Host and Tissue Affiliations of Culturable Endophytic Fungi Associated with Xerophytic Plants in the Desert Region of Northwest China


Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling Site
2.2. Plant Materials and Soil Sampling
2.3. Soil Physico-Chemical Characteristics
2.4. Isolation of Endophytic Fungi
2.5. Molecular Identification of Endophytic Fungi
2.6. Diversity of the Endophytic Fungal Community
2.7. Statistical Analysis
3. Results
3.1. Soil Properties
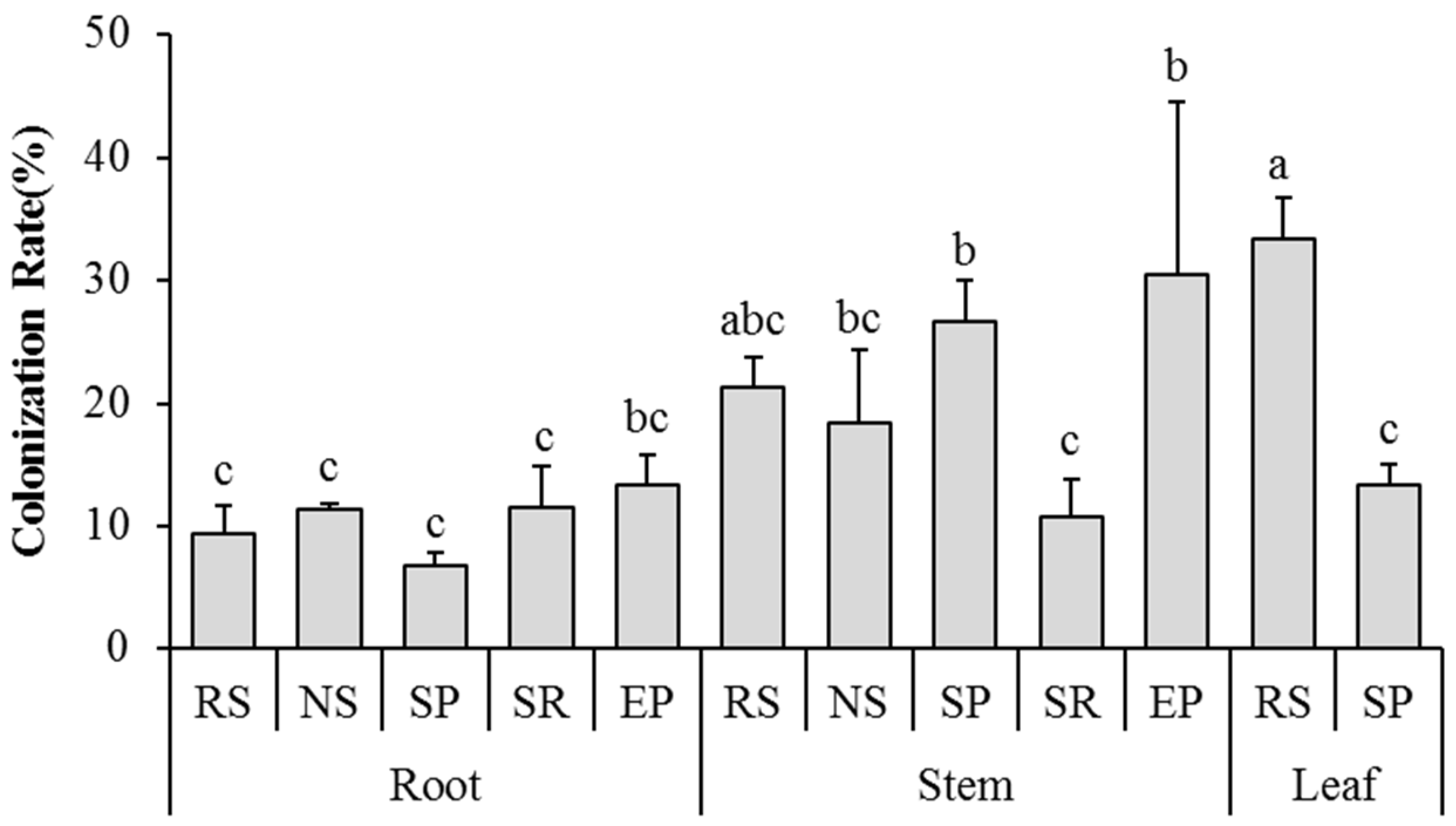
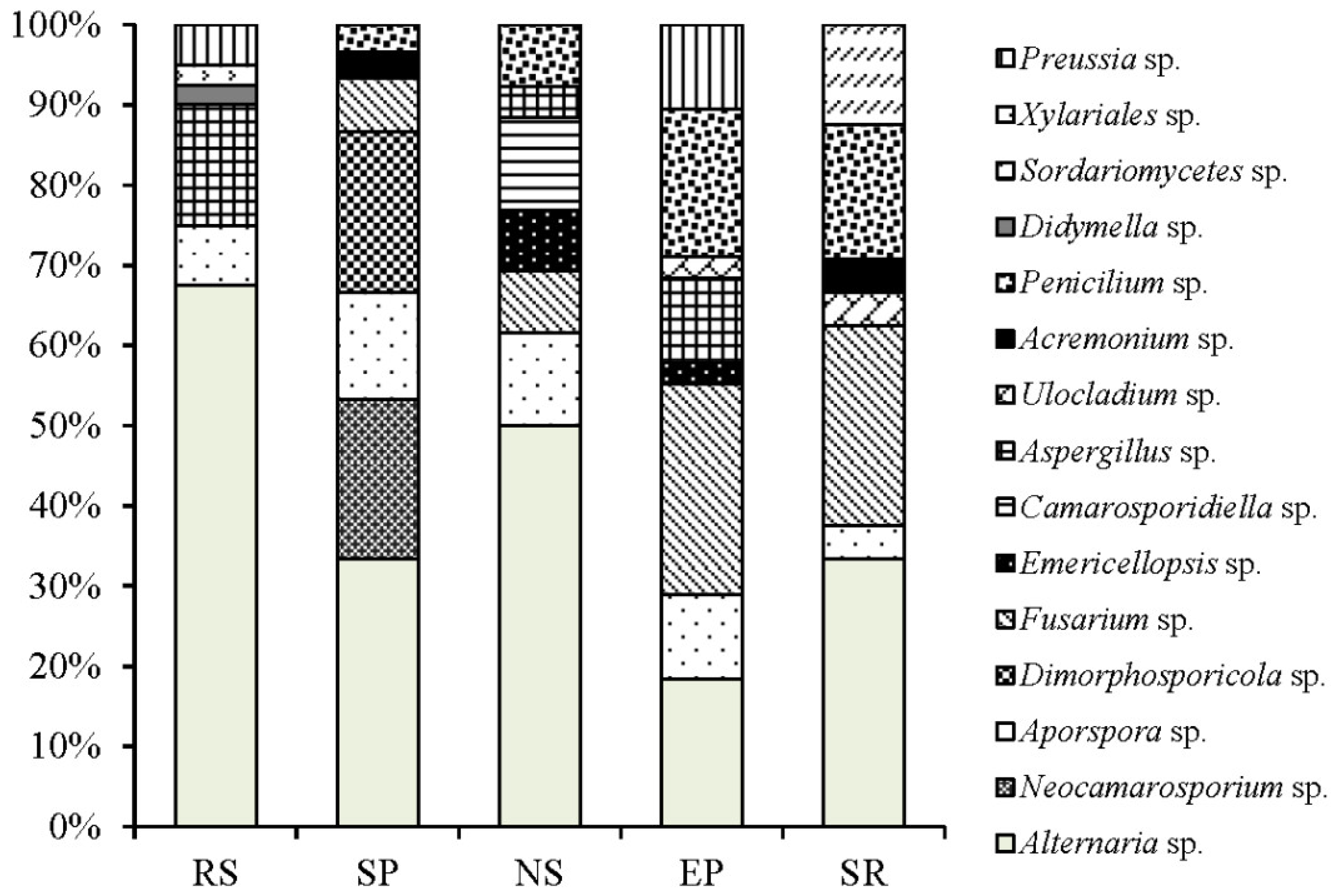
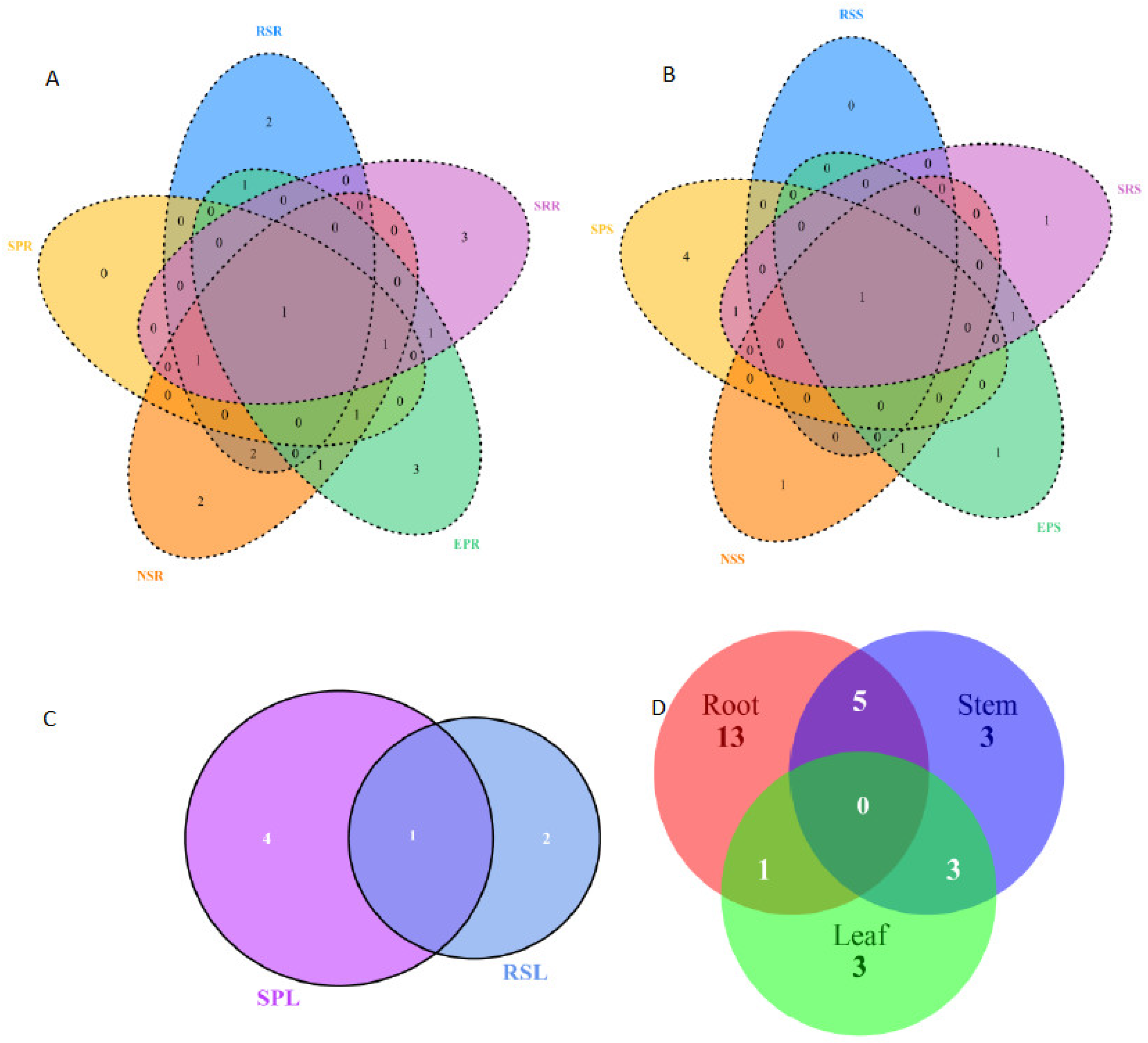
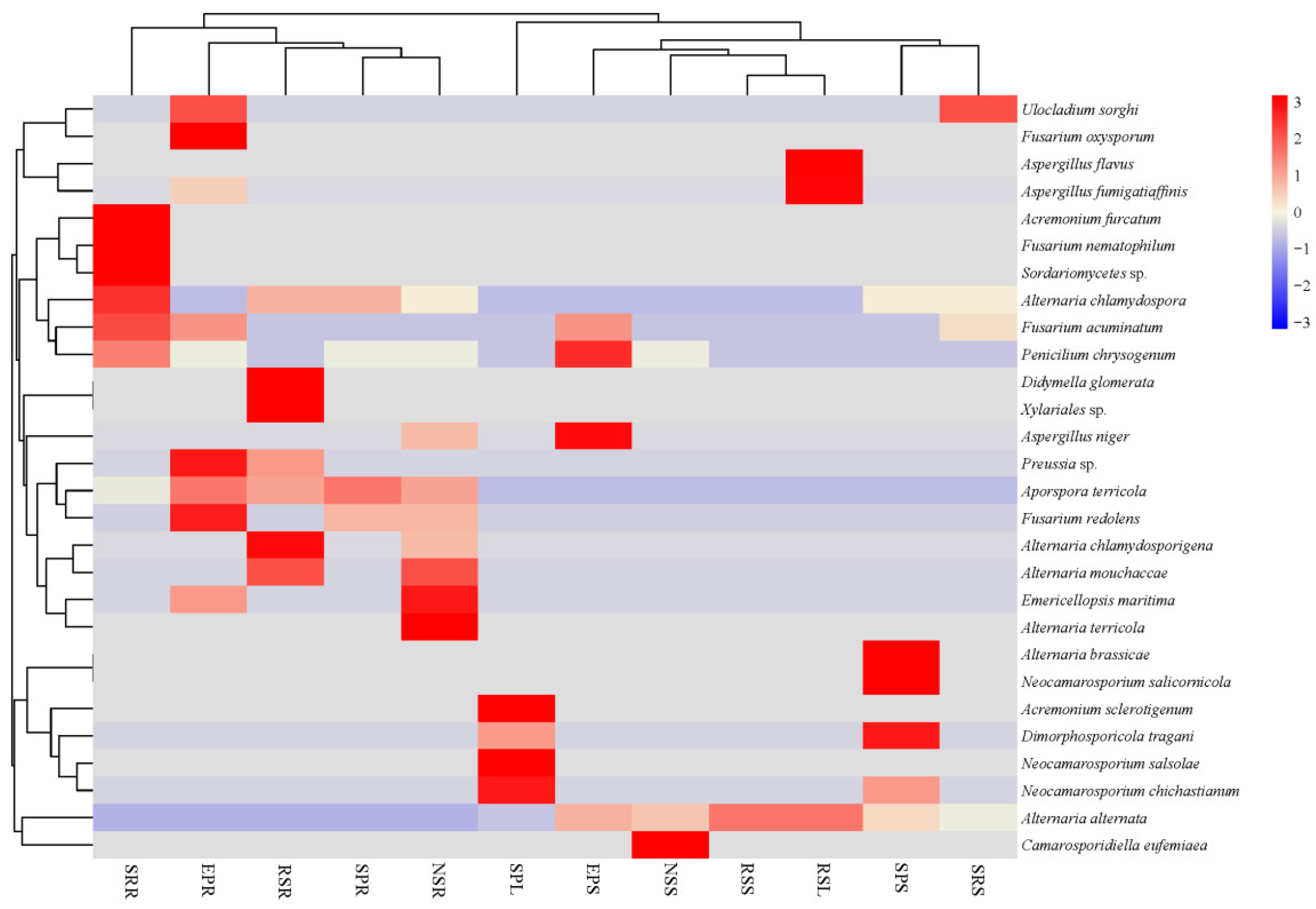
3.2. Community of Endophytic Fungi
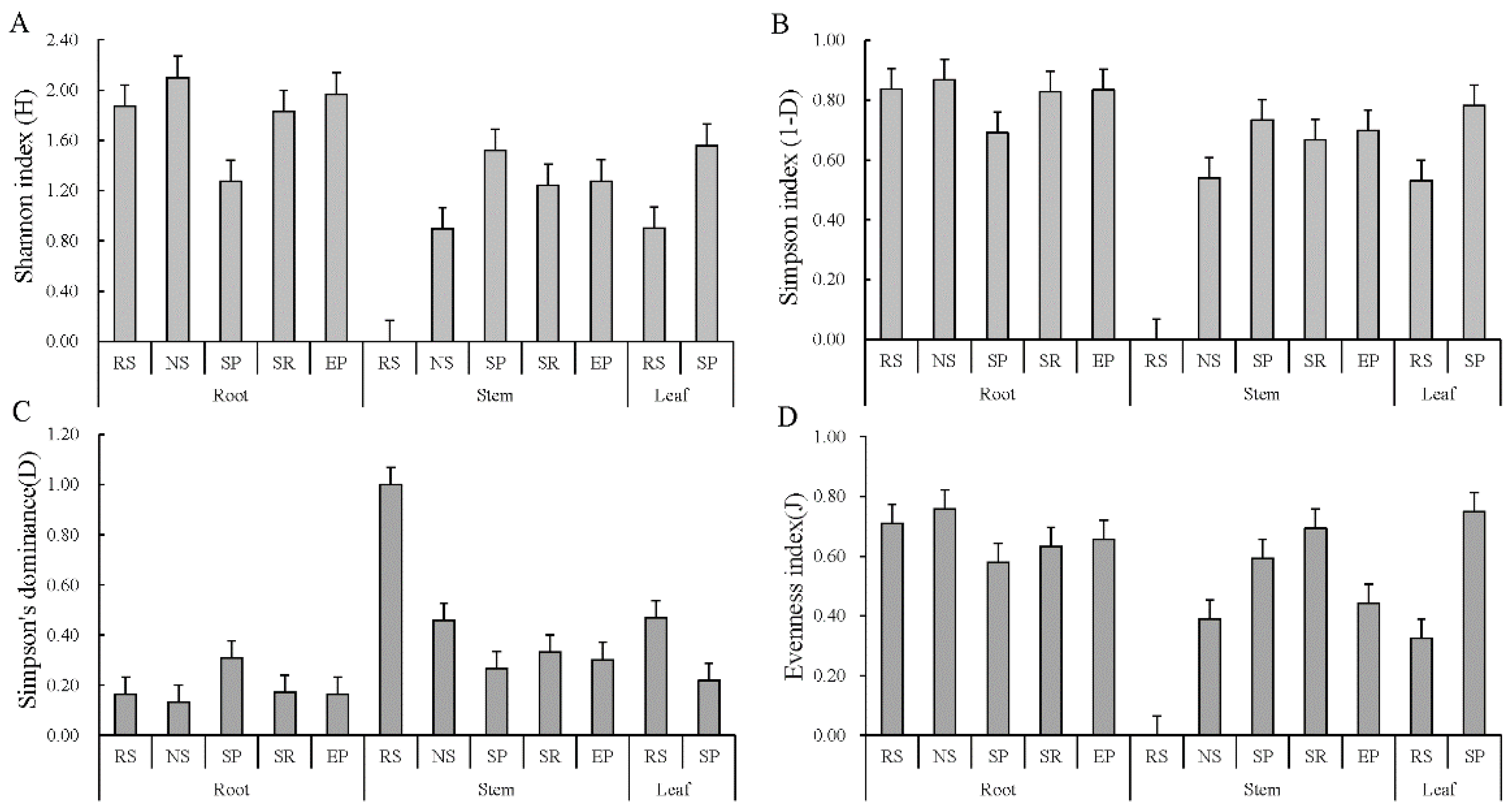
3.3. Diversity Indices of Endophytic Fungi
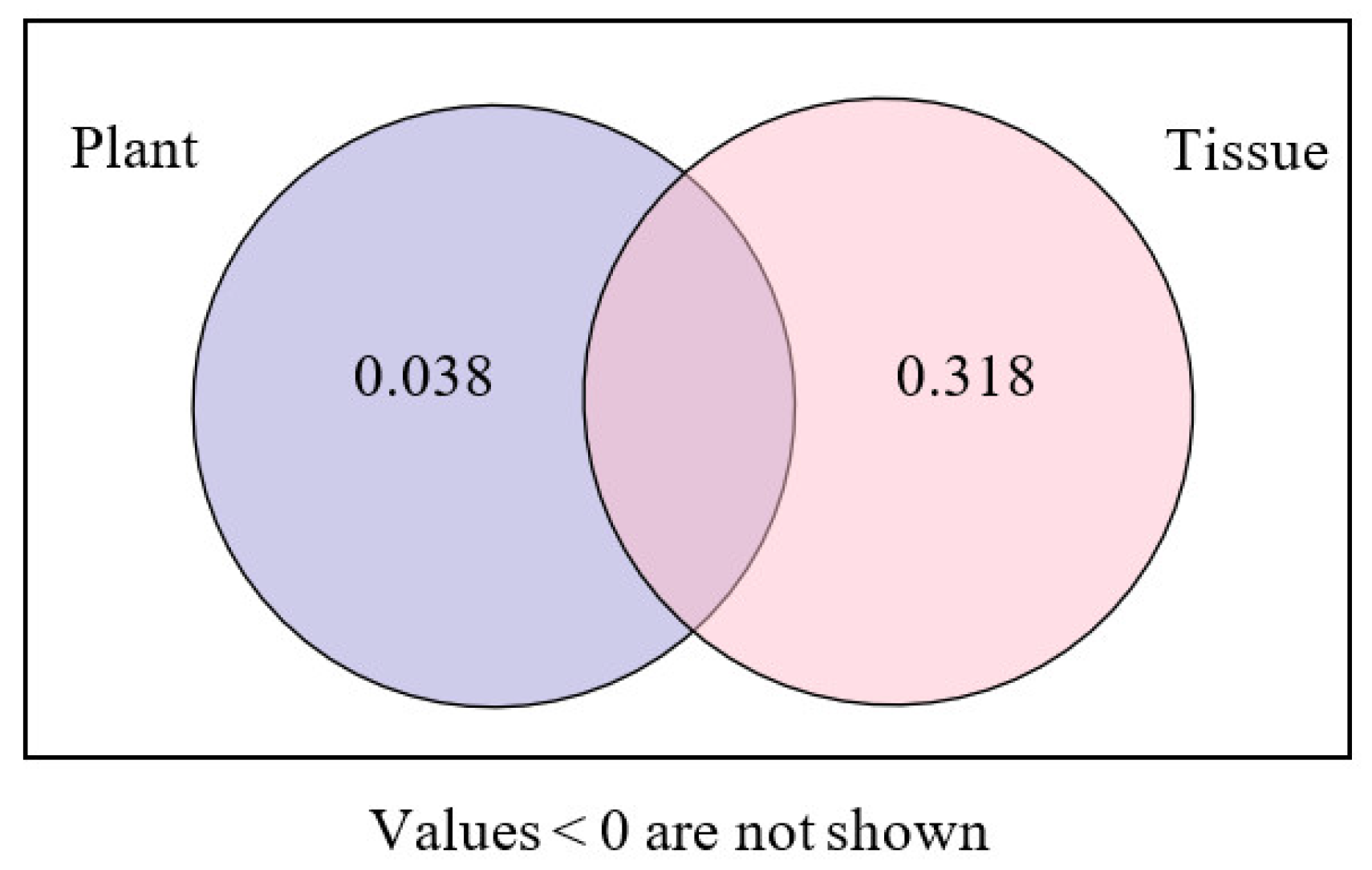
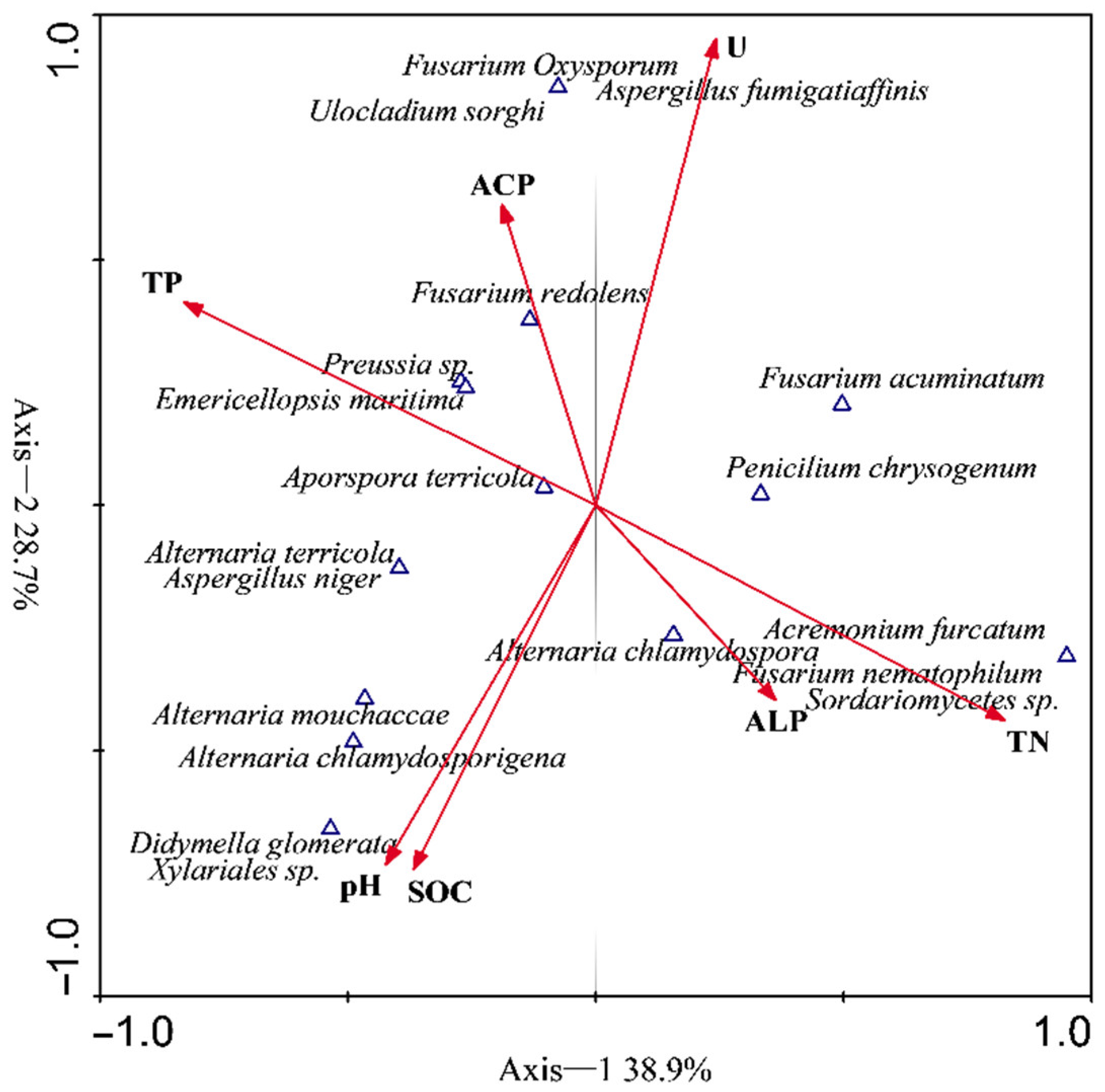
3.4. Variation Partitioning and Canonical Correlation Analysis
4. Discussion
4.1. Endophytic Fungal Community
4.2. Host Affiliations and Tissue Specificity
4.3. Root Endophytic Assemblages and Influence of Soil Factors
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Petrini, O. Fungal Endophytes of Tree Leaves. In Microbial Ecology of Leaves; Brock/Springer Series in Contemporary Bioscience; Andrews, J.H., Hirano, S.S., Eds.; Springer: New York, NY, USA, 1991; pp. 179–197. [Google Scholar]
- Rodriguez, R.J.; White, J.; Arnold, A.E.; Redman, R. Fungal endophytes, diversity and ecological roles. New Phytol. 2009, 182, 314–330. [Google Scholar] [CrossRef]
- Balla, K.M.; Troemel, E.R. Caenorhabditis elegansas a model for intracellular pathogen infection. Cell. Microbiol. 2013, 15, 1313–1322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoo, J.-H. Review of Disinfection and Sterilization—Back to the Basics. Infect. Chemother. 2018, 50, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Stewart, C.R.A.; Doilom, M.; Taylor, J.E. Analysis of fungal endophytes in Scottish Sitka spruce plantations shows extensive infections, novel host partners and gives insights into origins. For. Pathol. 2019, 49, e12471. [Google Scholar] [CrossRef] [Green Version]
- Arnold, A.E.; Miadlikowska, J.; Higgins, K.L.; Sarvate, S.D.; Gugger, P.; Way, A.; Hofstetter, V.; Kauff, F.; Lutzoni, F. A phylogenetic estimation of trophic transition networks for ascomycetous fungi: Are lichens cradles of symbiotrophic fungal diver-sification? Syst. Biol. 2009, 58, 283–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pickles, B.J.; Gorzelak, M.A.; Green, D.S.; Egger, K.N.; Massicotte, H.B. Host and habitat filtering in seedling root-associated fungal communities: Taxonomic and functional diversity are altered in ‘novel’ soils. Mycorrhiza 2015, 25, 517–531. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Yao, Y.-F. Endophytic Fungal Communities Associated with Vascular Plants in the High Arctic Zone Are Highly Diverse and Host-Plant Specific. PLoS ONE 2015, 10, e0130051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, A.; Verma, J.P. Does plant—Microbe interaction confer stress tolerance in plants: A review? Microbiol. Res. 2018, 207, 41–52. [Google Scholar] [CrossRef]
- Gunatilaka, L.A.A. Natural products from plant-associated microorganisms, Distribution, structural diversity, bioactivity and implications of their occurrence. J. Nat. Prod. 2006, 69, 509–526. [Google Scholar] [CrossRef] [Green Version]
- Ali, A.H.; Radwan, U.; El-Zayat, S.; El-Sayed, M.A. Desert plant-fungal endophytic association: The beneficial aspects to their hosts. Biol. Forum Int. J. 2018, 10, 138–145. [Google Scholar]
- Marasco, R.; Rolli, E.; Ettoumi, B.; Vigani, G.; Mapelli, F.; Borin, S.; Abou-Hadid, A.F.; El-Behairy, U.A.; Sorlini, C.; Cherif, A.; et al. A Drought Resistance-Promoting Microbiome Is Selected by Root System under Desert Farming. PLoS ONE 2012, 7, e48479. [Google Scholar] [CrossRef] [PubMed]
- Sieber, T.N.; Grünig, C.R. Fungal Root Endophytes. In Plant Roots—The Hidden Half, 4th ed.; Eshel, A., Beeckman, T., Eds.; CRC Press: Boca Raton, FL, USA, 2013; pp. 38–49. [Google Scholar]
- Loro, M.; Valero-Jiménez, C.; Nozawa, S.; Márquez, L. Diversity and composition of fungal endophytes in semiarid Northwest Venezuela. J. Arid Environ. 2012, 85, 46–55. [Google Scholar] [CrossRef]
- Hubbard, M.; Germida, J.; Vujanovic, V. Fungal endophytes improve wheat seed germination under heat and drought stress. Botany 2012, 90, 137–149. [Google Scholar] [CrossRef]
- Singh, L.P.; Gill, S.S.; Tuteja, N. Unraveling the role of fungal symbionts in plant abiotic stress tolerance. Plant Signal. Behav. 2011, 6, 175–191. [Google Scholar] [CrossRef] [Green Version]
- Fellbaum, C.R.; Mensah, J.A.; Cloos, A.J.; Strahan, G.E.; Pfeffer, P.E.; Kiers, T.; Bücking, H. Fungal nutrient allocation in common mycorrhizal networks is regulated by the carbon source strength of individual host plants. New Phytol. 2014, 203, 646–656. [Google Scholar] [CrossRef]
- Prieto, I.; Armas, C.; Pugnaire, F.I. Water release through plant roots: New insights into its consequences at the plant and ecosystem level. New Phytol. 2012, 193, 830–841. [Google Scholar] [CrossRef]
- Rilling, M.C.; Mummey, D.L. Mycorrhizas and soil structure. New Phytol. 2006, 171, 41–53. [Google Scholar] [CrossRef]
- Sikes, B.A.; Cottenie, K.; Klironomos, J.N. Plant and fungal identity deteimines pathogen protection of plant roots by arbuscular mycorrhizas. J. Ecol. 2009, 97, 1274–1280. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Ding, Q.; Hyde, K.; Guo, L. Community structure and preference of endophytic fungi of three woody plants in a mixed forest. Fungal Ecol. 2012, 5, 624–632. [Google Scholar] [CrossRef]
- White, J.F.; Torres, M.S.; Johnson, H.; Irizarry, I.; Tadych, M.A. Functional View of Plant Microbiomes: Endosymbiotic Systems That Enhance Plant Growth and Survival. In Advances in Endophytic Research; Verma, V., Gange, A., Eds.; Springer: New Delhi, India, 2013; pp. 425–439. [Google Scholar]
- Rodriguez, R.J.; Woodward, C.; Redman, R.S. Adaptation and Survival of Plants in High Stress Habitats via Fungal Endophyte Conferred Stress Tolerance. In Cellular Origin, Life in Extreme Habitats and Astrobiology; Seckbach, J., Grube, M., Eds.; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 2010; Volume 17, pp. 461–476. [Google Scholar]
- Fracchia, S.; Krapovickas, L.; Aranda-Rickert, A.; Valentinuzzi, V. Dispersal of arbuscular mycorrhizal fungi and dark septate endophytes by Ctenomys cf. knighti (Rodentia) in the northern Monte Desert of Argentina. J. Arid Environ. 2011, 75, 1016–1023. [Google Scholar] [CrossRef]
- Herrera, J.; Khidir, H.H.; Eudy, D.M.; Porras-Alfaro, A.; Natvig, D.O.; Sinsabaugh, R.L. Shifting fungal endophyte communities colonize Bouteloua gracilis: Effect of host tissue and geographical distribution. Mycologia 2010, 102, 1012–1026. [Google Scholar] [PubMed] [Green Version]
- Hou, L.; He, X.; Li, X.; Wang, S.; Zhao, L. Species composition and colonization of dark septate endophytes are affected by host plant species and soil depth in the Mu Us sandland, northwest China. Fungal Ecol. 2019, 39, 276–284. [Google Scholar] [CrossRef]
- Massimo, N.C.; Devan, M.M.N.; Arendt, K.R.; Wilch, M.H.; Riddle, J.M.; Furr, S.H.; Steen, C.; U’Ren, J.M.; Sandberg, D.C.; Arnold, A.E. Fungal Endophytes in Aboveground Tissues of Desert Plants: Infrequent in Culture, but Highly Diverse and Distinctive Symbionts. Microb. Ecol. 2015, 70, 61–76. [Google Scholar] [CrossRef] [PubMed]
- Del Olmo-Ruiz, M.; Arnold, A.E. Interannual variation and host affiliations of endophytic fungi associated with ferns at La Selva, Costa Rica. Mycologia 2014, 106, 8–21. [Google Scholar] [CrossRef]
- Sandberg, D.C.; Battista, L.J.; Arnold, A.E. Fungal Endophytes of Aquatic Macrophytes: Diverse Host-Generalists Characterized by Tissue Preferences and Geographic Structure. Microb. Ecol. 2014, 67, 735–747. [Google Scholar] [CrossRef]
- Bi, J.; He, D. Research advances in effects of plant on soil microbial diversity. Chin. Agric. Sci. Bull. 2009, 9, 244–250. [Google Scholar]
- Sardans, J.; Peñuelas, J. The Role of Plants in the Effects of Global Change on Nutrient Availability and Stoichiometry in the Plant-Soil System. Plant Physiol. 2012, 160, 1741–1761. [Google Scholar] [CrossRef] [Green Version]
- Eschen, R.; Hunt, S.; Mykura, C.; Gange, A.C.; Sutton, B.C. The foliar endophytic fungal community composition in Cirsium arvense is affected by mycorrhizal colonization and soil nutrient content. Fungal Biol. 2010, 114, 991–998. [Google Scholar] [CrossRef]
- David, A.S.; Seabloom, E.W.; May, G. Plant Host Species and Geographic Distance Affect the Structure of Aboveground Fungal Symbiont Communities, and Environmental Filtering Affects Belowground Communities in a Coastal Dune Ecosystem. Microb. Ecol. 2016, 71, 912–926. [Google Scholar] [CrossRef]
- Hartley, S.E.; Gange, A.C. Impacts of Plant Symbiotic Fungi on Insect Herbivores: Mutualism in a Multitrophic Context. Annu. Rev. Entomol. 2009, 54, 323–342. [Google Scholar] [CrossRef]
- Liu, D.; Huang, Y.; Sun, H.; An, S. The restoration age of Robinia pseudoacacia plantation impacts soil microbial biomass and microbial community structure in the Loess Plateau. Catena 2018, 165, 192–200. [Google Scholar] [CrossRef]
- Strobel, G.A. Methods of discovery and techniques to study endophytic fungi producing fuel-related hydrocarbons. Nat. Prod. Rep. 2014, 31, 259–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Shi, S.; Lin, F.; Jiang, P. Response of the soil fungal community to multi-factor environmental changes in a temperate forest. Appl. Soil Ecol. 2014, 81, 45–56. [Google Scholar] [CrossRef]
- Liu, Y.B.; Li, X.R.; Li, M.M.; Liu, D.; Zhang, W.L. Leaf (or assimilation branch) epidermal micromorphology of desert plant in arid and semi-arid areas of China. Chin. J. Plant Ecol. 2016, 40, 1189–1207. [Google Scholar]
- Zhang, J.; Wu, B.; Zhu, Y.; Li, Y.; Lu, Q.; Yao, B. Responses of Nitraria tangutorum to water and photosynthetic physiology in rain enrichment scenario. Acta Ecol. Sin. 2013, 33, 172–177. [Google Scholar] [CrossRef]
- Li, S.; Su, P.; Zhang, H.; Zhou, Z.; Shi, R.; Gou, W. Hydraulic Conductivity Characteristics of Desert Plant Organs: Coping with Drought Tolerance Strategy. Water 2018, 10, 1036. [Google Scholar] [CrossRef] [Green Version]
- Alsharif, W.; Saad, M.M.; Hirt, H. Desert microbes for boosting sustainable sgriculture in extreme environments. Front. Microbiol. 2020, 11, 1666. [Google Scholar] [CrossRef]
- Lugo, M.A.; Reinhart, K.; Menoyo, E.; Crespo, E.M.; Urcelay, C. Plant functional traits and phylogenetic relatedness explain variation in associations with root fungal endophytes in an extreme arid environment. Mycorrhiza 2014, 25, 85–95. [Google Scholar] [CrossRef]
- Rodrigues, A.; Bacci, M.; Mueller, U.G.; Ortiz, A.; Pagnocca, F.C. Microfungal “weeds” in the leafcutter ant symbiosis. Microb. Ecol. 2008, 56, 604–614. [Google Scholar] [CrossRef]
- Wang, J.J.; Qiang, W.; Liu, H.Y.; Ge, J.L.; Zuo, Y.L.; He, X.L. Effects of plants pecies on diversity of arbuscular mycorrhizal fungi in extremely arid desert environment. Mycosystema 2017, 36, 861–869. [Google Scholar]
- Kimmins, J.P. Forest Ecology; China Forestry Publishing House: Beijing, China, 2005; Volume 258, pp. 17–43. [Google Scholar]
- Heiri, O.; Lotter, A.F.; Lemcke, G. Loss on ignition as a method for estimating organic and carbonate content in sediments: Reproducibility and comparability of results. J. Paleolimnol. 2001, 25, 101–110. [Google Scholar] [CrossRef]
- Bao, S.D. Agrochemical Analysis of Soil; Chinese Agricultural Press: Beijing, China, 2000; pp. 44–49. (In Chinese) [Google Scholar]
- Hoffmann, G.G.; Teicher, K. A colorimetric technique for determining urease activity in soil. Dung Boden 1961, 95, 55–63. [Google Scholar] [CrossRef]
- Tarafdar, J.C.; Marschner, H. Phosphatase activity in the rhizosphere and hyphosphere of VA mycorrhizal wheat supplied with inorganic and organic phosphorus. Soil Biol. Biochem. 1994, 26, 387–395. [Google Scholar] [CrossRef]
- Zhan, F.; He, Y.; Li, T.; Yang, Y.-Y.; Toor, G.S.; Zhao, Z. Tolerance and Antioxidant Response of a Dark Septate Endophyte (DSE), Exophiala pisciphila, to Cadmium Stress. Bull. Environ. Contam. Toxicol. 2014, 94, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Parmar, S.; Li, Q.; Wu, Y.; Li, X.; Yan, J.; Sharma, V.K.; Wei, Y.; Li, H. Endophytic fungal community of Dysphania ambrosioides from two heavy metal-contaminated sites: Evaluated by culture-dependent and culture-independent approaches. Microb. Biotechnol. 2018, 11, 1170–1183. [Google Scholar] [CrossRef] [PubMed]
- Maadon, S.N.; Wakid, S.A.; Zainudin, I.I.; Rusli, L.S.; Mohd zan, M.S.; Hasan, N.A.; Abu Shah, N.A.; Rohani, E.R. Isolation and identification of endophytic fungi from UiTM reserve forest, Negeri Sembilan. Sains Malays. 2018, 47, 3025–3030. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J.W. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In PCR Protocols, a Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press, Inc.: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6, molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Z.L.; Rao, L.B.; Chen, Y.C.; Zhang, C.L.; Wu, Y.G. From pattern to process, species and functional diversity in fungal endophytes of Abies beshanzuensis. Fungal Biol. 2011, 115, 197–213. [Google Scholar] [CrossRef]
- Shannon, C.E.; Weaver, W. The Mathematical Theory of Communication; University of Illinois Press: Urbana, IL, USA, 1949. [Google Scholar]
- Simpson, E.H. Measurement of species diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Caruso, T.; Pigino, G.; Bernini, F.; Bargagli, R.; Migliorini, M. The Berger–Parker Index as an Effective Tool for Monitoring the Biodiversity of Disturbed Soils: A Case Study on Mediterranean Oribatid (Acari: Oribatida) Assemblages. In Biodiversity and Conservation in Europe; Hawksworth, D.L., Bull, A.T., Eds.; Springer: Dordrecht, The Netherlands, 2007; Volume 7, pp. 35–43. [Google Scholar]
- Samaga, P.V.; Rai, V.R. Diversity and bioactive potential of endophytic fungi from Nothapodytes foetida, Hypericum mysorense and Hypericum japonicum collected from Western Ghats of India. Ann. Microbiol. 2016, 66, 229–244. [Google Scholar] [CrossRef]
- Borcard, D.; Gillet, F.; Legendre, P. Numerical Ecology with R; Springer: New York, NY, USA, 2011. [Google Scholar]
- Luo, J.; Walsh, E.; Miller, S.; Blystone, D.; Dighton, J.; Zhang, N. Root endophytic fungal communities associated with pitch pine, switchgrass, and rosette grass in the pine barrens ecosystem. Fungal Biol. 2017, 121, 478–487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johansen, R.B.; Johnston, P.; Mieczkowski, P.; Perry, G.L.W.; Robeson, M.S.; Vilgalys, R.; Burns, B.R. Scattered far and wide: A broadly distributed temperate dune grass finds familiar fungal root associates in its invasive range. Soil Biol. Biochem. 2017, 112, 177–190. [Google Scholar] [CrossRef]
- Bonfim, J.A.; Vasconcellos, R.L.F.; Baldesin, L.F.; Sieber, T.N.; Cardoso, E.J.B.N. Dark septate endophytic fungi of native plants along an altitudinal gradient in the Brazilian Atlantic forest. Fungal Ecol. 2016, 20, 202–210. [Google Scholar] [CrossRef]
- Arnold, A.E.; Lutzoni, F. Diversity and host range of foliar fungal endophytes: Are tropical leaves biodiversity hotspots? Ecology 2007, 88, 541–549. [Google Scholar] [CrossRef]
- Zhang, T.; Zhang, Y.Q.; Liu, H.Y.; Wei, Y.Z.; Li, H.L.; Su, J.; Zhao, L.X.; Yu, L.Y. Diversity and cold adaptation of culturable endophytic fungi from bryophytes in the Fildes Region, King George Island, maritime Antarctica. FEMS Microbiol. Lett. 2013, 341, 52–61. [Google Scholar] [CrossRef] [Green Version]
- Rosa, L.H.; Vaz, A.B.M.; Caligiorne, R.B.; Campolina, S.; Rosa, C.A. Endophytic fungi associated with the Antarctic grass Deschampsia antarctica Desv. (Poaceae). Polar Biol. 2009, 32, 161–167. [Google Scholar] [CrossRef] [Green Version]
- El-Zayat, S.A.; Nassar, M.S.M.; El-Hissy, F.T.; Abdel-Motaal, F.F.; Ito, S. Mycoflora associated with Hyoscyamus muticus growing under an extremely arid desert environment (Aswan region, Egypt). J. Basic Microb. 2008, 48, 82–92. [Google Scholar] [CrossRef]
- Porras-Alfaro, A.; Herrera, J.; Sinsabaugh, R.L.; Odenbach, K.J.; Lowrey, T.; Natvig, D.O. Novel root fungal consortium associated with a dominant desert grass. Appl. Environ. Microb. 2008, 74, 2805–2813. [Google Scholar] [CrossRef] [Green Version]
- Xie, L.; He, X.; Wang, K.; Hou, L.; Sun, Q. Spatial dynamics of dark septate endophytes in the roots and rhizospheres of Hedysarum scoparium in northwest China and the influence of edaphic variables. Fungal Ecol. 2017, 26, 135–143. [Google Scholar] [CrossRef]
- Qiang, W.; He, X.; Wang, J.; Zhao, L. Temporal and spatial variation of arbuscular mycorrhizal fungi under the canopy of Hedysarum scoparium in the northern desert, China. Appl. Soil Ecol. 2019, 136, 139–147. [Google Scholar] [CrossRef]
- Arnold, A.E.; Herre, E.A. Canopy cover and leaf age affect colonization by tropical fungal endophytes: Ecological pattern and process in Theobroma cacao (Malvaceae). Mycologia 2003, 95, 388–398. [Google Scholar] [CrossRef] [PubMed]
- Bayman, P.; Angulo-Sandoval, P.; Báez-Ortiz, Z.; Lodge, D.J. Distribution and dispersal of Xylaria endophytes in two tree species in Puerto Rico. Mycol. Res. 1998, 102, 944–948. [Google Scholar] [CrossRef]
- Zuo, Y.; He, C.; He, X.; Li, X.; Xue, Z.; Li, X.; Wang, S. Plant cover of Ammopiptanthus mongolicus and soil factors shape soil microbial community and catabolic functional diversity in the arid desert in Northwest China. Appl. Soil Ecol. 2020, 147, 103389. [Google Scholar] [CrossRef]
- Lau, M.K.; Arnold, A.E.; Johnson, N.C. Factors influencing communities of foliar fungal endophytes in riparian woody plants. Fungal Ecol. 2013, 6, 365–378. [Google Scholar] [CrossRef]
- U’Ren, J.M.; Lutzoni, F.; Miadlikowska, J.; Laetsch, A.D.; Arnold, A.E. Host and geographic structure of endophytic and endolichenic fungi at a continental scale. Am. J. Bot. 2012, 99, 898–914. [Google Scholar] [CrossRef] [Green Version]
- Knapp, D.; Kovacs, G.M.; Zajta, E.; Groenewald, J.; Crous, P. Dark septate endophytic pleosporalean genera from semiarid areas. Persoonia Mol. Phylogeny Evol. Fungi 2015, 35, 87–100. [Google Scholar] [CrossRef] [Green Version]
- Jia, T.; Wang, R.; Fan, X.; Chai, B. A Comparative Study of Fungal Community Structure, Diversity and Richness between the Soil and the Phyllosphere of Native Grass Species in a Copper Tailings Dam in Shanxi Province, China. Appl. Sci. 2018, 8, 1297. [Google Scholar] [CrossRef] [Green Version]
- Gond, S.K.; Mishra, A.; Sharma, V.K.; Verma, S.K.; Kumar, J.; Kharwar, R.N.; Kumar, A. Diversity and antimicrobial activity of endophytic fungi isolated from Nyctanthes arbor-tristis, a well-known medicinal plant of India. Mycoscience 2012, 53, 113–121. [Google Scholar] [CrossRef]
- Muhsin, T.M.; Booth, T. Fungi associated with halophytes of an inland salt marsh, Manitoba, Canada. Can. J. Bot. 1987, 65, 1137–1151. [Google Scholar] [CrossRef]
- Petrini, O.; Fisher, P. Fungal endophytes in Salicornia perennis. Trans. Br. Mycol. Soc. 1986, 87, 647–651. [Google Scholar] [CrossRef]
- Moghaddam, M.S.H.; Safaie, N.; Soltani, J.; Hagh-Doust, N. Desert-adapted fungal endophytes induce salinity and drought stress resistance in model crops. Plant Physiol. Biochem. 2021, 160, 225–238. [Google Scholar] [CrossRef] [PubMed]
- Han, L.; Shi, J.; He, C.; He, X. Temporal and Spatial Dynamics of Dark Septate Endophytes in the Roots of Lycium ruthenicum in the Desert Region of Northwest China. Agronomy 2021, 11, 648. [Google Scholar] [CrossRef]
- Li, J.-L.; Sun, X.; Zheng, Y.; Lü, P.-P.; Wang, Y.-L.; Guo, L.-D. Diversity and community of culturable endophytic fungi from stems and roots of desert halophytes in northwest China. MycoKeys 2020, 62, 75–95. [Google Scholar] [CrossRef] [PubMed]
- Jalili, B.; Bagheri, H.; Azadi, S.; Soltani, J. Identification and salt tolerance evaluation of endophyte fungi isolates from halophyte plants. Int. J. Environ. Sci. Technol. 2020, 17, 3459–3466. [Google Scholar] [CrossRef]
- Abdel-Hafez, S.I.I. Halophilic fungi of desert soils in Saudi Arabia. Mycopathol. Mycol. Appl. 1981, 75, 75–80. [Google Scholar] [CrossRef]
- Ameen, F.; Stephenson, S.L.; AlNadhari, S.; Yassin, M.A. Isolation, identification and bioactivity analysis of an endophytic fungus isolated from Aloe vera collected from Asir desert, Saudi Arabia. Bioprocess Biosyst. Eng. 2021, 44, 1063–1070. [Google Scholar] [CrossRef]
- Fonseca-García, C.; Coleman-Derr, D.; Garrido, E.; Visel, A.; Tringe, S.G.; Partida-Martínez, L.P. The cacti microbiome: Inter-play between habitat-filtering and host-specificity. Front. Microbiol. 2016, 7, 150. [Google Scholar] [CrossRef]
- Abdelfattah, A.; Wisniewski, M.; Schena, L.; Tack, A.J.M. Experimental evidence of microbial inheritance in plants and transmission routes from seed to phyllosphere and root. Environ. Microbiol. 2021, 23, 2199–2214. [Google Scholar] [CrossRef]
- Gautam, A.K. Diversity of fungal endophytes in some medicinal plants of Himachal Pradesh, India. Arch. Phytopathol. Plant Prot. 2014, 47, 537–544. [Google Scholar] [CrossRef]
- Wenzel, J.B.; García, A.; Filho, C.J.R.; Prioli, A.J.; Pamphile, J.A. Evaluation of foliar fungal endophyte diversity and colonization of medicinal plant Luehea divaricata (Martius et Zuccarini). Biol. Res. 2010, 43, 375–384. [Google Scholar] [CrossRef] [Green Version]
- Zhou, D.; Hyde, K.D. Host-specificity, host-exclusivity, and host-recurrence in saprobic fungi. Mycol. Res. 2001, 105, 1449–1457. [Google Scholar] [CrossRef]
- Sun, X.; Guo, L.-D.; Hyde, K.D. Community composition of endophytic fungi in Acer truncatum and their role in decomposition. Fungal Divers. 2011, 47, 85–95. [Google Scholar] [CrossRef]
- Rodriguez, R.; Redman, R. Fungal Life-Styles and Ecosystem Dynamics: Biological Aspects of Plant Pathogens, Plant Endophytes and Saprophytes. Adv. Bot. Res. 1997, 24, 169–193. [Google Scholar] [CrossRef]
- Suryanarayanan, T.S.; Thirunavukkarasu, N.; Govindarajulu, M.B.; Gopalan, V. Fungal endophytes: An untapped source of biocatalysts. Fungal Divers. 2012, 54, 19–30. [Google Scholar] [CrossRef]
- Kumar, D.S.S.; Hyde, K.D. Biodiversity and tissue-recurrence of endophytic fungi in Tripterygium wilfordii. Fungal Divers. 2004, 17, 69–90. [Google Scholar]
- Tahtamouni, M.E.; Khresat, S.; Lucero, M.; Sigala, J.; Unc, A. Diversity of endophytes across the soil-plant continuum for Atriplex spp. in arid environments. J. Arid Land 2015, 8, 241–253. [Google Scholar] [CrossRef]
- Aly, A.H.; Debbab, A.; Proksch, P. Fungal endophytes: Unique plant inhabitants with great promises. Appl. Microbiol. Biotechnol. 2011, 90, 1829–1845. [Google Scholar] [CrossRef]
- Wu, L.; Han, T.; Li, W.; Jia, M.; Xue, L.; Rahman, K.; Qin, L. Geographic and tissue influences on endophytic fungal communities of Taxus chinensis var. mairei in China. Curr. Microbiol. 2012, 66, 40–48. [Google Scholar] [CrossRef]
- Knapp, D.G.; Kovács, G.M. Interspecific metabolic diversity of root-colonizing endophytic fungi revealed by enzyme activity tests. FEMS Microbiol. Ecol. 2016, 92, 190. [Google Scholar] [CrossRef] [Green Version]
- Petrini, O.; Sieber, T.N.; Toti, L.; Viret, O. Ecology, metabolite production, and substrate utilization in endophytic fungi. Nat. Toxins 1993, 1, 185–196. [Google Scholar] [CrossRef]
- Schardl, C.L.; Leuchtmann, A.; Spiering, M.J. Symbioses of grasses with seed borne fungal endophytes. Annu. Rev. Plant Biol. 2004, 55, 315–340. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.Y.; Spiering, M.J.; Scott, V.; Lane, G.A.; Christensen, M.J.; Schmid, J. In Planta Regulation of Extension of an Endophytic Fungus and Maintenance of High Metabolic Rates in Its Mycelium in the Absence of Apical Extension. Appl. Environ. Microbiol. 2001, 67, 5377–5383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; He, X.; He, C.; Chen, Y.; Wang, X. Spatial dynamics of dark septate endophytes and soil factors in the rhizosphere of Ammopiptanthus mongolicus in Inner Mongolia, China. Symbiosis 2015, 65, 75–84. [Google Scholar] [CrossRef]
- Li, X.; He, X.; Hou, L.; Ren, Y.; Wang, S.; Su, F. Dark septate endophytes isolated from a xerophyte plant promote the growth of Ammopiptanthus mongolicus under drought condition. Sci. Rep. 2018, 8, 7896. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; He, C.; He, X.; Su, F.; Hou, L.; Ren, Y.; Hou, Y. Dark septate endophytes improve the growth of host and non-host plants under drought stress through altered root development. Plant Soil 2019, 439, 259–272. [Google Scholar] [CrossRef]
- Deveautour, C.; Chieppa, J.; Nielsen, U.N.; Boer, M.M.; Mitchell, C.; Horn, S.; Power, S.A.; Guillen, A.; Bennett, A.E.; Powell, J.R. Biogeography of arbuscular mycorrhizal fungal spore traits along an aridity gradient, and responses to experimental rainfall manipulation. Fungal Ecol. 2020, 46, 100899. [Google Scholar] [CrossRef]
- Hoffman, M.T.; Arnold, A.E. Geographic locality and host identity shape fungal endophyte communities in cupressaceous trees. Mycol. Res. 2008, 112, 331–344. [Google Scholar] [CrossRef]
- Sui, X.; Zhang, R.; Xu, N.; Liu, Y.; Chai, C.; Wang, J.; Fu, X.; Zhong, H.; Ni, H. Fungal community structure of different degeneration Deyeuxia angustifolia wetlands in Sanjiang Plain. Environ. Sci. 2016, 37, 3598–3605. [Google Scholar]
- Abbassi, K.; Atay-Kadiri, Z.; Ghaout, S. Biological effects of alkaloids extracted from three plants of Moroccan arid areas on the desert locust. Physiol. Entomol. 2003, 28, 232–236. [Google Scholar] [CrossRef]
- Ju, T.; Chen, Y.; Chang, C.; An, L. The diversity of soil fungi and its relations with fertility factors in Taxus chinenesis (Pilg.) Rehd community of Xiaolongshan of Tianshui City. Res. J. Environ. Sci. 2008, 21, 128–132. [Google Scholar]
- Paterson, E.; Gebbing, T.; Abel, C.; Sim, A.; Telfer, G. Rhizodeposition shapes rhizosphere microbial community structure in organic soil. New Phytol. 2007, 173, 600–610. [Google Scholar] [CrossRef] [PubMed]
- de Vries, F.T.; Manning, P.; Tallowin, J.R.B.; Mortimer, S.R.; Pilgrim, E.S.; Harrison, K.A.; Hobbs, P.J.; Quirk, H.; Shipley, B.; Cornelissen, J.H.C.; et al. Abiotic drivers and plant traits explain landscape-scale patterns in soil microbial communities. Ecol. Lett. 2012, 15, 1230–1239. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, S.; Parsons, A.; Bassett, S.; Christensen, M.J.; Hume, D.; Johnson, L.; Johnson, R.; Simpson, W.; Stacke, C.; Voisey, C.; et al. High nitrogen supply and carbohydrate content reduce fungal endophyte and alkaloid concentration in Lolium perenne. New Phytol. 2007, 173, 787–797. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxa | R. soongorica | S. passerina | N. sphaerocarpa | E. przewalskii | S. regelii | |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Root | Stem | Leaf | Root | Stem | Leaf | Root | Stem | Root | Stem | Root | Stem | |||||||||||||
| I | RF% | I | RF% | I | RF% | I | RF% | I | RF% | I | RF% | I | RF% | I | RF% | I | RF% | I | RF% | I | RF% | I | RF% | |
| Alternaria alternata | 10 | 100 | 10 | 62.5 | 5 | 38.46 | 1 | 12.5 | 6 | 60 | 7 | 38.89 | 3 | 50.00 | ||||||||||
| Alternaria brassicae | 1 | 7.69 | ||||||||||||||||||||||
| Neocamarosporium salicornicola | 1 | 7.69 | ||||||||||||||||||||||
| Alternaria chlamydospora | 2 | 14.29 | 2 | 22.22 | 1 | 7.69 | 1 | 6.25 | 4 | 22.22 | 1 | 16.67 | ||||||||||||
| Aporspora terricola | 3 | 21.43 | 4 | 44.44 | 3 | 18.75 | 4 | 20 | 1 | 5.56 | ||||||||||||||
| Neocamarosporium salsolae | 2 | 25 | ||||||||||||||||||||||
| Dimorphosporicola tragani | 4 | 30.77 | 2 | 25 | ||||||||||||||||||||
| Neocamarosporium chichastianum | 1 | 7.69 | 2 | 25 | ||||||||||||||||||||
| Fusarium nematophilum | 1 | 6.25 | 2 | 11.11 | ||||||||||||||||||||
| Emericellopsis maritima | 2 | 12.5 | 1 | 5 | ||||||||||||||||||||
| Alternaria chlamydosporigena | 3 | 21.43 | 1 | 6.25 | ||||||||||||||||||||
| Alternaria terricola | 3 | 18.75 | ||||||||||||||||||||||
| Camarosporidiella eufemiaea | 3 | 30 | ||||||||||||||||||||||
| Aspergillus niger | 3 | 16.67 | ||||||||||||||||||||||
| Ulocladium sorghi | 1 | 5 | 1 | 16.67 | ||||||||||||||||||||
| Fusarium redolens | 2 | 22.22 | 2 | 12.5 | 5 | 25 | 2 | 11.11 | 2 | 11.11 | ||||||||||||||
| Aspergillus flavus | 2 | 12.5 | ||||||||||||||||||||||
| Aspergillus fumigatiaffinis | 4 | 25 | 1 | 5 | ||||||||||||||||||||
| Acremonium furcatum | 1 | 5.56 | ||||||||||||||||||||||
| Fusarium acuminatum | 2 | 10 | 1 | 5.56 | 1 | 16.67 | ||||||||||||||||||
| Penicilium chrysogenum | 1 | 11.11 | 1 | 6.25 | 1 | 10 | 1 | 5 | 6 | 33.33 | 4 | 22.22 | ||||||||||||
| Alternaria mouchaccae | 2 | 14.29 | 2 | 12.5 | ||||||||||||||||||||
| Fusarium oxysporum | 1 | 5 | ||||||||||||||||||||||
| Acremonium sclerotigenum | 1 | 12.5 | ||||||||||||||||||||||
| Didymella glomerata | 1 | 7.14 | ||||||||||||||||||||||
| Sordariomycetes sp. | 3 | 16.67 | ||||||||||||||||||||||
| Xylariales sp. | 1 | 7.14 | ||||||||||||||||||||||
| Preussia sp. | 2 | 14.29 | 4 | 20 | ||||||||||||||||||||
| Total | 14 | 24 | 40 | 49 | 62 | 70 | 86 | 96 | 116 | 134 | 152 | 158 | ||||||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zuo, Y.; Hu, Q.; Zhang, K.; He, X. Host and Tissue Affiliations of Culturable Endophytic Fungi Associated with Xerophytic Plants in the Desert Region of Northwest China. Agronomy 2022, 12, 727. https://doi.org/10.3390/agronomy12030727
Zuo Y, Hu Q, Zhang K, He X. Host and Tissue Affiliations of Culturable Endophytic Fungi Associated with Xerophytic Plants in the Desert Region of Northwest China. Agronomy. 2022; 12(3):727. https://doi.org/10.3390/agronomy12030727
Chicago/Turabian StyleZuo, Yiling, Qiannan Hu, Kaixun Zhang, and Xueli He. 2022. "Host and Tissue Affiliations of Culturable Endophytic Fungi Associated with Xerophytic Plants in the Desert Region of Northwest China" Agronomy 12, no. 3: 727. https://doi.org/10.3390/agronomy12030727