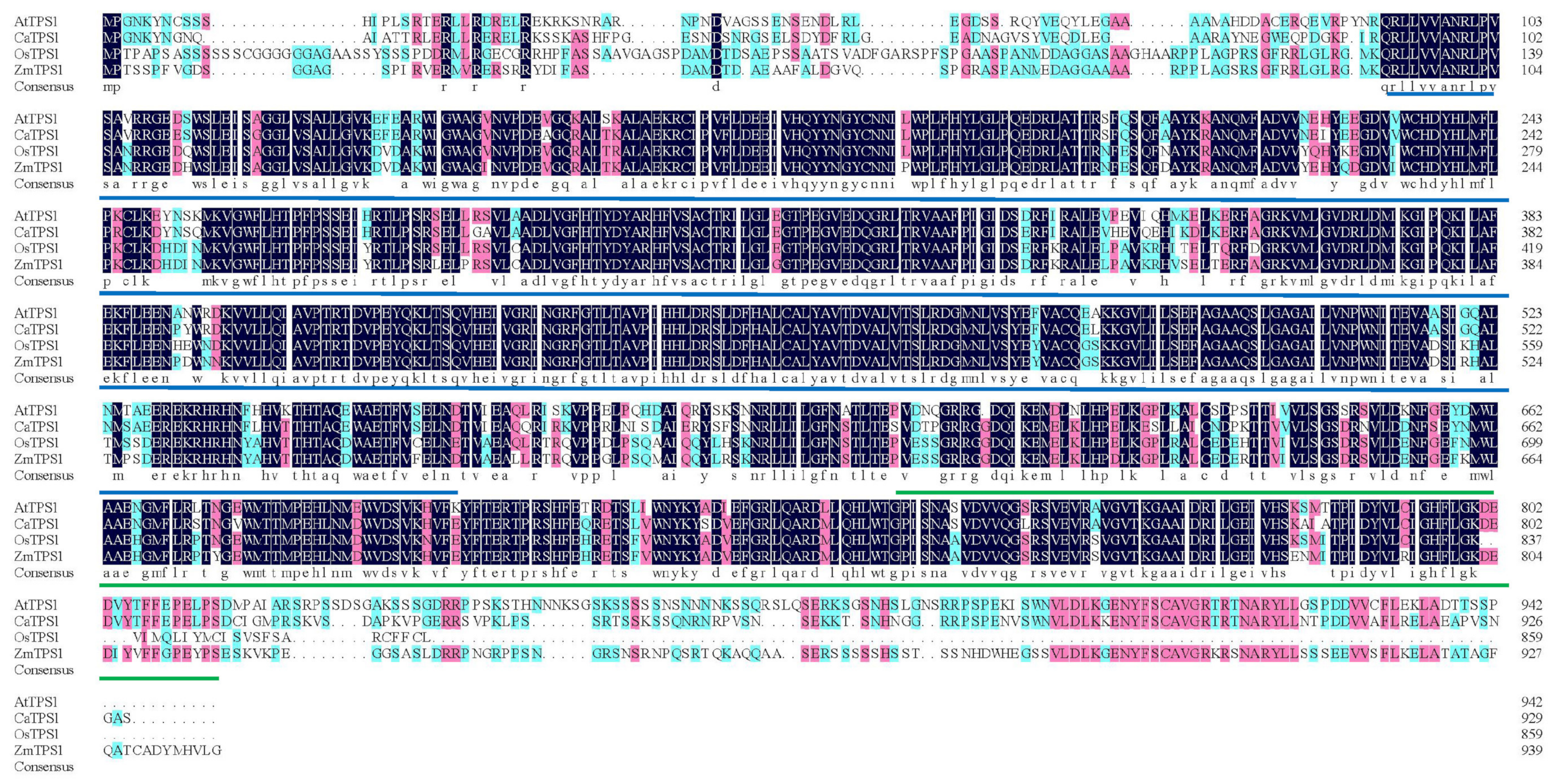
2.2. Homologous Sequence Alignment of CaTPS1
2.3. Construction of Silencing Vectors and Infection of Plants
2.4. The Plant Growth of Silenced Plants
2.5. RNA Isolation and qRT-PCR Analysis
2.6. Salt and Cold Stresses Assays
2.7. Determination of Chlorophyll and Trehalose Content, and Antioxidant Enzyme Activities
2.8. Statistical Analysis
3. Results
3.1. Homologous Sequence Alignment of CaTPS1
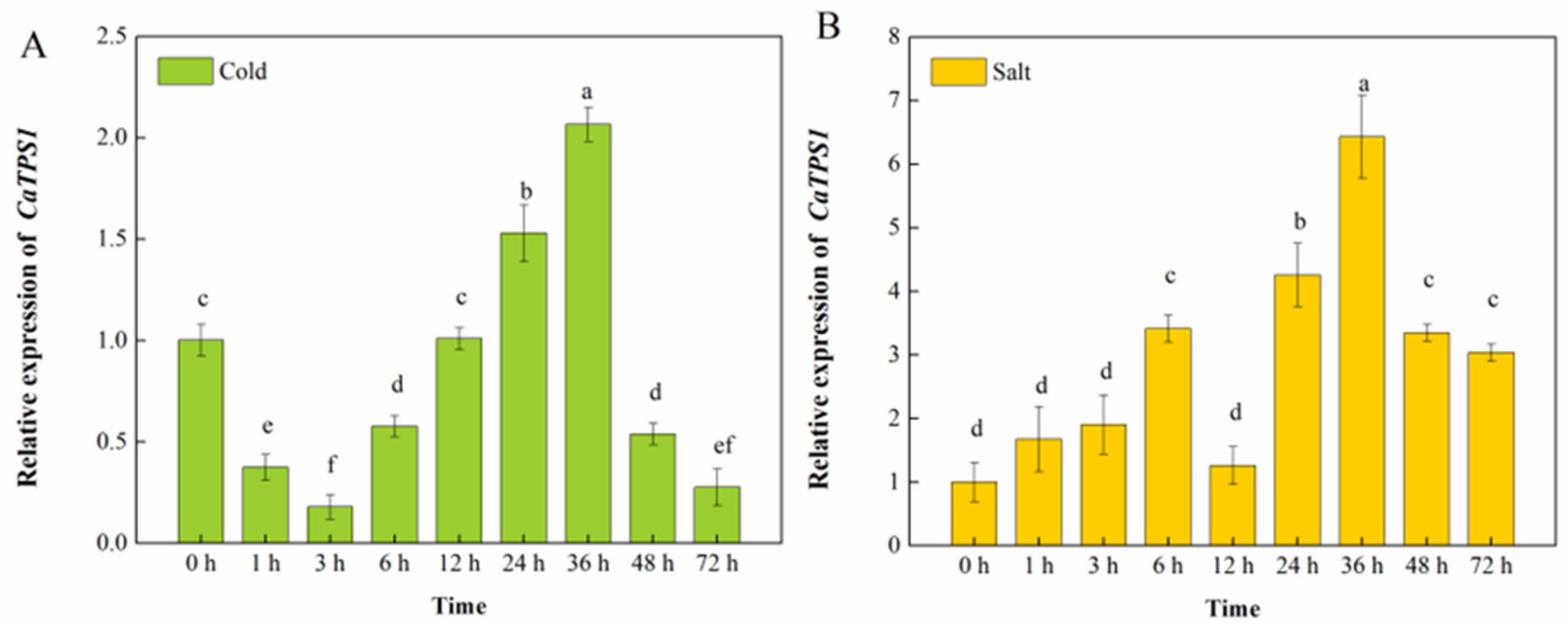
3.2. The Expression of CaTPS1 among Different Stress Treatments
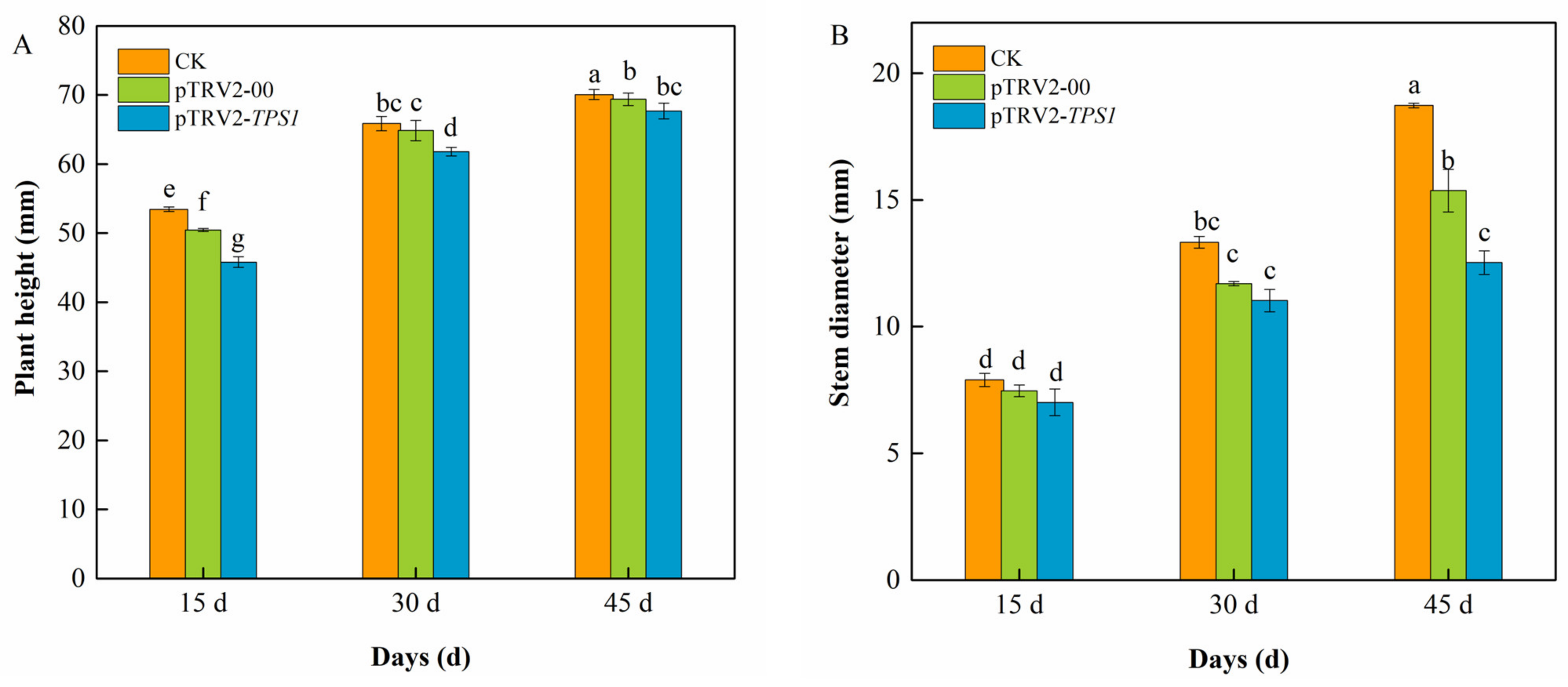
3.3. The Plant Growth of Silenced Plants
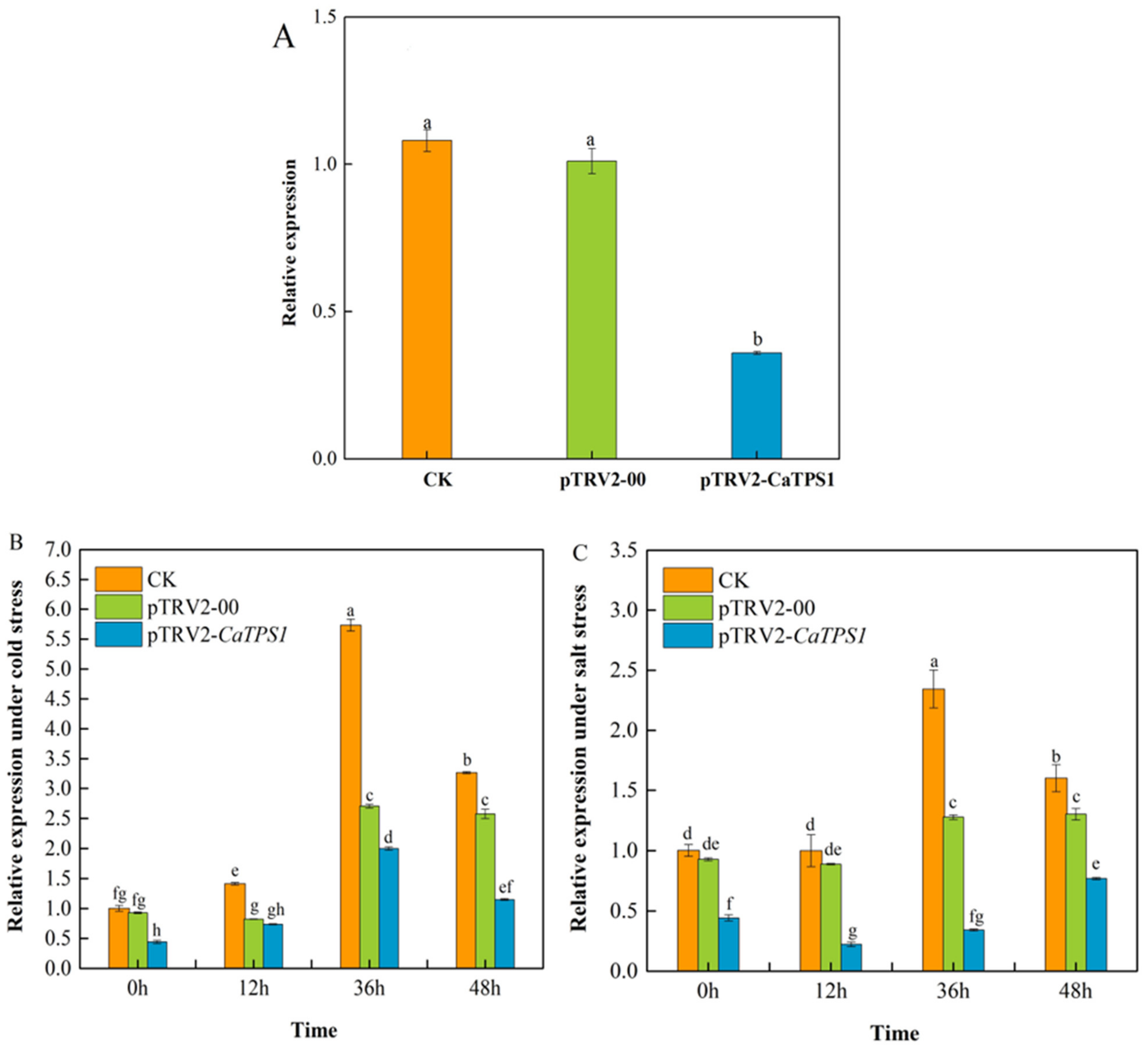
3.4. The Expression of CaTPS1 among Silenced Plants under Different Treatments
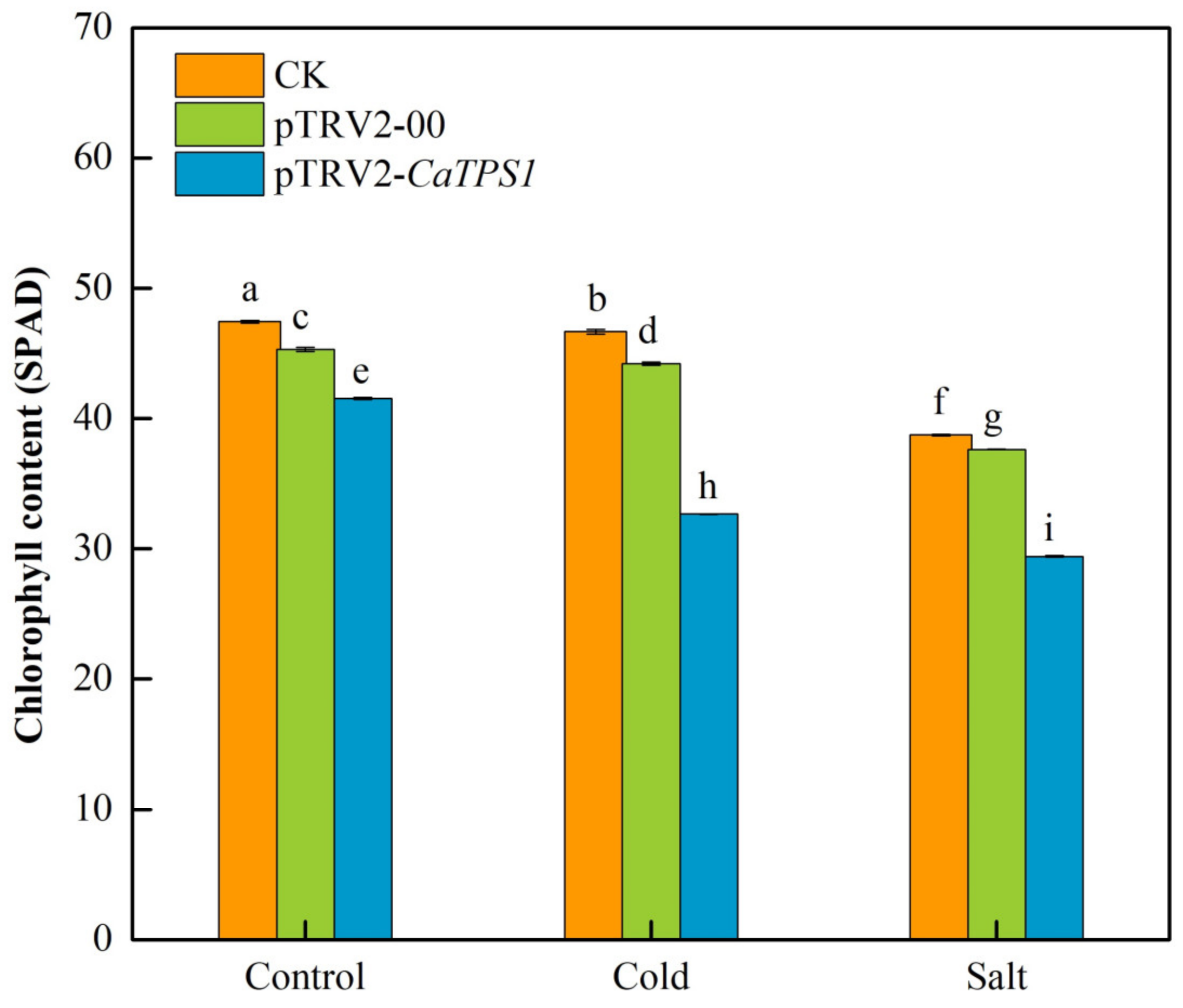
3.5. Chlorophyll Content of Silencing CaTPS1 under Cold and Salt Stresses
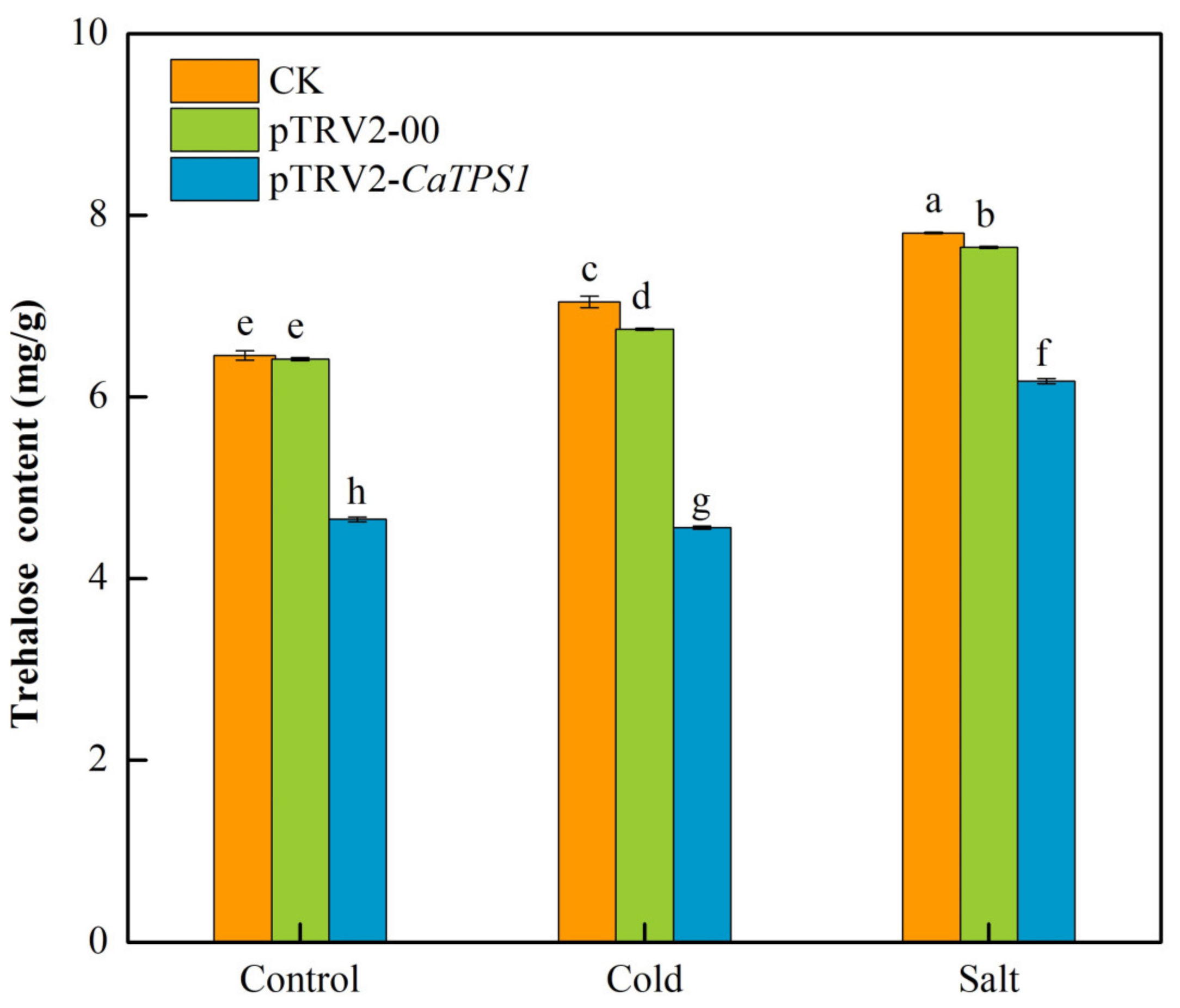
3.6. Trehalose Content of Silencing CaTPS1 under Cold and Salt Stresses
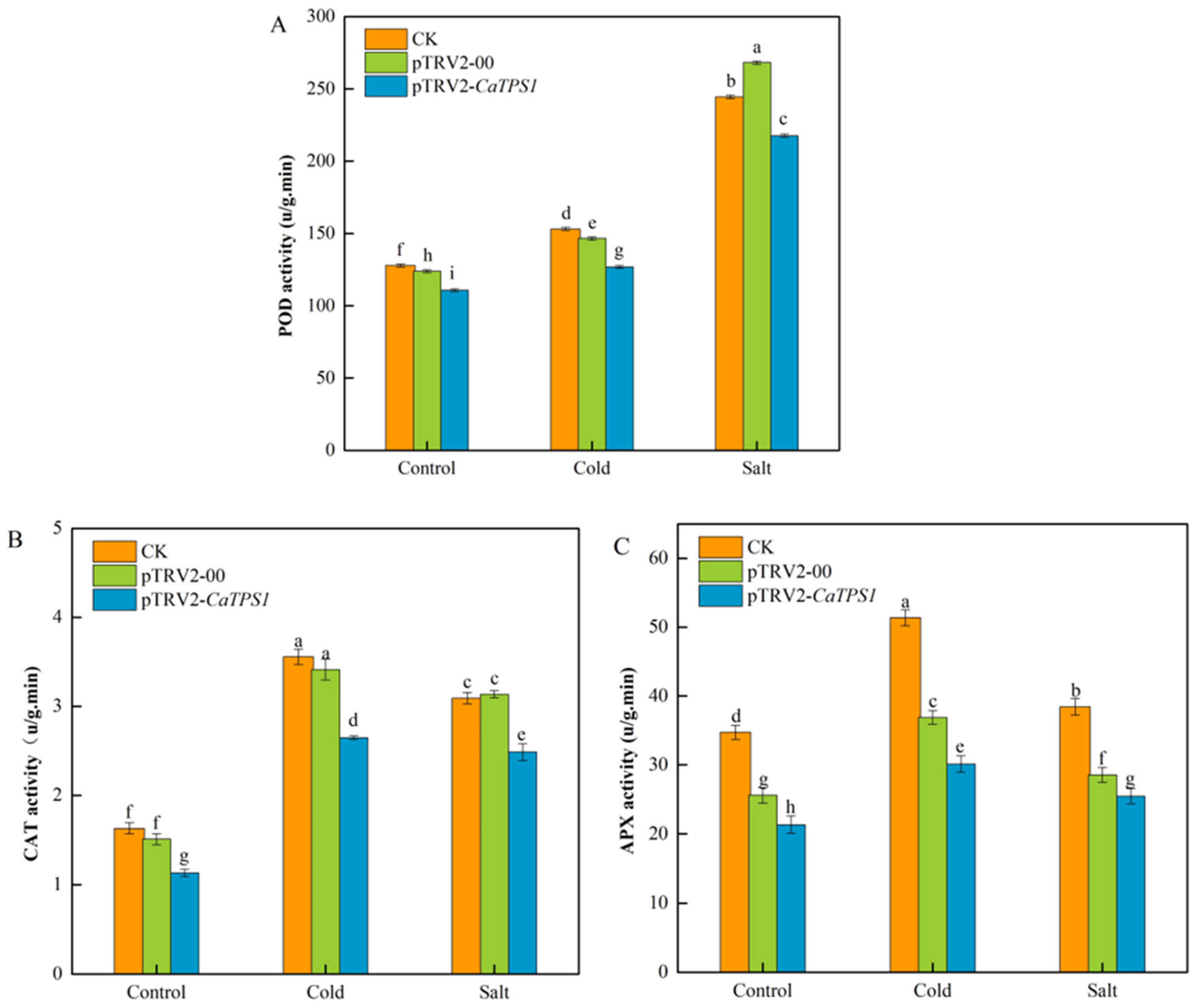
3.7. Antioxidant Enzymes of Silencing CaTPS1 under Cold and Salt Stresses
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Pino, J.; Gonzalez, M.; Ceballos, L.; Centurionyah, A.; Trujilloaguirre, J.; Latourneriemoreno, L.; Sauriduch, E. Characterization of total capsaicinoids, colour and volatile compounds of Habanero chilli pepper (Capsicum chinense Jack.) cultivars grown in Yucatan. Food Chem. 2007, 104, 1682–1686. [Google Scholar] [CrossRef]
- Pathirana, R. Peppers: Vegetable and Spice Capsicums, 2nd edition, by Paul W. Bosland and Eric J. Votava. N. Z. J. Crop Hortic. Sci. 2013, 41, 102–103. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.K. Abiotic Stress Signaling and Responses in Plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Genzel, F.; Dicke, M.D.; Junker-Frohn, L.V.; Neuwohner, A.; Thiele, B.; Putz, A.; Usadel, B.; Wormit, A.; Wiese-Klinkenberg, A. Impact of Moderate Cold and Salt Stress on the Accumulation of Antioxidant Flavonoids in the Leaves of Two Capsicum Cultivars. J. Agric. Food Chem. 2021, 69, 6431–6443. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Ding, Y.; Yang, Y.; Song, C.; Wang, B.; Yang, S.; Guo, Y.; Gong, Z. Protein kinases in plant responses to drought, salt, and cold stress. J. Integr. Plant Biol. 2021, 63, 53–78. [Google Scholar] [CrossRef] [PubMed]
- Trnka, M.; Rotter, R.P.; Ruiz-Ramos, M.; Kersebaum, K.C.; Olesen, J.E.; Zalud, Z.; Semenov, M.A. Adverse weather conditions for European wheat production will become more frequent with climate change. Nat. Clim. Change 2014, 4, 637–643. [Google Scholar] [CrossRef]
- Crimp, S.J.; Zheng, B.Y.; Khimashia, N.; Gobbett, D.L.; Chapman, S.; Howden, M.; Nicholls, N. Recent changes in southern Australian frost occurrence: Implications for wheat production risk. Crop Pasture Sci. 2016, 67, 801–811. [Google Scholar] [CrossRef]
- Vandesteene, L.; Lopez-Galvis, L.; Vanneste, K.; Feil, R.; Maere, S.; Lammens, W.; Rolland, F.; Lunn, J.E.; Avonce, N.; Beeckman, T.; et al. Expansive evolution of the trehalose-6-phosphate phosphatase gene family in Arabidopsis. Plant Physiol. 2012, 160, 884–896. [Google Scholar] [CrossRef] [Green Version]
- Wei, B.; Wang, L.; Zhang, R.; Chen, L.; Zhang, S.; Zhang, J. Identification of CaTPS Gene Family and Expression Analysis of CaTPS1 in Hot Pepper. Acta Hortic. Sin. 2016, 43, 1504–1512. (In Chinese) [Google Scholar]
- Nawaz, M.; Hassan, M.U.; Chattha, M.U.; Mahmood, A.; Shah, A.N.; Hashem, M.; Alamri, S.; Batool, M.; Rasheed, A.; Thabit, M.A.; et al. Trehalose: A promising osmo-protectant against salinity stress-physiological and molecular mechanisms and future prospective. Mol. Biol. Rep. 2022, 49, 11255–11271. [Google Scholar] [CrossRef]
- Elbein, A.D.; Pan, Y.T.; Pastuszak, I.; Carroll, D. New insights on trehalose: A multifunctional molecule. Glycobiology 2003, 13, 17R–27R. [Google Scholar] [CrossRef] [PubMed]
- Paul, M.J.; Oszvald, M.; Jesus, C.; Rajulu, C.; Griffiths, C.A. Increasing crop yield and resilience with trehalose 6-phosphate: Targeting a feast-famine mechanism in cereals for better source-sink optimization. J. Exp. Bot. 2017, 68, 4455–4462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pohane, A.A.; Carr, C.R.; Garhyan, J.; Swarts, B.M.; Siegrist, M.S. Trehalose Recycling Promotes Energy-Efficient Biosynthesis of the Mycobacterial Cell Envelope. Mbio 2021, 12, e02801-20. [Google Scholar] [CrossRef]
- Mu, M.; Lu, X.K.; Wang, J.J.; Wang, D.L.; Yin, Z.J.; Wang, S.; Fan, W.L.; Ye, W.W. Genome-wide Identification and analysis of the stress-resistance function of the TPS (Trehalose-6-Phosphate Synthase) gene family in cotton. BMC Genet. 2016, 17, 54. [Google Scholar]
- Liu, J.Y.; Huang, F.; Wang, X.; Zhang, M.; Zheng, R.; Wang, J.; Yu, D.Y. Genome-wide analysis of terpene synthases in soybean: Functional characterization of GmTPS3. Gene 2014, 544, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.C.; Wang, Y.J.; Mattson, N.; Yang, L.; Jin, Q.J. Genome-wide analysis of the Solanum tuberosum (potato) trehalose-6-phosphate synthase (TPS) gene family: Evolution and differential expression during development and stress. BMC Genom. 2017, 18, 926. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.L.; Liu, Y.J.; Wang, C.L.; Zeng, Q.Y. Molecular Evolution of Trehalose-6-Phosphate Synthase (TPS) Gene Family in Populus, Arabidopsis and Rice. PLoS ONE 2012, 7, e42438. [Google Scholar] [CrossRef] [PubMed]
- Zang, B.S.; Li, H.W.; Li, W.J.; Deng, X.W.; Wang, X.P. Analysis of trehalose-6-phosphate synthase (TPS) gene family suggests the formation of TPS complexes in rice. Plant Mol. Biol. 2011, 76, 507–522. [Google Scholar] [CrossRef]
- Vicente, R.L.; Spina, L.; Gomez, J.P.L.; Dejean, S.; Parrou, J.L.; Francois, J.M. Trehalose-6-phosphate promotes fermentation and glucose repression in Saccharomyces cerevisiae. Microb. Cell 2018, 5, 444–459. [Google Scholar] [CrossRef]
- Lin, M.; Jia, R.; Li, J.; Zhang, M.; Chen, H.; Zhang, D.; Zhang, J.; Chen, X. Evolution and expression patterns of the trehalose-6-phosphate synthase gene family in drumstick tree (Moringa oleifera Lam.). Planta 2018, 248, 999–1015. [Google Scholar] [CrossRef]
- Li, H.W.; Zang, B.S.; Deng, X.W.; Wang, X.P. Overexpression of the trehalose-6-phosphate synthase gene OsTPS1 enhances abiotic stress tolerance in rice. Planta 2011, 234, 1007–1018. [Google Scholar] [CrossRef]
- Stephen, K.; Beena, R.; Kiran, A.G.; Shanija, S.; Saravanan, R. Changes in physiological traits and expression of key genes involved in sugar signaling pathway in rice under high temperature stress. 3 Biotech 2022, 12, 183. [Google Scholar] [CrossRef] [PubMed]
- Stiller, I.; Dulai, S.; Kondrak, M.; Tarnai, R.; Szabo, L.; Toldi, O.; Banfalvi, Z. Effects of drought on water content and photosynthetic parameters in potato plants expressing the trehalose-6-phosphate synthase gene of Saccharomyces cerevisiae. Planta 2008, 227, 299–308. [Google Scholar] [CrossRef] [PubMed]
- Lyu, J.I.; Min, S.R.; Lee, J.H.; Lim, Y.H.; Kim, J.K.; Bae, C.H.; Liu, J.R. Overexpression of a trehalose-6-phosphate synthase/phosphatase fusion gene enhances tolerance and photosynthesis during drought and salt stress without growth aberrations in tomato. Plant Cell Tissue Org. Cult. 2013, 112, 257–262. [Google Scholar] [CrossRef]
- Xie, D.W.; Wang, X.N.; Fu, L.S.; Sun, J.; Zheng, W.; Li, Z.F. Identification of the trehalose-6-phosphate synthase gene family in winter wheat and expression analysis under conditions of freezing stress. J. Genet. 2015, 94, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Dai, Z.G.; Zhang, X.Y.; Tang, Q.; Cheng, C.H.; Liu, C.; Yu, Y.; Xu, G.C.; Xie, D.W.; Su, J.G. Identification of TPS and TPP gene families in Cannabis sativa and their expression under abiotic stresses. Biol. Plant. 2022, 66, 14–23. [Google Scholar] [CrossRef]
- Zhang, Z.Y.; Li, J.H.; Li, F.; Liu, H.H.; Yang, W.S.; Chong, K.; Xu, Y.Y. OsMAPK3 Phosphorylates OsbHLH002/OsICE1 and Inhibits Its Ubiquitination to Activate OsTPP1 and Enhances Rice Chilling Tolerance. Dev. Cell 2017, 43, 731–743.e5. [Google Scholar] [CrossRef] [Green Version]
- Xia, C.X.; Gong, Y.S.; Chong, K.; Xu, Y.Y. Phosphatase OsPP2C27 directly dephosphorylates OsMAPK3 and OsbHLH002 to negatively regulate cold tolerance in rice. Plant Cell Environ. 2021, 44, 491–505. [Google Scholar] [CrossRef]
- Godge, M.R.; Purkayastha, A.; Dasgupta, I.; Kumar, P.P. Virus-induced gene silencing for functional analysis of selected genes. Plant Cell Rep. 2008, 27, 209–219. [Google Scholar] [CrossRef]
- Tomar, M.; Sundaresha, S.; Singh, B.; Bhardwaj, V.; Sood, S.; Singh, B.; Salaria, N.; Thakur, K.; Kumar, A.; Sharma, N.; et al. Validation of molecular response of tuberization in response to elevated temperature by using a transient Virus Induced Gene Silencing (VIGS) in potato. Funct. Integr. Genom. 2021, 21, 215–229. [Google Scholar] [CrossRef]
- Zhang, H.J.; Zheng, D.W.; Yin, L.F.; Song, F.M.; Jiang, M. Functional Analysis of OsMED16 and OsMED25 in Response to Biotic and Abiotic Stresses in Rice. Front. Plant Sci. 2021, 12, 652453. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.X.; Guang, Y.L.; Wang, F.; Chen, Y.; Yang, W.T.; Xiao, X.F.; Luo, S.; Zhou, Y. Characterization of Phytochrome-Interacting Factor Genes in Pepper and Functional Analysis of CaPIF8 in Cold and Salt Stress. Front. Plant Sci. 2021, 12, 746517. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.N.; Mehari, T.G.; Xu, Y.C.; Umer, M.J.; Hou, Y.Q.; Wang, Y.H.; Peng, R.H.; Wang, K.B.; Cai, X.Y.; Zhou, Z.L.; et al. GhGLK1 a Key Candidate Gene From GARP Family Enhances Cold and Drought Stress Tolerance in Cotton. Front. Plant Sci. 2021, 12, 759312. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Fu, F.-L.; Zhang, S.-Z.; Wu, L.; Li, W.-C. Cloning and Characterization of Functional Trehalose-6-Phosphate Synthase Gene in Maize. J. Plant Biol. 2010, 53, 134–141. [Google Scholar] [CrossRef]
- Dong, Y.; Zhang, L.; Chang, X.; Wang, X.; Li, G.; Chen, S.; Jin, S. Overexpression of LpCPC from Lilium pumilum confers saline-alkali stress (NaHCO(3)) resistance. Plant Signal. Behav. 2022, 17, 2057723. [Google Scholar] [CrossRef]
- Letunic, I.; Khedkar, S.; Bork, P. SMART: Recent updates, new developments and status in 2020. Nucleic Acids Res. 2021, 49, D458–D460. [Google Scholar] [CrossRef]
- Guo, W.L.; Chen, R.G.; Gong, Z.H.; Yin, Y.X.; Li, D.W. Suppression Subtractive Hybridization Analysis of Genes Regulated by Application of Exogenous Abscisic Acid in Pepper Plant (Capsicum annuum L.) Leaves under Chilling Stress. PLoS ONE 2013, 8, e66667. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Araz, O.; Ekinci, M.; Yuce, M.; Shams, M.; Agar, G.; Yildirim, E. Low-temperature modified DNA methylation level, genome template stability, enzyme activity, and proline content in pepper (Capsicum annuum L.) genotypes. Sci. Hortic. Amst. 2022, 294, 110761. [Google Scholar] [CrossRef]
- Zhao, S.; Gou, B.; Wang, Y.; Yang, N.; Duan, P.; Wei, M.; Zhang, G.; Wei, B. Identification and relative expression analysis of CaFRK gene family in pepper. 3 Biotech 2022, 12, 137. [Google Scholar] [CrossRef]
- Pompelli, M.F.; Martins, S.C.; Celin, E.F.; Ventrella, M.C.; Damatta, F.M. What is the influence of ordinary epidermal cells and stomata on the leaf plasticity of coffee plants grown under full-sun and shady conditions? Braz. J. Biol. 2010, 70, 1083–1088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, K.; Nahakpam, S. Heat exposure alters the expression of SOD, POD, APX and CAT isozymes and mitigates low cadmium toxicity in seedlings of sensitive and tolerant rice cultivars. Plant Physiol. Biochem. 2012, 57, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Goulder, D.M.; Harte, F.M. Prevention of low-temperature gelation in milk protein concentrates by calcium-binding salts. J. Dairy Sci. 2022, 105, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Cha, S.J.; Park, H.J.; Kwon, S.J.; Lee, J.K.; Park, J.H. Early detection of plant stress using the internal electrical conductivity of Capsicum annuum in response to temperature and salinity stress. Plant Growth Regul. 2021, 95, 371–380. [Google Scholar] [CrossRef]
- Liu, L.; Li, H.; Li, N.; Li, S.X.; Guo, J.H.; Li, X.N. Parental salt priming improves the low temperature tolerance in wheat offspring via modulating the seed proteome. Plant Sci. 2022, 324, 111428. [Google Scholar] [CrossRef]
- Fichtner, F.; Olas, J.J.; Feil, R.; Watanabe, M.; Krause, U.; Hoefgen, R.; Stitt, M.; Lunn, J.E. Functional Features of TREHALOSE-6-PHOSPHATE SYNTHASE1, an Essential Enzyme in Arabidopsis. Plant Cell 2020, 32, 1949–1972. [Google Scholar] [CrossRef] [Green Version]
- Van Leemputte, F.; Vanthienen, W.; Wijnants, S.; Van Zeebroeck, G.; Thevelein, J.M. Aberrant Intracellular pH Regulation Limiting Glyceraldehyde-3-Phosphate Dehydrogenase Activity in the Glucose-Sensitive Yeast tps1 Delta Mutant. Mbio 2020, 11, e02199-20. [Google Scholar] [CrossRef]
- Zhang, H.F.; Liu, S.Y.; Ma, J.H.; Wang, X.K.; Haq, S.U.; Meng, Y.C.; Zhang, Y.M.; Chen, R.G. CaDHN4, a Salt and Cold Stress-Responsive Dehydrin Gene from Pepper Decreases Abscisic Acid Sensitivity in Arabidopsis. Int. J. Mol. Sci. 2019, 21, 26. [Google Scholar] [CrossRef] [Green Version]
- Gancedo, C.; Flores, C.L. The importance of a functional trehalose biosynthetic pathway for the life of yeasts and fungi. FEMS Yeast Res. 2004, 4, 351–359. [Google Scholar] [CrossRef] [Green Version]
- Avonce, N.; Mendoza-Vargas, A.; Morett, E.; Iturriaga, G. Insights on the evolution of trehalose biosynthesis. BMC Evol. Biol. 2006, 6, 109. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Usage | Primer Name | Primer Sequence(5′-3′) |
|---|---|---|
| pTRV2-CaTPS1 construction | V2-CaTPS1 | F: tgtgagtaaggttaccgaattcAACTTTTGTGAGTGAACTAAATGA |
| R: tgagctcggtaccggatccTACTCACTGAAGTTATCATCTAAG | ||
| qRT-PCR | q-CaTPS1 | F: TAAGTGGTGGAGGTCTCGTCAGC |
| R: ATTCACACCTGCCCAACCAATCC | ||
| qRT-PCR | q-Actin | F: CCCGGAAGAGCACCCTGTC |
| R: ATGCTGCTGGGAGCCAACG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gou, B.; Duan, P.; Wei, M.; Zhao, S.; Wang, Y.; Yang, N.; Zhang, G.; Wei, B. Silencing CaTPS1 Increases the Sensitivity to Low Temperature and Salt Stresses in Pepper. Agronomy 2023, 13, 319. https://doi.org/10.3390/agronomy13020319
Gou B, Duan P, Wei M, Zhao S, Wang Y, Yang N, Zhang G, Wei B. Silencing CaTPS1 Increases the Sensitivity to Low Temperature and Salt Stresses in Pepper. Agronomy. 2023; 13(2):319. https://doi.org/10.3390/agronomy13020319
Chicago/Turabian StyleGou, Bingdiao, Panpan Duan, Min Wei, Shufang Zhao, Yongfu Wang, Nan Yang, Gaoyuan Zhang, and Bingqiang Wei. 2023. "Silencing CaTPS1 Increases the Sensitivity to Low Temperature and Salt Stresses in Pepper" Agronomy 13, no. 2: 319. https://doi.org/10.3390/agronomy13020319