
Sustainable Growing Media Blends with Woody Green Composts: Optimizing the N Release with Organic Fertilizers and Interaction with Microbial Biomass

 ,
,
Abstract
:1. Introduction
1.1. New Materials in Growing Media Blends
1.2. The Microbiome of Growing Media
1.3. Interaction between Growing Media and Organic Fertilizers
1.4. Aim
2. Materials and Methods
2.1. Characterization of Materials and Blends
2.2. Microbial Biomass and Activity and Net N Release of Composts versus Wood Fiber and Bark
2.3. Net N Release from Composts Blended with Blood Meal
2.4. Effect of Blend and Organic Fertilizer on N Release
3. Results
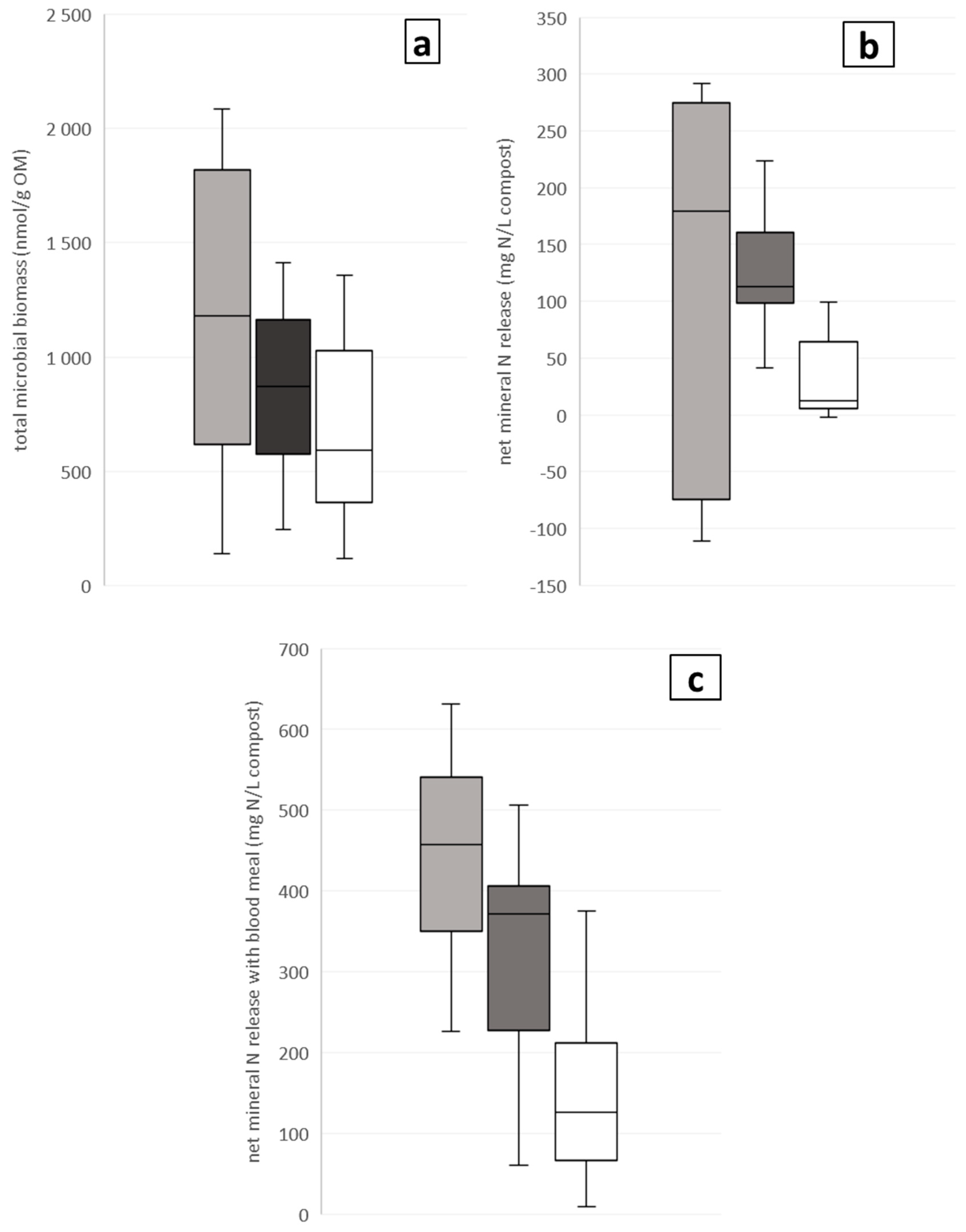
3.1. Step 1: Microbial Biomass and Activity and Net N Release of Composts versus Wood Fiber and Bark
3.2. Step 2: Net N Release from Composts Blended with Blood Meal
3.3. Step 3: Effect of Blend and Organic Fertilizer on N Release
3.3.1. Characteristics of the Materials and the Blends
3.3.2. Total Mineral N Concentrations and Microbial Biomass
3.3.3. Plant Trial with Cucumber: Seed Emergence
4. Discussion
- -
- What is the N release of composts selected for a better fit for use in growing media, both for the composts with or without added organic fertilizers?
- -
- Can the blend with woody compost be optimized with organic fertilizers to increase the mineral N availability? What is the interaction between N release and microbial biomass?
4.1. Compost: Role as Bulk Material vs. Role as Organic Fertilizer
4.2. Organic Fertilizers: N Release and Microbial Activity
4.3. N Release versus Blend Composition: How to Provide Sufficient N?
4.4. Total Microbial Biomass: Value as Indicator and Link with N Release
4.5. Implications for Use of the Growing Media Blends with Woody Composts
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Agarwal, P.; Saha, S.; Hariprasad, P. Agro-industrial-residues as potting media: Physicochemical and biological characters and their influence on plant growth. Biomass Convers. Biorefinery 2021, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Barrett, G.E.; Alexander, P.D.; Robinson, J.S.; Bragg, N.C. Achieving environmentally sustainable growing media for soilless plant cultivation systems—A review. Sci. Hortic. 2016, 212, 220–234. [Google Scholar] [CrossRef] [Green Version]
- Atzori, G.; Pane, C.; Zaccardelli, M.; Cacini, S.; Massa, D. The Role of Peat-Free Organic Substrates in the Sustainable Management of Soilless Cultivations. Agronomy 2021, 11, 1236. [Google Scholar] [CrossRef]
- Vandecasteele, B.; Pot, S.; Maenhout, K.; Delcour, I.; Vancampenhout, K.; Debode, J. Acidification of composts versus woody management residues: Optimizing biological and chemical characteristics for a better fit in growing media. J. Environ. Manag. 2021, 277, 111444. [Google Scholar] [CrossRef] [PubMed]
- Neher, D.A.; Fang, L.; Weicht, T.R. Ecoenzymes as indicators of compost to suppress Rhizoctonia solani. Compost. Sci. Util. 2017, 25, 251–261. [Google Scholar] [CrossRef] [Green Version]
- Herren, G.; Binnemans, I.; Joos, L.; Viaene, N.; Ehlers, R.-U.; Vandecasteele, B.; Bert, W.; Steel, H. Compost as a carrier medium for entomopathogenic nematodes: The influence of compost maturity on their virulence and survival. Biol. Control. 2018, 125, 29–38. [Google Scholar] [CrossRef]
- Cambardella, C.A.; Richard, T.; Russell, A. Compost mineralization in soil as a function of composting process conditions. Eur. J. Soil Biol. 2003, 39, 117–127. [Google Scholar] [CrossRef] [Green Version]
- Blok, C.; van der Salm, C.; Hofland-Zijlstra, J.; Streminska, M.; Eveleens, B.; Regelink, I.; Fryda, L.; Visser, R. Biochar for horticultural rooting media improvement: Evaluation of biochar from gasification and slow pyrolysis. Agronomy 2017, 7, 6. [Google Scholar] [CrossRef] [Green Version]
- Escuer, O.; Karp, K.; Escuer-Gatius, J.; Raave, H.; Teppand, T.; Shanskiy, M. Hardwood biochar as an alternative to reduce peat use for seed germination and growth of Tagetes patula. Acta Agric. Scand. Sect. B Soil Plant Sci. 2021, 71, 1–14. [Google Scholar] [CrossRef]
- Dittrich, C.; Pecenka, R.; Løes, A.-K.; Cáceres, R.; Conroy, J.; Rayns, F.; Schmutz, U.; Kir, A.; Kruggel-Emden, H. Extrusion of Different Plants into Fibre for Peat Replacement in Growing Media: Adjustment of Parameters to Achieve Satisfactory Physical Fibre-Properties. Agronomy 2021, 11, 1185. [Google Scholar] [CrossRef]
- Vandecasteele, B.; Muylle, H.; De Windt, I.; Van Acker, J.; Ameloot, N.; Moreaux, K.; Coucke, P.; Debode, J. Plant fibers for renewable growing media: Potential of defibration, acidification or inoculation with biocontrol fungi to reduce the N drawdown and plant pathogens. J. Clean. Prod. 2018, 203, 1143–1154. [Google Scholar] [CrossRef]
- Hartung, C.; Meinken, E. Fen plant biomass as growing media constituent—Reduction of nitrogen immobilization by composting. Acta Hortic. 2021, 1317, 93–98. [Google Scholar] [CrossRef]
- Montagne, V.; Charpentier, S.; Cannavo, P.; Capiaux, H.; Grosbellet, G.; Lebeau, T. Structure and activity of spontaneous fungal communities in organic substrates used for soilless crops. Sci. Hortic. 2015, 192, 148–157. [Google Scholar] [CrossRef]
- Montagne, V.; Capiaux, H.; Barret, M.; Cannavo, P.; Charpentier, S.; Grosbellet, G.; Lebeau, T. Bacterial and fungal communities vary with the type of organic substrate: Implications for biocontrol of soilless crops. Environ. Chem. Lett. 2017, 15, 537–545. [Google Scholar] [CrossRef]
- Pot, S.; De Tender, C.; Ommeslag, S.; Delcour, I.; Ceusters, J.; Gorrens, E.; Debode, J.; Vandecasteele, B.; Vancampenhout, K. Understanding the Shift in the Microbiome of Composts That Are Optimized for a Better Fit-for-Purpose in Growing Media. Front. Microbiol. 2021, 12, 757. [Google Scholar] [CrossRef] [PubMed]
- Taparia, T.; Hendrix, E.; Nijhuis, E.; de Boer, W.; van der Wolf, J. Circular alternatives to peat in growing media: A microbiome perspective. J. Clean. Prod. 2021, 327, 129375. [Google Scholar] [CrossRef]
- Grunert, O.; Reheul, D.; Van Labeke, M.; Perneel, M.; Hernandez-Sanabria, E.; Vlaeminck, S.E.; Boon, N. Growing media constituents determine the microbial nitrogen conversions in organic growing media for horticulture. Microb. Biotechnol. 2016, 9, 389–399. [Google Scholar] [CrossRef] [Green Version]
- Cacini, S.; Di Lonardo, S.; Orsenigo, S.; Massa, D. Managing pH of Organic Matrices and New Commercial Substrates for Ornamental Plant Production: A Methodological Approach. Agronomy 2021, 11, 851. [Google Scholar] [CrossRef]
- Verhagen, J.B.G.M. Using organic fertilisers in growing media for young plant production. Acta Hortic. 2021, 1317, 141–148. [Google Scholar] [CrossRef]
- Gonnella, M.; Renna, M. The Evolution of Soilless Systems towards Ecological Sustainability in the Perspective of a Circular Economy. Is It Really the Opposite of Organic Agriculture? Agronomy 2021, 11, 950. [Google Scholar] [CrossRef]
- Paillat, L.; Cannavo, P.; Barraud, F.; Huché-Thélier, L.; Guénon, R. Growing Medium Type Affects Organic Fertilizer Mineralization and CNPS Microbial Enzyme Activities. Agronomy 2020, 10, 1955. [Google Scholar] [CrossRef]
- Lashermes, G.; Nicolardot, B.; Parnaudeau, V.; Thuries, L.; Chaussod, R.; Guillotin, M.L.; Lineres, M.; Mary, B.; Metzger, L.; Morvan, T.; et al. Typology of exogenous organic matters based on chemical and biochemical composition to predict potential nitrogen mineralization. Bioresour. Technol. 2010, 101, 157–164. [Google Scholar] [CrossRef]
- De Tender, C.; Vandecasteele, B.; Verstraeten, B.; Ommeslag, S.; De Meyer, T.; De Visscher, J.; Dawyndt, P.; Clement, L.; Kyndt, T.; Debode, J. Chitin in strawberry cultivation: Foliar growth and defense response promotion, but reduced fruit yield and disease resistance by nutrient imbalances. Mol. Plant-Microbe Interact. 2021, 34, 227–239. [Google Scholar] [CrossRef] [PubMed]
- Vandecasteele, B.; Amery, F.; Ommeslag, S.; Vanhoutte, K.; Visser, R.; Robbens, J.; De Tender, C.; Debode, J. Chemically versus thermally processed brown shrimp shells or Chinese mitten crab as a source of chitin, nutrients or salts and as microbial stimulant in soilless strawberry cultivation. Sci. Total Environ. 2021, 771, 145263. [Google Scholar] [CrossRef] [PubMed]
- De Tender, C.; Mesuere, B.; Van der Jeugt, F.; Haegeman, A.; Ruttink, T.; Vandecasteele, B.; Dawyndt, P.; Debode, J.; Kuramae, E.E. Peat substrate amended with chitin modulates the N-cycle, siderophore and chitinase responses in the lettuce rhizobiome. Sci. Rep. 2019, 9, 9890. [Google Scholar] [CrossRef]
- El Knidri, H.; Belaabed, R.; Abdellah, A.; Laajeb, A.; Lahsini, A. Extraction, chemical modification and characterization of chitin and chitosan: A review. Int. J. Biol. Macromol. 2018, 120, 1181–1189. [Google Scholar] [CrossRef]
- Hartz, T.K.; Mitchell, J.P.; Giannini, C. Nitrogen and Carbon Mineralization Dynamics of Manures and Composts. HortScience 2000, 35, 209–212. [Google Scholar] [CrossRef]
- Hartz, T.K.; Giannini, C. Duration of Composting of Yard Wastes Affects Both Physical and Chemical Characteristics of Compost and Plant Growth. HortScience 1998, 33, 1192–1196. [Google Scholar] [CrossRef] [Green Version]
- Cáceres, R.; Malinska, K.; Marfà, O. Nitrification within composting: A review. Waste Manag. 2017, 72, 119–137. [Google Scholar] [CrossRef]
- Grigatti, M.; Cavani, L.; Ciavatta, C. The evaluation of stability during the composting of different starting materials: Comparison of chemical and biological parameters. Chemosphere 2011, 83, 41–48. [Google Scholar] [CrossRef]
- Blok, C.; Verkerke, W.; Boedijn, A.; Streminska, M.; Eveleens, B. Recirculation, circular fertilizers and resilience: The potential of growing media systems for circular production. Acta Hortic. 2021, 1317, 189–206. [Google Scholar] [CrossRef]
- Chamuris, G.P.; Koziol-Kotch, S.; Brouse, T.M. Screening Fungi Isolated from Woody Compost for Lignin-Degrading Potential. Compost. Sci. Util. 2000, 8, 6–11. [Google Scholar] [CrossRef]
- Cytryn, E.; Levkovitch, I.; Negreanu, Y.; Dowd, S.; Frenk, S.; Silber, A. Impact of short-term acidification on nitrification and nitrifying bacterial community dynamics in soilless cultivation media. Appl. Environ. Microbiol. 2012, 78, 6576–6582. [Google Scholar] [CrossRef] [Green Version]
- Dion, P.; Jeanne, T.; Thériault, M.; Hogue, R.; Pepin, S.; Dorais, M. Nitrogen release from five organic fertilizers commonly used in greenhouse organic horticulture with contrasting effects on bacterial communities. Can. J. Soil Sci. 2020, 100, 120–135. [Google Scholar] [CrossRef]
- Grunert, O.; Robles-Aguilar, A.A.; Hernandez-Sanabria, E.; Schrey, S.; Reheul, D.; Van Labeke, M.C.; Vlaeminck, S.; Vandekerckhove, T.; Mysara, M.; Monsieurs, P.; et al. Tomato plants rather than fertilizers drive microbial community structure in horticultural growing media. Sci. Rep. 2019, 9, 9561. [Google Scholar] [CrossRef] [PubMed]
- Ciavatta, C.; Govi, M.; Sitti, L.; Gessa, C. Influence of blood meal organic fertilizer on soil organic matter: A laboratory study. J. Plant Nutr. 1997, 20, 1573–1591. [Google Scholar] [CrossRef]
- Ebrahimi, N.; Viaene, N.; Vandecasteele, B.; D’Hose, T.; Debode, J.; Cremelie, P.; De Tender, C.; Moens, M. Traditional and new soil amendments reduce survival and reproduction of potato cyst nematodes, except for biochar. Appl. Soil Ecol. 2016, 107, 191–204. [Google Scholar] [CrossRef]
- Debode, J.; Ebrahimi, N.; D’Hose, T.; Cremelie, P.; Viaene, N.; Vandecasteele, B. Has compost with biochar added during the process added value over biochar or compost to increase disease suppression? Appl. Soil Ecol. 2020, 153, 103571. [Google Scholar] [CrossRef]
- Debode, J.; De Tender, C.; Soltaninejad, S.; Van Malderghem, C.; Haegeman, A.; Van der Linden, I.; Cottyn, B.; Heyndrickx, M.; Maes, M. Chitin Mixed in Potting Soil Alters Lettuce Growth, the Survival of Zoonotic Bacteria on the Leaves and Associated Rhizosphere Microbiology. Front. Microbiol. 2016, 7, 565. [Google Scholar] [CrossRef] [PubMed]
- Blok, C.; Eveleens, B.; van Winkel, A. Oxygen use in compost storage as influenced by moisture, temperature and degradability. Acta Hortic. 2019, 1266, 291–300. [Google Scholar] [CrossRef]
- Diaz-Perez, J.C.; Keith Jenkins, W.; Pitchay, D.; Gunawan, G. Detrimental Effects of Blood Meal and Feather Meal on Tomato (Solanum lycopersicon L.) Seed Germination. HortScience 2017, 52, 138–141. [Google Scholar] [CrossRef] [Green Version]
- Cayuela, M.L.; Mondini, C.; Insam, H.; Sinicco, T.; Franke-Whittle, I. Plant and animal wastes composting: Effects of the N source on process performance. Bioresour. Technol. 2009, 100, 3097–3106. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Category | n | IC | pH-H2O | EC | Nmin | Nimmob | P-AmAc | K-AmAc | OM | Dry Bulk Density | OUR | C/N | Total Biomass |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| %/DM | - | µS/cm | mg N/L Compost | % | mg/L Compost | %/DM | g DM/L Compost | mmol O2/kg OM/hr | - | nmol/g OM | |||
| VFG compost | 3 | 0.4 (c) | 8.4 (bc) | 2240 (b) | 560 (b) | −19 | 621 (b) | 3287 (b) | 39 (a) | 287 (c) | 5.0 (ab) | 9 (a) | 1461 (c) |
| Green compost | 6 | 0.31 (bc) | 8.4 (c) | 1185 (b) | 128 (ab) | −4 | 367 (b) | 3304 (b) | 38 (a) | 285 (c) | 4.2 (ab) | 12 (a) | 992 (c) |
| Woody compost | 5 | 0.24 (abc) | 8.4 (c) | 708 (b) | 37 (a) | 9 | 234 (b) | 1847 (b) | 42 (a) | 261 (c) | 4.9 (b) | 19 (a) | 898 (bc) |
| Bark compost | 5 | 0.08 (a) | 6.4 (a) | 150 (a) | 13 (a) | 20 | 37 (a) | 457 (a) | 86 (b) | 143 (b) | 4.1 (ab) | 75 (b) | 341 (b) |
| Wood fiber | 4 | 0.13 (ab) | 7.0 (ab) | 39 (a) | 10 (a) | −4 | 9 (a) | 35 (a) | 99 (b) | 50 (a) | 1.1 (a) | 436 (c) | 77 (a) |
| Category | Score | Net N Release | Nmin | IC | pH-H2O | EC | Nimmob | P-AmAc | K-AmAc | OM | Dry Bulk Density | OUR | C/N | Total Biomass |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| mg N/L Compost | %/DM | - | µS/cm | % | mg/L Compost | %/DM | g DM/L | mmol O2/kg OM/hr | - | nmol/g OM | ||||
| Composts | ||||||||||||||
| VFG compost 1 | 5 | 229 | 1069 | 0.50 | 8.5 | 3030 | 36 | 708 | 3031 | 38 | 341 | 7.2 | 9.7 | 1527 |
| VFG compost 2 | 6 | 124 | 364 | 0.35 | 8.5 | 2200 | −58 | 600 | 4017 | 33 | 297 | 2.4 | 8.6 | 1013 |
| VFG compost 3 | 9 | 59 | 247 | 0.34 | 8.3 | 1490 | −34 | 554 | 2812 | 45 | 223 | 5.4 | 9.8 | 1843 |
| Green compost 1 | 5 | 209 | 233 | 0.50 | 8.9 | 1558 | 44 | 538 | 5048 | 41 | 334 | 4.8 | 11 | 411 |
| Green compost 2 | 7 | 12 | 204 | 0.44 | 7.9 | 1273 | −3 | 355 | 3618 | 31 | 381 | 3.1 | 11 | 616 |
| Green compost 3 | 8 | <5 | <5 | 0.16 | 7.5 | 361 | 21 | 49 | 901 | 37 | 266 | 2.8 | 17 | 1525 |
| Green compost 4 | 8 | 380 | 231 | 0.31 | 8.3 | 1721 | −28 | 339 | 4141 | 39 | 237 | 3.0 | 10 | 986 |
| Green compost 5 | 9 | 37 | 85 | 0.29 | 8.1 | 894 | −38 | 586 | 2658 | 50 | 190 | 2.2 | 12 | 1194 |
| Green compost 6 | 10 | −19 | <5 | <0.1 | 8.7 | 810 | −5 | 127 | 1861 | 40 | 236 | 5.9 | 15 | 1216 |
| Woody compost 1 | 9 | <5 | <5 | <0.1 | 7.6 | 343 | 13 | 69 | 1138 | 46 | 261 | 6.8 | 29 | 214 |
| Woody compost 2 | 9 | −20 | 31 | 0.24 | 8.4 | 697 | 12 | 268 | 2147 | 41 | 291 | 1.9 | 18 | 890 |
| Woody compost 3 | 9 | 25 | 40 | 0.37 | 8.8 | 1051 | 8 | 318 | 1697 | 47 | 208 | 6.4 | 16 | 1123 |
| Woody compost 4 | 10 | 17 | 20 | 0.35 | 8.6 | 939 | 13 | 259 | 2817 | 42 | 213 | 7.1 | 16 | 1248 |
| Woody compost 5 | 10 | 50 | 83 | 0.16 | 8.6 | 511 | 0 | 259 | 1439 | 33 | 332 | 2.3 | 17 | 1015 |
| Compost from SGM | 13 | 40 | 401 | < 0.1 | 6.9 | 1253 | 10 | 34 | 84 | 82 | 101 | 1.2 | 24 | 295 |
| Other materials used in the growing media blends | ||||||||||||||
| Bark compost | 14 | 3 | 23 | <0.1 | 6.3 | 94 | −27 | 19 | 373 | 87 | 145 | 2.5 | 56 | 238 |
| Wood fiber | 9 | NA | <5 | <0.1 | 7.5 | 33 | −7 | <4 | <23 | 99 | 36 | 0.8 | 624 | 71 |
| M1 | M2 | M3 | M4 | M5 | M0 | |
|---|---|---|---|---|---|---|
| Composition | ||||||
| Woody compost 1 (vol%) | 40 | 40 | ||||
| Woody compost 2 (vol%) | 40 | 40 | ||||
| Black peat (vol%) | 15 | 15 | 15 | 50 | ||
| White peat (vol%) | 15 | 15 | 15 | 50 | ||
| Bark compost (vol%) | 30 | 30 | 30 | 30 | 30 | |
| Wood Fiber (vol%) | 30 | 30 | 20 | |||
| Coir fiber (vol%) | 20 | |||||
| Lime (g/L) | 0 | 0 | 0 | 0 | 0 | 1.4 |
| Elemental S (g/L) | 0 | 0 | 0.5 | 1.0 | 0 | 0 |
| Initial mineral N | ||||||
| Without fertilizer | 11 | 59 | 10 | 18 | 25 | <5 |
| Total mineral N after 50 day incubation at 15 °C (n = 3) | ||||||
| Blood meal (3.5 g/L) | 11 (a) | 40 (b) | 10 (a) | 11 (a) | 15 (a) | 30 (b) |
| Chitin (2 g/L) | 29 (B) | 92 (D) | 9 (A) | 64 (C) | 21 (B) | 26 (B) |
| Total mineral N after 100 day incubation at 15 °C (n = 3) | ||||||
| Blood meal (3.5 g/L) | 9 (a) | 57 (c) | 9 (a) | 10 (a) | 32 (b) | 95 (d) |
| Chitin (2 g/L) | 27 (B) | 119 (D) | 8 (A) | 96 (C) | 26 (B) | 43 (B) |
| Total mineral N after 250 day incubation at 15 °C (n = 1) | ||||||
| Chitin (2 g/L) | 42 | 170 | 7 | 142 | 41 | 106 |
| Microbial biomass (nmol/g OM) (n = 1) | ||||||
| Without fertilizer | 265 | 346 | 457 | 574 | 228 | 150 |
| Blood meal (3.5 g/L) | 452 | 457 | 639 | 907 | 357 | 149 |
| Chitin (2 g/L) | 374 | 305 | 699 | 439 | 363 | 140 |
| M1 | M2 | M3 | M4 | M5 | ||
|---|---|---|---|---|---|---|
| pH-H2O | - | 6.7 | 6.5 | 7.4 | 7.3 | 5.5 |
| EC | µS/cm | 174 | 380 | 225 | 403 | 95 |
| Mineral N | mg/L substrate | <10 | 25 | <10 | <10 | 20 |
| P-AmAc | 35 | 126 | 47 | 156 | <14 | |
| K-AmAc | 578 | 1064 | 701 | 1256 | 158 | |
| Dry bulk density | g/L dry substrate | 227 | 237 | 203 | 226 | NA |
| Total pore volume | volume% | 88 | 87 | 89 | 88 | NA |
| Air volume −10 cm | 16 | 17 | 39 | 34 | NA | |
| Air volume −50 cm | 44 | 47 | 57 | 55 | NA | |
| Air volume −100 cm | 50 | 52 | 61 | 59 | NA | |
| Water volume −10 cm | 71 | 70 | 50 | 54 | NA | |
| Water volume −50 cm | 43 | 41 | 32 | 33 | NA | |
| Water volume −100 cm | 38 | 35 | 28 | 30 | NA | |
| Easily available water | 28 | 29 | 19 | 21 | NA | |
| Organic matter | %/DM | 61 | 60 | 63 | 57 | 90 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vandecasteele, B.; Van Loo, K.; Ommeslag, S.; Vierendeels, S.; Rooseleer, M.; Vandaele, E. Sustainable Growing Media Blends with Woody Green Composts: Optimizing the N Release with Organic Fertilizers and Interaction with Microbial Biomass. Agronomy 2022, 12, 422. https://doi.org/10.3390/agronomy12020422
Vandecasteele B, Van Loo K, Ommeslag S, Vierendeels S, Rooseleer M, Vandaele E. Sustainable Growing Media Blends with Woody Green Composts: Optimizing the N Release with Organic Fertilizers and Interaction with Microbial Biomass. Agronomy. 2022; 12(2):422. https://doi.org/10.3390/agronomy12020422
Chicago/Turabian StyleVandecasteele, Bart, Koen Van Loo, Sarah Ommeslag, Siebert Vierendeels, Maxim Rooseleer, and Elke Vandaele. 2022. "Sustainable Growing Media Blends with Woody Green Composts: Optimizing the N Release with Organic Fertilizers and Interaction with Microbial Biomass" Agronomy 12, no. 2: 422. https://doi.org/10.3390/agronomy12020422