
Conversion of Thermal Energy to Gas Flow Kinetic Energy in the Bionic Leaf Stomata


Abstract
:1. Introduction
2. Materials and Methods
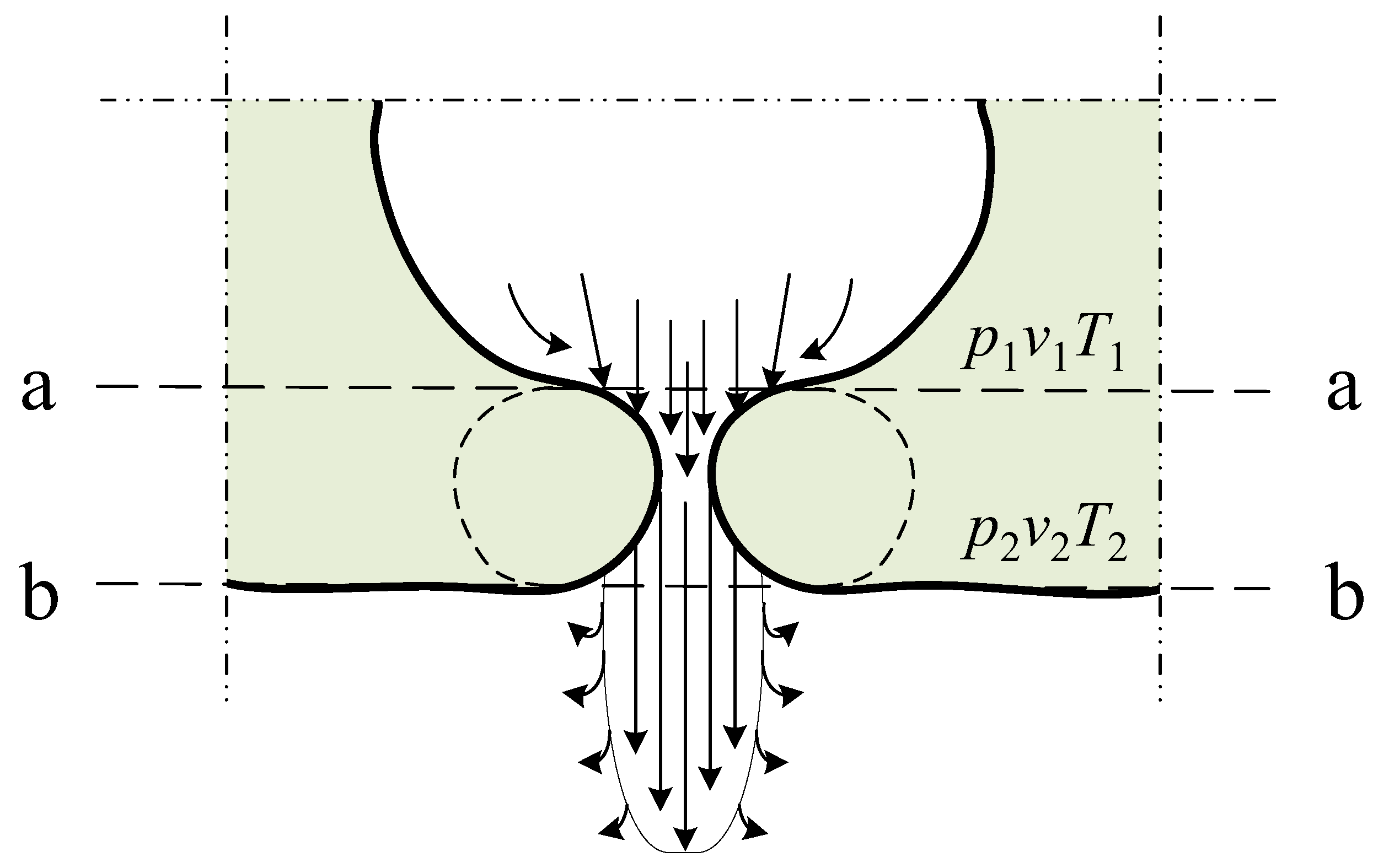
3. Theoretical Substantiation of Gas Flow in Leaf Stomata
- -
- The torque around the leaf suspension axis depends on the leaf surface area. Torque increases with a higher leaf surface area. Subsequently, the rotational speed of the leaf also increases.
- -
- Torque depends on the difference in gas pressures ∆p = p2 − p1. With a higher difference between the pressures in the leaf’s gas exchange system and the environment, the outgoing gas flow increases and the rotational force acting on the leaf also increases. Consequently, the rotational speed of the leaf depends on differential pressure ∆p = p2 − p1. Gas pressure p1 in the plant leaf stomata framework changes depending on the plant leaf temperature, i.e., the pressure increases at a higher temperature. Temperature pulsations in the plant leaf produce pressure pulsations in the gas exchange system of the plant leaf. Therefore, when the plant leaf is exposed to heat flow in the form of a light source, the leaf’s temperature increases and the pressure in the leaf’s gas exchange system also increases. With higher pressure, the gas flow displaced through the plant leaf stomata generates a higher reactive force on the illuminated side of the suspended leaf. The rotational speed of the leaf will also increase.
- -
- The velocity of the gas flow leaving the plant leaf stomata has a substantive effect on the rotational speed of the leaf. This factor alone cannot be evaluated as it is directly related and depends on ∆p = p2 − p1.
4. Results
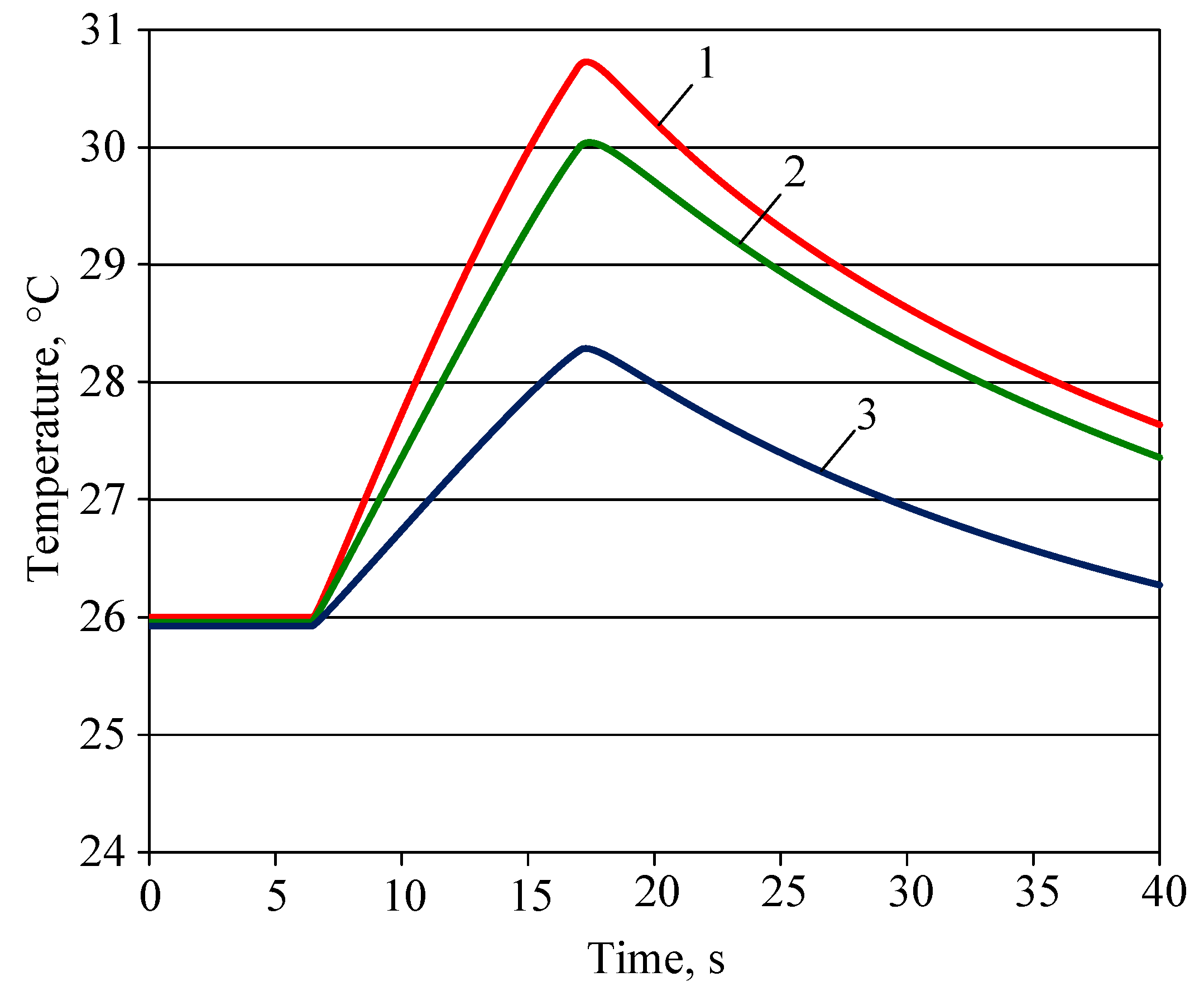
4.1. Variation of Plant Leaf Temperature for Lighting Using Different Light Sources
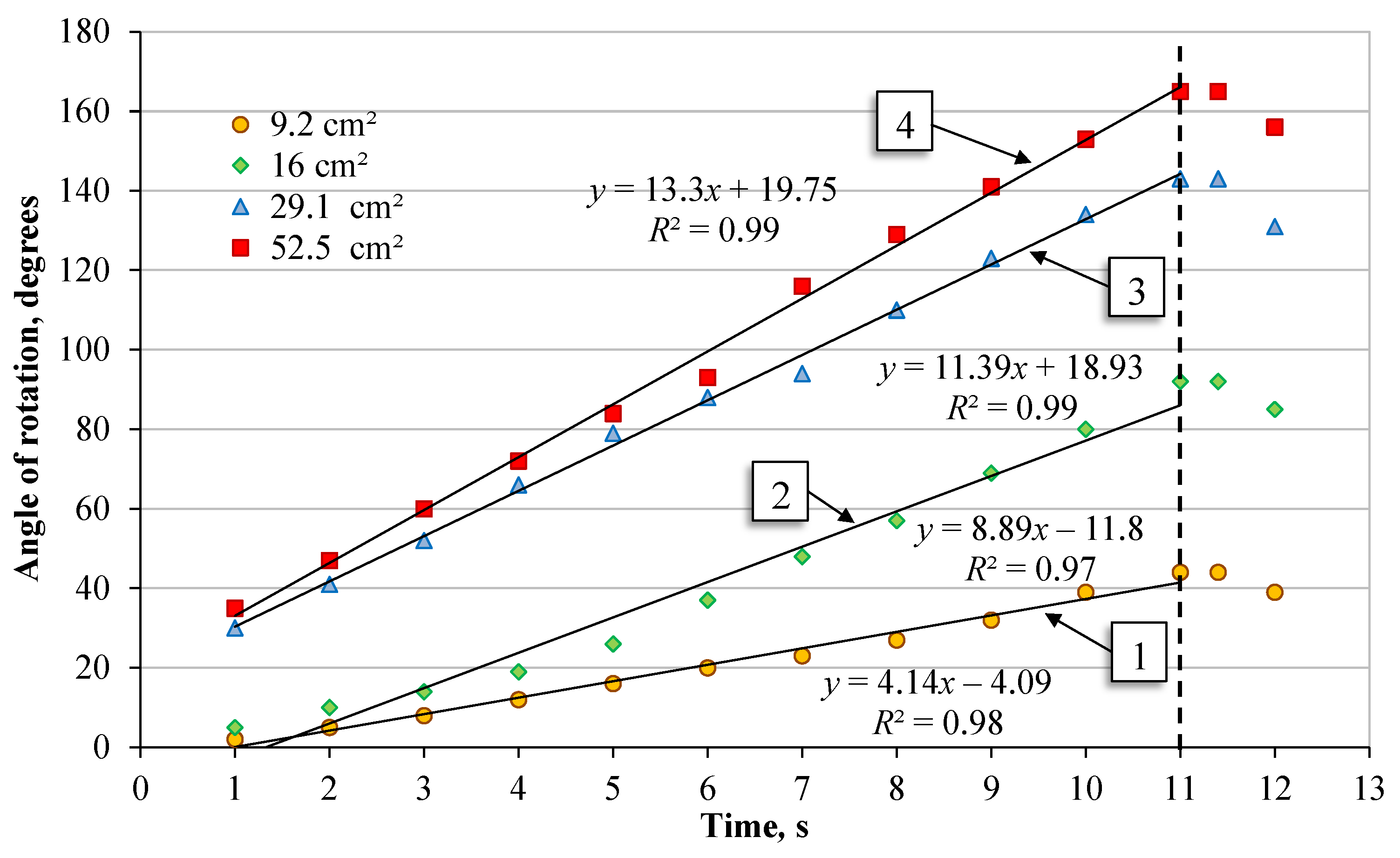
4.2. The Relationship between the Rotation of the Plant Leaf and the Leaf Blade Surface Area
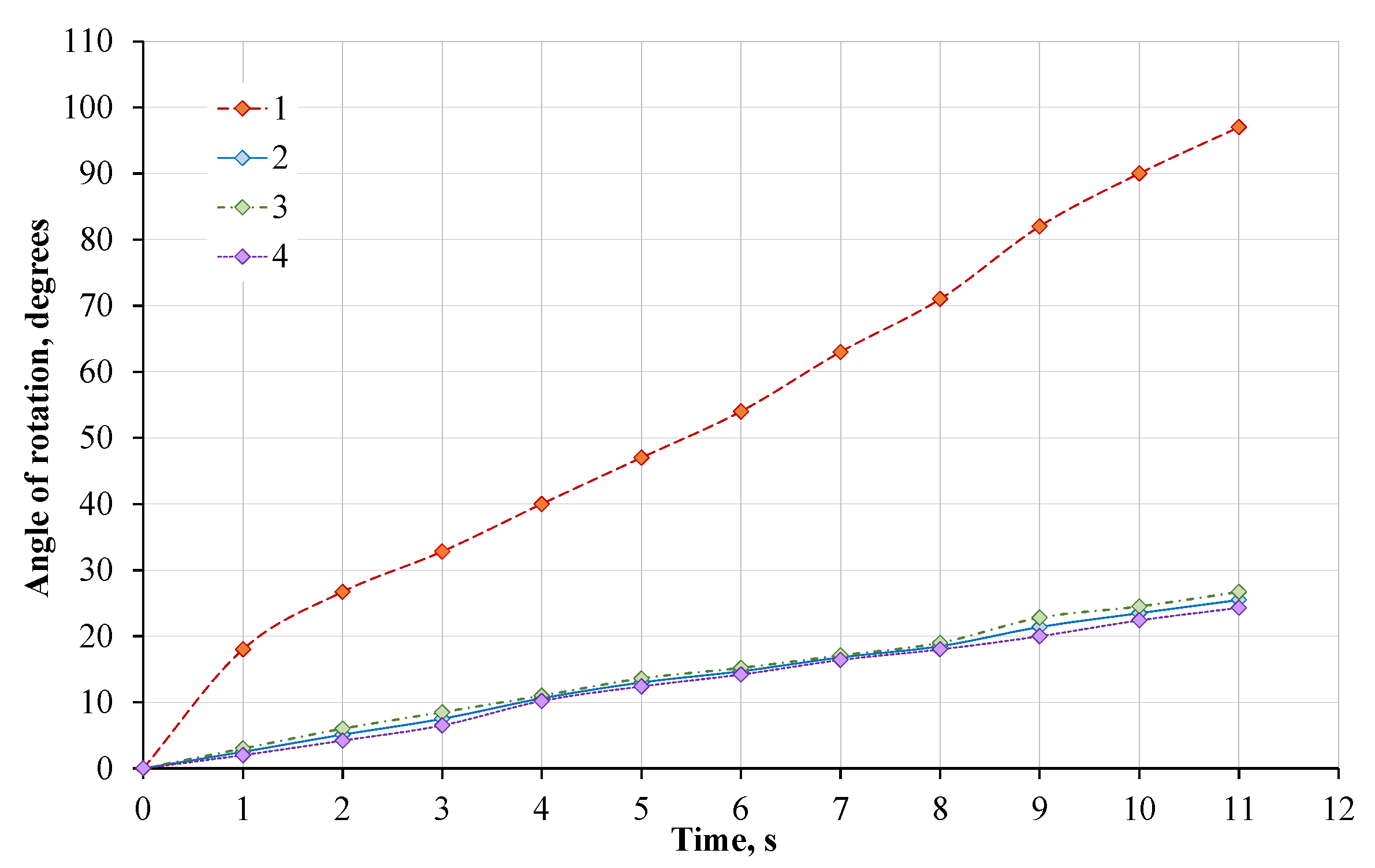
4.3. The Relationship between the Rotation of the Plant Leaf and the Power of Heat-Emitting Source
5. Discussion
6. Conclusions
- The presented experimental studies of the rotation of a suspended plant leaf (from the surface area of the plant leaf and the light flux given to the leaf) confirm the theoretical statements about the internal engine of the plant leaf, which activates the assimilation process with its surrounding environment.
- In a natural or artificial environment, the low-potential heat energy is converted into kinetic—mechanical energy of the gas flow in the petiole of the plant leaf. When the energy of the light flow is supplied to the plant leaf, the motor of the plant leaf begins to work, which intensifies the energy and gas exchange with the environment. This is shown by the observed difference between the live plant leaf and simulated plant leaf models, which ranged from 70.3° to 72.7° (72.5–74.9%).
- It was established that after providing the plant leaf with a light source flow (in the form of heat rays) with a power of 150 W and causing a temperature change in the tissues of the plant leaf, the hanging plant leaf rotated around the hanging axis, whereby a plant leaf with an area of 9.2 ± 0.6 cm2 had a rotation angle of 44°, 16.0 ± 0.8 cm2—92°, 29.1 ± 1.1 cm2—143° and 52.5 ± 1.9 cm2—165°. As the plant leaf area increased, so did the angular rotation speed, which ranged from 0.070–0.262 rad/s, respectively.
- By analyzing the influence of light sources on the rotation angle of a plant leaf, it was found that when illuminating a plant leaf with an active leaf area of 25.0 ± 1.0 cm2 with a 40 W light source, after 11 s, the rotation angle reaches 31°, with a 60 W light source it reaches 97°, with a 100 W light source it reaches 131° and with a 150 W light source it reaches 134°. When the power of the light source changes from 40 to 150 W, the angular rotation speed of the plant leaf also changes—0.049–0.213 rad/s.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sirvydas, P.A.; Kucinskas, V.; Kerpauskas, P.; Uksas, T. The Principle of Transforming the Solar Energy to Mechanical Energy in the Plant Leaf Stomata Engine. Energetika 2014, 60, 19–26. [Google Scholar]
- Kramer, P.J.; Boyer, J.S. Stomata and Gas Exchange. In Water Relations of Plants and Soils; Chapter 8; Academic Press, Inc.: Cambridge, MA, USA, 1995; pp. 257–282. [Google Scholar]
- Urban, J.; Ingwers, M.W.; McGuire, M.A.; Teskey, R.O. Increase in Leaf Temperature Opens Stomata and Decouples Net Photosynthesis from Stomatal Conductance in Pinus Taeda and Populus Deltoides X Nigra. J. Exp. Bot. 2017, 68, 1757–1767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Chen, G.; Dai, F.; Wang, Y.; Hills, A.; Ruan, Y.; Zhang, G.; Franks, P.J.; Nevo, E.; Blatt, M.R. Molecular Evolution of Grass Stomata. Trends Plant Sci. 2017, 22, 124–139. [Google Scholar] [CrossRef]
- Ye, H.; Yuan, Z.; Zhang, S. The Heat and Mass Transfer Analysis of a Leaf. J. Bionic Eng. 2013, 10, 170–176. [Google Scholar] [CrossRef]
- Yuan, Z.; Ye, H.; Li, S. Bionic Leaf Simulating the Thermal Effect of Natural Leaf Transpiration. J. Bionic Eng. 2014, 11, 90–97. [Google Scholar] [CrossRef]
- Yang, Y.; Liu, Z.; Hu, B.; Man, Y.; Wu, W. Bionic Composite Material Simulating the Optical Spectra of Plant Leaves. J. Bionic Eng. 2010, 7, S43–S49. [Google Scholar] [CrossRef]
- He, J.; Mao, M.; Li, D.; Liu, Y.; Jin, Z. Characterization of Leaf-Inspired Microfluidic Chips for Pumpless Fluid Transport. J. Bionic Eng. 2014, 11, 109–114. [Google Scholar] [CrossRef]
- Peng, Y.; Liu, W.; Chen, W.; Wang, N. A Conceptual Structure for Heat Transfer Imitating the Transporting Principle of Plant Leaf. Int. J. Heat Mass Transf. 2014, 71, 79–90. [Google Scholar] [CrossRef]
- Faisal, T.R.; Khalil Abad, E.M.; Hristozov, N.; Pasini, D. The Impact of Tissue Morphology, Cross-Section and Turgor Pressure on the Mechanical Properties of the Leaf Petiole in Plants. J. Bionic Eng. 2010, 7, S11–S23. [Google Scholar] [CrossRef]
- Wang, S.; Ren, L.; Liu, Y.; Han, Z.; Yang, Y. Mechanical Characteristics of Typical Plant Leaves. J. Bionic Eng. 2010, 7, 294–300. [Google Scholar] [CrossRef]
- Vialet-Chabrand, S.R.M.; Matthews, J.S.A.; McAusland, L.; Blatt, M.R.; Griffiths, H.; Lawson, T. Temporal Dynamics of Stomatal Behavior. Plant Physiol. 2017, 174, 603–613. [Google Scholar] [CrossRef] [Green Version]
- Matthews, J.S.A.; Vialet-Chabrand, S.; Lawson, T. Acclimation to Fluctuating Light Impacts the Rapidity of Response and Diurnal Rhythm of Stomatal Conductance. Plant Physiol. 2018, 176, 1939–1951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woolfenden, H.C.; Baillie, A.L.; Gray, J.E.; Hobbs, J.K.; Morris, R.J.; Fleming, A.J. Models and Mechanisms of Stomatal Mechanics. Trends Plant Sci. 2018, 23, 822–832. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, P.; Or, D. Effects of Stomata Clustering on Leaf Gas Exchange. New Phytol. 2015, 207, 1015–1025. [Google Scholar] [CrossRef] [PubMed]
- Dow, G.J.; Berry, J.A.; Bergmann, D.C. The Physiological Importance of Developmental Mechanisms that Enforce Proper Stomatal Spacing in Arabidopsis Thaliana. New Phytol. 2014, 201, 1205–1217. [Google Scholar] [CrossRef] [PubMed]
- Franks, P.J.; Berner, R.A.; Beerling, D.J. Maximum Leaf Conductance Driven by CO2 Effects on Stomatal Size and Density Over Geologic Time. Proc. Nat. Acad. Sci. USA 2009, 106, 10343–10347. [Google Scholar] [CrossRef] [Green Version]
- Zarinkamar, F. Stomatal Observations in Dicotyledons. Pak. J. Biol. Sci. 2007, 10, 199–219. [Google Scholar] [CrossRef] [Green Version]
- Drake, P.L.; Froend, R.H.; Franks, P.J. Smaller, Faster Stomata: Scaling of Stomatal Size, Rate of Response, and Stomatal Conductance. J. Exp. Bot. 2013, 64, 495–505. [Google Scholar] [CrossRef] [Green Version]
- Nobel, P.S. Physicochemical and Environmental Plant Physiology; Elsevier Science & Technology: London, UK, 2005. [Google Scholar]
- Šlapakauskas, V. Augalų Ekofiziologija; Lututė: Kaunas, Lithuania, 2006; p. 413. [Google Scholar]
- Düring, H. Stomatal and Mesophyll Conductances Control CO2 Transfer to Chloroplasts in Leaves of Grapevine (Vitis Vinifera L.). Vitis Geilweilerhof 2003, 42, 65–68. [Google Scholar]
- Voelker, S.L.; Brooks, J.R.; Meinzer, F.C.; Anderson, R.; Bader, M.K.-F.; Battipaglia, G.; Becklin, K.M.; Beerling, D.; Bert, D.; Betancourt, J.L.; et al. A Dynamic Leaf Gas-Exchange Strategy is Conserved in Woody Plants Under Changing Ambient CO2: Evidence from Carbon Isotope Discrimination in Paleo and CO2 Enrichment Studies. Glob. Chang. Biol. 2016, 22, 889–902. [Google Scholar] [CrossRef]
- Ache, P.; Bauer, H.; Kollist, H.; Al-Rasheid, K.A.S.; Lautner, S.; Hartung, W.; Hedrich, R. Stomatal Action Directly Feeds Back on Leaf Turgor: New Insights into the Regulation of the Plant Water Status from Non-invasive Pressure Probe Measurements. Plant J. Cell Mol. Biol. 2010, 62, 1072–1082. [Google Scholar] [CrossRef] [PubMed]
- Rahnama, A.; James, R.A.; Poustini, K.; Munns, R. Stomatal Conductance as a Screen for Osmotic Stress Tolerance in Durum Wheat Growing in Saline Soil. Funct. Plant Biol. FPB 2010, 37, 255. [Google Scholar] [CrossRef]
- Cano, F.J.; Sanchez-Gomez, D.; Rodrigues-Calcerrada, J.; Warren, C.R.; Gil, L.; Ismael, A. Effects of Drought on Mesophyll Conductance and Photosynthetic Limitations at Different Tree Canopy Layers. Plant Cell Environ. 2013, 36, 1961–1980. [Google Scholar] [CrossRef] [PubMed]
- Fanourakis, D.; Bouranis, D.; Giday, H.; Carvalho, D.R.A.; Rezaei Nejad, A.; Ottosen, C. Improving Stomatal Functioning at Elevated Growth Air Humidity: A Review. J. Plant Physiol. 2016, 207, 51–60. [Google Scholar] [CrossRef]
- Varela, S.A.; Weigandt, M.N.; Willems, P.; Bianchi, E.; Diez, J.P.; Gyenge, J.E. Physiological Status of Conifer Seedlings Treated with Radiation, Drought and Frost Stress Mitigation Techniques: A Laboratory Assessment. New For. 2015, 47, 87–103. [Google Scholar] [CrossRef]
- Kim, H.; Goins, G.D.; Wheeler, R.M.; Sager, J.C. Stomatal Conductance of Lettuce Grown Under or Exposed to Different Light Qualities. Ann. Bot. 2004, 94, 691–697. [Google Scholar] [CrossRef] [Green Version]
- Hedrich, R.; Shabala, S. Stomata in a Saline World. Curr. Opin. Plant Biol. 2018, 46, 87–95. [Google Scholar] [CrossRef]
- Jarvis, P.G. Stomatal Physiology; Cambridge University. Press: Cambridge, UK, 1981. [Google Scholar]
- Willmer, C.; Fricker, M. Stomata; Springer: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Lambers, H.; Oliveira, R.S. Plant Physiological Ecology, 3rd ed.; Springer: Cham, Switzerland, 2019. [Google Scholar]
- Franks, P.J.; Farquhar, G.D. The Effect of Exogenous Abscisic Acid on Stomatal Development, Stomatal Mechanics, and Leaf Gas Exchange in Tradescantia Virginiana. Plant Physiol. 2001, 125, 935–942. [Google Scholar] [CrossRef] [Green Version]
- Boer, H.J.; Price, C.A.; Wagner-Cremer, F.; Dekker, S.C.; Franks, P.J.; Veneklaas, E.J. Optimal Allocation of Leaf Epidermal Area for Gas Exchange. New Phytol. 2016, 210, 1219–1228. [Google Scholar] [CrossRef]
- Švenčianas, P.; Adomavičius, A. Inžinerinė Termodinamika; Technologija: Kaunas, Lithuania, 2011; p. 308. [Google Scholar]
- Hetherington, A.M.; Woodward, F.I. The Role of Stomata in Sensing and Driving Environmental Change. Nature 2003, 424, 901–908. [Google Scholar] [CrossRef]
- Shimazaki, K.; Doi, M.; Assmann, S.M.; Kinoshita, T. Light Regulation of Stomatal Movement. Ann. Rev. Plant Biol. 2007, 58, 219–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harrison, E.L.; Arce Cubas, L.; Gray, J.E.; Hepworth, C. The Influence of Stomatal Morphology and Distribution on Photosynthetic Gas Exchange. Plant J. 2020, 101, 768–779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Planck, M. Treatise on Thermodynamics; Dover Publications: Dover, UK, 1987. [Google Scholar]
- Moran, M.J. Fundamentals of Engineering Thermodynamics, 8th [revised] ed.; Wiley: Hoboken, NJ, USA, 2014. [Google Scholar]
- Drobavičius, A.; Garbavičius, J.; Gimbutis, G.S.; Kajutis, K.; Krukonis, V. Bendroji Šiluminė Technika; Mintis: Vilnius, Lithuania, 1974; p. 572. [Google Scholar]
- Von Caemmerer, S.; Farquhar, G.D. Some Relationships between the Biochemistry of Photosynthesis and the Gas Exchange of Leaves. Planta 1981, 153, 376–387. [Google Scholar] [CrossRef]
- Ziegler, H. The evolution of stomata. In Stomatal Function; Zeiger, E., Farquhar, G.D., Cowan, I.R., Eds.; Stanford University Press: Stanford, CA, USA, 1987; pp. 29–57. [Google Scholar]
- Franks, P.J.; Farquhar, G.D. The Mechanical Diversity of Stomata and its Significance in Gas-Exchange Control. Plant Physiol. 2007, 143, 78–87. [Google Scholar] [CrossRef] [Green Version]
- Gimbutis, G.; Kajutis, K.; Krukonis, V. Šiluminė Technika; Mokslas: Vilnius, Lithuanisa, 1993. [Google Scholar]
- Van Wylen, G.J.; Sonntag, R.E. Fundamentals of Classical Thermodynamics, 2ed. ed.; Wiley: New York, NY, USA, 1973. [Google Scholar]
- Martinaitis, V. Techninė Termodinamika ir Šilumokaita; Technika: Vilnius, Lithuania, 2003; p. 163. [Google Scholar]
- Baehr, H.D. Thermodynamik, 3rd ed.; Springer: Berlin/Heidelberg, Germany, 1973; p. 440. [Google Scholar]
- Kleshnin, A.F. Die Pflanze und das Licht. Neu bearbeitete und ergänzte Ausgabe; Akademie–Verlag: Berlin, Germany, 1960; p. 620. [Google Scholar]
- Merlot, S.; Mustilli, A.; Genty, B.; North, H.; Lefebvre, V.; Sotta, B.; Vavasseur, A.; Giraudat, J. Use of Infrared Thermal Imaging to Isolate Arabidopsis Mutants Defective in Stomatal Regulation. Plant J. Cell Mol. Biol 2002, 30, 601–609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daszkowska-Golec, A.; Szarejko, I. Open or Close the Gate—Stomata Action Under the Control of Phytohormones in Drought Stress Conditions. Front. Plant Sci. 2013, 4, 138. [Google Scholar] [CrossRef] [Green Version]
- Meinzer, F.C.; McCulloh, K.A. Xylem Recovery from Drought-Induced Embolism: Where is the Hydraulic Point of no Return? Tree Physiol. 2013, 33, 331–334. [Google Scholar] [CrossRef] [Green Version]
- Brodribb, T.J.; McAdam, S.A.M.; Jordan, G.J.; Martins, S.C.V. Conifer Species Adapt to Low-Rainfall Climates by Following One of Two Divergent Pathways. Proc. Nat. Acad. Sci. USA 2014, 111, 14489–14493. [Google Scholar] [CrossRef] [Green Version]
- Schymanski, S.J.; Or, D.; Zwieniecki, M. Stomatal Control and Leaf Thermal and Hydraulic Capacitances Under Rapid Environmental Fluctuations. PLoS ONE 2013, 8, e54231. [Google Scholar] [CrossRef]
- Page, G.F.M.; Liénard, J.F.; Pruett, M.J.; Moffett, K.B. Spatiotemporal Dynamics of Leaf Transpiration Quantified with Time-Series Thermal Imaging. Agric. For. Meteorol. 2018, 256–257, 304–314. [Google Scholar] [CrossRef]
- Berry, J.; Bjorkman, O. Photosynthetic Response and Adaptation to Temperature in Higher Plants. Ann. Rev. Plant Physiol. 1980, 31, 491–543. [Google Scholar] [CrossRef]
- Wang, L.; Good, S.P.; Caylor, K.K. Global Synthesis of Vegetation Control on Evapotranspiration Partitioning. Geophys. Res. Lett. 2014, 41, 6753–6757. [Google Scholar] [CrossRef]
- Wang, R.; Yu, G.; He, N.; Wang, Q.; Zhao, N.; Xu, Z.; Ge, J. Latitudinal Variation of Leaf Stomatal Traits from Species to Community Level in Forests: Linkage with Ecosystem Productivity. Sci. Rep. 2015, 5, 14454. [Google Scholar] [CrossRef]
- Assmann, S.M.; Wang, X. From Milliseconds to Millions of Years: Guard Cells and Environmental Responses. Curr. Opin. Plant Biol. 2001, 4, 421–428. [Google Scholar] [CrossRef]
- Dubbert, M.; Piayda, A.; Cuntz, M.; Correia, A.C.; Costa E Silva, F.; Pereira, J.S.; Werner, C. Stable Oxygen Isotope and Flux Partitioning Demonstrates Understory of an Oak Savanna Contributes Up to Half of Ecosystem Carbon and Water Exchange. Front. Plant Sci. 2014, 5, 530. [Google Scholar] [CrossRef] [PubMed]
- Chazdon, R.L.; Pearcy, R.W. The Importance of Sunflecks for Forest Understory Plants. Bioscience 1991, 41, 760–766. [Google Scholar] [CrossRef]
- Way, D.A.; Pearcy, R.W. Sunflecks in Trees and Forests: From Photosynthetic Physiology to Global Change Biology. Tree Physiol. 2012, 32, 1066–1081. [Google Scholar] [CrossRef] [Green Version]
- Hartt, C.E.; Kortschak, H.P. Translocation of 14C in the Sugarcane Plant during the Day and Night. Plant Physiol. 1967, 42, 89–94. [Google Scholar] [CrossRef] [Green Version]
- Blonder, B.; Michaletz, S.T. A Model for Leaf Temperature Decoupling from Air Temperature. Agric. For. Meteorol. 2018, 262, 354–360. [Google Scholar] [CrossRef]
- Cerasoli, S.; Wertin, T.; McGuire, M.A.; Rodrigues, A.; Aubrey, D.P.; Pereira, J.S.; Teskey, R.O. Poplar Saplings Exposed to Recurring Temperature Shifts of Different Amplitude Exhibit Differences in Leaf Gas Exchange and Growth Despite Equal Mean Temperature. AoB Plants 2014, 6, 2014. [Google Scholar] [CrossRef] [Green Version]
- von Caemmerer, S.; Evans, J.R. Temperature Responses of Mesophyll Conductance Differ Greatly between Species. Plant Cell Environ. 2015, 38, 629–637. [Google Scholar] [CrossRef] [PubMed]
- Weston, D.J.; Bauerle, W.L. Inhibition and Acclimation of C(3) Photosynthesis to Moderate Heat: A Perspective from Thermally Contrasting Genotypes of Acer Rubrum (Red Maple). Tree Physiol. 2007, 27, 1083–1092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lahr, E.C.; Schade, G.W.; Crossett, C.C.; Watson, M.R. Photosynthesis and Isoprene Emission from Trees Along an Urban-Rural Gradient in Texas. Glob. Chang. Biol. 2015, 21, 4221–4236. [Google Scholar] [CrossRef]
- Leakey, A.D.B.; Scholes, J.D.; Press, M.C. Physiological and Ecological Significance of Sunflecks for Dipterocarp Seedlings. J. Exp. Bot. 2005, 56, 469–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaňa, R.; Vass, I. Thermoimaging as a Tool for Studying Light-Induced Heating of Leaves Correlation of Heat Dissipation with the Efficiency of Photosystem II Photochemistry and Non-Photochemical Quenching. Environ. Exp. Bot. 2008, 64, 90–96. [Google Scholar] [CrossRef]
- Vialet-Chabrand, S.; Lawson, T. Dynamic Leaf Energy Balance: Deriving Stomatal Conductance from Thermal Imaging in a Dynamic Environment. J. Exp. Bot. 2019, 70, 2839–2855. [Google Scholar] [CrossRef] [Green Version]
- Bajons, P.; Klinger, G.; Schlosser, V. Determination of Stomatal Conductance by Means of Infrared Thermography. Infrared Phys. Technol. 2005, 46, 429–439. [Google Scholar] [CrossRef]
- Karlsson, P.E.; Assmann, S.M. Rapid and Specific Modulation of Stomatal Conductance by Blue Light in Ivy (Hedera Helix): An Approach to Assess the Stomatal Limitation of Carbon Assimilation. Plant Physiol. 1990, 94, 440–447. [Google Scholar] [CrossRef] [Green Version]
- Grantz, D.A.; Assmann, S.M. Stomatal Response to Blue Light: Water use Efficiency in Sugarcane and Soybean. Plant Cell Environ. 1991, 14, 683–690. [Google Scholar] [CrossRef]
- Assmann, S.M.; Lee, D.M.; Malkus, P. Rapid Stomatal Response to Redlight in Zea Mays. Photochem. Photobiol. 1992, 56, 685–689. [Google Scholar] [CrossRef]
- Lawson, T.; von Caemmerer, S.; Baroli, I. Photosynthesis and Stomatal Behaviour. In Progress in Botany 72; Springer: Berlin/Heidelberg, Germany, 2010; pp. 265–304. [Google Scholar]
- Raven, J.A. Selection Pressures on Stomatal Evolution. New Phytol. 2002, 153, 371–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berry, J.A.; Beerling, D.J.; Franks, P.J. Stomata: Key Players in the Earth System, Past and Present. Curr. Opin. Plant Biol. 2010, 13, 232–239. [Google Scholar] [CrossRef] [PubMed]
- De Boer, H.J.; Lammertsma, E.I.; Wagner-Cremer, F.; Dilcher, D.L.; Wassen, M.J.; Dekker, S.C. Climate Forcing due to Optimization of Maximal Leaf Conductance in Subtropical Vegetation Under Rising CO. Proc. Nat. Acad. Sci. USA 2011, 108, 4041–4046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Assouline, S.; Or, D. Plant Water use Efficiency Over Geological Time—Evolution of Leaf Stomata Configurations Affecting Plant Gas Exchange. PLoS ONE 2013, 8, e67757. [Google Scholar] [CrossRef]
- Buckley, T.N.; Schymanski, S.J. Stomatal Optimisation in Relation to Atmospheric CO2. New Phytol. 2014, 201, 372–377. [Google Scholar] [CrossRef]
- Woolley, J.T. Reflectance and Transmittance of Light by Leaves. Plant Physiol. 1971, 47, 656–662. [Google Scholar] [CrossRef] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Light source power, W | 150 | 100 | 60 | 40 |
| Energy content under normal conditions W/m2 | 1135.9 | 1011.8 | 330.6 | 181.8 |
| Energy content under normal conditions W/cm2 | 0.114 | 0.101 | 0.033 | 0.018 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ūksas, T.; Sirvydas, P.A.; Paulikienė, S.; Čingienė, R. Conversion of Thermal Energy to Gas Flow Kinetic Energy in the Bionic Leaf Stomata. Agronomy 2022, 12, 1742. https://doi.org/10.3390/agronomy12081742
Ūksas T, Sirvydas PA, Paulikienė S, Čingienė R. Conversion of Thermal Energy to Gas Flow Kinetic Energy in the Bionic Leaf Stomata. Agronomy. 2022; 12(8):1742. https://doi.org/10.3390/agronomy12081742
Chicago/Turabian StyleŪksas, Tomas, Povilas Algimantas Sirvydas, Simona Paulikienė, and Rasa Čingienė. 2022. "Conversion of Thermal Energy to Gas Flow Kinetic Energy in the Bionic Leaf Stomata" Agronomy 12, no. 8: 1742. https://doi.org/10.3390/agronomy12081742