
Rhizobium Symbiosis Leads to Increased Drought Tolerance in Chinese Milk Vetch (Astragalus sinicus L.)


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Growth and Treatments
2.2. Determination of Nitrogenase Activity
2.3. Evaluation on Drought Tolerance
2.4. Determination of Free Amino Acids
2.5. Determination of Polyamines
2.6. Determinations of Enzyme Activity
2.7. Statistical Analysis
3. Results
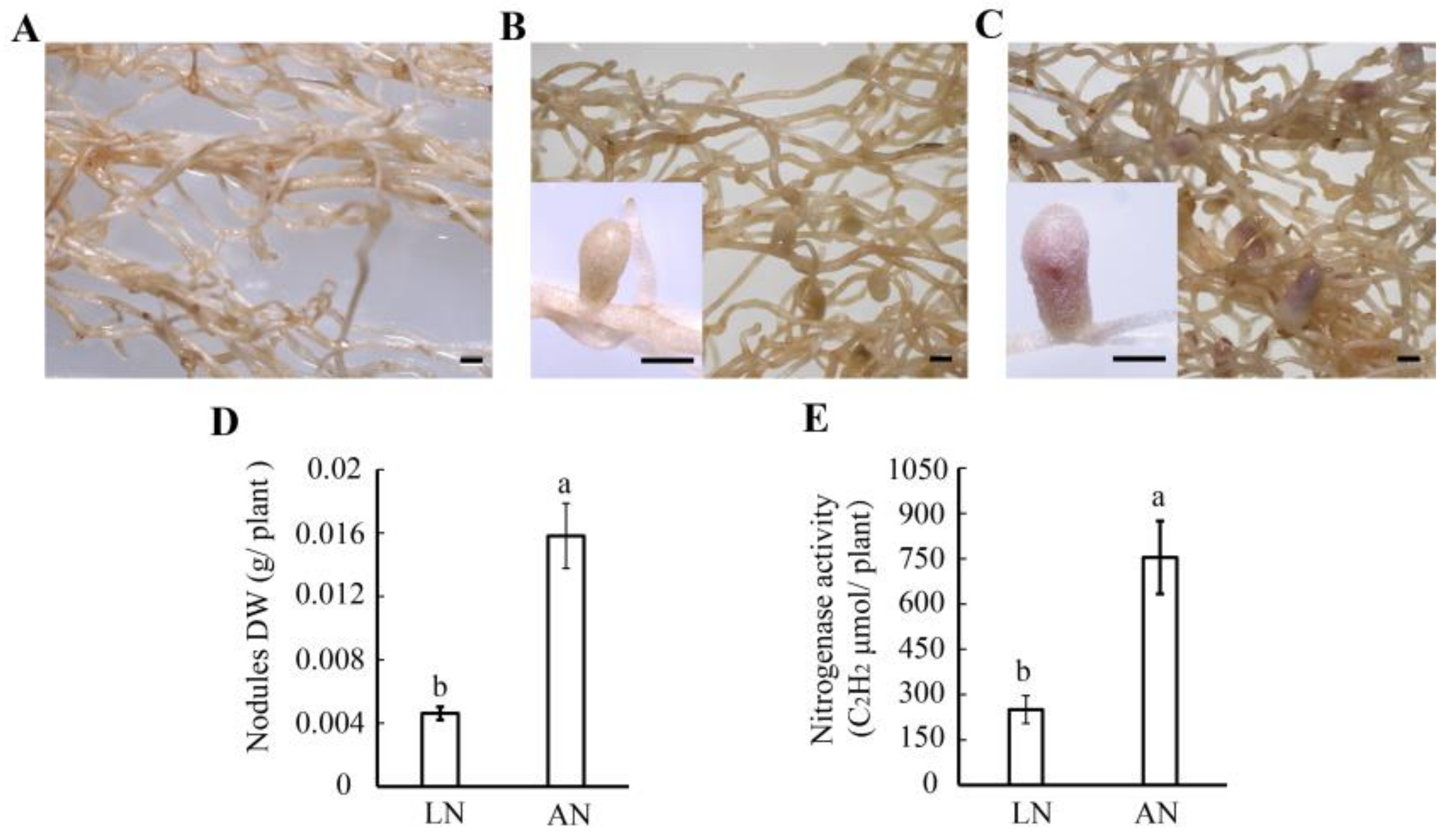
3.1. Nodulation in Milk Vetch under Different Levels of Nitrogen
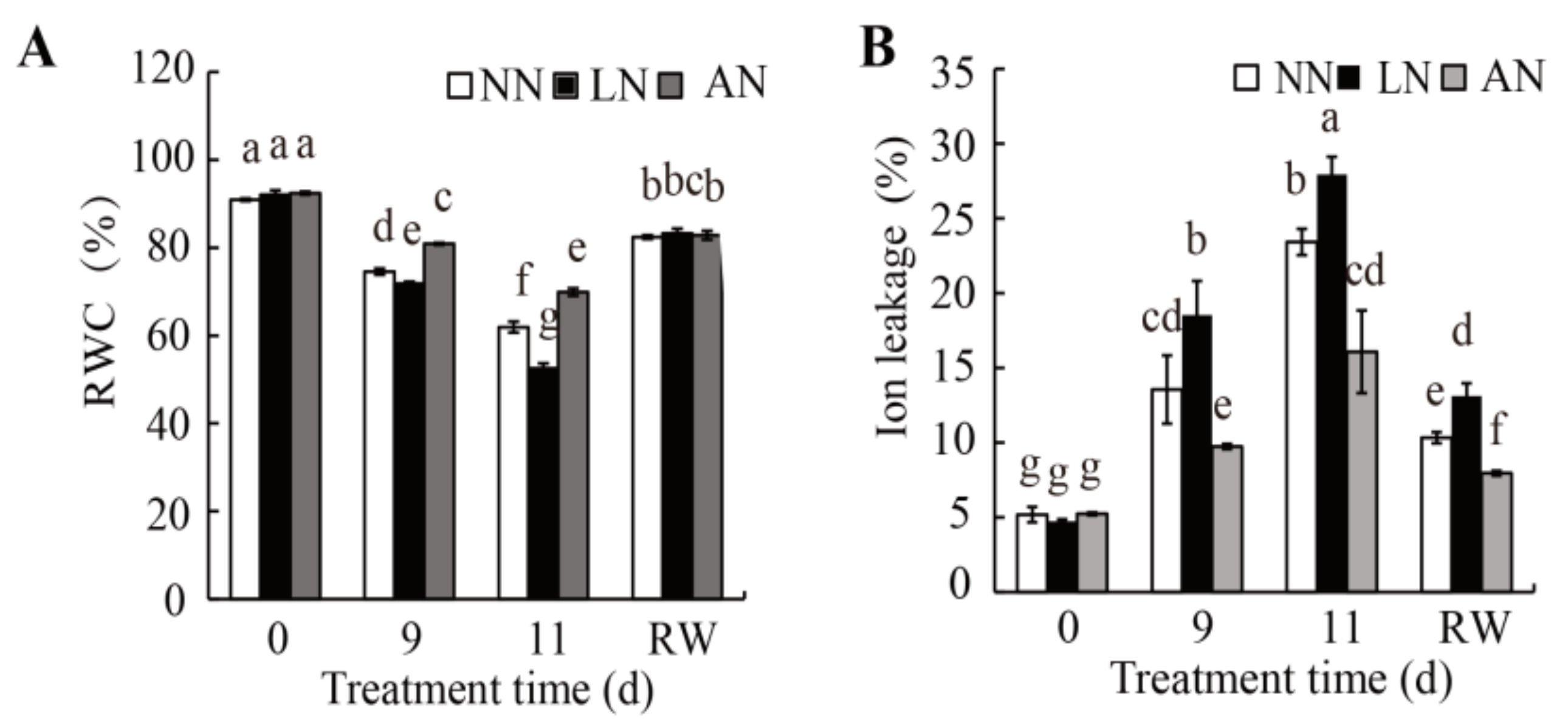
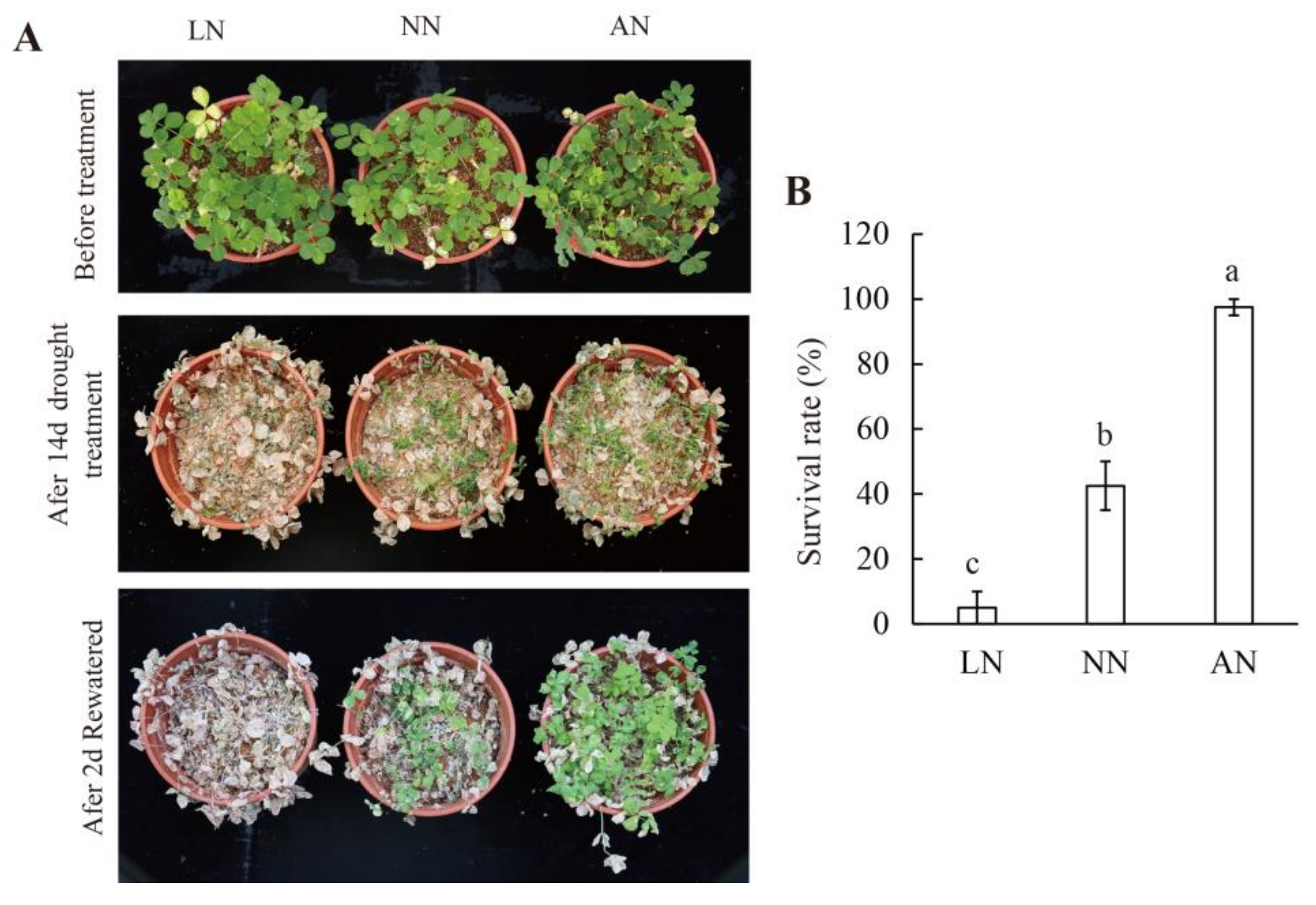
3.2. Nodulation Led to Increased Drought Tolerance in Milk Vetch
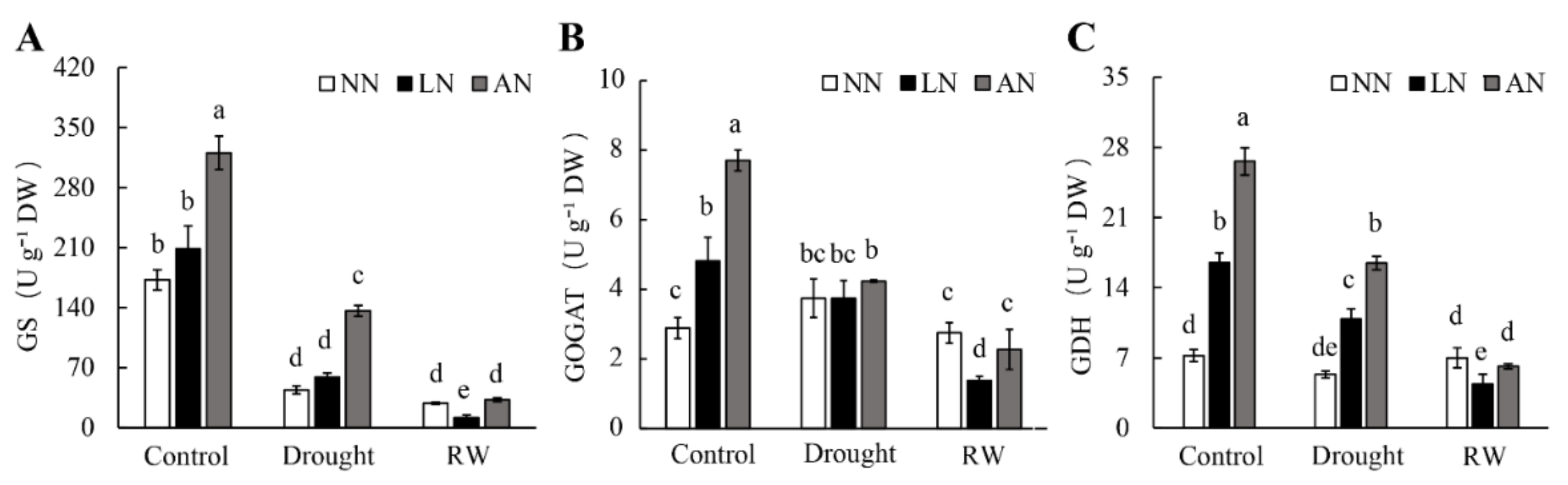
3.3. The Nodulated Milk Vetch Had Increased GOGAT, GS, and GDH Activities under Drought Conditions
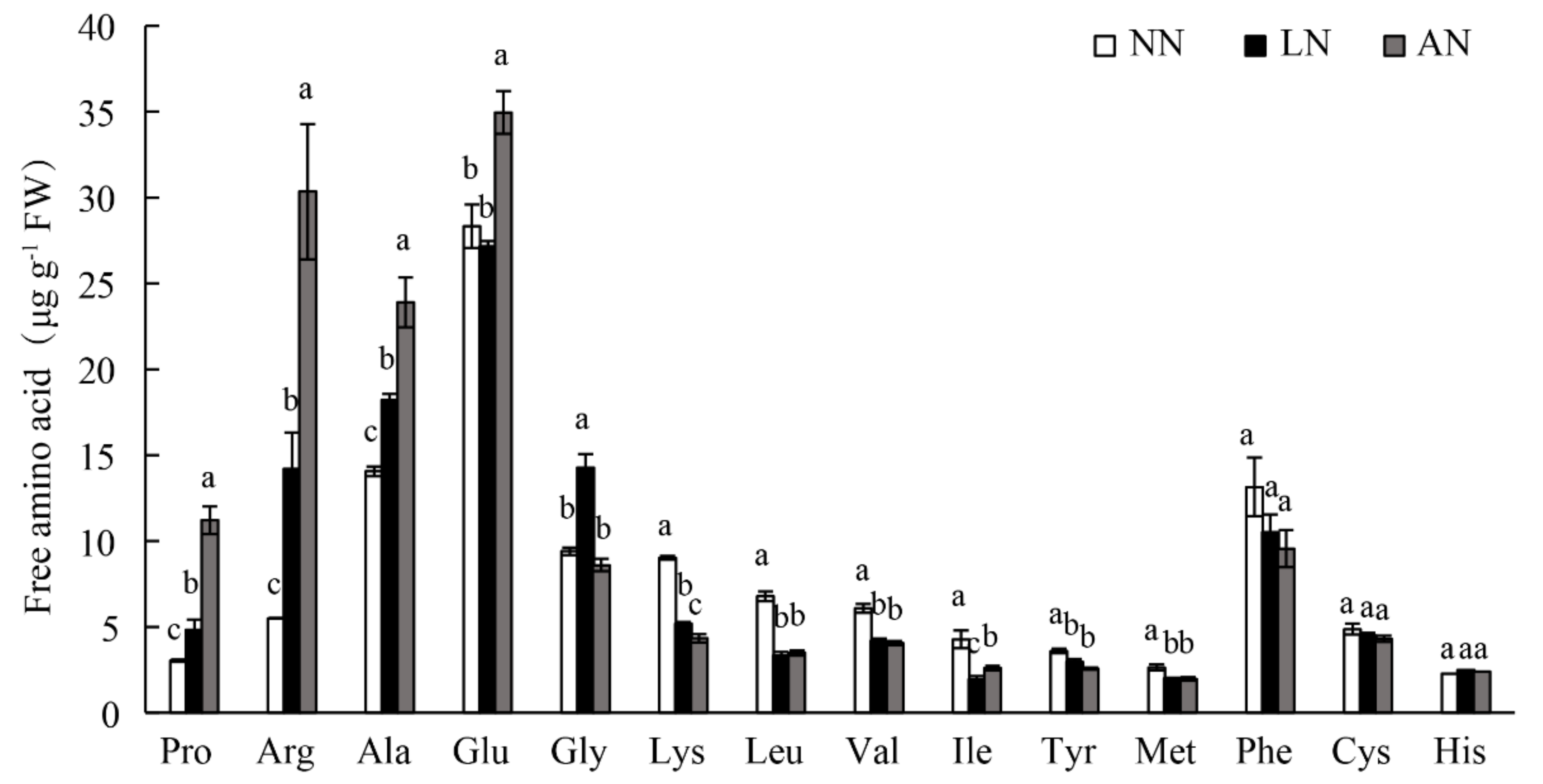
3.4. Nodulation Led to Altered Free Amino Acids in Milk Vetch
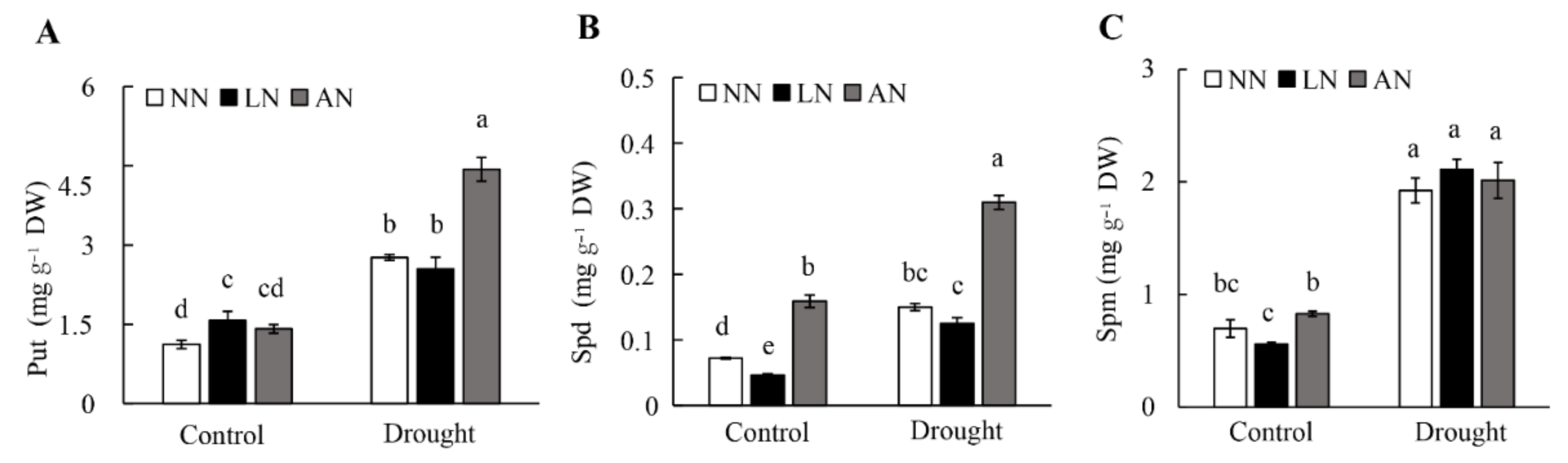
3.5. Polyamines in Milk Vetch as Affected by Nodulation and Drought Stress
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hoffman, B.M.; Lukoyanov, D.; Yang, Z.; Dean, D.R.; Seefeldt, L.C. Mechanism of nitrogen fixation by nitrogenase: The next stage. Chem. Rev. 2014, 114, 4041–4062. [Google Scholar] [CrossRef] [PubMed]
- Lea, P.J.; Miflin, B.J. Nitrogen assimilation and its relevance to crop improvement. In Annual Plant Reviews, Nitrogen Metabolism in Plants in the Post-Genomic Era; Foyer, C.H., Zhang, H., Eds.; Wiley-Blackwell: New York, NY, USA, 2011; Volume 42, pp. 1–40. [Google Scholar]
- Debouba, M.; Dguimi, H.M.; Ghorbel, M.; Gouia, H.; Suzuki, A. Expression pattern of genes encoding nitrate and ammonium assimilating enzymes in Arabidopsis thaliana exposed to short term NaCl stress. Plant Physiol. 2013, 170, 155–160. [Google Scholar] [CrossRef] [PubMed]
- Buchanan, B.B.; Gruissem, W.; Jnoes, R.L. Biochemistry and Molecular Biology of Plants, 2nd ed.; John Wiley & Sons: Hoboken, NJ, USA, 2015. [Google Scholar]
- Harrison, J.; Brugiere, N.; Phillipson, B.; Ferrario-Mery, S.; Becker, T.; Limami, A.; Hirel, B. Manipulating the pathway of ammonia assimilation through genetic engineering and breeding: Consequences to plant physiology and plant development. Plant Soil 2000, 221, 81–93. [Google Scholar] [CrossRef]
- Arrese-Igor, C.; González, E.M.; Marino, D.; Ladrera, R.; Larrainzar, E.; Gil-Quintana, R. Physiological response of legume nodules to drought. Plant Stress. 2011, 5, 24–31. Available online: https://hdl.handle.net/2454/23973 (accessed on 7 February 2022).
- Kunert, K.J.; Vorster, B.J.; Fenta, B.A.; Kibido, T.; Dionisio, G.; Foyer, C.H. Drought stress responses in soybean roots and nodules. Front. Plant Sci. 2016, 7, 1015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wadisirisuk, P.; Danso, S.K.A.; Hardarson, G.; Bowen, G.D. Influence of Bradyrhizobium japonicum location and movement on nodulation and nitrogen fixation in soybeans. Appl. Environ. Microbiol. 1989, 35, 1711–1716. [Google Scholar] [CrossRef] [Green Version]
- Mnasri, B.; Aouani, M.E.; Mhamdi, R. Nodulation and growth of common bean (Phaseolus vulgaris) under water deficiency. Soil Boil. Biochem. 2007, 39, 1744–1750. [Google Scholar] [CrossRef]
- Zahran, H.H. Rhizobium-legume symbiosis and nitrogen fixation under severe conditions and in an arid climate. Microbiol. Mol. Biol. Rev. 1999, 63, 968–989. [Google Scholar] [CrossRef] [Green Version]
- Elboutahiri, N.; Thami-Alami, I.; Udupa, S.M. Phenotypic and genetic diversity in Sinorhizobium meliloti and S. medicae from drought and salt affected regions of Morocco. BMC Microbiol. 2010, 10, 15. [Google Scholar] [CrossRef] [Green Version]
- Sulieman, S.; Tran, L.P. Legume Nitrogen Fixation in a Changing Environment; Springer: Berlin/Heidelberg, Germany, 2015. [Google Scholar]
- Ashraf, M.; Iram, A. Drought stress induced changes in some organic substances in nodules and other plant parts of two potential legumes differing in salt tolerance. Flora 2005, 200, 535–546. [Google Scholar] [CrossRef]
- Puppo, A.; Groten, K.; Bastian, F.; Carzaniga, R.; Foyer, C.H. Legume nodule senescence: Roles for redox and hormone signalling in the orchestration of the natural aging process. New Phytol. 2005, 165, 683–701. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo, M.V.; Burity, H.A.; Martinez, C.R.; Chanway, C.P. Alleviation of drought stress in the common bean (Phaseolus vulgaris L.) by co-inoculation with Paenibacillus polymyxa and Rhizobium tropici. Appl. Soil Ecol. 2008, 40, 182–188. [Google Scholar] [CrossRef]
- Yang, P.; Zhang, P.; Li, B.; Hu, T. Effect of nodules on dehydration response in alfalfa (Medicago sativa L.). Environ. Exp. Bot. 2013, 86, 29–34. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, Z.; Zhang, P.; Cao, Y.; Hu, T.; Yang, P. Rhizobium symbiosis contribution to short-term salt stress tolerance in alfalfa (Medicago sativa L.). Plant Soil 2016, 402, 247–261. [Google Scholar] [CrossRef]
- Song, T.; Xu, H.; Sun, N.; Jiang, L.; Tian, P.; Yong, Y.; Yang, W.; Cai, H.; Cui, G. Metabolomic analysis of alfalfa (Medicago sativa L.) root-symbiotic rhizobia responses under alkali stress. Front. Plant Sci. 2017, 8, 1208. [Google Scholar] [CrossRef] [PubMed]
- Staudinger, C.; Mehmeti-Tershani, V.; Gil-Quintana, E.; Gonzalez, E.M.; Hofhansl, F.; Bachmann, G.; Wienkoop, S. Evidence for a rhizobia-induced drought stress response strategy in Medicago truncatula. J. Proteom. 2016, 136, 202–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frechilla, S.; González, E.M.; Royuela, M.; Minchin, F.R.; Aparicio-Tejo, P.M.; Arrese-Igor, C. Source of nitrogen nutrition (nitrogen fixation or nitrate assimilation) is a major factor involved in pea response to moderate water stress. Plant Physiol. 2000, 157, 609–617. [Google Scholar] [CrossRef]
- Borsani, O.; Díaz, P.; Monza, J. Proline is involved in water stress responses of Lotus corniculatus nitrogen fixing and nitrate fed plants. J. Plant Physiol. 1999, 155, 269–273. [Google Scholar] [CrossRef]
- Lodeiro, A.R.; González, P.; Hernández, A.; Balagué, L.J.; Favelukes, G.F. Comparison of drought tolerance in nitrogen-fixing and inorganic nitrogen-grown common beans. Plant Sci. 2000, 154, 31–41. [Google Scholar] [CrossRef]
- Li, Y.; Zhou, L.; Li, Y.; Chen, D.; Tan, X.; Lei, L.; Zhou, J. A nodule-specific plant cysteine proteinase, AsNODF32, is involved in nodule senescence and nitrogen fixation activity of the green manure legume Astragalus sinicus. New Phytol. 2008, 180, 185–192. [Google Scholar] [CrossRef]
- Xie, Z.; Tu, S.; Shah, F.; Xu, C.; Chen, J.; Han, D.; Liu, G.; Li, H.; Muhammad, I.; Cao, W. Substitution of fertilizer-N by green manure improves the sustainability of yield in double-rice cropping system in south China. Field Crops Res. 2016, 188, 142–149. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Zhou, X.; Liao, Y.; Lu, Y.; Nie, J.; Cao, W. Co-incorporation of rice straw and green manure benefits rice yield and nutrient uptake. Crop Sci. 2019, 59, 749–759. [Google Scholar] [CrossRef]
- Gao, S.; Zhou, G.; Liao, Y.; Lu, Y.; Cao, W. Contributions of ammonia-oxidising bacteria and archaea to nitrification under long-term application of green manure in alkaline paddy soil. Geoderma 2020, 374, 114419. [Google Scholar] [CrossRef]
- Zhou, G.; Gao, S.; Lu, Y.; Liao, Y.; Nie, J.; Cao, W. Co-incorporation of green manure and rice straw improves rice production, soil chemical, biochemical and microbiological properties in a typical paddy field in southern China. Soil Tillage Res. 2020, 197, 104499. [Google Scholar] [CrossRef]
- Gao, S.; Cao, W.; Zhou, G.; Rees, R.M. Bacterial communities in paddy soils changed by milk vetch as green manure: A study conducted across six provinces in South China. Pedosphere 2021, 31, 521–530. [Google Scholar] [CrossRef]
- Hoagland, D.R.; Arnon, D.I. The water-culture method for growing plants without soil. Circular. Calif. Agric. Exp. Stn. 1950, 347, 32. [Google Scholar]
- David, K.A.; Apte, S.K.; Banerji, A.; Thomas, J. Acetylene reduction assay for nitrogenase activity: Gas chromatographic determination of ethylene per sample in less than one minute. Appl. Environ. Microbiol. 1980, 39, 1078–1080. [Google Scholar] [CrossRef] [Green Version]
- Lu, S.; Wang, Z.; Niu, Y.; Guo, Z.; Huang, B. Antioxidant responses of radiation-induced dwarf mutants of bermudagrass to drought stress. J. Am. Soc. Hortic. Sci. 2008, 133, 360–366. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Chen, J.; Guo, Z.; Lu, S. Differential responses of polyamines and antioxidants to drought in a centipedegrass mutant in comparison to its wild type plants. Front. Plant Sci. 2017, 8, 792. [Google Scholar] [CrossRef] [Green Version]
- Zhuo, C.; Wang, T.; Guo, Z.; Lu, S. Overexpression of MfPIP2-7 from Medicago falcata promotes cold tolerance and growth under NO3− deficiency in transgenic tobacco plants. BMC Plant Biol. 2016, 16, 138. [Google Scholar] [CrossRef] [Green Version]
- Guo, Z.; Tan, J.; Zhuo, C.; Wang, C.; Xiang, B.; Wang, Z. Abscisic acid, H2O2 and nitric oxide interactions mediated cold-induced S-adenosylmethionine synthetase in Medicago sativa subsp. falcata that confers cold tolerance through up-regulating polyamine oxidation. Plant Biotechnol. J. 2014, 12, 601–612. [Google Scholar] [CrossRef] [PubMed]
- Corbin, J.L.; Marsh, B.H.; Peters, G.A. An improved method for analysis of polyamines in plant tissue by precolumn derivatization with o-phthalaldehyde and separation by high performance liquid chromatography. Plant Physiol. 1989, 90, 434–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagy, Z.; Németh, E.; Guóth, A.; Bona, L.; Wodala, B.; Pécsváradi, A. Metabolic indicators of drought stress tolerance in wheat: Glutamine synthetase isoenzymes and Rubisco. Plant Physiol. Biochem. 2013, 67, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.P.; Srivastava, H.S. Regulation of glutamate dehydrogenase activity by amino acids in maize seedlings. Physiol. Plant. 1983, 57, 549–554. [Google Scholar] [CrossRef]
- Ohyama, T.; Kumazawa, K. Nitrogen assimilation in soybean nodules. Soil Sci. Plant Nutr. 1980, 26, 109–115. [Google Scholar] [CrossRef] [Green Version]
- Dwivedi, S.L.; Sahrawat, K.L.; Upadhyaya, H.D.; Mengoni, A.; Galardini, M.; Bazzicalupo, M.; Biondi, E.G.; Hungria, M.; Kaschuk, G.; Blair, M.W.; et al. Advances in host plant and rhizobium genomics to enhance symbiotic nitrogen fixation in grain legumes. Adv. Agron. 2015, 129, 1–116. [Google Scholar] [CrossRef]
- Hirel, B.; Lea, P.J. Ammonia assimilation. In Plant Nitrogen; Lea, P.J., Morot-Gaudry, J.-F., Eds.; Springer: Berlin/Heidelberg, Germany, 2001; pp. 79–99. [Google Scholar]
- Guan, M.; Bang, T.D.; Pedersen, C.; Schijoerring, J.K. Cytosolic glutamine synthetase Gln1;2 is the main isozyme contributing to GS1 activity in Arabidopsis shoots and can be up-regulated to relieve ammonium toxicity. Plant Physiol. 2016, 171, 1921. [Google Scholar] [CrossRef] [Green Version]
- Ashraf, M.; Ad, M.F. Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environ. Exp. Bot. 2007, 59, 206–216. [Google Scholar] [CrossRef]
- Kaur, G.; Asthir, B. Proline: A key player in plant abiotic stress tolerance. Biol. Plant. 2015, 59, 609–619. [Google Scholar] [CrossRef]
- Rai, V.K. Role of amino acids in plant responses to stresses. Biol. Plant. 2002, 45, 481–487. [Google Scholar] [CrossRef]
- Rai, V.K.; Bapat, C.M. Water stress effects on amino acid metabolism during early vegetative growth in Phaseolus mungo L. Geobios 1977, 4, 231–234. [Google Scholar]
- Mori, T.; Sakagami, Y.; Doi, K. Changes in free amino acid contents in Cryptomeria japonica transplants under various soil moisture conditions. J. For. Res. 1971, 52, 350–354. [Google Scholar]
- Yang, C.W.; Lin, C.C.; Kao, C.H. Proline, ornithine, arginine and glutamic acid contents in detached rice leaves. Biol. Plant. 2000, 43, 305–307. [Google Scholar] [CrossRef]
- Haghighi, M.; Saadat, S.; Abbey, L. Effect of exogenous amino acids application on growth and nutritional value of cabbage under drought stress. Sci. Hortic. 2020, 272, 109561. [Google Scholar] [CrossRef]
- Bowne, J.B.; Erwin, T.A.; Juttner, J.; Schnurbusch, T.; Langridge, P.; Bacic, A.A.; Roessner, U. Drought responses of leaf tissues from wheat cultivars of differing drought tolerance at the metabolite level. Mol. Plant. 2012, 5, 418–429. [Google Scholar] [CrossRef] [Green Version]
- You, J.; Zhang, Y.; Liu, A.; Li, E.; Zhang, X. Transcriptomic and metabolomic profiling of drought-tolerant and susceptible sesame genotypes in response to drought stress. BMC Plant Biol. 2019, 19, 267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanfrey, C.; Sommer, S.; Mayer, M.J.; Burtin, D.; Michael, A.J. Arabidopsis polyamine biosynthesis: Absence of ornithine decarboxylase and the mechanism of arginine decarboxylase activity. Plant J. 2010, 27, 551–560. [Google Scholar] [CrossRef] [Green Version]
- Miller-Fleming, L.; Olin-Sandoval, V.; Campbell, K.; Ralser, M. Remaining mysteries of molecular biology: The role of polyamines in the cell. J. Mol. Cell Biol. 2015, 427, 3389–3406. [Google Scholar] [CrossRef] [PubMed]
- Minocha, R.; Majumdar, R.; Minocha, S.C. Polyamines and abiotic stress in plants: A complex relationship. Front. Plant Sci. 2014, 5, 175. [Google Scholar] [CrossRef] [Green Version]
- Todorova, D.; Katerova, Z.; Alexieva, V.; Sergiev, I. Polyamines—Possibilities for application to increase plant tolerance and adaptation capacity to stress. Genet. Plant Physiol. 2015, 5, 123–144. [Google Scholar]
- Alcázar, R.; Bueno, M.; Tiburcio, A.F. Polyamines: Small amines with large effects on plant abiotic stress tolerance. Cells 2020, 9, 2373. [Google Scholar] [CrossRef] [PubMed]
- Galston, A.W.; Kaur-Sawhney, R.; Altabella, T.; Tiburcio, A.F. Plant polyamines in reproductive activity and response to abiotic stress. Bot. Acta 2015, 110, 197–207. [Google Scholar] [CrossRef]
- Capell, T.; Bassie, L.; Christou, P. Modulation of the polyamine biosynthetic pathway in transgenic rice confers tolerance to drought stress. Proc. Natl. Acad. Sci. USA 2014, 101, 9909–9914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, D.; Shao, Q.; Yin, L.; Younis, A.; Zheng, B. Polyamine function in plants: Metabolism, regulation on development, and roles in abiotic stress responses. Front. Plant Sci. 2019, 9, 1945. [Google Scholar] [CrossRef]
- Satish, L.; Rency, A.S.; Ramesh, M. Spermidine sprays alleviate the water deficit-induced oxidative stress in finger millet (Eleusine coracana L. Gaerth.) plants. 3 Biotech 2018, 8, 63. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Wang, L.; Yang, R.; Han, Y.; Hao, J.; Liu, C.; Fan, S. Effects of exogenous putrescine on the ultrastructure of and calcium ion flow rate in lettuce leaf epidermal cells under drought stress. Hortic. Environ. Biotechnol. 2019, 60, 479–490. [Google Scholar] [CrossRef]
- Liu, J.H.; Wang, W.; Wu, H. Polyamines function in stress tolerance: From synthesis to regulation. Front. Plant Sci. 2015, 6, 827. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Guo, Z.; Shi, H. Rhizobium Symbiosis Leads to Increased Drought Tolerance in Chinese Milk Vetch (Astragalus sinicus L.). Agronomy 2022, 12, 725. https://doi.org/10.3390/agronomy12030725
Liu Y, Guo Z, Shi H. Rhizobium Symbiosis Leads to Increased Drought Tolerance in Chinese Milk Vetch (Astragalus sinicus L.). Agronomy. 2022; 12(3):725. https://doi.org/10.3390/agronomy12030725
Chicago/Turabian StyleLiu, Yajie, Zhenfei Guo, and Haifan Shi. 2022. "Rhizobium Symbiosis Leads to Increased Drought Tolerance in Chinese Milk Vetch (Astragalus sinicus L.)" Agronomy 12, no. 3: 725. https://doi.org/10.3390/agronomy12030725