
The Application of Tomato Plant Residue Compost and Plant Growth-Promoting Rhizobacteria Improves Soil Quality and Enhances the Ginger Field Soil Bacterial Community

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Test Field Conditions
2.2. Experimental Materials
2.3. Experimental Design
2.4. Soil Sample Collection
2.5. Soil Nutrient Content and Enzyme Activity Determination
2.6. Soil DNA Extraction, 16S rRNA Gene Amplicon Sequencing
2.7. Soil Microbial Community Analysis
2.8. Data Analysis
3. Results and Analyses
3.1. Effect of Applying Tomato Plant Residue Compost and PGPR on the Nutrient Content of Ginger Field Soil
3.2. Effect of Applying Tomato Plant Residue Compost and PGPR on the Enzyme Activity of Ginger Field Soil
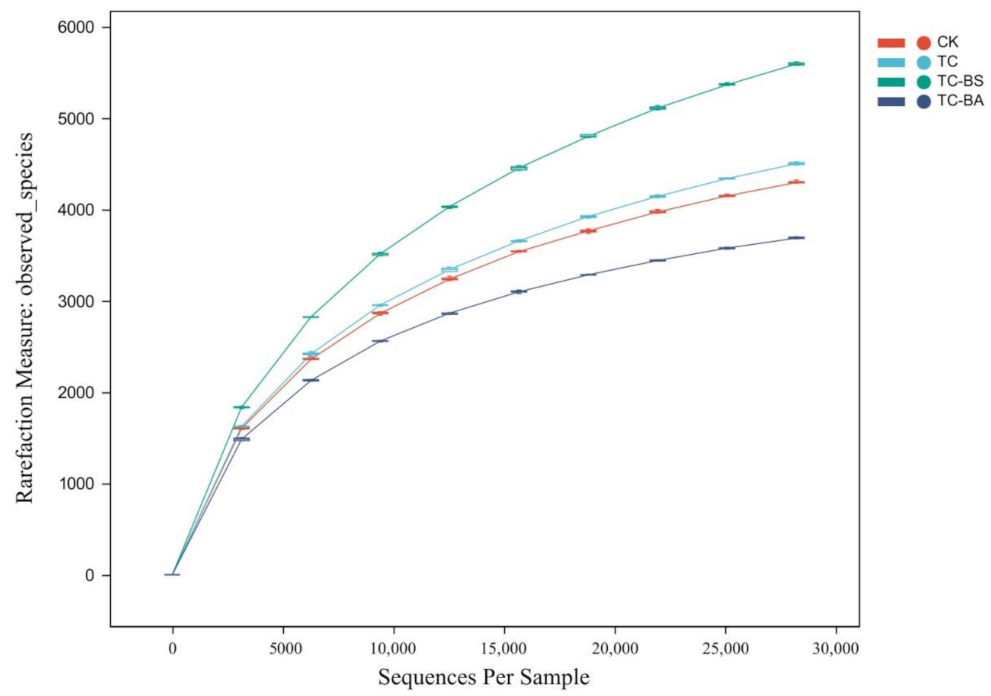
3.3. Effect of Applying Tomato Plant Residue Compost and PGPR on the Alpha Diversity of Ginger Field Soil Bacteria
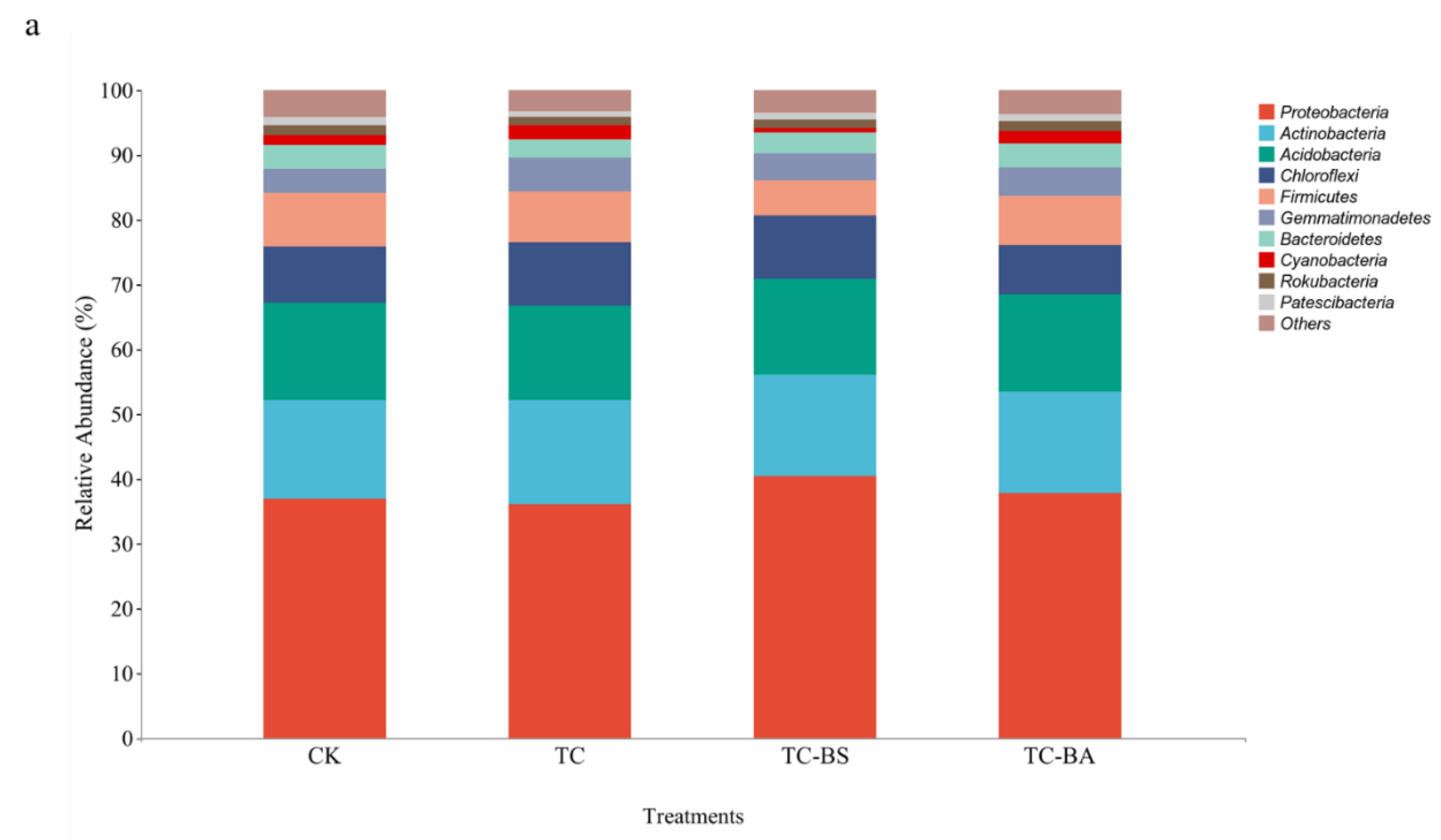
3.4. Effect of Applying Tomato Plant Residue Compost and PGPR on the Relative Abundance of Soil Bacteria in Ginger Field at the Phylum and Genus Level
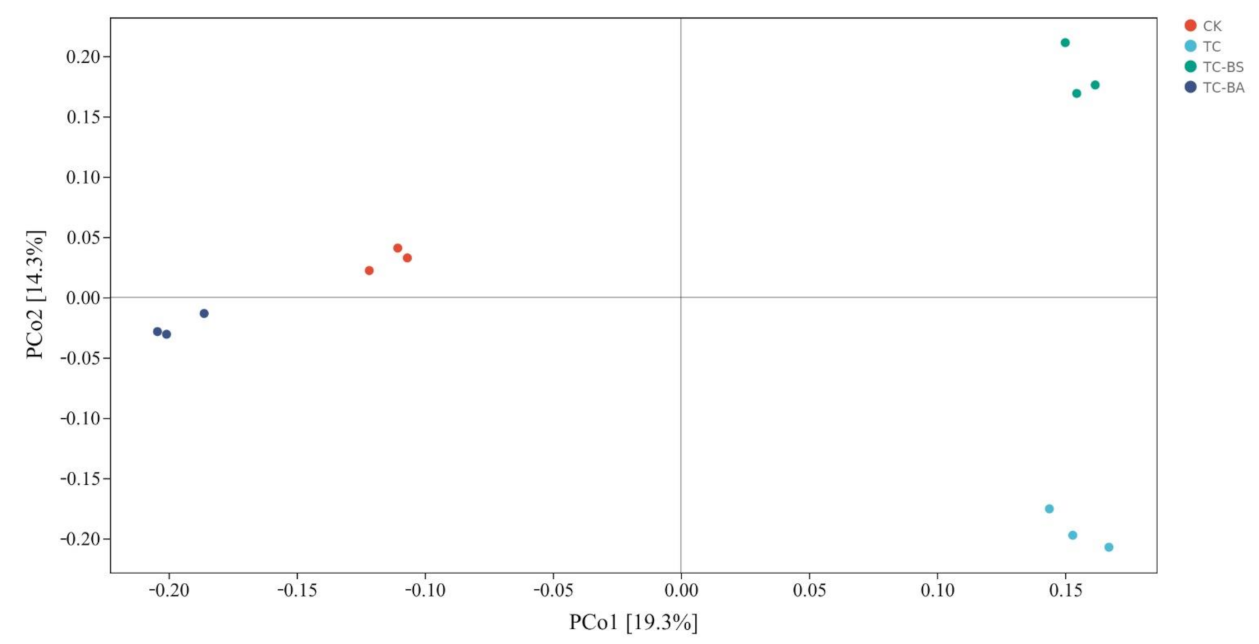
3.5. Principal Coordinate Analysis (PCoA) of the Ginger Field Soil Bacterial Community
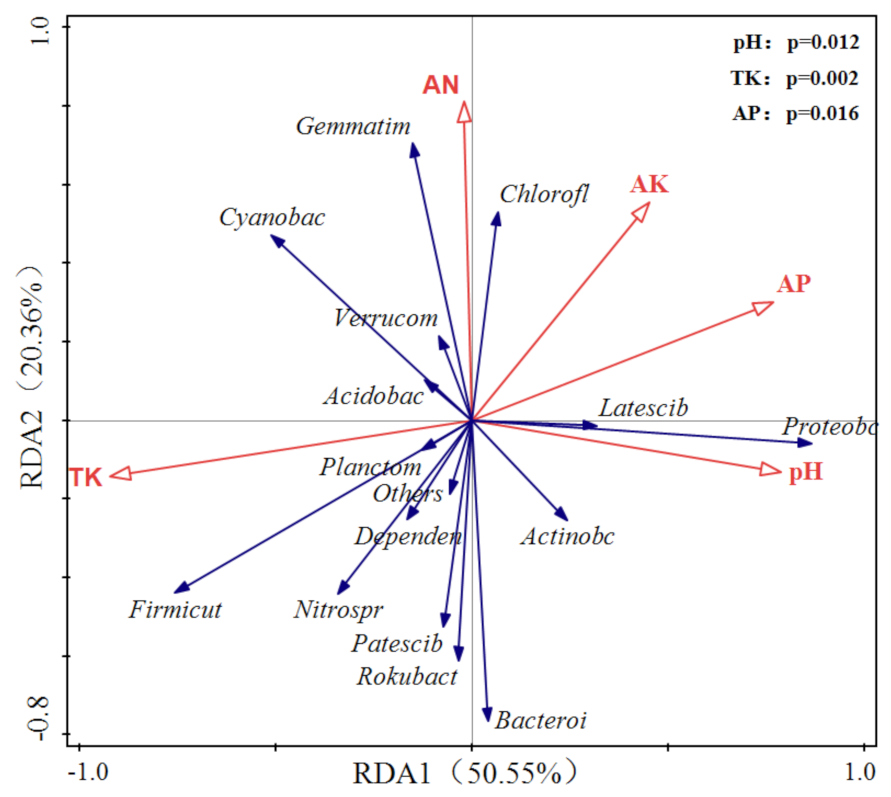
3.6. Redundancy Analysis (RDA) of the Soil Bacterial Community of Ginger Field Soil and Environmental Factors
4. Discussion
4.1. Effect of Applying Tomato Plant Residue Compost and PGPR on Ginger Field Soil Nutrient Content
4.2. Effect of Applying Tomato Plant Residue Compost and PGPR on Ginger Field Soil Enzyme Activity
4.3. Effect of Applying Tomato Plant Residue Compost and PGPR on the Ginger Field Soil Bacterial Community
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Han, L.; Li, Y.S.; Yu, X.C.; He, C.X. Effects of vegetable residue compost returning to soil on soil properties and vegetable yield in solar greenhouse. Chin. J. Appl. Ecol. 2016, 27, 1553–1559. [Google Scholar] [CrossRef]
- Wang, R.; Zhao, Y.; Xie, X.; Mohamed, T.A.; Zhu, L.; Tang, Y.; Chen, Y.; Wei, Z. Role of NH3 recycling on nitrogen fractions during sludge composting. Bioresour. Technol. 2020, 295, 122175. [Google Scholar] [CrossRef] [PubMed]
- Meena, R.S.; Meena, V.S.; Meena, S.K.; Verma, J.P. The needs of healthy soils for a healthy world. J. Clean. Prod. 2015, 102, 560–561. [Google Scholar] [CrossRef]
- Kianpoor Kalkhajeh, Y.; Huang, B.; Hu, W.; Ma, C.; Gao, H.; Thompson, M.L.; Bruun Hansen, H.C. Environmental soil quality and vegetable safety under current greenhouse vegetable production management in China. Agric. Ecosyst. Environ. 2021, 307, 107230. [Google Scholar] [CrossRef]
- Fernández-Gómez, M.J.; Romero, E.; Nogales, R. Feasibility of vermicomposting for vegetable greenhouse waste recycling. Bioresour. Technol. 2010, 101, 9654–9660. [Google Scholar] [CrossRef] [PubMed]
- Sokol, N.W.; Slessarev, E.; Marschmann, G.L.; Nicolas, A.; Blazewicz, S.J.; Brodie, E.L.; Firestone, M.K.; Foley, M.M.; Hestrin, R.; Hungate, B.A.; et al. Life and death in the soil microbiome: How ecological processes influence biogeochemistry. Nat. Rev. Microbiol. 2022, 20, 415–430. [Google Scholar] [CrossRef] [PubMed]
- Postma-Blaauw, M.B.; de Goede, R.G.; Bloem, J.; Faber, J.H.; Brussaard, L. Soil biota community structure and abundance under agricultural intensification and extensification. Ecology 2010, 91, 460–473. [Google Scholar] [CrossRef]
- Zhao, S.; Li, K.; Zhou, W.; Qiu, S.; Huang, S.; He, P. Changes in soil microbial community, enzyme activities and organic matter fractions under long-term straw return in north-central China. Agric. Ecosyst. Environ. 2016, 216, 82–88. [Google Scholar] [CrossRef]
- Ai, C.; Liang, G.; Sun, J.; Wang, X.; Zhou, W. Responses of extracellular enzyme activities and microbial community in both the rhizosphere and bulk soil to long-term fertilization practices in a fluvo-aquic soil. Geoderma 2012, 173–174, 330–338. [Google Scholar] [CrossRef]
- Navarro-Noya, Y.E.; Gómez-Acata, S.; Montoya-Ciriaco, N.; Rojas-Valdez, A.; Suárez-Arriaga, M.C.; Valenzuela-Encinas, C.; Jiménez-Bueno, N.; Verhulst, N.; Govaerts, B.; Dendooven, L. Relative impacts of tillage, residue management and crop-rotation on soil bacterial communities in a semi-arid agroecosystem. Soil Biol. Biochem. 2013, 65, 86–95. [Google Scholar] [CrossRef]
- Meng, F.; Dungait, J.A.J.; Xu, X.; Bol, R.; Zhang, X.; Wu, W. Coupled incorporation of maize (Zea mays L.) straw with nitrogen fertilizer increased soil organic carbon in Fluvic Cambisol. Geoderma 2017, 304, 19–27. [Google Scholar] [CrossRef]
- Kong, P.J.; Zheng, J.; Luan, L.; Chen, Z.Y.; Xue, J.R.; Sun, B.; Jiang, Y.J. Effects of Different Types of Straw Returning on the Bacterial Community, Organic Carbon Mineralization and Maize Yield in Upland Red Soil. Environ. Sci. 2021, 42, 6047–6057. (In Chinese) [Google Scholar] [CrossRef]
- Olasupo, I.O.; Liang, Q.; Zhang, C.; Islam, M.S.; Li, Y.; Yu, X.; He, C. Agronomic Biofortification of Cayenne Pepper Cultivars with Plant Growth-Promoting Rhizobacteria and Chili Residue in a Chinese Solar Greenhouse. Microorganisms 2021, 9, 2398. [Google Scholar] [CrossRef] [PubMed]
- Gray, E.J.; Smith, D.L. Intracellular and extracellular PGPR: Commonalities and distinctions in the plant-bacterium signaling processes. Soil Biol. Biochem. 2005, 37, 395–412. [Google Scholar] [CrossRef]
- Senkovs, M.; Nikolajeva, V.; Makarenkova, G.; Petrina, Z. Influence of Trichoderma asperellum and Bacillus subtilis as biocontrol and plant growth promoting agents on soil microbiota. Ann. Microbiol. 2021, 71, 34. [Google Scholar] [CrossRef]
- Miljaković, D.; Marinković, J.; Balešević-Tubić, S. The Significance of Bacillus spp. in Disease Suppression and Growth Promotion of Field and Vegetable Crops. Microorganisms 2020, 8, 1037. [Google Scholar] [CrossRef]
- Kaspar, F.; Neubauer, P.; Gimpel, M. Bioactive Secondary Metabolites from Bacillus subtilis: A Comprehensive Review. J. Nat. Prod. 2019, 82, 2038–2053. [Google Scholar] [CrossRef]
- Lu, Y.F.; Xu, H.; Duan, H.T.; Mei, X.L.; Li, R.; Guan, X.J.; Zhang, N.; Shen, Q.R. Foliar Spraying of Liquid Compound Microbial Fertilizer Containing Bacillus amyloliquefaciens SQR9 Promoted Plant Foliar Spraying of Liquid Compound Microbial Fertilizer Containing Bacillus amyloliquefaciens SQR9 Promoted Plant Growth. Soils 2019, 51, 903–909. (In Chinese) [Google Scholar] [CrossRef]
- Zhao, W.; Guo, Q.; Li, S.; Wang, P.; Dong, L.; Su, Z.; Zhang, X.; Lu, X.; Ma, P. Effects of Bacillus subtilis NCD-2 and broccoli residues return on potato Verticillium wilt and soil fungal community structure. Biol. Control. 2021, 159, 104628. [Google Scholar] [CrossRef]
- Hui, C.; Sun, P.; Guo, X.; Jiang, H.; Zhao, Y.; Xu, L. Shifts in microbial community structure and soil nitrogen mineralization following short-term soil amendment with the ammonifier Bacillus amyloliquefaciens DT. Int. Biodeterior. Biodegrad. 2018, 132, 40–48. [Google Scholar] [CrossRef]
- Huang, X.F.; Zhou, D.; Guo, J.; Manter, D.K.; Reardon, K.F.; Vivanco, J.M. Bacillus spp. from rainforest soil promote plant growth under limited nitrogen conditions. J. Appl. Microbiol. 2015, 118, 672–684. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Qiu, Y.; Yao, T.; Ma, Y.; Zhang, H.; Yang, X. Effects of PGPR microbial inoculants on the growth and soil properties of Avena sativa, Medicago sativa, and Cucumis sativus seedlings. Soil Tillage Res. 2020, 199, 104577. [Google Scholar] [CrossRef]
- Pane, C.; Celano, G.; Piccolo, A.; Villecco, D.; Spaccini, R.; Palese, A.M.; Zaccardelli, M. Effects of on-farm composted tomato residues on soil biological activity and yields in a tomato cropping system. Chem. Biol. Technol. Agric. 2015, 2, 4. [Google Scholar] [CrossRef] [Green Version]
- Ghaly, A.E.; Alkoaik, F.; Snow, A. Inactivation of Botrytis cinerea during thermophilic composting of greenhouse tomato plant residues. Appl. Biochem. Biotechnol. 2006, 133, 59–75. [Google Scholar] [CrossRef]
- Bao, S.D. Analysis Method of Soil and Agricultural Chemistry, 3rd ed.; China Agricultural Press: Beijing, China, 2000; pp. 25–108. [Google Scholar]
- Guan, S.Y. Soil Enzyme and Its Study Method; China Agriculture Press: Beijing, China, 1986; pp. 270–285. [Google Scholar]
- Liu, Z.; Xie, W.; Yang, Z.; Huang, X.; Zhou, H. Effects of Manure and Chemical Fertilizer on Bacterial Community Structure and Soil Enzyme Activities in North China. Agronomy 2021, 11, 1017. [Google Scholar] [CrossRef]
- Zhang, X.; Tian, X.; Ma, L.; Feng, B.; Liu, Q.; Yuan, L.; Fan, C.; Huang, H.; Huang, H.; Yang, Q. Biodiversity of the Symbiotic Bacteria Associated with Toxic Marine Dinoflagellate Alexandrium tamarense. J. Biosci. Med. 2015, 3, 23–28. [Google Scholar] [CrossRef] [Green Version]
- Lozupone, C.; Knight, R. UniFrac: A New Phylogenetic Method for Comparing Microbial Communities. Appl. Environ. Microbiol. 2005, 72, 8228–8235. [Google Scholar] [CrossRef] [Green Version]
- Ramette, A. Multivariate analyses in microbial ecology. FEMS Microbiol. Ecol. 2007, 62, 142–160. [Google Scholar] [CrossRef] [Green Version]
- Kong, T.; Li, B.; Ke, Y.; Zhu, H.Y.; Ma, Y.; Han, Y.N.; Li, J.F.; Wen, J.F.; Zhang, X. Effect of vegetables waste compost on protected vegetable yield and soil microbial property. Soil Fertil. Sci. China 2017, 5, 157–160. (In Chinese) [Google Scholar] [CrossRef]
- Tabassum, B.; Khan, A.; Tariq, M.; Ramzan, M.; Iqbal Khan, M.S.; Shahid, N.; Aaliya, K. Bottlenecks in commercialisation and future prospects of PGPR. Appl. Soil Ecol. 2017, 121, 102–117. [Google Scholar] [CrossRef]
- Sánchez, Ó.J.; Ospina, D.A.; Montoya, S. Compost supplementation with nutrients and microorganisms in composting process. Waste Manag. 2017, 69, 136–153. [Google Scholar] [CrossRef] [PubMed]
- Tanwar, A.; Aggarwal, A.; Parkash, V. Effect of bioinoculants and superphosphate fertilizer on the growth and yield of broccoli (Brassicaoleracea L. var italica Plenck). N. Z. J. Crop Hortic. Sci. 2014, 42, 288–302. [Google Scholar] [CrossRef] [Green Version]
- Yildirim, E.; Karlidag, H.; Turan, M. Growth, Nutrient Uptake, and Yield Promotion of Broccoli by Plant Growth Promoting Rhizobacteria with Manure. HortsSience 2011, 46, 932–936. [Google Scholar] [CrossRef] [Green Version]
- Liu, X. Application of microbial fertilizer in agricultural production. Agric. Henan 2021, 17, 14–15. (In Chinese) [Google Scholar] [CrossRef]
- Bashir, S.; Bakhsh Gulshan, A.; Iqbal, J.; Husain, A.; Alwahibi, M.S.; Alkahtani, J.; Dwiningsih, Y.; Bakhsh, A.; Ahmed, N.; Jamal Khan, M.; et al. Comparative role of animal manure and vegetable waste induced compost for polluted soil restoration and maize growth. Saudi J. Biol. Sci. 2021, 28, 2534–2539. [Google Scholar] [CrossRef]
- Babalola, O.O. Beneficial bacteria of agricultural importance. Biotechnol. Lett. 2010, 32, 1559–1570. [Google Scholar] [CrossRef]
- Sheng, X.F.; He, L.Y. Solubilization of potassium-bearing minerals by a wild-type strain of Bacillus edaphicus and its mutants and increased potassium uptake by wheat. Saudi J. Biol. Sci. 2006, 52, 66–72. [Google Scholar] [CrossRef]
- Du, J.; Hou, F.; Zhou, Q. Response of soil enzyme activity and soil bacterial community to PCB dissipation across different soils. Chemosphere 2021, 283, 131229. [Google Scholar] [CrossRef]
- Paz-Ferreiro, J.; Trasar-Cepeda, C.; Leirós, M.C.; Seoane, S.; Gil-Sotres, F. Biochemical properties in managed grassland soils in a temperate humid zone: Modifications of soil quality as a consequence of intensive grassland use. Biol. Fertil. Soils 2009, 45, 711–722. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Xiong, J.; Ru, S.; Patil, S.M.; Kurade, M.B.; Govindwar, S.P.; Oh, S.; Jeon, B. Toxicity of benzophenone-3 and its biodegradation in a freshwater microalga Scenedesmus obliquus. J. Hazard. Mater. 2020, 389, 122149. [Google Scholar] [CrossRef]
- Chen, H.; Liang, Q.; Gong, Y.; Kuzyakov, Y.; Fan, M.; Plante, A.F. Reduced tillage and increased residue retention increase enzyme activity and carbon and nitrogen concentrations in soil particle size fractions in a long-term field experiment on Loess Plateau in China. Soil Tillage Res. 2019, 194, 104296. [Google Scholar] [CrossRef]
- Asghar, W.; Kataoka, R. Effect of co-application of Trichoderma spp. with organic composts on plant growth enhancement, soil enzymes and fungal community in soil. Arch. Microbiol. 2021, 203, 4281–4291. [Google Scholar] [CrossRef]
- Ning, C.C.; Wang, J.H.; Cai, K.Z. The Effects of Organic Fertilizers on Soil Fertility and Soil Environmental Quality: A Review. Ecol. Environ. Sci. 2016, 25, 175–181. (In Chinese) [Google Scholar] [CrossRef]
- Pascual, J.A.; García, C.; Hernandez, T. Lasting microbiological and biochemical effects of the addition of municipal solid waste. Arid. Soil. Biol. Fertil. Soils 1999, 30, 1–6. [Google Scholar] [CrossRef]
- Lu, K.Z.; Jin, Q.; Cao, Q.; Li, S.S.; Sun, S.; Jiang, Q.Y.; JIin, J.R.; Ling, L.C.; Fu, X.H.; Du, X.; et al. Effects of different microbial agents on tomato quality and soil enzyme activity. J. Shanghai Norm. Univ. Nat. Sci. 2019, 48, 197–206. (In Chinese) [Google Scholar] [CrossRef]
- Li, J.H.; Gao, K.X.; Wan, L.; Cao, G.Y.; Jiao, F.Y.; Wang, Y.P.; Tian, Y.H.; Zhao, B.J.; Li, C.R. Effects of microbial agent on the growth of Catalpa bungei seedlings and the diversity of bacterial community in rhizosphere soil. Acta Ecol. Sin. 2020, 40, 7588–7607. (In Chinese) [Google Scholar] [CrossRef]
- Yan, Y.; Kuramae, E.E.; de Hollander, M.; Klinkhamer, P.G.; van Veen, J.A. Functional traits dominate the diversity-related selection of bacterial communities in the rhizosphere. ISME J. 2017, 11, 56–66. [Google Scholar] [CrossRef] [Green Version]
- Du, T.; He, H.; Zhang, Q.; Lu, L.; Mao, W.; Zhai, M. Positive effects of organic fertilizers and biofertilizers on soil microbial community composition and walnut yield. Appl. Soil Ecol. 2022, 175, 104457. [Google Scholar] [CrossRef]
- Olasupo, I.O.; Wang, J.; Wei, X.; Sun, M.; Li, Y.; Yu, X.; Yan, Y.; He, C. Chili residue and Bacillus laterosporus synergy impacts soil bacterial microbiome and agronomic performance of leaf mustard (Brassica juncea L.) in a solar greenhouse. Plant Soil 2022. [Google Scholar] [CrossRef]
- Song, Y.L.; Yu, J.; Chen, S.G.; Xiao, C.Z.; Li, Y.H.; Su, X.R.; Ding, F.J. Effects of Complex Microbial Agent on Cotton Physiological Characteristics, Microorganism and Physicochemical Properties in Rhizosphere Soil. Soils 2019, 51, 477–487. (In Chinese) [Google Scholar] [CrossRef]
- McKenney, P.T.; Driks, A.; Eichenberger, P. The Bacillus subtilis endospore: Assembly and functions of the multilayered coat. Nat. Rev. Microbiol. 2013, 11, 33–44. [Google Scholar] [CrossRef] [PubMed]
- Delgado-Baquerizo, M.; Oliverio, A.M.; Brewer, T.E.; Benavent-González, A. A global atlas of the dominant bacteria found in soil. Science 2018, 359, 320–325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sui, J.; Ji, C.; Wang, X.; Liu, Z.; Sa, R.; Hu, Y.; Wang, C.; Li, Q.; Liu, X. A plant growth-promoting bacterium alters the microbial community of continuous cropping poplar trees’ rhizosphere. J. Appl. Microbiol. 2019, 126, 1209–1220. [Google Scholar] [CrossRef]
- Wei, X.; Li, Y.; Fan, X.; He, C.; Yan, Y.; Sun, M.; Ding, C.; Wang, J.; Yu, X. Techno-Economic Feasibility of In Situ Vegetable Residue Return in the Chinese Solar Greenhouse. Agronomy 2021, 11, 1828. [Google Scholar] [CrossRef]
- Xun, W.; Huang, T.; Zhao, J.; Ran, W.; Wang, B.; Shen, Q.; Zhang, R. Environmental conditions rather than microbial inoculum composition determine the bacterial composition, microbial biomass and enzymatic activity of reconstructed soil microbial communities. Soil Biol. Biochem. 2015, 90, 10–18. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | SOM g/kg | TN % | TP % | TK % | AN mg/kg | AP mg/kg | AK mg/kg | pH | EC us/cm |
|---|---|---|---|---|---|---|---|---|---|
| CK | 33.43 ± 1.33 b | 0.16 ± 0.01 b | 0.17 ± 0.01 a | 1.86 ± 0.07 a | 137.00 ± 2.00 c | 158.67 ± 6.66 c | 381.67 ± 6.51 c | 7.2 ± 0.01 a | 120.00 ± 0.01 b |
| TC | 39.23 ± 3.06 a | 0.19 ± 0.01 a | 0.18 ± 0.01 a | 1.80 ± 0.23 a | 166.67 ± 6.51 a | 189.67 ± 4.62 b | 523.67 ± 18.18 a | 7.2 ± 0.02 a | 123.33 ± 1.15 ab |
| TC-BS | 36.33 ± 0.85 ab | 0.18 ± 0.01 a | 0.18 ± 0.01 a | 1.51 ± 0.16 b | 152.00 ± 1.00 b | 208.00 ± 8.54 a | 524.33 ± 28.01 a | 7.3 ± 0.01 a | 130.33 ± 4.51 a |
| TC-BA | 34.43 ± 0.78 b | 0.16 ± 0.01 b | 0.17 ± 0.01 a | 1.78 ± 0.09 a | 137.67 ± 1.53 c | 188.67 ± 5.78 b | 453.33 ± 22.37 b | 7.3 ± 0.01 a | 126.67 ± 4.93 b |
| Treatments | Effective Sequence Number | Chao1 Index | Shannon Index | Simpson Index | Observed_Species | Goods_Coverage |
|---|---|---|---|---|---|---|
| CK | 61282 | 5561.30 ± 581.21 b | 10.96 ± 0.10 b | 0.9988 ± 0.00 b | 4406.63 ± 324.11 b | 0.95 ± 0.01 a |
| TC | 58745 | 5891.18 ± 86.66 b | 10.98 ± 0.04 b | 0.9988 ± 0.00 b | 4539.30 ± 61.51 b | 0.95 ± 0.00 a |
| TC-BS | 90548 | 7446.96 ± 250.02 a | 11.43 ± 0.02 a | 0.9992 ± 0.00 a | 5590.23 ± 107.54 a | 0.96 ± 0.00 a |
| TC-BA | 38885 | 4162.51 ± 50.14 c | 10.69 ± 0.06 c | 0.9986 ± 0.00 b | 3609.3± 54.23 c | 0.95 ± 0.00 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xie, K.; Sun, M.; Shi, A.; Di, Q.; Chen, R.; Jin, D.; Li, Y.; Yu, X.; Chen, S.; He, C. The Application of Tomato Plant Residue Compost and Plant Growth-Promoting Rhizobacteria Improves Soil Quality and Enhances the Ginger Field Soil Bacterial Community. Agronomy 2022, 12, 1741. https://doi.org/10.3390/agronomy12081741
Xie K, Sun M, Shi A, Di Q, Chen R, Jin D, Li Y, Yu X, Chen S, He C. The Application of Tomato Plant Residue Compost and Plant Growth-Promoting Rhizobacteria Improves Soil Quality and Enhances the Ginger Field Soil Bacterial Community. Agronomy. 2022; 12(8):1741. https://doi.org/10.3390/agronomy12081741
Chicago/Turabian StyleXie, Kunhao, Mintao Sun, Aokun Shi, Qinghua Di, Ru Chen, Duo Jin, Yansu Li, Xianchang Yu, Shuangchen Chen, and Chaoxing He. 2022. "The Application of Tomato Plant Residue Compost and Plant Growth-Promoting Rhizobacteria Improves Soil Quality and Enhances the Ginger Field Soil Bacterial Community" Agronomy 12, no. 8: 1741. https://doi.org/10.3390/agronomy12081741