
Responses of N2O Production and Abundances of Associated Microorganisms to Soil Profiles and Water Regime in Two Paddy Soils

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Location and Soils Sampling
2.2. Microcosm Experiment
2.3. Physical and Chemical Analysis
2.4. DNA Extraction and Real-Time Quantitative PCR
2.5. Statistical analysis
3. Results
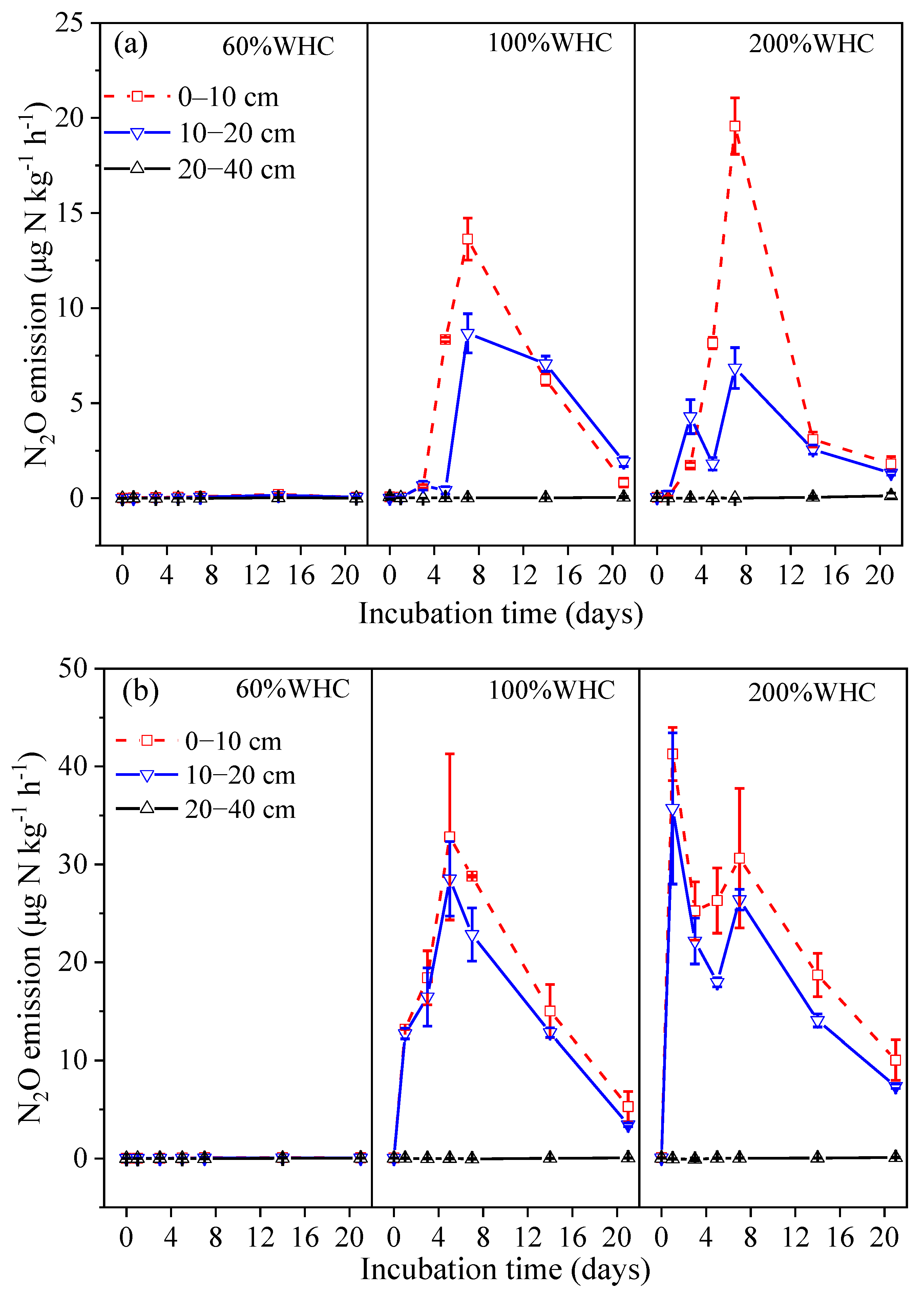
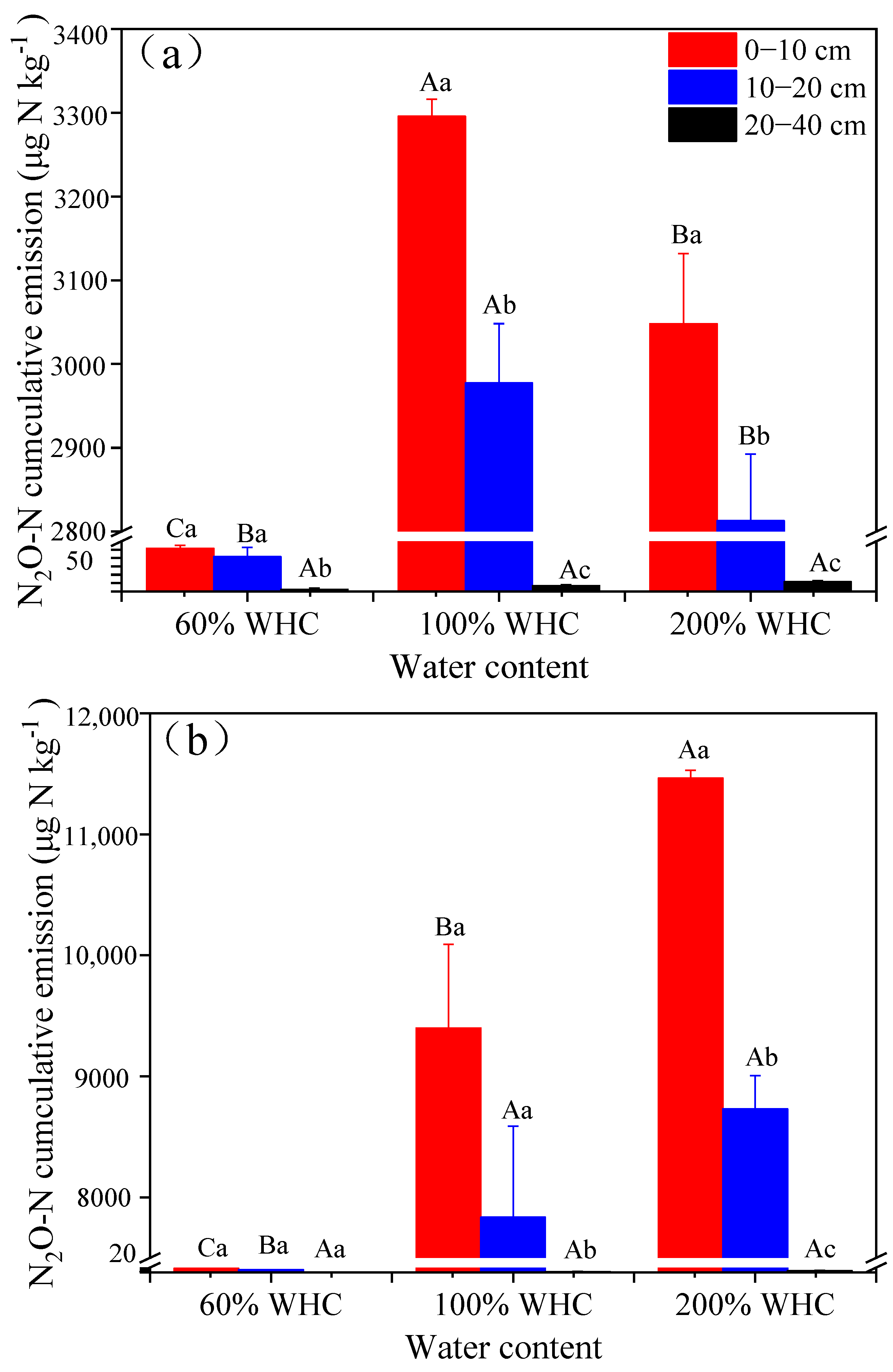
3.1. Soil N2O Emissions
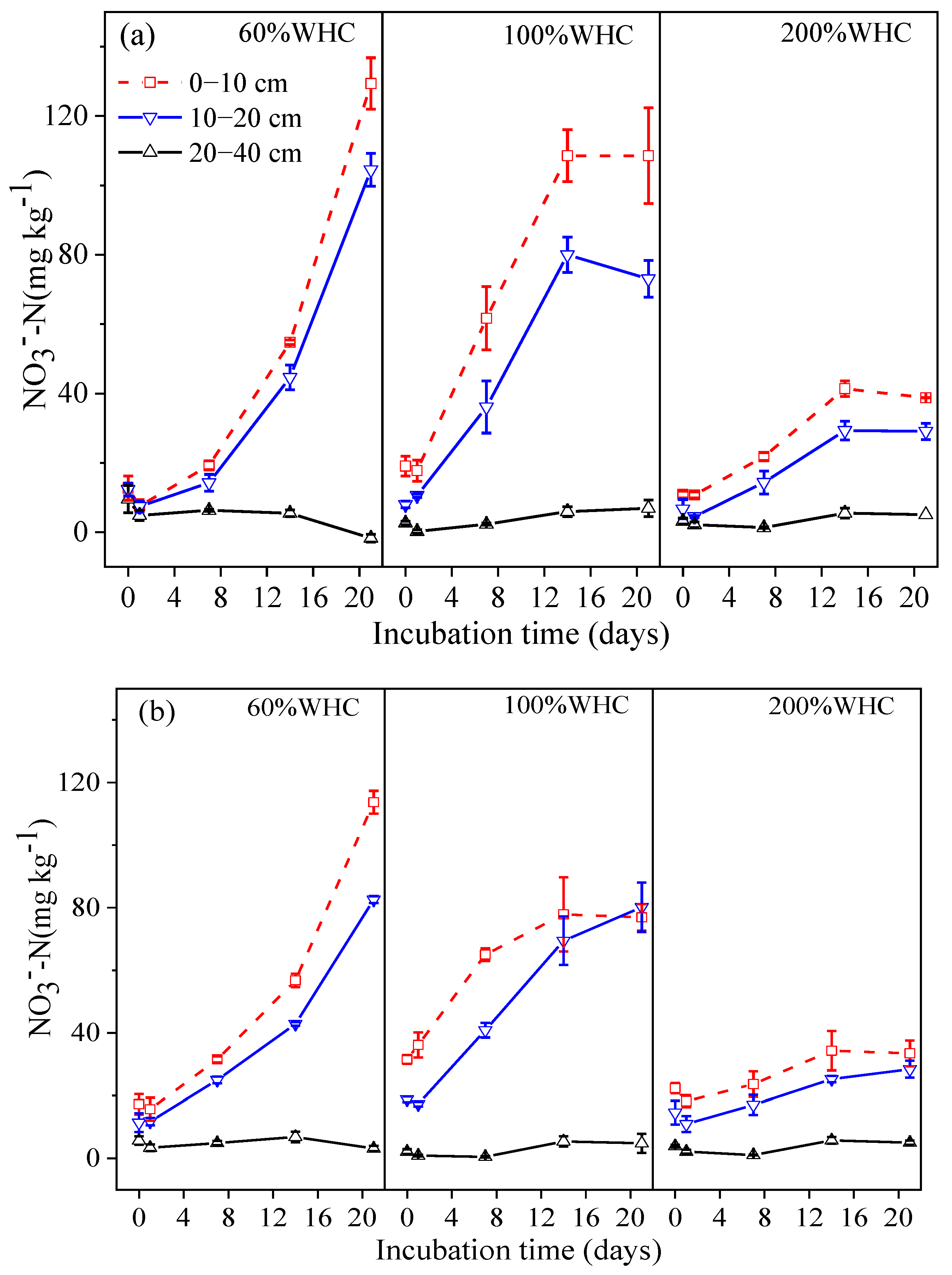
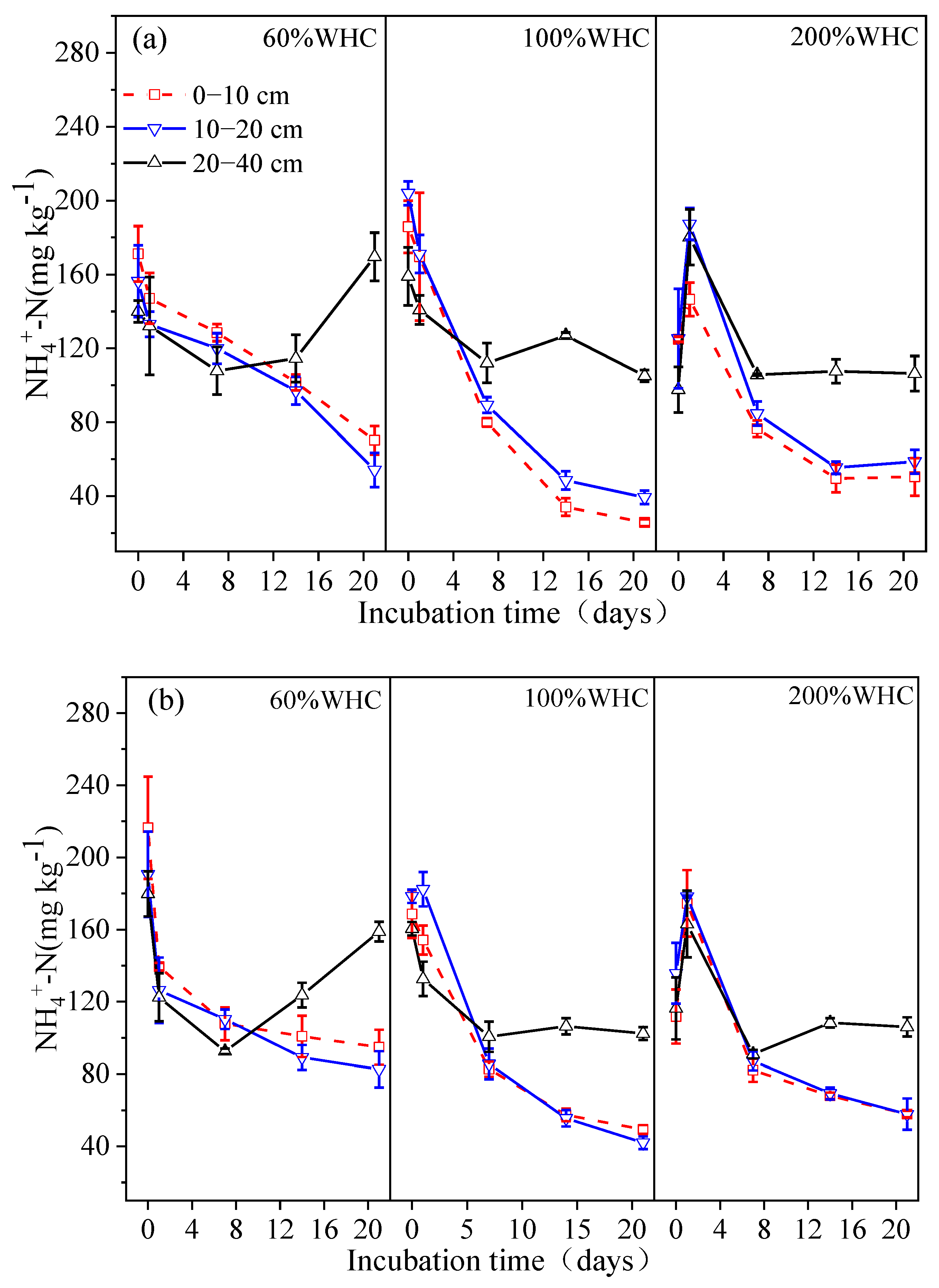
3.2. Soil Inorganic Nitrogen
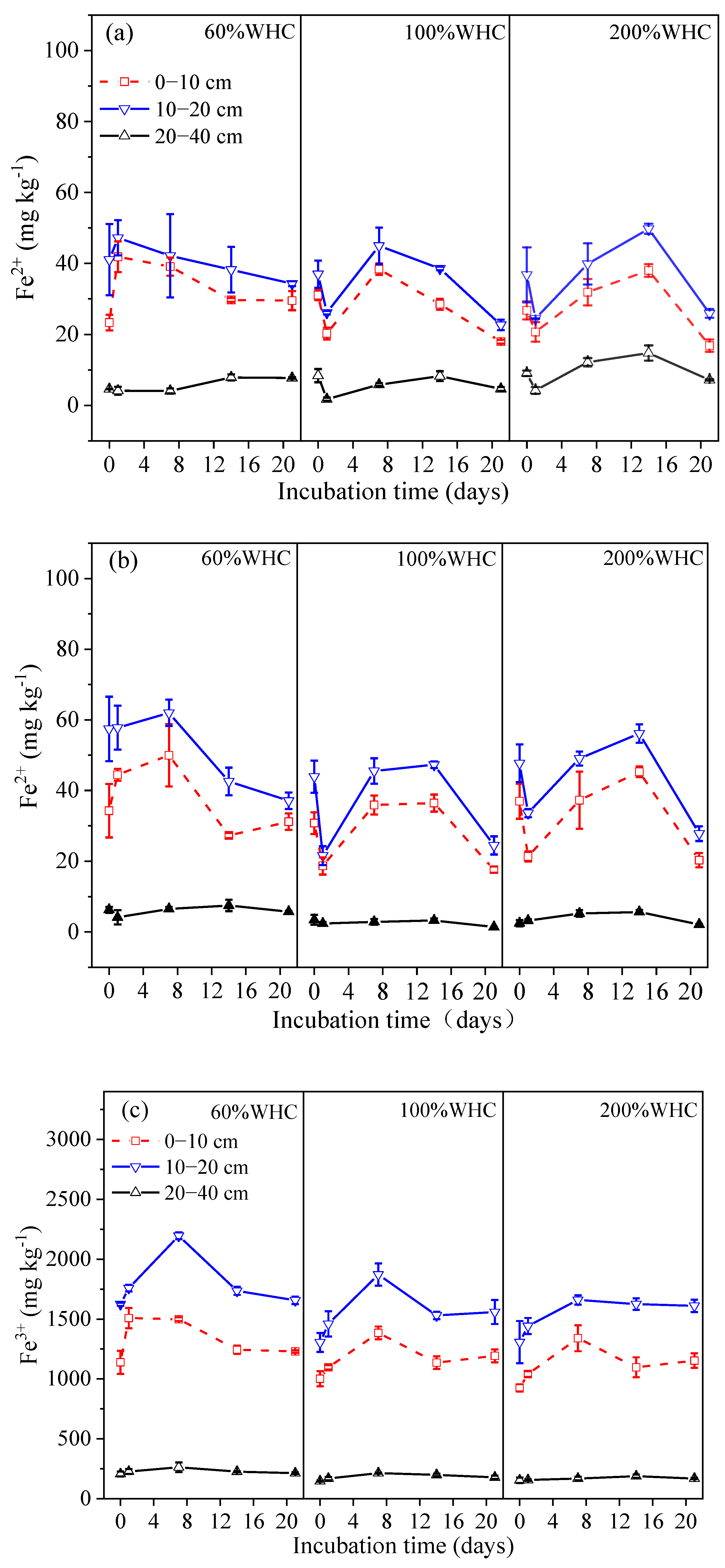
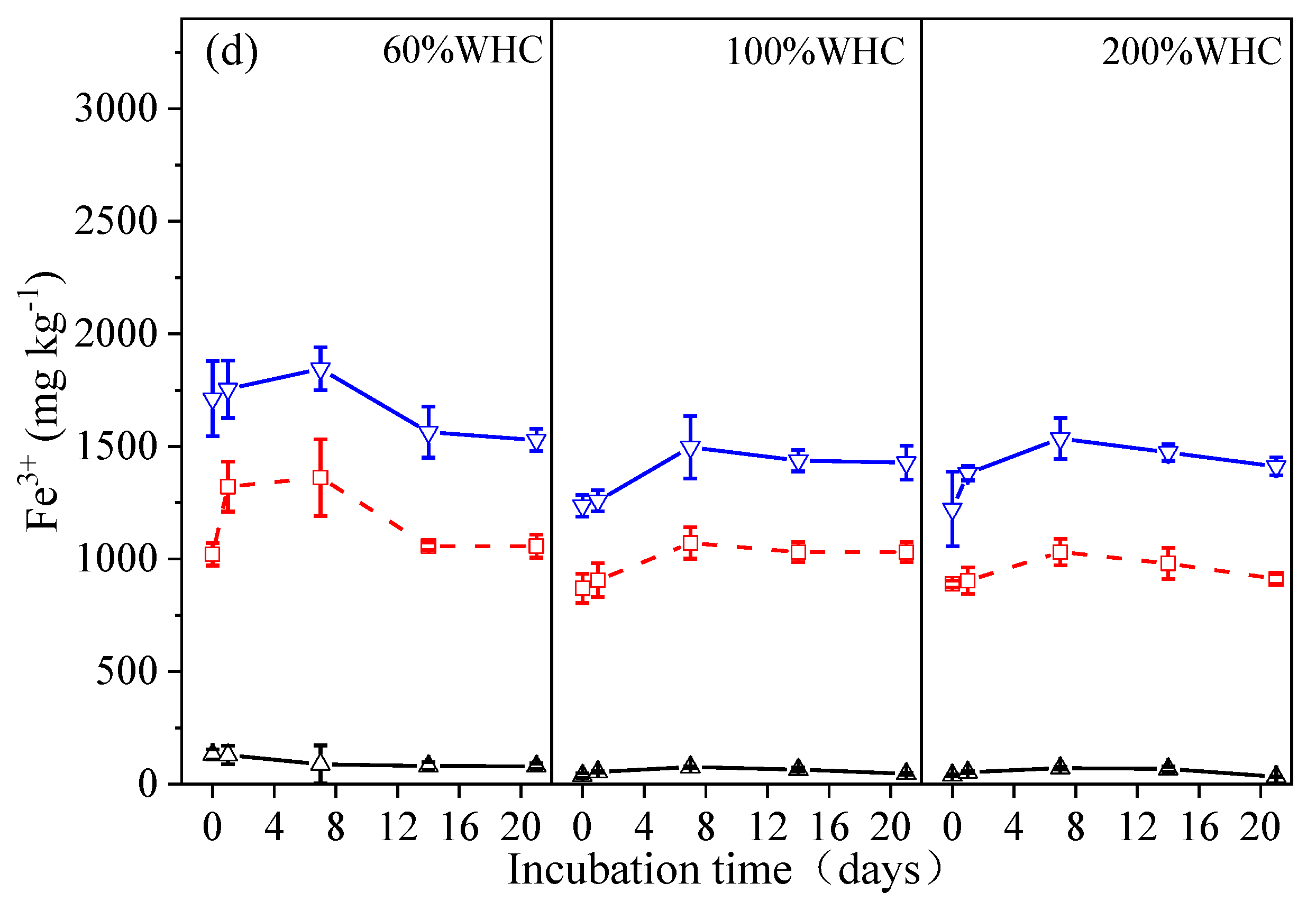
3.3. Dynamics of Fe Concentrations
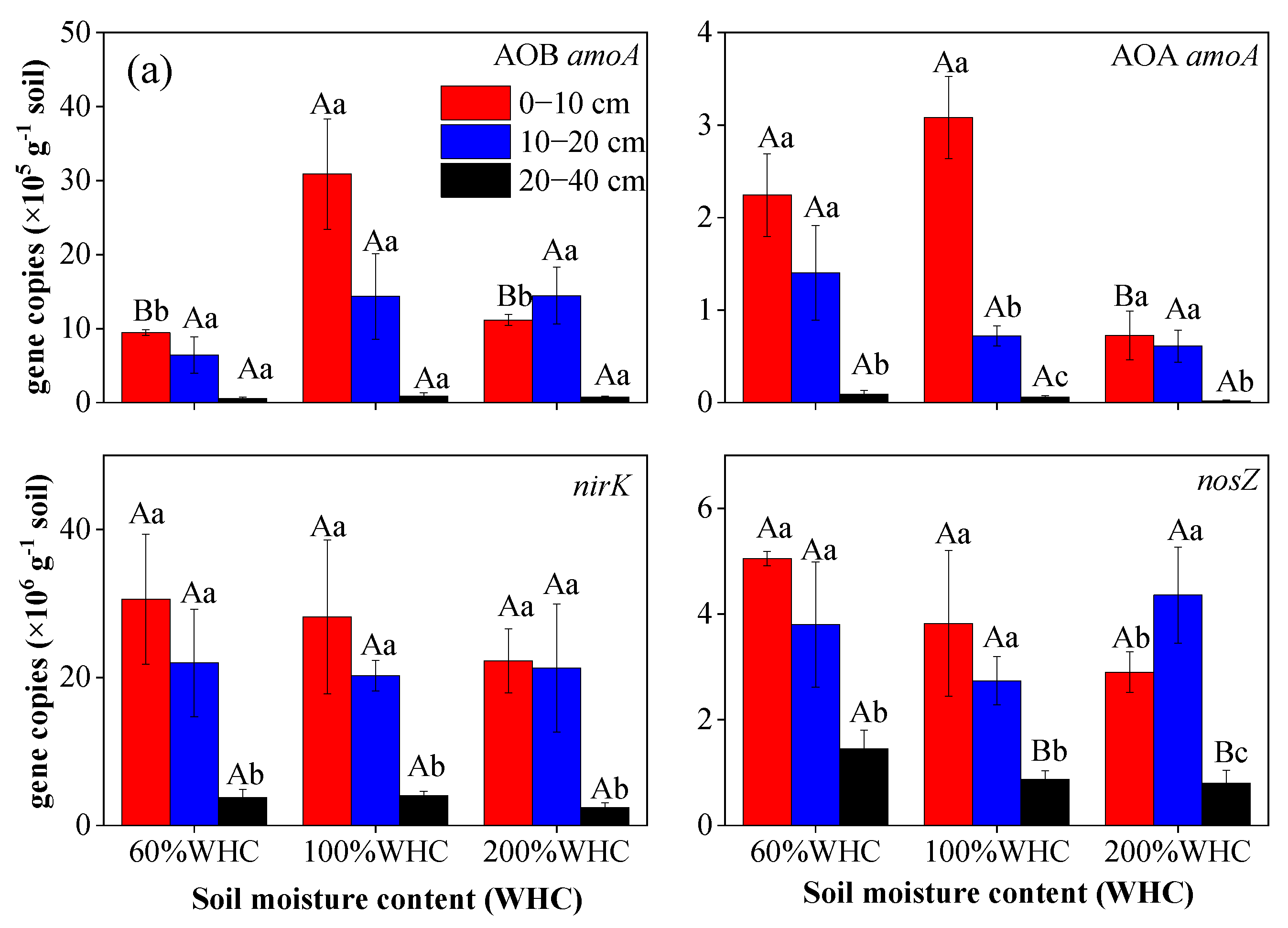
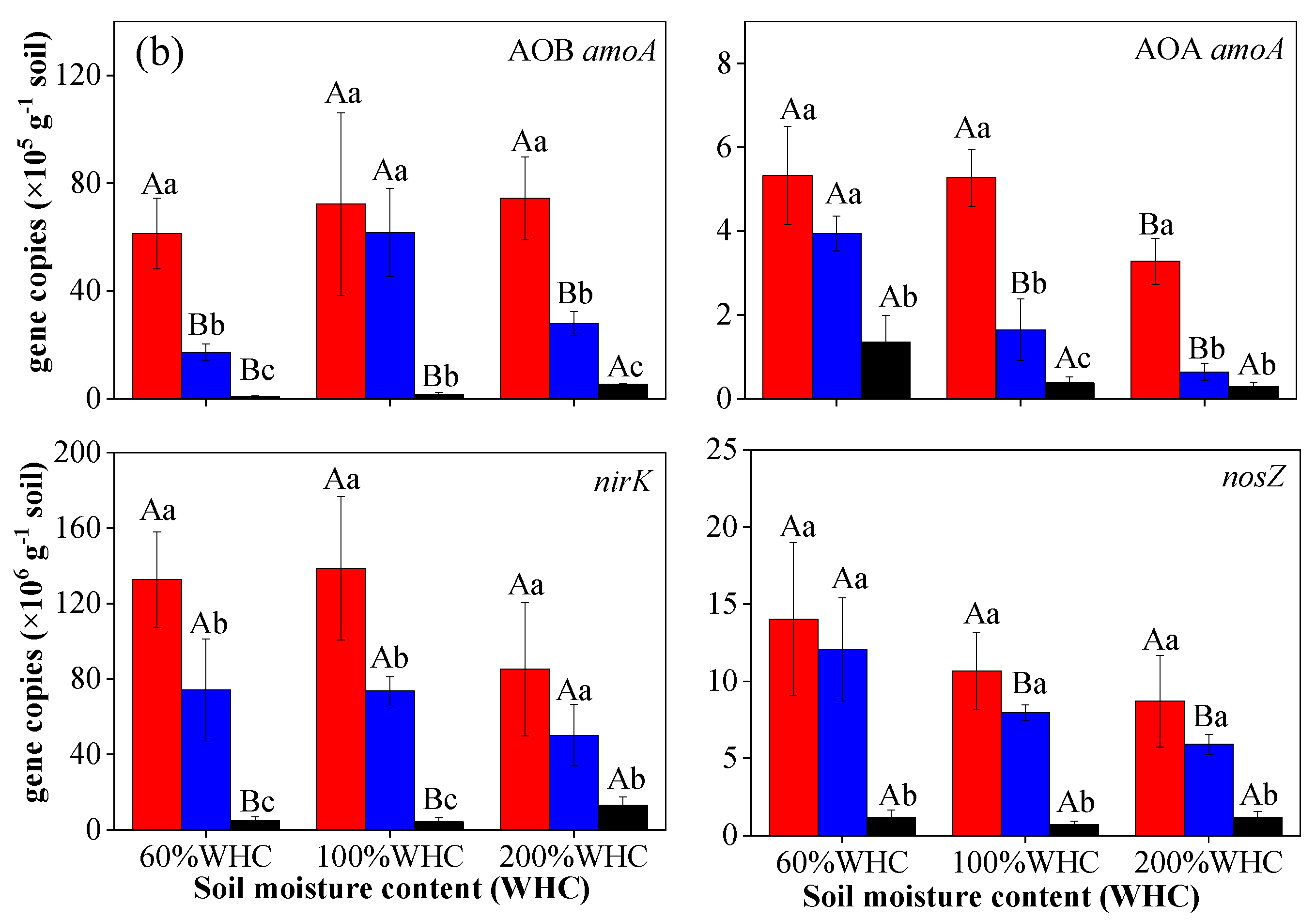
3.4. Abundance of AOA, AOB, nosZ, and nirK
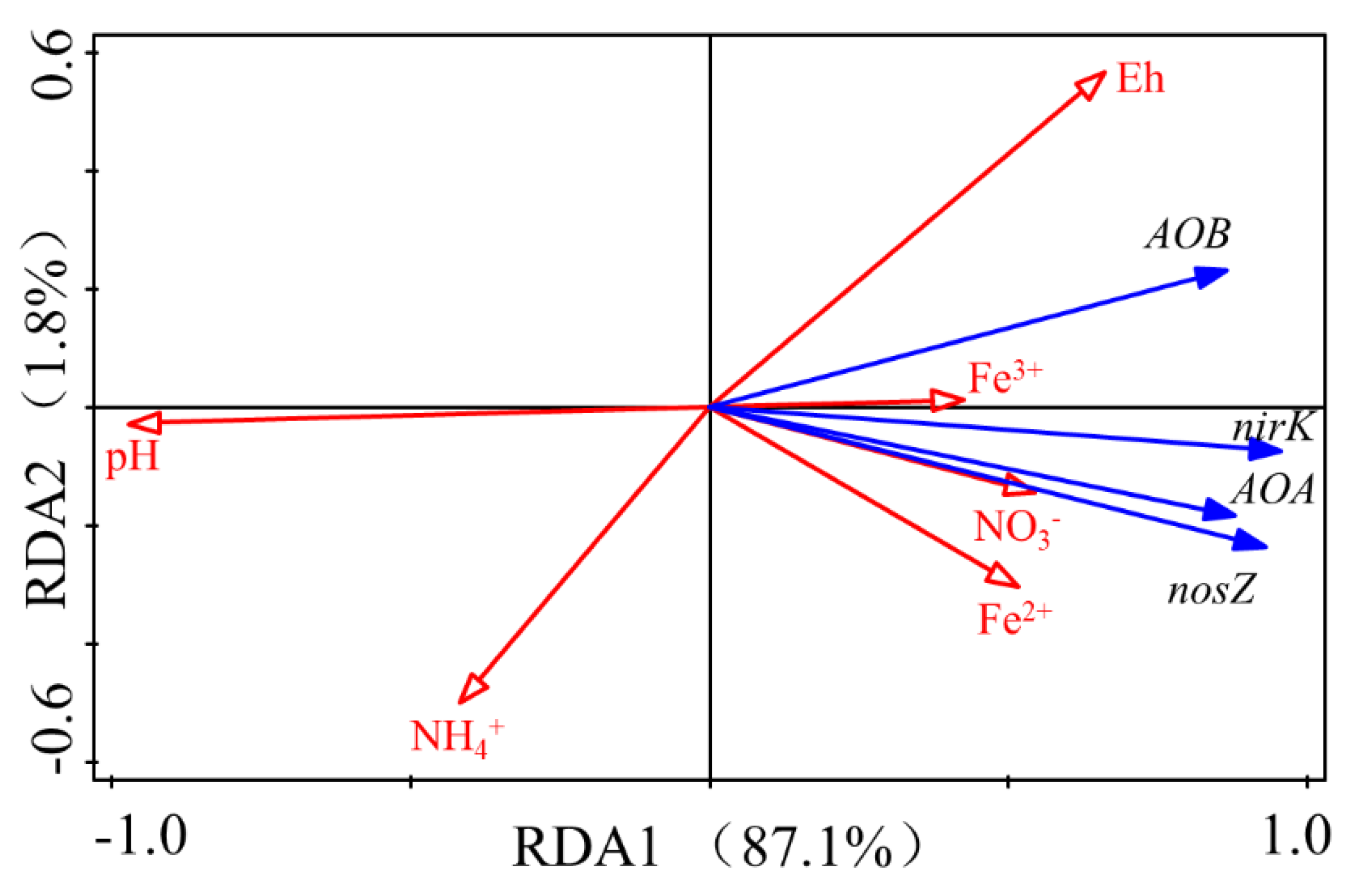
3.5. Correlation of Environmental Factors with N2O Emissions and the Abundance of Nitrifiers and Denitrifiers Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- IPCC. Refinement to the 2006 IPCC guidelines for National Greenhouse Gas Inventories, Advance version; Intergovernmental Panel on Climate Change IPCC: Geneva, Switzerland, 2019. [Google Scholar]
- Zou, J.W.; Huang, Y.; Zheng, X.H.; Wang, Y.S. Quantifying direct N2O emissions in paddy fields during rice growing season in mainland China: Dependence on water regime. Atmos. Environ. 2007, 41, 8030–8042. [Google Scholar] [CrossRef]
- Wang, Q.; Liu, Y.R.; Zhang, C.J.; Zhang, L.M.; Han, L.L.; Shen, J.P.; He, J.Z. Responses of soil nitrous oxide production and abundances and composition of associated microbial communities to nitrogen and water amendment. Biol. Fertil. Soils 2017, 53, 601–611. [Google Scholar] [CrossRef]
- Hoben, J.P.; Gehl, R.J.; Millar, N.; Grace, P.R.; Robertson, G.P. Nonlinear nitrous oxide (N2O) response to nitrogen fertilizer in on-farm corn crops of the US Midwest. Glob. Chang. Biol. 2011, 17, 1140–1152. [Google Scholar] [CrossRef]
- Liu, C.Y.; Wang, K.; Meng, S.X.; Zheng, X.H.; Zhou, Z.X.; Han, S.H.; Chen, D.; Yang, Z.P. Effects of irrigation, fertilization and crop straw management on nitrous oxide and nitric oxide emission from a wheat-maize rotation field in northern China. Agric. Ecosyst. Environ. 2012, 140, 226–233. [Google Scholar] [CrossRef]
- Wang, M.L.; Hu, R.G.; Zhao, J.S.; Kuzyakov, Y.; Liu, S.R. Iron oxidation affects nitrous oxide emissions via donating electrons to denitrification in paddy soils. Geoderma 2016, 271, 173–180. [Google Scholar] [CrossRef]
- Liu, Y.R.; Manuel, D.B.; Pankaj, T.; He, J.Z.; Brajesh, S. Species identity of biocrust-forming lichens drives the response of soil nitrogen cycle to altered precipitation frequency and nitrogen amendment. Soil Biol. Biochem. 2016, 96, 128–136. [Google Scholar] [CrossRef]
- Wrage, N.; Horn, M.A.; Well, R.; Müller, C.; Velthof, G.; Oenema, O. The role of nitrifier denitrification in the production of nitrous oxide revisited. Soil Biol. Biochem. 2018, 123, 3–16. [Google Scholar] [CrossRef]
- Butterbach, B.K.; Baggs, E.M.; Dannenmann, M.; Kiese, R.; Zechmeister, B.S. Review article: Nitrous oxide emissions from soils: How well do we understand the processes and their controls? Philos. Trans. R. Soc. B 2013, 368, 20130122. [Google Scholar] [CrossRef]
- Braker, G.; Conrad, R. Diversity, structure, and size of N2O-producing microbial communities in soils-what matters for their functioning? Adv. Appl. Microbiol. 2011, 75, 33–70. [Google Scholar]
- Baggs, E.M. Soil microbial sources of nitrous oxide: Recent advances in knowledge, emerging challenges and future direction. Curr. Opin. Environ. Sustain. 2011, 3, 321–327. [Google Scholar] [CrossRef]
- Cao, Z.H.; Huang, J.F.; Zhang, C.S.; Li, A.F. Soil quality evolution after land use change from paddy soil to vegetable land. Environ. Geochem. Health 2004, 26, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Fu, Q.L.; Xi, R.Z.; Zhu, J.; Hu, H.Q.; Xing, Z.Q.; Zuo, J.C. The relative contribution of ammonia oxidizing bacteria and archaea to N2O emission from two paddy soils with different fertilizer N sources: A microcosm study. Geoderma 2020, 375, 114486. [Google Scholar] [CrossRef]
- Han, X.G.; Xu, C.C.; Nie, Y.X.; He, J.H.; Wang, W.J.; Deng, Q.; Shen, W.J. Seasonal variations in N2O emissions in a subtropical forest with exogenous nitrogen enrichment are predominately influenced by the abundances of soil nitrifiers and denitrifiers. J. Geophys. Res.-Biogeosci. 2019, 124, 3635–3651. [Google Scholar] [CrossRef]
- Antonio, C.H.; Jesus, G.L.; Eulogio, J.B. Distinct effect of nitrogen fertilization and soil depth on nitrous oxide emissions and nitrifiers and denitrifiers abundance. Biol. Fertil. Soils 2018, 54, 829–840. [Google Scholar]
- Chen, X.Q.; Zhou, J.M.; Wang, H.Y. Productivity and K-supplying power change by an eight-season potash application in different patterns on two paddy soils. Geoderma 2003, 115, 65–74. [Google Scholar] [CrossRef]
- Huang, L.L.; Jia, X.X.; Shao, M.A.; Chen, L.M.; Han, G.Z.; Zhang, G.L. Phases and rates of iron and magnetism changes during paddy soil development on calcareous marine sediment and acid Quaternary red-clay. Sci. Rep. 2018, 8, 444. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.C.; Yu, S.; Strong, J.; Wang, H.L. Are the biogeochemical cycles of carbon, nitrogen, sulfur, and phosphorus driven by the “FeIII–FeII redox wheel” in dynamic redox environments? J. Soils Sediments 2012, 12, 683–693. [Google Scholar] [CrossRef]
- Zhu, B.X.; Cavazos, A.R.; Ostrom, N.E.; Horwath, W.R.; Glass, J.B. The importance of abiotic reactions for nitrous oxide production. Biogeochemistry 2015, 126, 251–267. [Google Scholar]
- Huang, X.R.; Zhu, B.X.; Horwath, W.R.; Faeflen, S.J.; Luo, H.Y.; Xin, X.P.; Jiang, X.J. Effect of iron oxide on nitrification in two agricultural soils with different pH. Biogeosciences 2016, 13, 5609–5617. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.; Silva, L.C.R.; Doane, T.A.; Horwath, W.R. Iron: The forgotten driver of nitrous oxide production in agricultural soil. PLoS ONE 2013, 8, e60146. [Google Scholar] [CrossRef] [Green Version]
- Senn, D.B.; Hemond, H.F. Nitrate controls on iron and arsenic in an urban lake. Science 2002, 296, 2373–2376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muehe, E.M.; Gerhardt, S.; Schink, B.; Kappler, A. Ecophysiology and the energetic benefit of mixotrophic Fe(II) oxidation by various strains of nitrate-reducing bacteria. FEMS Microbiol. Ecol. 2009, 70, 335–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melton, E.D.; Schmidt, C.; Kappler, A. Microbial iron(II) oxidation in littoral freshwater lake sediment: The potential for competition between phototrophic vs. nitrate-reducing iron(II)-oxidizers. Front. Microbiol. 2012, 3, 197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sawayama, S. Possibility of anoxic ferric ammonium oxidation. J. Biosci. Bioeng. 2006, 101, 70–72. [Google Scholar] [CrossRef]
- Shrestha, J.; Rich, J.J.; Ehrenfeld, J.G.; Jaffe, P.R. Oxidation of ammonium to nitrite under iron-reducing conditions in wetland soils: Laboratory, field demonstrations, and push-pull rate determination. Soil Sci. 2009, 174, 156–164. [Google Scholar] [CrossRef]
- Yang, W.H.; Weber, K.A.; Silver, W.L. Nitrogen loss from soil through anaerobic ammonium oxidation coupled to iron re-duction. Nat. Geosci. 2012, 5, 538–541. [Google Scholar] [CrossRef] [Green Version]
- Shaw, L.J.; Nicol, G.W.; Smith, Z.; Fear, J.; Prosser, J.I.; Baggs, E.M. Nitrosospira spp. can produce nitrous oxide via a nitrifier denitrification pathway. Environ. Microbiol. 2006, 8, 214–222. [Google Scholar] [CrossRef]
- Kim, S.W.; Miyahara, M.; Fushinobu, S.; Wakagi, T.; Shoun, H. Nitrous oxide emission from nitrifying activated sludge de-pendent on denitrification by ammonia-oxidizing bacteria. Bioresour. Technol. 2010, 101, 3958–3963. [Google Scholar] [CrossRef]
- Hu, H.W.; Chen, D.; He, J.Z. Microbial regulation of terrestrial nitrous oxide formation: Understanding the biological pathways for prediction of emission rates. FEMS Microbiol. Rev. 2015, 39, 729–749. [Google Scholar] [CrossRef]
- Chen, S.M.; Wang, F.H.; Zhang, Y.M.; Qin, S.P.; Wei, S.C.; Wang, S.Q.; Hu, C.S.; Liu, B.B. Organic carbon availability limiting microbial denitrification in the deep vadose zone. Environ. Microbiol. 2018, 20, 980–992. [Google Scholar] [CrossRef] [Green Version]
- Deirdre, B.G.; Christoph, M.; Samiran, B.; Wei, M.; Steven, D.S.; Daniel, V.M. Response of ammonia oxidizing archaea and bacteria to changing water filled pore space. Soil Biol. Biochem. 2010, 42, 1888–1891. [Google Scholar]
- Hu, H.W.; Macdonald, C.A.; Trivedi, P.; Holmes, B.; Bodrossy, L.; He, J.Z.; Singh, B.K. Water addition regulates the metabolic activity of ammonia oxidizers responding to environmental perturbations in dry subhumid ecosystems. Environ. Microbiol. 2014, 17, 444–461. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, S.; Helgason, B.; Wang, L.; Winsley, T.; Ferrari, B.C.; Siciliano, S.D. Legacy effects of soil moisture on microbial community structure and N2O emissions. Soil Biol. Biochem. 2016, 95, 40–50. [Google Scholar] [CrossRef]
- Liu, Y.; Li, C.; Nelson, W.C.; Shi, L.; Xu, F.; Liu, Y.; Yan, A.; Zhong, L.; Thompson, C.J.; Fredrickson, J.K.; et al. Effect of water chemistry and hydrodynamics on nitrogen transformation activity and microbial community functional potential in hyporheic zone sediment columns. Environ. Sci. Technol. 2017, 51, 4877–4886. [Google Scholar] [CrossRef]
- Cawse, P.A. The determination of nitrate in soil solutions by ultraviolet spectrophotometry. Analyst 1967, 92, 311–315. [Google Scholar] [CrossRef]
- Scheiner, D. Determination of ammonia and kjeldahl nitrogen by indophenol method. Water Res. 1976, 10, 31–36. [Google Scholar] [CrossRef]
- Viollier, E.; Inglett, P.W.; Hunter, K.; Roychoudhury, A.N.; Van, C.P. The ferrozine method revisited: Fe(II)/Fe(III) determina-tion in natural waters. Appl. Geochem. 2000, 15, 785–790. [Google Scholar] [CrossRef]
- Cheng, Y.; Wang, J.; Wang, S.Q.; Zhang, J.B.; Cai, Z.C. Effects of soil moisture on gross N transformations and N2O emission in acid subtropical forest soils. Biol. Fertil. Soils 2014, 50, 1099–1108. [Google Scholar] [CrossRef]
- Wu, L.; Tang, S.R.; He, D.R.; Wu, X.; Shaaban, M.; Wang, M.L.; Zhao, J.S.; Khan, I.; Zheng, X.H.; Hu, R.G. Conversion from rice to vegetable production increases N2O emission via increased soil organic matter mineralization. Sci. Total Environ. 2017, 583, 190–201. [Google Scholar] [CrossRef]
- Bateman, E.J.; Baggs, E.M. Contributions of nitrification and denitrification to N2O emissions from soils at different water-filled pore space. Biol. Fertil. Soils 2005, 41, 379–388. [Google Scholar] [CrossRef]
- Liu, R.; Helen, H.; Suter, H.; Hu, H.W.; Chen, D. The effect of temperature and moisture on the source of N2O and contributions from ammonia oxidizers in an agricultural soil. Biol. Fertil. Soils 2016, 53, 141–152. [Google Scholar] [CrossRef]
- Kool, D.M.; Dolfing, J.; Wrage, N.; Groenigen, J.W.V. Nitrifier denitrification as a distinct and significant source of nitrous oxide from soil. Soil. Biol. Biochem. 2011, 43, 174–178. [Google Scholar] [CrossRef]
- Huang, T.; Gao, B.; Hu, X.K.; Lu, X.; Well, R.; Christie, P.; Bakken, L.R.; Ju, X.T. Ammonia-oxidation as an engine to generate nitrous oxide in an intensively managed calcareous Fluvo-aquic soil. Sci. Rep. 2014, 4, 3950. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.L.; Hu, R.G.; Ruser, R.; Schmidt, C.; Kappler, A. The role of chemodenitrification for N2O emissions from nitrate reduction in rice paddy soils. ACS Earth Space Chem. 2020, 4, 122–132. [Google Scholar] [CrossRef]
- Ma, L.; Cheng, Y.; Wang, J.Y.; Yan, X.Y. Mechanical insights into the effect of fluctuation in soil moisture on nitrous oxide emissions from paddy soil. Paddy Water Environ. 2017, 15, 359–369. [Google Scholar] [CrossRef]
- Zhang, Y.; Ji, G.D.; Wang, C.; Zhang, X.R.; Xu, M. Importance of denitrification driven by the relative abundances of microbial communities in coastal wetlands. Environ. Pollut. 2018, 244, 47–54. [Google Scholar] [CrossRef]
- Kim, H.; Bae, H.S.; Reddy, K.R.; Ogram, A. Distributions, abundances and activities of microbes associated with the nitrogen cycle in riparian and stream sediments of a river tributary. Water Res. 2016, 106, 51–61. [Google Scholar] [CrossRef] [Green Version]
- Klueglein, N.; Kappler, A. Abiotic oxidation of Fe(II) by reactive nitrogen species in cultures of the nitrate-reducing Fe(II) oxi-dizer Acidovorax sp. BoFeN1—Questioning the existence of enzymatic Fe(II) oxidation. Geobiology 2013, 11, 180–190. [Google Scholar] [CrossRef]
- Zuo, J.C.; Hu, H.Q.; Fu, Q.L.; Zhu, J.; Xing, Z.Q. Biological-chemical comprehensive effects of goethite addition on nitrous oxide emissions in paddy soils. J. Soils Sediments 2020, 20, 3580–3590. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Types and Layers | pH | SOM (g kg−1) | NH4+-N (mg kg−1) | NO3−-N (mg kg−1) | Bulk Density (g cm−3) | Total Fe (g kg−1) | HCl-Extractable Fe (g kg−1) | |
|---|---|---|---|---|---|---|---|---|
| Hydromorphic paddy soil | 0–10 cm | 6.8 b (0.02) | 20.8 bc (0.43) | 39.5 a (0.65) | 8.4 b (0.87) | 1.46 b (0.04) | 31.5 b (0.50) | 2.2 b (0.09) |
| 10–20 cm | 6.9 b (0.06) | 18.7 c (0.78) | 28.4 c (0.55) | 8.1 b (0.58) | 1.54 ab (0.02) | 33.0 a (0.18) | 3.1 a (0.15) | |
| 20–40 cm | 7.0 a (0.01) | 7.3 d (1.05) | 11.2 e (0.39) | 3.9 c (0.32) | 1.61 a (0.03) | 26.4 d (0.29) | 0.4 d (0.04) | |
| Gleyed paddy soil | 0–10 cm | 6.1 e (0.06) | 27.1 a (2.07) | 34.2 b (0.97) | 14.8 a (0.43) | 1.15 d (0.01) | 26.8 d (0.18) | 1.8 c (0.13) |
| 10–20 cm | 6.3 d (0.03) | 23.2 b (2.31) | 22.5 d (0.19) | 9.6 b (0.37) | 1.34 c (0.01) | 30.7 c (0.47) | 3.1 a (0.21) | |
| 20–40 cm | 6.6 c (0.09) | 7.1 d (0.88) | 10.0 e (1.21) | 5.6 c (0.67) | 1.45 b (0.06) | 33.4 a (0.69) | 0.10 e (0.01) | |
| Factors | N2O | AOB amoA | AOA amoA | nirK | nosZ |
|---|---|---|---|---|---|
| Water | ˂0.001 | 0.016 | ˂0.001 | 0.012 | 0.335 |
| Soil layers | ˂0.001 | ˂0.001 | ˂0.001 | ˂0.001 | ˂0.001 |
| Water × Soil layers | ˂0.001 | 0.297 | ˂0.001 | 0.475 | ˂0.565 |
| N2O Total Emission | NH4+-N | NO3--N | Fe2+ | Fe3+ | pH | Eh | AOB | AOA | nirK | nosZ | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| N2O total emission | |||||||||||
| NH4+-N | −0.563 ** | ||||||||||
| NO3−-N | 0.375 ** | −0.290 * | |||||||||
| Fe2+ | 0.516 ** | −0.465 ** | 0.703 ** | ||||||||
| Fe3+ | 0.360 ** | −0.446 ** | 0.706 ** | 0.937 ** | |||||||
| pH | −0.842 ** | 0.424 ** | −0.418 ** | −0.477 ** | −0.265 | ||||||
| Eh | 0.753 ** | −0.278 * | 0.312 * | 0.409 ** | 0.185 | −0.931 ** | |||||
| AOB | 0.742 ** | −0.419 ** | 0.640 ** | 0.535 ** | 0.369 ** | −0.815 ** | 0.724 ** | ||||
| AOA | 0.277 * | 0.023 | 0.756 ** | 0.487 ** | 0.394 ** | −0.489 ** | 0.453 ** | 0.661 ** | |||
| nirK | 0.491 ** | −0.111 | 0.668 ** | 0.523 ** | 0.373 ** | −0.695 ** | 0.614 ** | 0.778 ** | 0.888 ** | ||
| nosZ | 0.336 * | −0.035 | 0.638 ** | 0.650 ** | 0.517 ** | −0.556 ** | 0.536 ** | 0.649 ** | 0.874 ** | 0.906 ** |
| Explains % | Pseudo-F | p | |
|---|---|---|---|
| pH | 81.9 | 72.6 | 0.002 |
| Eh | 2.2 | 2.1 | 0.146 |
| NO3− | 1 | 0.9 | 0.326 |
| Fe3+ | 3.2 | 3.6 | 0.078 |
| NH4+ | 0.5 | 0.5 | 0.592 |
| Fe2+ | 0.2 | 0.2 | 0.79 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zuo, J.; Hu, H.; Fu, Q.; Zhu, J.; Zheng, H.; Mo, M.; Tu, A. Responses of N2O Production and Abundances of Associated Microorganisms to Soil Profiles and Water Regime in Two Paddy Soils. Agronomy 2022, 12, 743. https://doi.org/10.3390/agronomy12030743
Zuo J, Hu H, Fu Q, Zhu J, Zheng H, Mo M, Tu A. Responses of N2O Production and Abundances of Associated Microorganisms to Soil Profiles and Water Regime in Two Paddy Soils. Agronomy. 2022; 12(3):743. https://doi.org/10.3390/agronomy12030743
Chicago/Turabian StyleZuo, Jichao, Hongqing Hu, Qingling Fu, Jun Zhu, Heng Zheng, Minghao Mo, and Anguo Tu. 2022. "Responses of N2O Production and Abundances of Associated Microorganisms to Soil Profiles and Water Regime in Two Paddy Soils" Agronomy 12, no. 3: 743. https://doi.org/10.3390/agronomy12030743