
Effect of Plant Growth-Promoting Bacteria Azospirillum brasilense on the Physiology of Radish (Raphanus sativus L.) under Waterlogging Stress



Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Inoculant and Inoculation Method
2.3. Physiological Measurements
2.4. Growth Measurements
2.5. Statistical Analysis
3. Results
3.1. Physiological Parameters
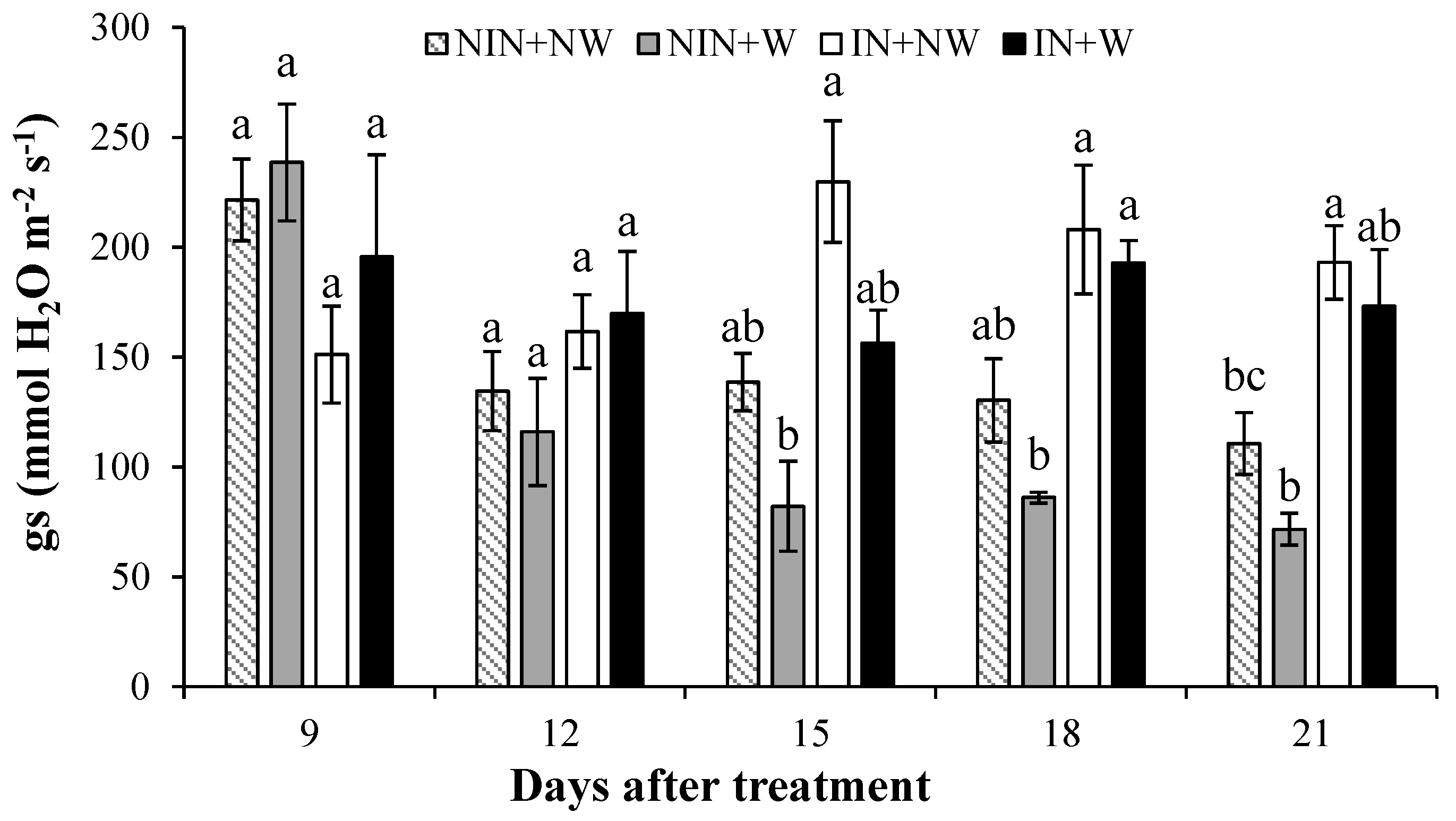
3.1.1. Stomatal Conductance
3.1.2. Chlorophyll Index
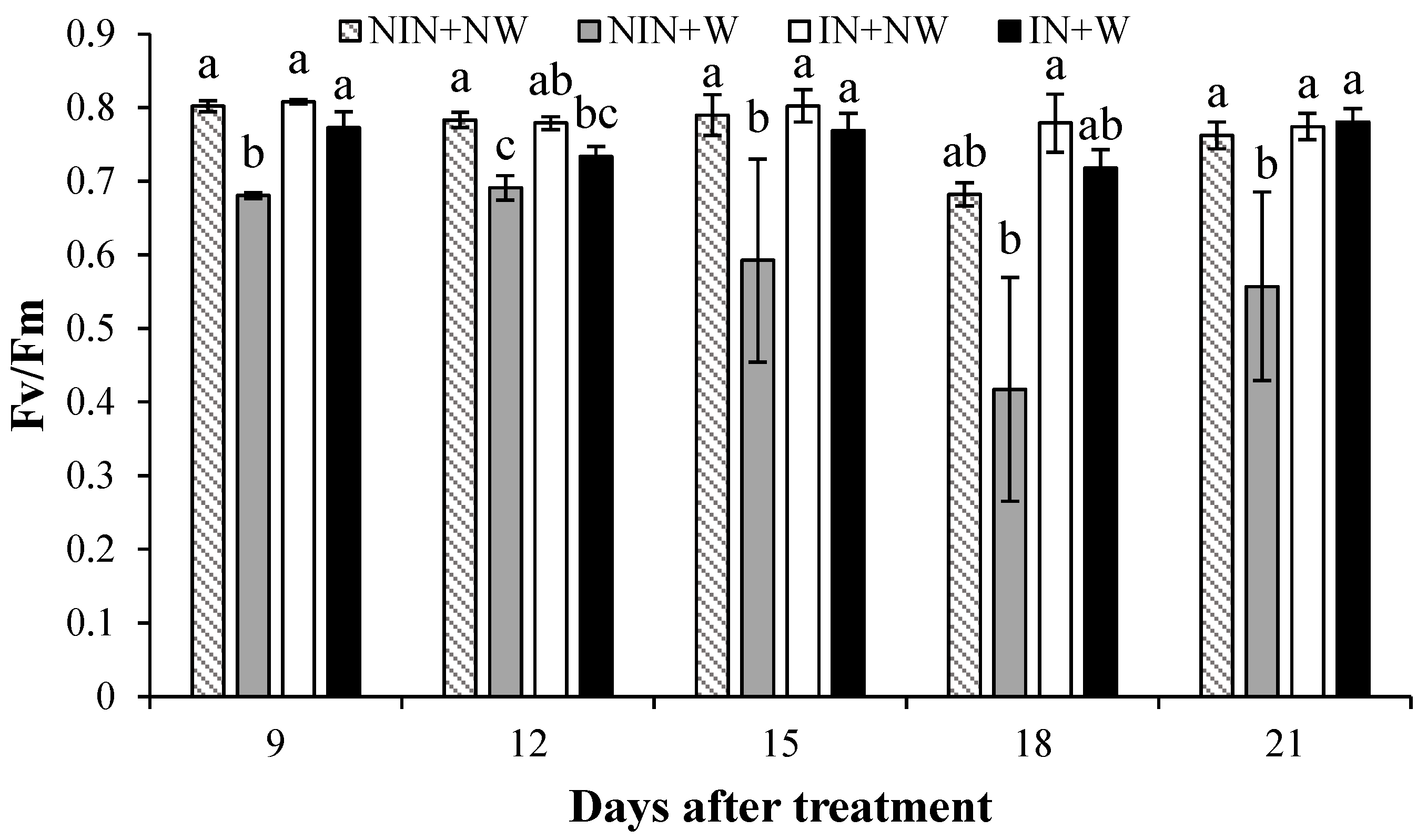
3.1.3. Maximum Efficiency of PSII
3.1.4. Photochemical Quenching (Pq)
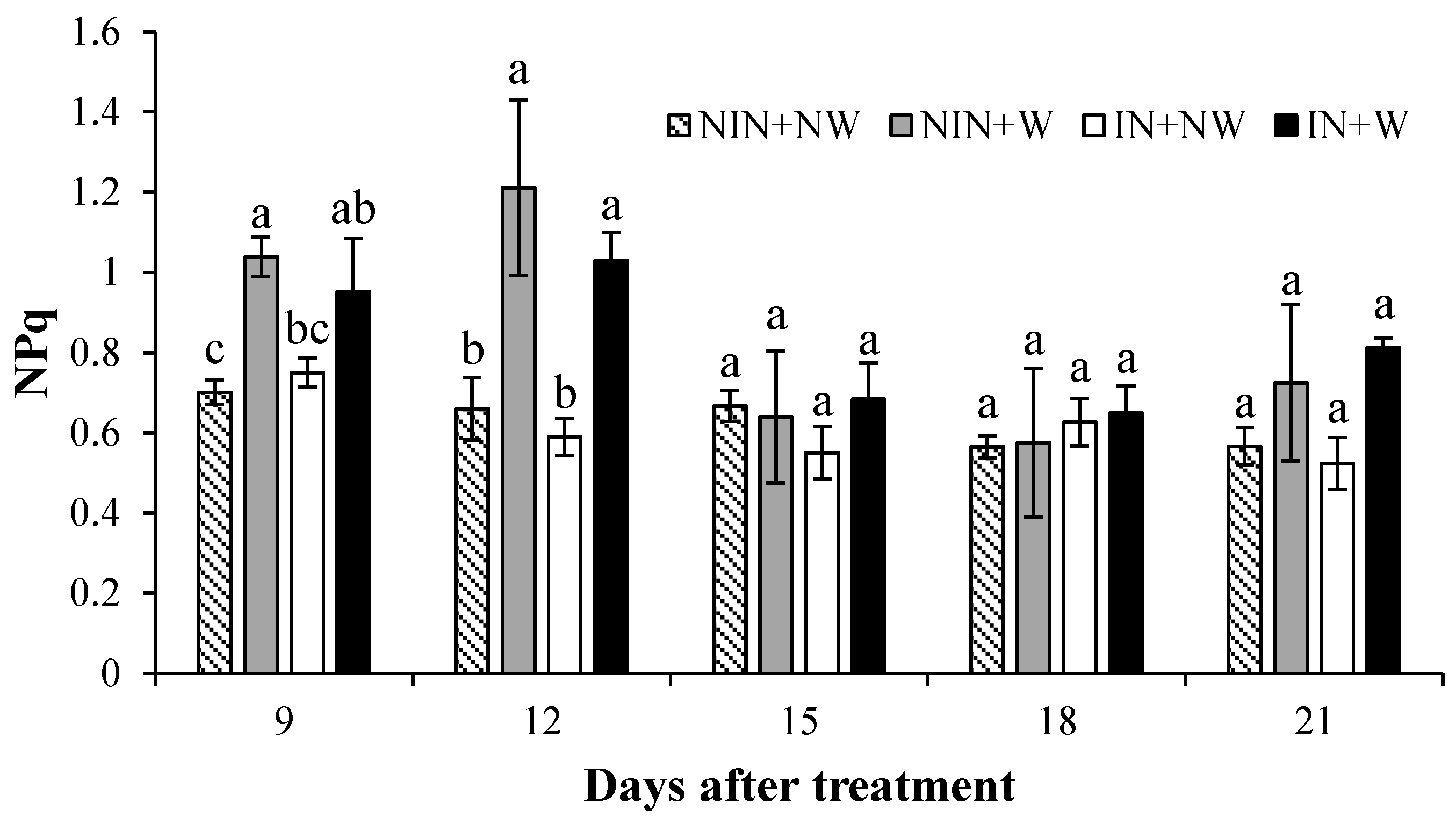
3.1.5. Non-Photochemical Quenching (NPq)
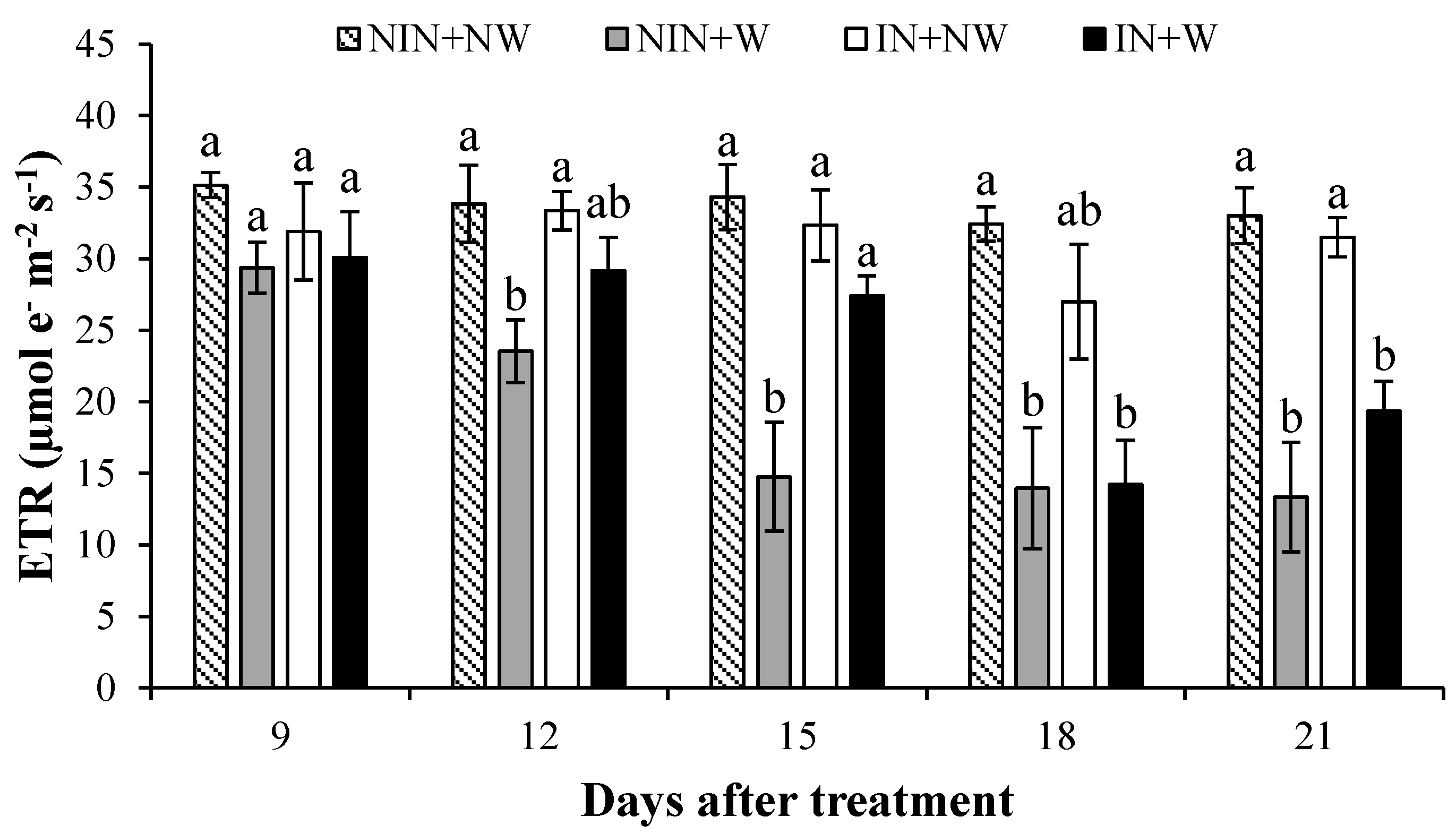
3.1.6. Electron Transport Rate (ETR)
3.2. Growth Parameters
4. Discussion
4.1. Stomatal Conductance
4.2. Chlorophyll Index
4.3. Maximum Efficiency of PSII
4.4. Photochemical Quenching (Pq)
4.5. Non-Photochemical Quenching (NPq)
4.6. Electron Transport Rate (ETR)
4.7. Plant Growth
4.7.1. Tuberous Root Diameter
4.7.2. Number of Leaves
4.7.3. Leaf Area
4.7.4. Fresh and Dry Weight
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Criollo, H.; García, J. Efecto de la densidad de siembra sobre el crecimiento de plantas de rábano (Raphanus sativus L.) bajo invernadero. Rev. Colomb. Cienc. Hortíc. 2009, 3, 210–222. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.B.V.; Saila, E.S.; Narang, P.; Aishwarya, M.; Raina, R.; Gautam, M.; Shankar, E.G. Biofunctionalization and biological synthesis of the ZnO nanoparticles: The effect of Raphanus sativus (white radish) root extract on antimicrobial activity against MDR strain for wound healing applications. Inorg. Chem. Commun. 2019, 100, 101–106. [Google Scholar] [CrossRef]
- Jani, D.K.; Goswami, S. Antidiabetic activity of Cassia angustifolia Vahl. and Raphanus sativus Linn. leaf extracts. J. Trad. Complement. Med. 2019, 10, 124–131. [Google Scholar] [CrossRef]
- Jani, D.; Goswami, S.S. Ameliorative effect of Raphanus sativus and Cassia angustifolia in experimentally induced hyperlipidemia and cardiovascular risk reduction. Int. J. PharmTech Res. 2017, 10, 273–279. [Google Scholar] [CrossRef]
- Banihani, S.A. Radish (Raphanus sativus) and diabetes. Nutrients 2017, 9, 1014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filgueira, F.A.R. Novo Manual de Olericultura: Agrotecnologia Moderna Na Produção e Comercialização de Hortaliças, 3rd ed.; UFV: Viçosa, Brazil, 2008. [Google Scholar]
- Basilio, A.; de Sousa, L.; da Silva, T.; de Moura, J.; Gonçalves, A.; de Melo Filho, J.; Leal, Y.; Dias, T. Morfofisiología del rábano (Raphanus sativus L.) bajo estrés salino y tratamientos con ácido ascórbico. Agron. Colomb. 2018, 36, 257–265. [Google Scholar]
- Parent, C.; Capelli, N.; Berger, A.; Crèvecoeur, M.; Dat, J.F. An overview of plant responses to soil waterlogging. Plant Stress 2008, 2, 20–27. [Google Scholar]
- Fischer, G.; Ramírez, F.; Casierra-Posada, F. Ecophysiological aspects of fruit crops in the era of climate change. A review. Agron. Colomb. 2016, 34, 190–199. [Google Scholar] [CrossRef]
- Cardona, W.A.; Bautista, L.G.; Flórez-Velasco, N.; Fischer, G. Desarrollo de la biomasa y raíz en plantas de lulo (Solanum quitoense var. septentrionale) en respuesta al sombrío y anegamiento. Rev. Colomb. Cienc. Hortíc. 2016, 10, 53–65. [Google Scholar] [CrossRef] [Green Version]
- Cassán, F.; Diaz-Zorita, M. Azospirillum sp. in current agriculture: From the laboratory to the field. Soil Biol. Biochem. 2016, 103, 117–130. [Google Scholar] [CrossRef]
- Barnawal, D.; Bharti, N.; Maji, D.; Chanotiya, C.S.; Kalra, A. 1-Aminocyclopropane-1-carboxylic acid (ACC) deaminase-containing rhizobacteria protect Ocimum sanctum plants during waterlogging stress via reduced ethylene generation. Plant Physiol. Biochem. 2012, 58, 227–235. [Google Scholar] [CrossRef]
- Olanrewaju, O.S.; Glick, B.R.; Babalola, O.O. Mechanisms of action of plant growth promoting bacteria. World J. Microbiol. Biotechnol. 2017, 33, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glick, B.R. Introduction to plant growth-promoting bacteria. In Beneficial Plant-Bacterial Interactions; Springer: Cham, Switzerland, 2020; pp. 1–37. [Google Scholar]
- Calzavara, A.K.; Paiva, P.H.G.; Gabriel, L.C.; Oliveira, A.L.M.; Milani, K.; Oliveira, H.C.; Bianchini, E.; Pimenta, J.A.; de Oliveira, M.C.N.; Stolf-Moreira, R. Associative bacteria influence maize (Zea mays L.) growth, physiology and root anatomy under different nitrogen levels. Plant Biol. 2018, 20, 870–878. [Google Scholar] [CrossRef]
- Camilios-Neto, D.; Bonato, P.; Wassem, R.; Tadra-Sfeir, M.Z.; Brusamarello-Santos, L.C.; Valdameri, G.; Donatti, L.; Faoro, H.; Weiss, V.A.; Chubatsu, L.S.; et al. Dual RNA-seq transcriptional analysis of wheat roots colonized by Azospirillum brasilense reveals up-regulation of nutrient acquisition and cell cycle genes. BMC Genom. 2014, 15, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Vidotti, M.S.; Lyra, D.H.; Morosini, J.S.; Granato, Í.S.C.; Quecine, M.C.; Azevedo, J.L.D.; Fritsche-Neto, R. Additive and heterozygous (dis) advantage GWAS models reveal candidate genes involved in the genotypic variation of maize hybrids to Azospirillum brasilense. PLoS ONE 2019, 14, e0222788. [Google Scholar] [CrossRef]
- Peng, H.; de-Bashan, L.E.; Higgins, B.T. Comparison of algae growth and symbiotic mechanisms in the presence of plant growth promoting bacteria and non-plant growth promoting bacteria. Algal Res. 2021, 53, 102156. [Google Scholar] [CrossRef]
- Bashan, Y.; de-Bashan, L.E. How the plant growth-promoting bacterium Azospirillum promotes plant growth—a critical assessment. In Advances in Agronomy; Academic Press: Cambridge, MA, USA, 2010; Volume 108, pp. 77–136. [Google Scholar]
- Bacilio, M.; Hernandez, J.P.; Bashan, Y. Restoration of giant cardon cacti in barren desert soil amended with common compost and inoculated with Azospirillum brasilense. Biol. Fertil. Soils 2006, 43, 112–119. [Google Scholar] [CrossRef]
- de-Bashan, L.E.; Hernandez, J.P.; Bashan, Y.; Maier, R.M. Bacillus pumilus ES4: Candidate plant growth-promoting bacterium to enhance establishment of plants in mine tailings. Environ. Exp. Bot. 2010, 69, 343–352. [Google Scholar] [CrossRef] [Green Version]
- Lopez, B.R.; Bashan, Y.; Trejo, A.; de-Bashan, L.E. Amendment of degraded desert soil with wastewater debris containing immobilized Chlorella sorokiniana and Azospirillum brasilense significantly modifies soil bacterial community structure, diversity, and richness. Biol. Fertil. Soils 2013, 49, 1053–1063. [Google Scholar] [CrossRef]
- Bashan, Y.; Levanony, H. An improved selection technique and medium for the isolation and enumeration of Azospirillum brasilense. Can. J. Microbiol. 1985, 31, 947–952. [Google Scholar] [CrossRef]
- de-Bashan, L.E.; Hernandez, J.P.; Bashan, Y. Interaction of Azospirillum spp. with microalgae: A basic eukaryotic-prokaryotic model and its biotechnological applications. In Handbook for Azospirillum; Springer: Cham, Switzerland, 2015; pp. 367–388. [Google Scholar]
- Flórez-Velasco, N.; Balaguera-López, H.E.; Restrepo-Díaz, H. Effects of foliar urea application on lulo (Solanum quitoense cv. septentrionale) plants grown under different waterlogging and nitrogen conditions. Sci. Hortic. 2015, 186, 154–162. [Google Scholar] [CrossRef]
- Ahmed, S.; Nawata, E.; Sakuratani, T. Changes of endogenous ABA and ACC, and their correlations to photosynthesis and water relations in mungbean (Vigna radiata (L.) Wilczak cv. KPS1) during waterlogging. Environ. Exp. Bot. 2006, 57, 278–284. [Google Scholar] [CrossRef]
- Cohen, A.C.; Bottini, R.; Piccoli, P.N. Azosporillum brasilense Sp 245 produces ABA in chemically-defined culture medium and increases ABA content in Arabidopsis plants. Plant Growth Regul. 2008, 54, 97–103. [Google Scholar] [CrossRef]
- Cassán, F.; Vanderleyden, J.; Spaepen, S. Physiological and agronomical aspects of phytohormone production by model plant-growth-promoting rhizobacteria (PGPR) belonging to the genus Azospirillum. J. Plant Growth Regul. 2014, 33, 440–459. [Google Scholar] [CrossRef]
- Ruiz-Sánchez, M.; Armada, E.; Muñoz, Y.; García-de Salamone, I.; Aroca, R.; Ruíz-Lozano, J.; Azcón, R. Azospirillum and arbuscular mycorrhizal colonization enhance rice growth and physiological traits under well-watered and drought conditions. J. Plant Physiol. 2011, 168, 1031–1037. [Google Scholar] [CrossRef] [PubMed]
- Villareal, A. Evaluación fisiológica de plantas de uchuva (Physalis peruviana L.), en la respuesta al estrés por anegamiento e infección de Fusarium oxysporum. In Tesis de Maestría Facultad de Ciencias Agrarias; Universidad Nacional de Colombia: Bogotá, Colombia, 2014. [Google Scholar]
- Yetisir, H.; Caliscan, M.; Soylu, S.; Sakar, M. Some physiological and growth responses of watermelon (Citrullus lanatus (Thunb.) Matsum. and Nakai) grafted onto Lagenaria siceraria to flooding. Environ. Exp. Bot. 2006, 58, 1–8. [Google Scholar] [CrossRef]
- Ren, B.; Zhang, J.; Dong, S.; Liu, P.; Zhao, B. Effects of waterlogging on leaf mesophyll cell ultrastructure and photosynthetic characteristics of summer maize. PLoS ONE 2016, 11, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Kumar, P.; Gupta, A.; Sharma, L.; Bakade, R. Mechanisms of Azospirillum in plant growth promotion. Sch. J. Agri. Vet. Sci. 2017, 4, 338–343. [Google Scholar]
- Davaran-Hagh, E.; Mirshekari, B.; Reza-Ardakani, M.; Farahvash, F.; Rejali, F. Azospirillum lipoferum and nitrogen fertilization effect on chlorophyll content, nutrients uptake and biometric properties of Zea mays L. Agrociencia 2015, 49, 889–897. [Google Scholar]
- Castro-Duque, N.E.; Chávez-Arias, C.C.; Restrepo-Díaz, H. Foliar glycine betaine or hydrogen peroxide sprays ameliorate waterlogging stress in cape gooseberry. Plants 2020, 9, 644. [Google Scholar] [CrossRef]
- Striker, G.G. Flooding stress on plants: Anatomical, morphological and physiological responses. Botany 2012, 1, 3–28. [Google Scholar]
- Jiménez-Suancha, S.C.; Alvarado, O.H.; Balaguera-López, H.E. Fluorescencia como indicador de estrés en Helianthus annuus L. Una revisión. Rev. Colomb. Cienc. Hortíc. 2015, 9, 149–160. [Google Scholar] [CrossRef] [Green Version]
- Maxwell, K.; Johnson, G. Chlorophyll fluorescence—A practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef]
- Nauman, J.; Young, D.; Anderson, J. Leaf chlorophyll fluorescence, reflectance, and physiological response to freshwater and saltwater flooding in the evergreen shrub, Myrica cerifera. Environ. Exp. Bot. 2008, 63, 402–409. [Google Scholar] [CrossRef]
- Liu, Z.; Cheng, R.; Xiao, W.; Guo, Q.; Wang, N. Effect of off-season flooding on growth, photosynthesis, carbohydrate partitioning, and nutrient uptake in Distylium chinense. PLoS ONE 2014, 9, 1–9. [Google Scholar] [CrossRef]
- Sánchez-Reinoso, A.D.; Jiménez-Pulido, Y.; Martínez-Pérez, J.P.; Pinilla, C.S.; Fischer, G. Parámetros de fluorescencia de la clorofila y otros parámetros fisiológicos como indicadores del estrés por anegamiento y sombrío en plántulas de lulo (Solanum quitoense var. septentrionale). Rev. Colomb. Cienc. Hortíc. 2019, 13, 325–335. [Google Scholar] [CrossRef]
- Else, M.; Janowiak, F.; Atkinson, C.; Jackson, M. Root signals and stomatal closure in relation to photosynthesis, chlorophyll a fluorescence and adventitious rooting of flooded tomato plants. Ann. Bot. 2009, 103, 313–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heidari, M.; Golpayegani, A. Effects of water stress and inoculation with plant growth promoting rhizobacteria (PGPR) on antioxidant status and photosynthetic pigments in basil (Ocimum basilicum L.). J. Saudy Soc. Agric. Sci. 2012, 11, 57–61. [Google Scholar] [CrossRef] [Green Version]
- Mohammed, G.H.; Zarco-Tejada, P.; Miller, J.R. Applications of chlorophyll fluorescence in forestry and ecophysiology. In Practical Applications of Chlorophyll Fluorescence in Plant Biology; Springer: Boston, MA, USA, 2003; pp. 79–124. [Google Scholar]
- Goltsev, V.; Kalaji, H.; Paunov, M.; Baba, W.; Horaczek, T.; Mojski, J.; Kociel, H.; Allakhverdiev, S. Variable chlorophyll fluorescence and its use for assessing physiological condition of plant photosynthetic apparatus. Russ. J. Plant Physiol. 2016, 63, 869–893. [Google Scholar] [CrossRef]
- Dall’Osto, L.; Holt, N.; Kaligotla, S.; Fuciman, M.; Cazzaniga, S.; Carbonera, D.; Frank, H.A.; Alric, J.; Basi, R. Zeaxanthin protects plant photosynthesis by modulating chlorophyll triplet yield in specific light-harvesting antenna subunits. J. Biol. Chem. 2012, 287, 41820–41834. [Google Scholar] [CrossRef] [Green Version]
- Velasco, N.F.; Ligarreto, G.A.; Díaz, H.R.; Fonseca, L.P.M. Photosynthetic responses and tolerance to root-zone hypoxia stress of five bean cultivars (Phaseolus vulgaris L.). S. Afr. J. Bot. 2019, 123, 200–207. [Google Scholar] [CrossRef]
- Azooz, M.; Ahmad, P. Plant Environment Interaction: Responses and Approaches to Mitigate Stress; Wiley-Blackwell: Hoboken, NJ, USA, 2015. [Google Scholar]
- Bashan, Y.; Bustillos, J.J.; Leyva, L.A.; Hernandez, J.P.; Bacilio, M. Increase in auxiliary photoprotective photosynthetic pigments in wheat seedlings induced by Azospirillum brasilense. Biol. Fertil. Soils 2006, 42, 279–285. [Google Scholar] [CrossRef]
- D’Angioli, A.; Gorne, R.; Lambers, H.; Frankland, A.; Silva, R. Inoculation with Azospirillum brasilense (Ab-V4, Ab-V5) increases Zea mays root carboxylate-exudation rates, dependent on soil phosphorus supply. Plant Soil 2017, 410, 499–507. [Google Scholar] [CrossRef]
- Méndez-Gómez, M.; Castro-Mercado, E.; García-Pineda, E. Azospirillum una rizobacteria con uso potencial en la agricultura. Biologicas 2014, 16, 11–18. [Google Scholar]
- Rood, S.; Nielsen, J.; Shenton, L.; Gill, K.; Letts, M. Effects of flooding on leaf development, transpiration, and photosynthesis in narrow leaf cottonwood, a willow-like poplar. Photosynth. Res. 2010, 104, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Baracaldo, A.; Carvajal, R.; Romero, A.; Prieto, A.; Garcia, F.; Fisher, G.; Miranda, D. El anegamiento afecta el crecimiento y producción de biomasa en tomate chonto (Solanum lycopersicum L.), cultivado bajo sombrío. Rev. Colomb. Cienc. Hortíc. 2014, 8, 92–102. [Google Scholar] [CrossRef] [Green Version]
- Aguirre-Medina, J.; Mendoza-López, A.; Cadena-Iniguez, J.; Avendaño-Arrazate, C. Efecto de la biofertilización en vivero del cacao (Theobroma cacao L) con Azospirillum brasilense tarrand, Krieg et Döbereiner y Glomus intraradices Schenk et Smith. Interciencia 2007, 32, 541–556. [Google Scholar]
- De la Cruz-Jiménez, J.; Moreno, P.; Magnitskiy, S. Respuesta de las plantas a estrés por inundación. Una revisión. Rev. Colomb. Cienc. Hortíc. 2012, 6, 96–109. [Google Scholar] [CrossRef] [Green Version]
- Pardos, J.A. Respuestas de las plantas al anegamiento del suelo. Invest Agrar. Sist. Recur. 2008, 13, 101–107. [Google Scholar] [CrossRef] [Green Version]
- Moreno, A.; Fischer, G. Efectos del anegamiento en los frutales. Una Revisión. Temas Agrar. 2014, 19, 106–123. [Google Scholar] [CrossRef] [Green Version]
- Aldana, F.; García, P.; Fischer, G. Effect of waterlogging stress on the growth, development and symptomatology of cape gooseberry (Physalis peruviana L.) plants. Rev. Acad. Colomb. Cienc. Ex. Fis. Nat. 2014, 38, 393–400. [Google Scholar] [CrossRef] [Green Version]
- Schramm, M.; Schaffer, B. Photosynthetic and growth responses of Eugenia uniflora L. seedlings to soil flooding and light intensity. Environ. Exp. Bot. 2010, 68, 113–121. [Google Scholar]
- Veresoglou, S.; Menexes, G. Impact of inoculation with Azospirillum spp. on growth properties and seed yield of wheat: A meta-analysis of studies in the ISI Web of Science from 1981 to 2008. Plant Soil 2010, 337, 469–480. [Google Scholar] [CrossRef]







{kind=link}
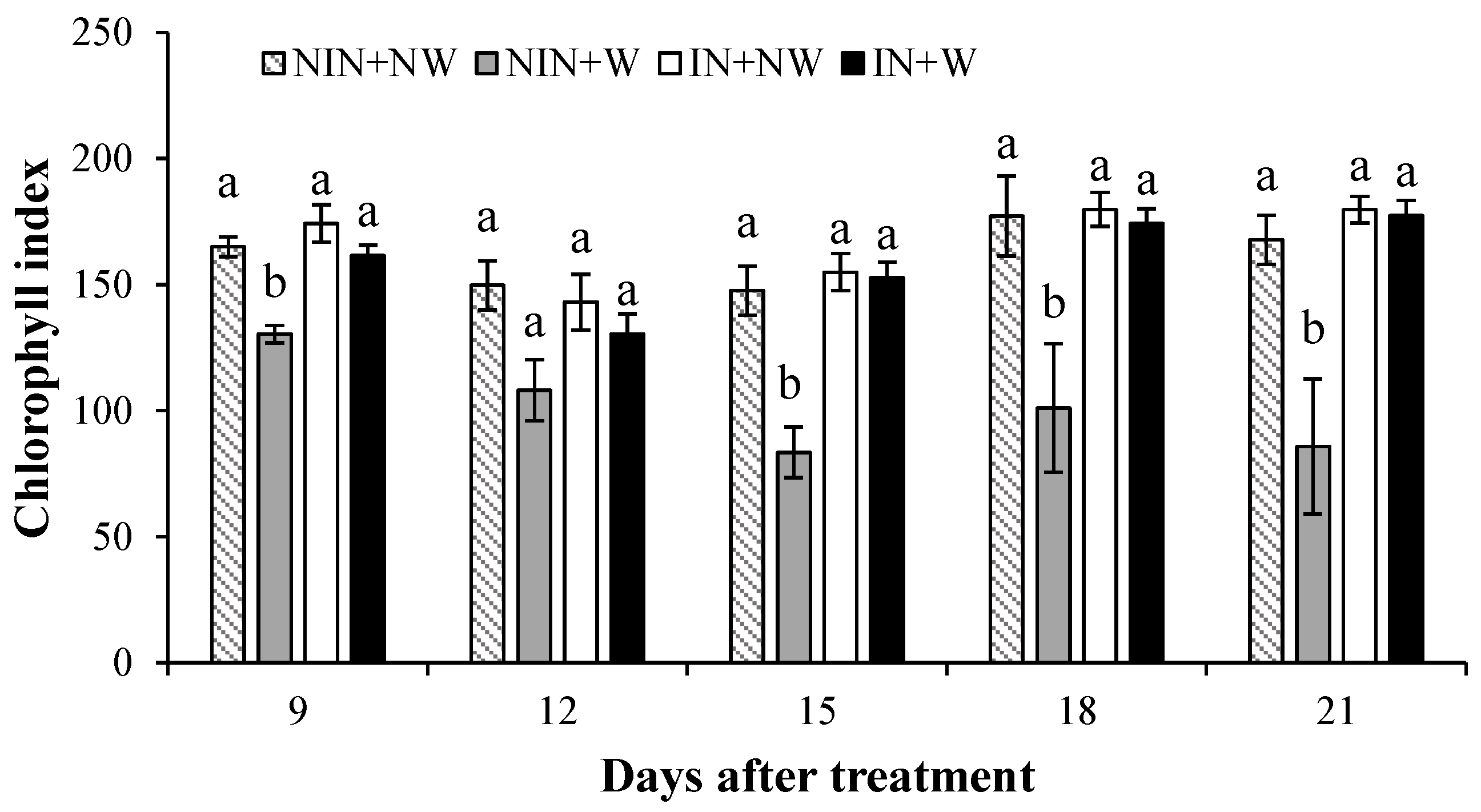{kind=link}
{kind=link}
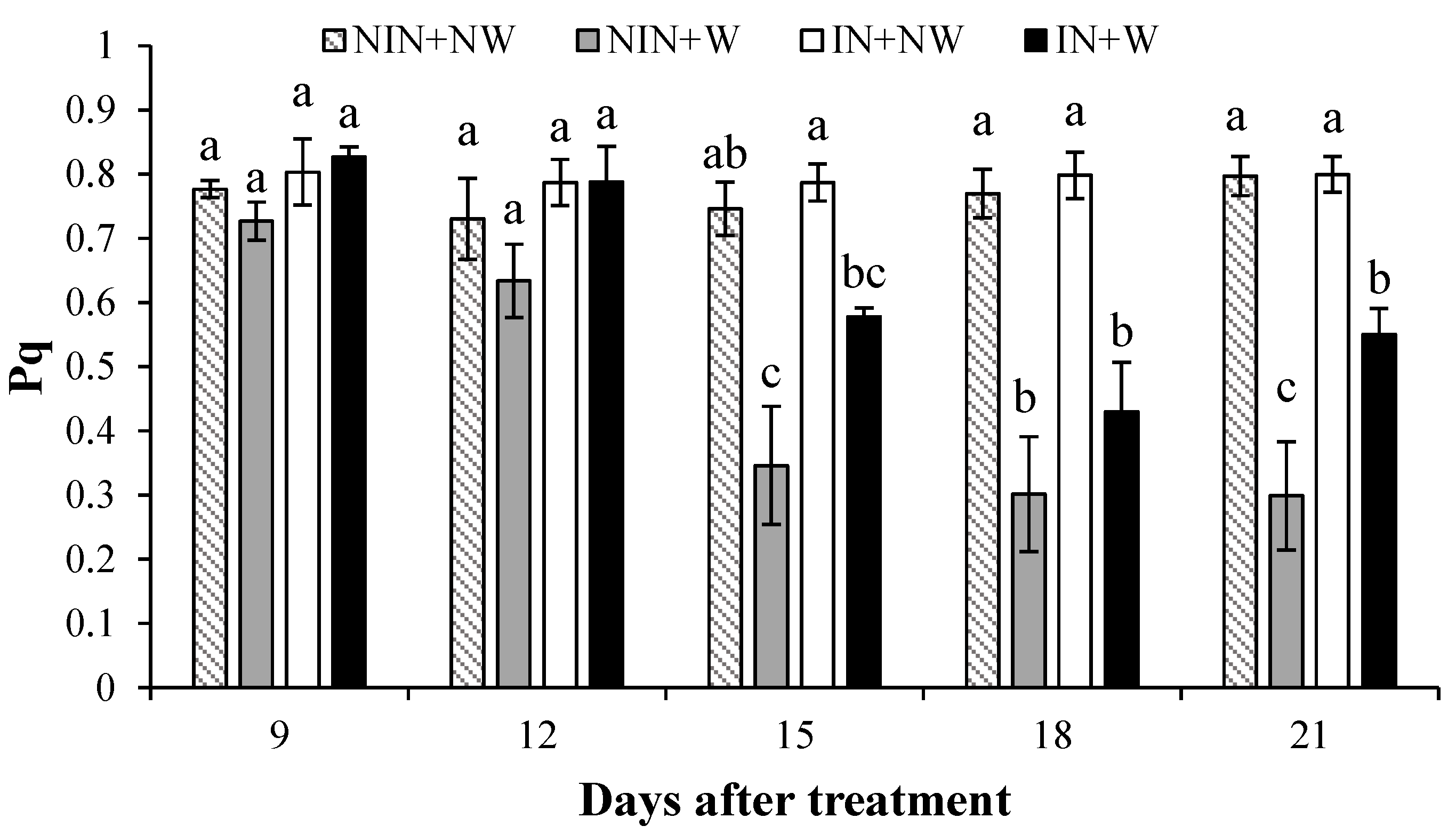{kind=link}
{kind=link}
{kind=link}
| Treatment | Definitions | Symbols |
|---|---|---|
| T1 | Non-inoculated, non-waterlogged (control) | NIN+NW |
| T2 | Non-inoculated, waterlogged | NIN+W |
| T3 | Inoculated, non-waterlogged | IN+NW |
| T4 | Inoculated, waterlogged | IN+W |
| Treatment | Diameter of Tuberous Root (mm) | Number of Leaves | Leaf Area (cm2) | Fresh Weight (g) | Dry Weight (g) |
|---|---|---|---|---|---|
| NIN+NW | 31.86 ± 4.15 a | 8.11 ± 0.56 a | 277.34 ± 37.06 a | 29.87 ± 2.07 ab | 1.90 ± 0.17 a |
| NIN+W | 12.08 ± 3.53 b | 4.67 ± 1.03 b | 86.03 ± 26.67 b | 3.42 ± 1.30 c | 0.16 ± 0.06 c |
| IN+NW | 37.53 ± 1.45 a | 9.33 ± 0.37 a | 274.22 ± 11.94 a | 34.62 ± 1.65 a | 1.83 ± 0.09 a |
| IN+W | 33.04 ± 1.64 a | 8.89 ± 0.26 a | 232.43 ± 14.69 a | 23.68 ± 2.78 b | 1.25 ± 0.17 b |
| Significance | * | * | * | * | * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salazar-Garcia, G.; Balaguera-Lopez, H.E.; Hernandez, J.P. Effect of Plant Growth-Promoting Bacteria Azospirillum brasilense on the Physiology of Radish (Raphanus sativus L.) under Waterlogging Stress. Agronomy 2022, 12, 726. https://doi.org/10.3390/agronomy12030726
Salazar-Garcia G, Balaguera-Lopez HE, Hernandez JP. Effect of Plant Growth-Promoting Bacteria Azospirillum brasilense on the Physiology of Radish (Raphanus sativus L.) under Waterlogging Stress. Agronomy. 2022; 12(3):726. https://doi.org/10.3390/agronomy12030726
Chicago/Turabian StyleSalazar-Garcia, Gisselle, Helber Enrique Balaguera-Lopez, and Juan Pablo Hernandez. 2022. "Effect of Plant Growth-Promoting Bacteria Azospirillum brasilense on the Physiology of Radish (Raphanus sativus L.) under Waterlogging Stress" Agronomy 12, no. 3: 726. https://doi.org/10.3390/agronomy12030726