
Beneficial Effects of Silicon Fertilizer on Growth and Physiological Responses in Oil Palm



Abstract
:1. Introduction
2. Materials and Methods
2.1. Pre-Plantation Soil Analysis
2.2. Seedling Transplantation and Adaptation
2.3. Treatment Application
2.4. Data Collection
2.5. Determination of Silicon and Nitrogen Contents in Oil Palm Seedlings
2.6. Statistical Analysis
3. Results
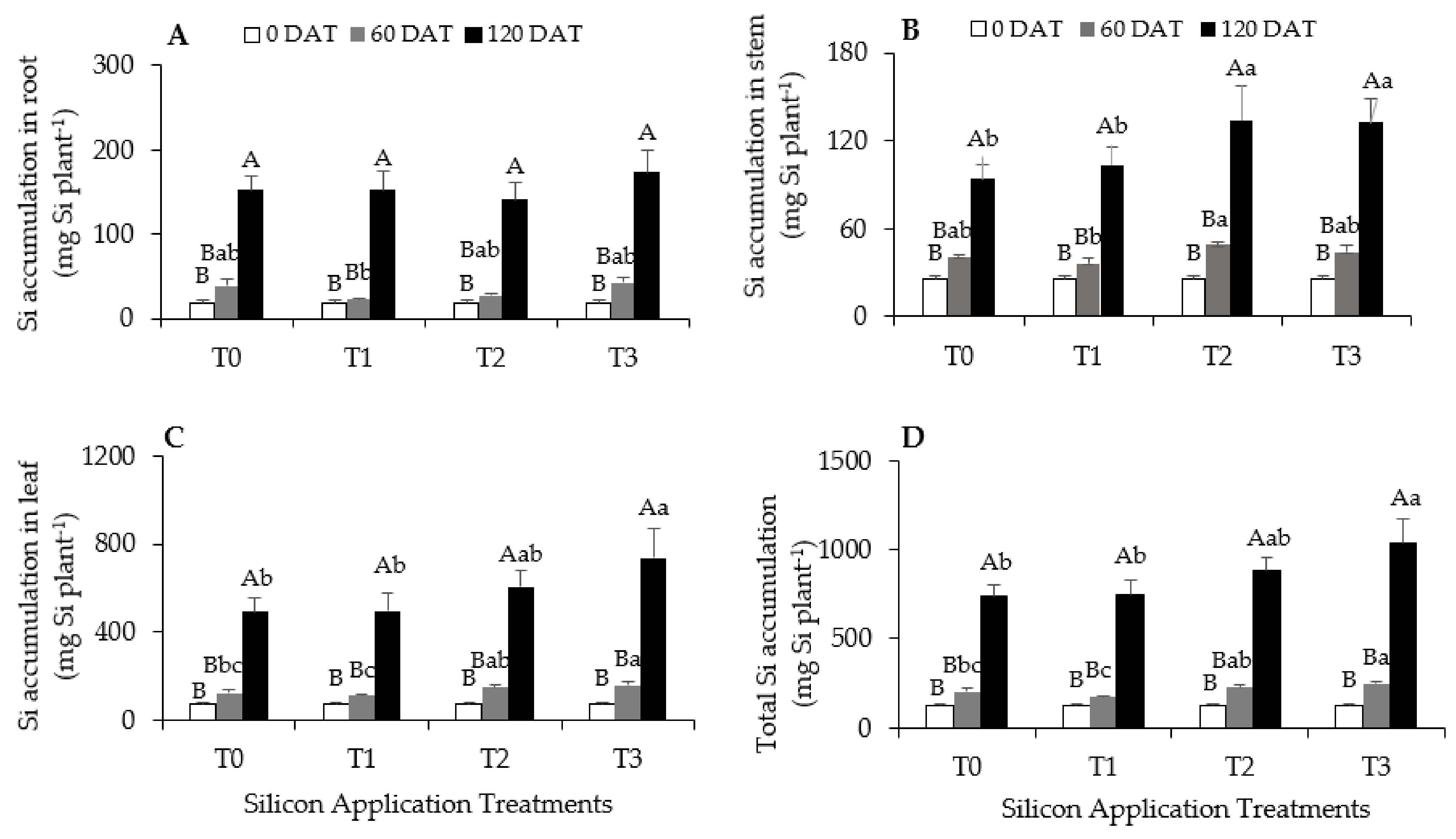
3.1. Accumulation and Distribution of Si in Oil Palm Seedlings
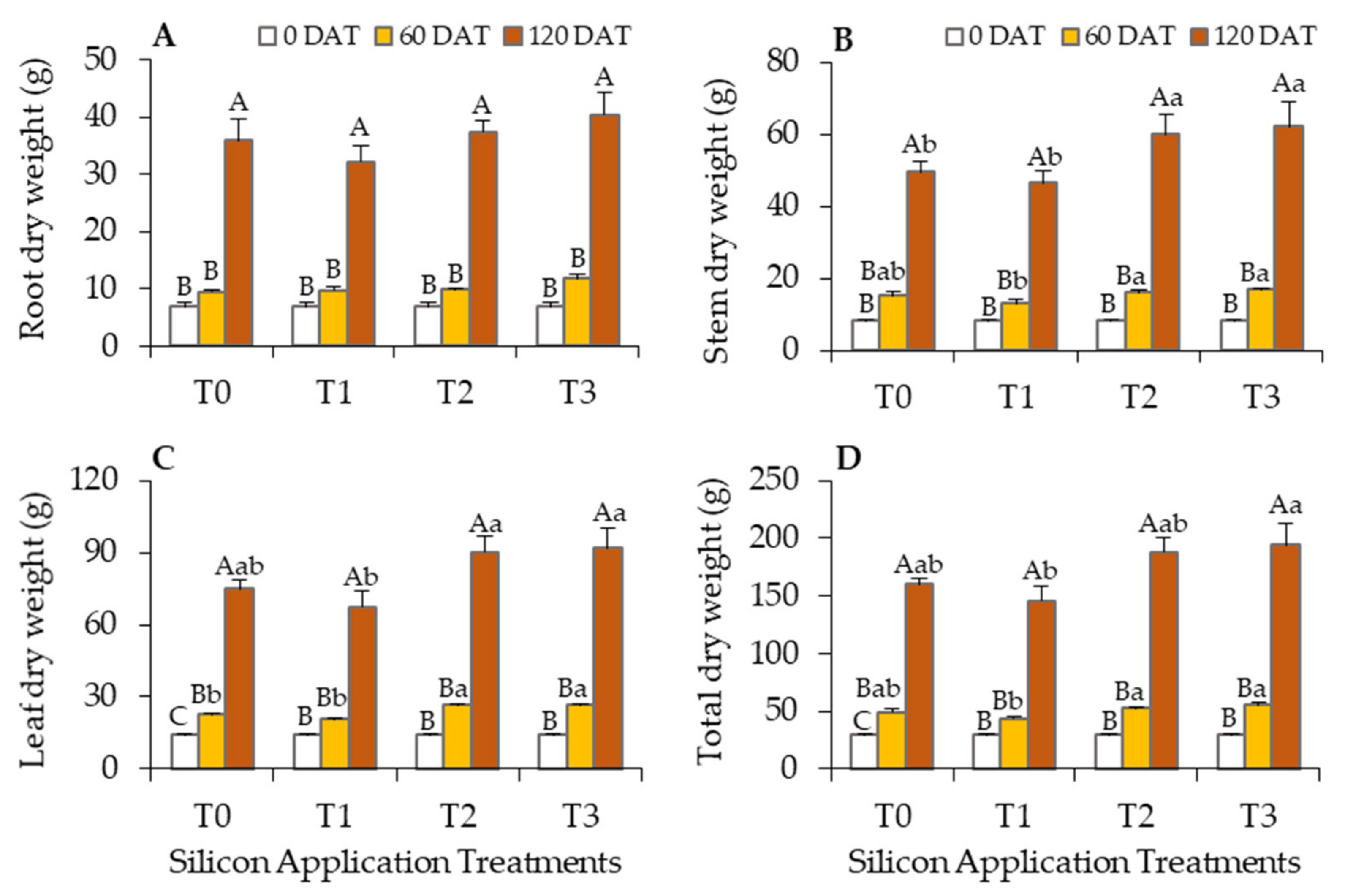
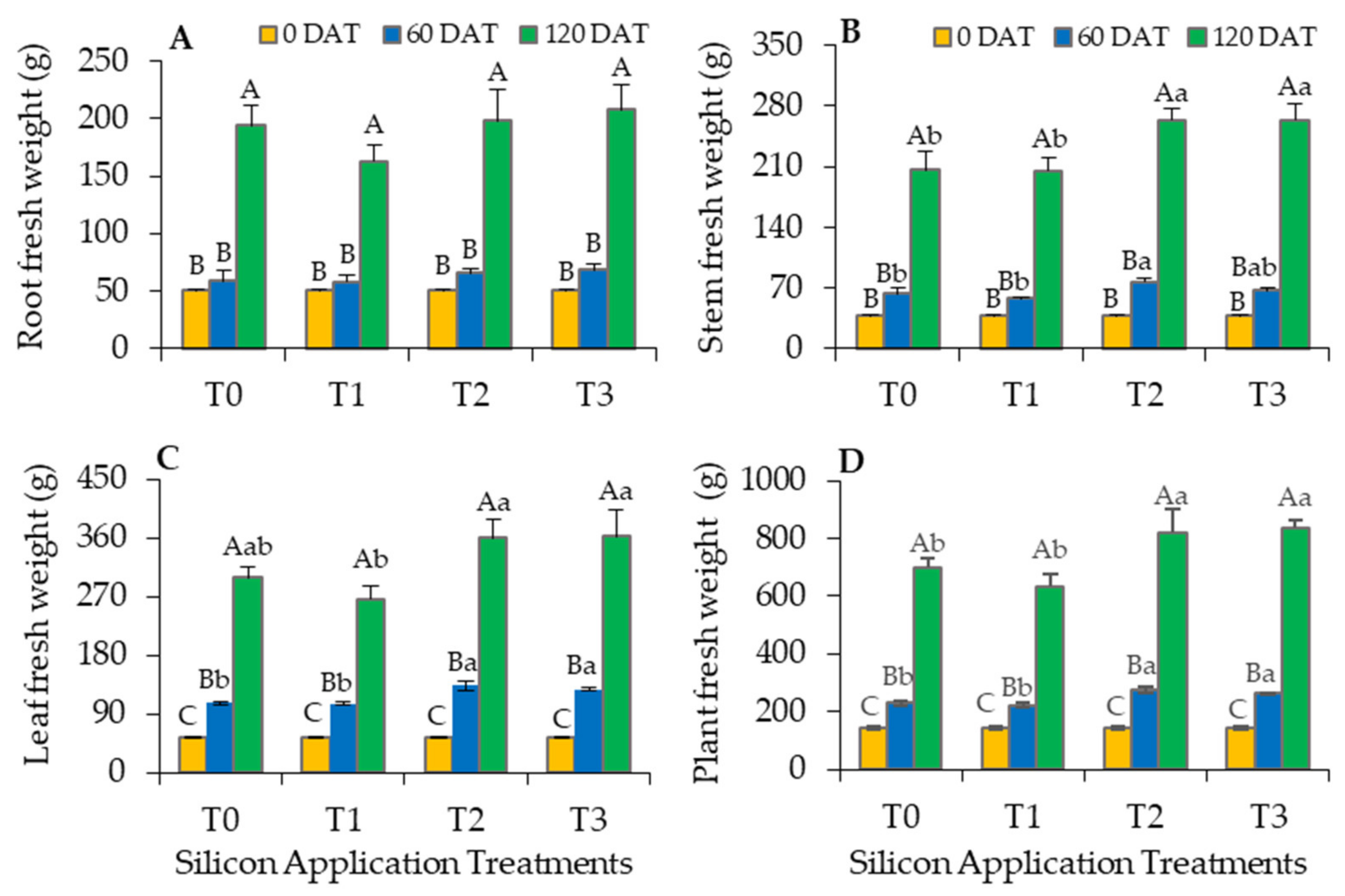
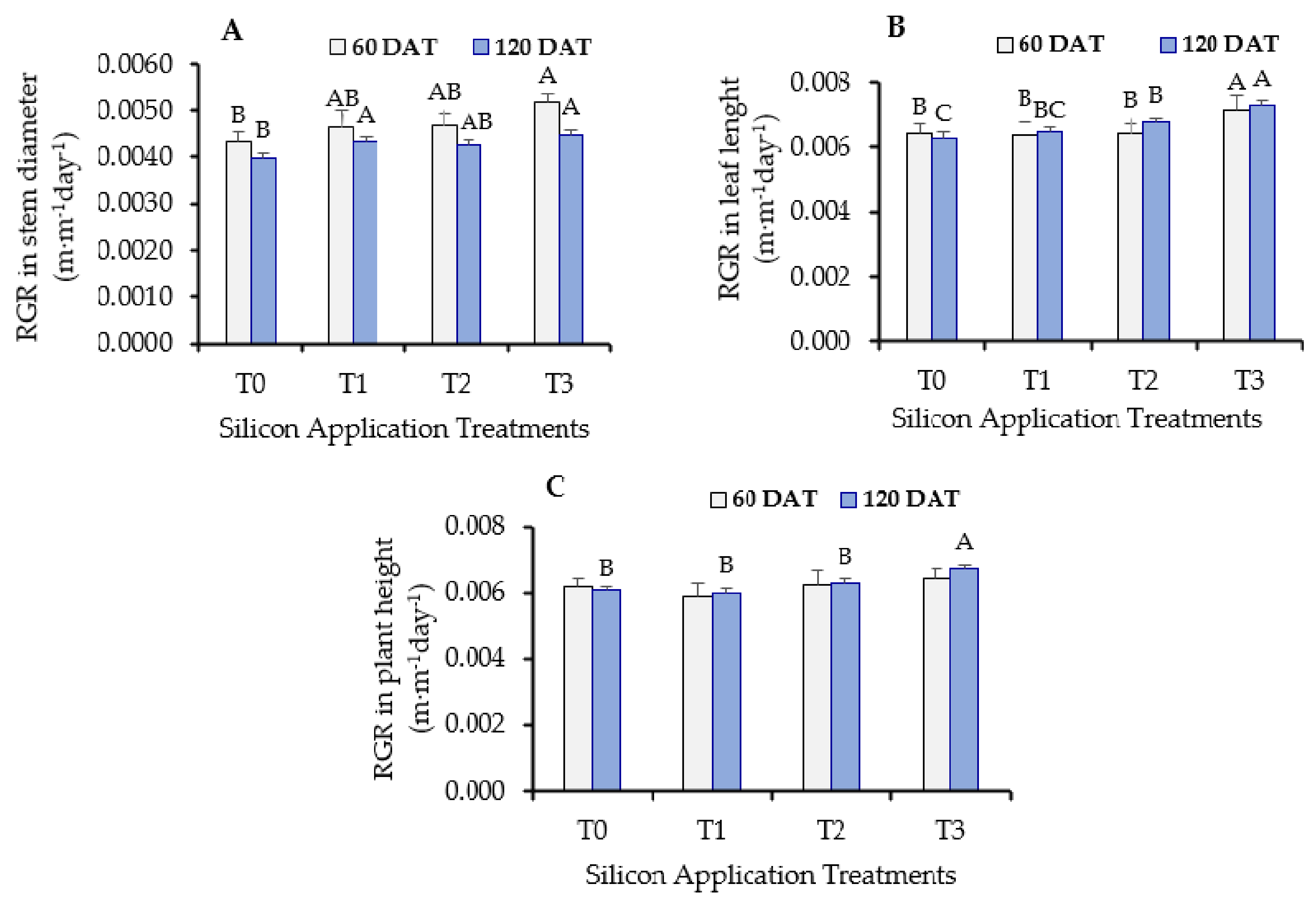
3.2. Positive Impacts of Silicon on Oil Palm Growth
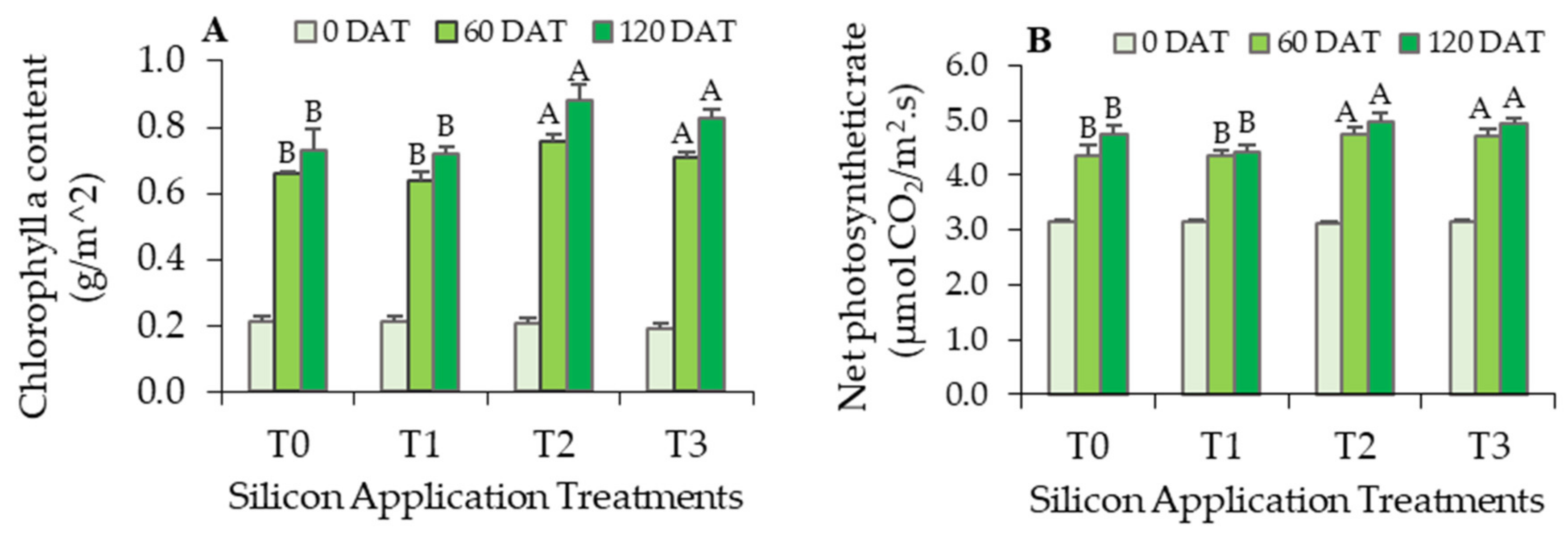
3.3. Effects of Silicon Fertilization on Physiological Responses of Oil Palm Seedlings
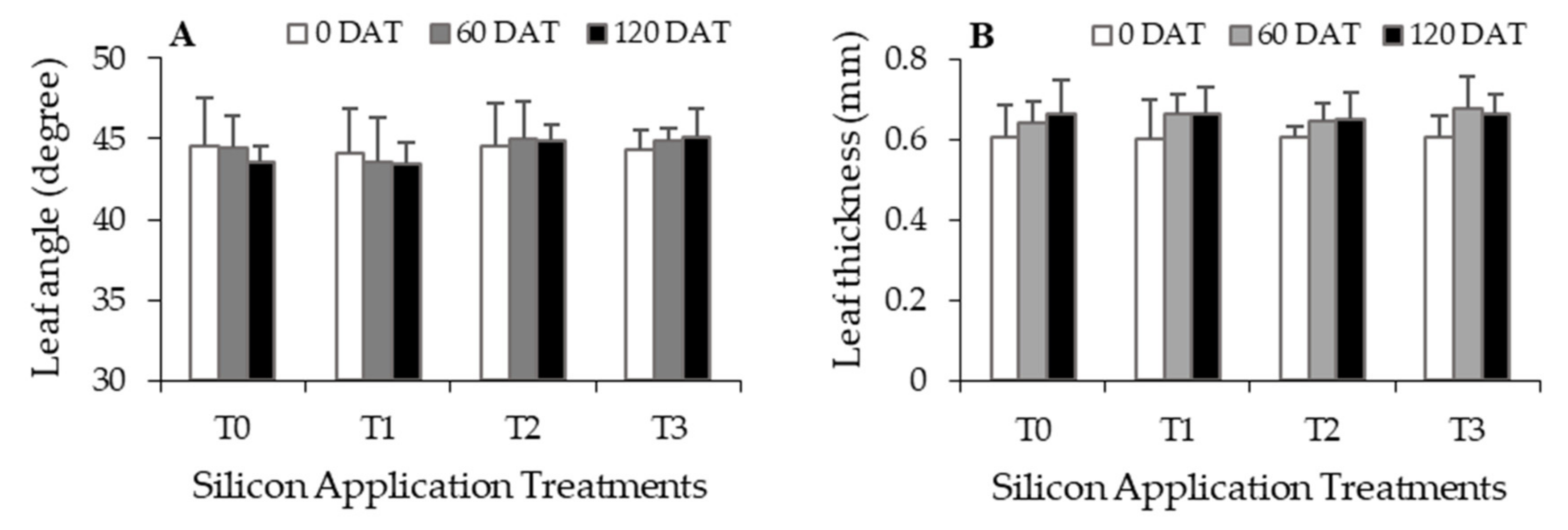
3.4. Effects of Silicon Fertilization on Oil Palm Leaf Morphology
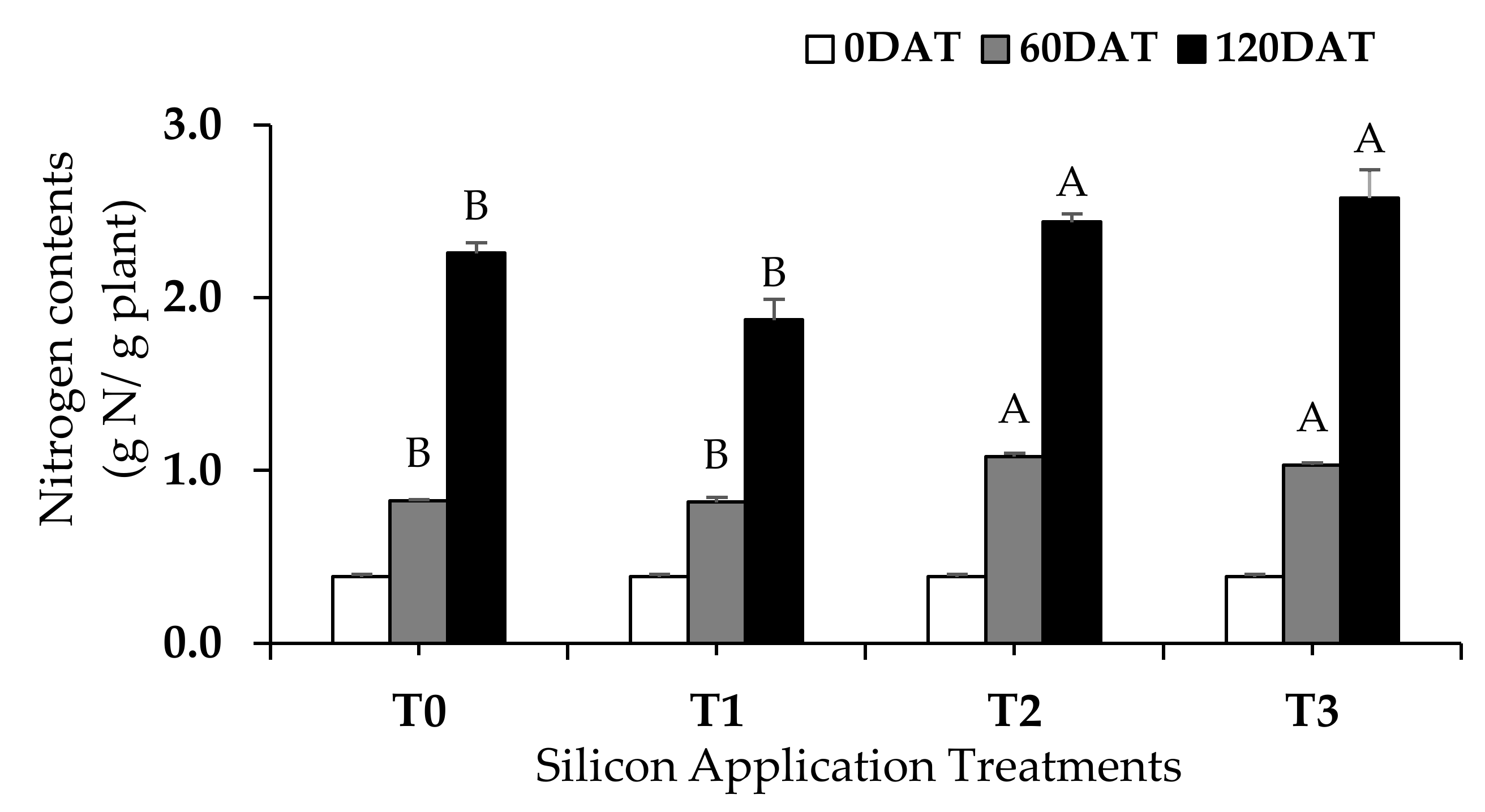
3.5. Effects of Silicon Fertilization on Nitrogen Accumulation in Oil Palm Seedlings
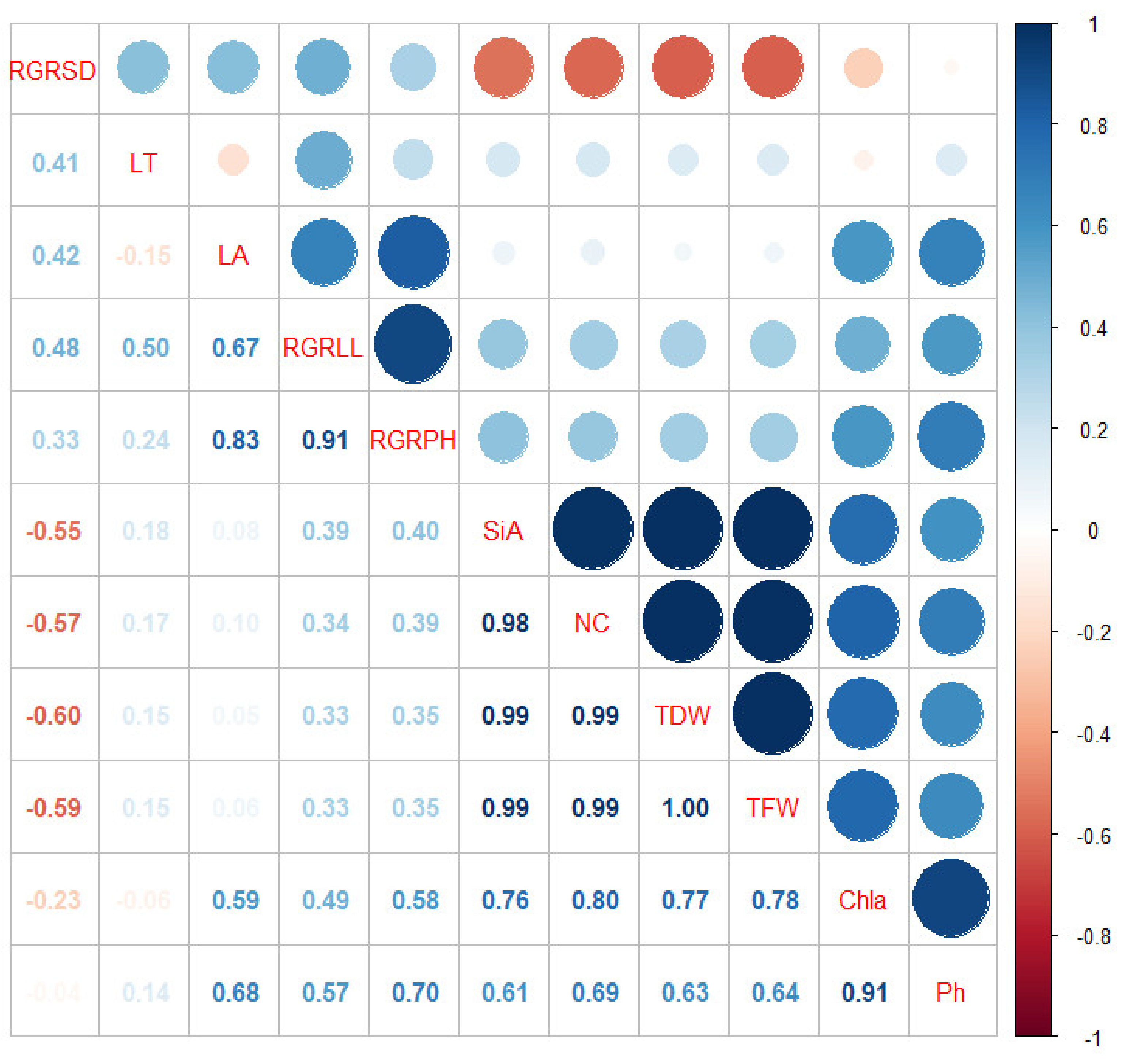
3.6. Correlation Assessment
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hodson, M.J.; Evans, D.E. Aluminium/silicon interactions in higher plants. J. Exp. Bot. 1995, 46, 161–171. [Google Scholar] [CrossRef]
- Hans Wedepohl, K. The composition of the continental crust. Geochim. Cosmochim. Acta 1995, 59, 1217–1232. [Google Scholar] [CrossRef]
- Sommer, M.; Kaczorek, D.; Kuzyakov, Y.; Breuer, J. Silicon pools and fluxes in soils and landscapes—A review. J. Soil Sci. Plant Nutr. 2006, 169, 310–329. [Google Scholar]
- Camargo, M.S.; Keeping, M.G. Silicon in sugarcane: Availability in soil, fertilization, and uptake. Silicon 2021, 13, 3691–3701. [Google Scholar] [CrossRef]
- Meena, V.D. A case for silicon fertilization to improve crop yields in tropical soils. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci 2014, 84, 505–518. [Google Scholar] [CrossRef] [Green Version]
- Dobermann, A.; Fairhurst, T. Rice: Nutrient Disorder & Nutrient Management; Potash & Phosphate Institute (PPI), Potash & Phosphate Institute of Canada (PPIC) and International Rice Research Institute: Laguna, Philippines, 2000; p. 191. [Google Scholar]
- Bari, M.A.; Prity, S.A.; Das, U.; Akther, M.S.; Sajib, S.A.; Reza, M.A.; Kabir, A.H. Silicon induces phytochelatin and ROS scavengers facilitating cadmium detoxification in rice. Plant Biol. 2020, 22, 472–479. [Google Scholar] [CrossRef]
- Chaiwong, N.; Rekasem, B.; Pusadee, T. Silicon application improves caryopsis development and yield in rice. J. Sci. Food Agric. 2021, 101, 220–228. [Google Scholar] [CrossRef]
- Geng, A.; Wang, X.; Wu, L.; Wang, F.; Wu, Z.; Yang, H.; Chen, Y.; Wen, D.; Liu, X. Silicon improves growth and alleviates oxidative stress in rice seedlings (Oryza sativa L.) by strengthening antioxidant defense and enhancing protein metabolism under arsanilic acid exposure. Ecotoxicol. Environ. Saf. 2018, 15, 266–273. [Google Scholar]
- Ahmad, M.; Ahmad, M.; El-Saeid, M.H.; Akram, M.A.; Ahmad, H.R.; Hussain, H.H.A. Silicon fertilization—A tool to boost up drought tolerance in wheat (Triticum aestivum L.) crop for better yield. J. Plant Nutr. 2016, 39, 1283–1291. [Google Scholar] [CrossRef]
- Verma, K.K.; Song, X.P.; Zeng, Y.; Guo, D.J.; Singh, M.; Rajput, V.D.; Malviya, M.K.; Wei, K.J.; Sharma, A.; Li, D.P.; et al. Foliar application of silicon boosts growth, photosynthetic leaf gas exchange, antioxidative response and resistance to limited water irrigation in sugarcane (Saccharum officinarum L.). Plant Physiol. Biochem. 2021, 166, 582–592. [Google Scholar] [CrossRef]
- Sun, X.; Liu, Q.; Tang, T.; Chen, X.; Luo, X. Silicon fertilizer application promotes phytolith accumulation in rice plants. Front. Plant Sci. 2019, 10, 425. [Google Scholar] [CrossRef]
- Ma, J.F.; Miyake, Y.; Takahashi, E. Silicon as a beneficial element for crop plants. In Studies in Plant Science; Datnoff, L.E., Snyder, G.H., Korndörfer, G.H., Eds.; Elsevier: Amsterdam, The Netherlands, 2001; pp. 17–39. [Google Scholar]
- Adatia, M.H.; Besford, R. The effects of silicon on cucumber plants grown in recirculating nutrient solution. Ann. Bot. 1986, 58, 343–351. [Google Scholar] [CrossRef]
- Hodson, M. Phylogenetic variation in the silicon composition of plants. Ann. Bot. 2005, 96, 1027–1046. [Google Scholar] [CrossRef] [Green Version]
- Mitani, N. Uptake system of silicon in different plant species. J. Exp. Bot. 2005, 56, 1255–1261. [Google Scholar] [CrossRef] [Green Version]
- Hossain, M.T.; Mori, R.; Soga, K.; Wakabayashi, K.; Kamisaka, S.; Fujii, S.; Yamamoto, R.; Hoson, T. Growth promotion and an increase in cell wall extensibility by silicon in rice and some other Poaceae seedlings. J. Plant Res. 2002, 115, 23–27. [Google Scholar] [CrossRef]
- Ahmed, M.; Qadeer, U.; Hassan, F.; Fahad, S.; Naseem, W.; Duangpan, S.; Ahmad, S. Abiotic Stress Tolerance in Wheat, and the Role of Silicon: An Experimental Evidence. In Agronomic Crops; Hasanuzzaman, M., Ed.; Springer: Singapore, 2020. [Google Scholar]
- Ahmad, T.; Haddad, R. Study of silicon effects on antioxidant enzyme activities and osmotic adjustment of wheat under drought stress. Czech J. Genet. Plant Breed. 2011, 47, 17–27. [Google Scholar] [CrossRef] [Green Version]
- Berahim, Z.; Omar, H.M.; Zakaria, N.; Ismail, M.R.; Rosle, R.; Roslin, N.A.; Che’ya, N.N. Silicon improves yield performance by enhancement in physiological responses, crop imagery, and leaf and culm sheath morphology in new rice line, PadiU Putra. BioMed Res. Int. 2021, 2021. [Google Scholar] [CrossRef]
- Verma, K.K.; Liu, X.H.; Wu, K.C.; Singh, K.R.; Song, Q.; Malviya, M.K.; Song, X.; Singh, P.; Verma, C.L.; Li, Y. The impact of silicon on photosynthetic and biochemical responses of sugarcane under different soil moisture levels. Silicon 2020, 12, 1355–1367. [Google Scholar] [CrossRef]
- Dorairaj, D.; Ismail, M.R.; Sinniah, U.R.; Ban, T.K. Influence of silicon on growth, yield, and lodging resistance of MR219, a lowland rice of Malaysia. J. Plant Nutr. 2017, 40, 1111–1124. [Google Scholar] [CrossRef]
- Song, A.; Li, P.; Li, Z.; Liang, Y. The effect of Silicon on photosynthesis and expression of its relevant genes in rice (Oryza sativa L.) under high-zinc stress. PLoS ONE 2014, 9, e113782. [Google Scholar]
- Ávila, F.; Baliza, D.; Faquin, V.; Araujo, J.; Ramos, S. Silicon-nitrogen interaction in rice cultivated under nutrient solution. Cienc. Agron. 2010, 41, 184–190. [Google Scholar] [CrossRef]
- Reynolds, O.L. Silicon: Potential to promote direct and indirect effects on plant defense against arthropod pests in agriculture. Front. Plant Sci. 2016, 7, 744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Massey, F.P.; Hartley, S.E. Experimental demonstration of the antiherbivore effects of silica in grasses: Impacts on foliage digestibility and vole growth rates. Proc. Biol. Sci. 2006, 273, 2299–2304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Massey, F.P.; Hartley, S.E. Physical defences wear you down: Progressive and irreversible impacts of silica on insect herbivores. J. Anim. Ecol. 2009, 78, 281–291. [Google Scholar] [CrossRef]
- Yang, R.; Howe, J.A.; Golden, B.R. Calcium silicate slag reduces drought stress in rice (Oryza sativa L.). J. Agron. Crop. Sci. 2019, 205, 353–361. [Google Scholar] [CrossRef]
- Pei, Z.F.; Ming, D.F.; Liu, D.; Wan, G.L.; Geng, X.X.; Gong, H.J.; Zhou, W.J. Silicon improves the tolerance to water-deficit stress induced by polyethylene glycol in wheat (Triticum aestivum L.) seedlings. J. Plant Growth Regul. 2010, 29, 106–115. [Google Scholar] [CrossRef]
- Parveen, A.; Liu, W.; Hussain, S.; Asghar, J.; Perveen, S.; Xiong, Y. Silicon priming regulates morpho-physiological growth and oxidative metabolism in maize under drought stress. Plants 2019, 8, 431. [Google Scholar] [CrossRef] [Green Version]
- Torabi, F.; Majd, A.; Enteshari, S. The effect of silicon on alleviation of salt stress in borage (Borago officinalis L.). Soil Sci. Plant Nutr. 2015, 61, 788–798. [Google Scholar] [CrossRef] [Green Version]
- Woittiez, L.; Wijk, M.; Slingerland, M.; van Noordwijk, M.; Giller, K. Yield gaps in oil palm: A quantitative review of contributing factors. Eur. J. Agron. 2017, 83, 57–77. [Google Scholar] [CrossRef]
- Corley, R.H.V.; Tinker, P.B. The Oil Palm, 4th ed.; Blackwell Publishing: Oxford, UK, 2003; p. 284. [Google Scholar]
- Xin, Y.; Sun, L.; Hansen, M.C. Biophysical and socioeconomic drivers of oil palm expansion in Indonesia. Environ. Res. Lett. 2021, 16, 034048. [Google Scholar] [CrossRef]
- Pilon-Smits, E.A.; Quinn, C.F.; Tapken, W.; Malagoli, M.; Schiavon, M. Physiological functions of beneficial elements. Curr. Opin. Plant Biol. 2009, 12, 267–274. [Google Scholar] [CrossRef]
- Munevar, M.; Romero, F.A. Soil and plant silicon status in oil palm crops in Colombia. Exp. Agric. 2015, 51, 382–392. [Google Scholar] [CrossRef]
- Onthong, J.; Osaki, M.; Nilnond, C.; Tadano, T. Phosphorus status of some highly weathered soils in Peninsular Thailand and availability in Relation to citrate and oxamate application. Soil Sci. Plant Nutr. 1999, 45, 627–637. [Google Scholar] [CrossRef] [Green Version]
- Bouyoucos, G.J. Directios for making mechanical analyses of soils by the hydrometer method. Soil Sci. 1936, 42, 225–230. [Google Scholar] [CrossRef]
- Kjeldahl, J. Neue Methode zur Bestimmung des Stickstoffs in organischen Korpern. Z. Anal. Chem. 1883, 22, 366–382. [Google Scholar] [CrossRef] [Green Version]
- Bray, R.H.; Kurtz, L.T. Determination of toal, organic, and available forms of phosphorus in soils. Soil Sci. 1945, 59, 39–46. [Google Scholar] [CrossRef]
- Estefan, G.; Sommer, R.; Ryan, J. Methods of Soil, Plant, and Water Analysis: A Manual for the West Asia and North Africa Region; International Center for Agricultural Research in the Dry Areas: Beirut, Lebanon, 2013. [Google Scholar]
- Hoffmann, W.; Poorter, H. Avoiding bias in calculations of relative growth rate. Ann. Bot. 2002, 90, 37–42. [Google Scholar] [CrossRef] [Green Version]
- Lepeduš, H.; Vidaković-Cifrek, Z.; Šebalj, I.; Dunić, J.A.; Cesar, V. Effects of low and high irradiation levels on growth and PSII efficiency in Lemna minor L. Acta Bot. Croat. 2020, 79, 185–192. [Google Scholar] [CrossRef]
- Wei, T.; Simko, V. R Package “Corrplot”: Visualization of a Correlation Matrix. Available online: https://github.com/taiyun/corrplot (accessed on 10 November 2021).
- Hussain, T.; Hussain, N.; Ahmed, M.; Nualsri, C.; Duangpan, S. Responses of lowland rice genotypes under terminal water stress and identification of drought tolerance to stabilize rice productivity in southern Thailand. Plants 2021, 10, 2565. [Google Scholar] [CrossRef]
- Duangpan, S.; Buapet, P.; Sujitto, S.; Eksomtramage, T. Early assessment of drought tolerance in oil palm D × P progenies using growth and physiological characters in seedling stage. Plant Genet. Resour. C 2018, 16, 544–554. [Google Scholar] [CrossRef]
- Guo, Z.G.; Liu, H.X.; Tian, F.P.; Zhang, Z.H.; Wang, S.M. Effect of silicon on the morphology of shoots and roots of alfalfa (Medicago sativa). Anim. Prod. Sci. 2006, 46, 1161–1166. [Google Scholar] [CrossRef]
- Costa, B.N.S.; Dias, G.M.G.; Costa, I.J.S.; Assis, F.A.; Silveira, F.A.S.; Pasqual, M. Effects of silicon on the growth and genetic stability of passion fruit. Acta Sci. 2016, 38, 503–511. [Google Scholar] [CrossRef] [Green Version]
- Shamshiripour, M.; Motesharezadeh, B.; Rahmani, H.A.; Alikhani, H.A.; Etesami, H. Optimal concentrations of silicon enhance the growth of soybean (Glycine Max, L.) cultivars by improving nodulation, root system architecture, and soil biological properties. Silicon 2021. [Google Scholar] [CrossRef]
- Tongchu, C.; Onthong, J.; Duangpan, S. Effects of calcium silicate fertilizer on silicon accumulation of oil palm seedling. Thai J. Soils Fertil. 2018, 41, 18–25. [Google Scholar]
- Putra, E.T.S.; Issukindarsyah, I.; Taryono, T.; Purwanto, B.H.; Indradewa, D. Role of boron and silicon in inducing mechanical resistance of oil palm seedlings to drought stress. J. Appl. Sci. 2016, 16, 242–251. [Google Scholar] [CrossRef] [Green Version]
- Luyckx, M.; Hausman, J.F.; Lutts, S.; Guerriero, G. Impact of silicon in plant biomass production: Focus on bast fibres, hypotheses, and perspectives. Plants 2017, 6, 37. [Google Scholar] [CrossRef] [Green Version]
- Pavlovic, J.; Kostic, L.; Bosnic, P.; Kirkby, E.A.; Nikolic, M. Interactions of silicon with essential and beneficial elements in plants. Front. Plant Sci. 2021, 12, 697592. [Google Scholar] [CrossRef]
- Bokor, B.; Ondoš, S.; Vaculík, M.; Bokorová, S.; Weidinger, M.; Lichtscheidl, I.; Turňa, J.; Lux, A. Expression of genes for Si uptake, accumulation, and correlation of Si with other elements in Ionome of Maize Kernel. Front. Plant Sci. 2017, 8, 1063. [Google Scholar] [CrossRef] [Green Version]
- Greger, M.; Landberg, T.; Vaculík, M. Silicon influences soil availability and accumulation of mineral nutrients in various plant species. Plants 2018, 7, 41. [Google Scholar] [CrossRef] [Green Version]
- Pati, S.; Pal, B.; Badole, S.; Hazra, G.C.; Mandal, B. Effect of silicon fertilization on growth, yield, and nutrient uptake of rice. Commun. Soil Sci. Plant Anal. 2016, 47, 284–290. [Google Scholar] [CrossRef]
- Cuong, T.X.; Ullar, H.; Datta, A.; Hanh, T.C. Effects of silicon-based fertilizer on growth, yield and nutrient uptake of rice in tropical zone of Vietnam. Rice Sci. 2017, 24, 283–290. [Google Scholar] [CrossRef]
- Gou, T.; Yang, L.; Hu, W.; Chen, X.; Zhu, Y.; Guo, J.; Gong, H. Silicon improves the growth of cucumber under excess nitrate stress by enhancing nitrogen assimilation and chlorophyll synthesis. Plant Physiol. Biochem. 2020, 152, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Gao, F.; Yang, J.; Zhen, X.; Li, Y.; Zhao, J.; Li, J.; Qian, B.; Yang, D.; Li, X. Photosynthetic characteristics and uptake and translocation of nitrogen in peanut in a wheat-peanut rotation system under different fertilizer management regimes. Front. Plant Sci. 2019, 10, 86. [Google Scholar] [CrossRef] [PubMed]
- Bassi, D.; Menossi, M.; Mattiello, L. Nitrogen supply influences photosynthesis establishment along the sugarcane leaf. Sci. Rep. 2018, 8, 2327. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Properties | Values/ Description | Methods | References/Instrument |
|---|---|---|---|
| Texture | Sandy clay loam | Hydrometer | Bouyoucos, 1936 [38] |
| pH | 4.76 | pH meter, soil/water = 1:5 | Seven Easy (Mettler Toledo) |
| Electrical conductivity (ds·m−1) | 0.303 | EC meter, soil/water = 1:5 | Seven Easy EC Meter (Mettler Toledo) |
| Total N (g·kg−1) | 0.262 | Kjeldahl method | Kjeldahl, 1883 [39] |
| Available P (mg·kg−1) | 1.729 | Bray II, molybdenum blue method | Bray, 1945 [40] |
| Extractable K+ (cmol·kg−1) | 0.073 | 1 M-NH4OAc (pH 7) atomic absorption spectrophotometry | Spectrophotometer |
| Extractable Mg (cmol·kg−1) | 0.020 | 1 M-NH4OAc (pH 7) atomic absorption spectrophotometry | Spectrophotometer |
| Extractable Ca (cmol·kg−1) | 0.034 | 1 M-NH4OAc (pH 7) atomic absorption spectrophotometry | Spectrophotometer |
| Available Si (mg·kg−1) | 8.774 | Yellow molybdenum blue method | Estefan et al., 2013 [41] |
| Si Fertilization Treatments | Si Concentration (% Dry Weight) | ||
|---|---|---|---|
| Root | Stem | Leaf | |
| T0 | 0.45 ± 0.01 | 0.17 ± 0.09b | 0.66 ± 0.02b |
| T1 | 0.44 ± 0.02 | 0.22 ± 0.01ab | 0.63 ± 0.05b |
| T2 | 0.45 ± 0.02 | 0.25 ± 0.01a | 0.74 ± 0.02ab |
| T3 | 0.40 ± 0.05 | 0.24 ± 0.01a | 0.89 ± 0.04a |
| F-test | ns | * | * |
| CV (%) | 11.60 | 7.80 | 8.54 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duangpan, S.; Tongchu, Y.; Hussain, T.; Eksomtramage, T.; Onthong, J. Beneficial Effects of Silicon Fertilizer on Growth and Physiological Responses in Oil Palm. Agronomy 2022, 12, 413. https://doi.org/10.3390/agronomy12020413
Duangpan S, Tongchu Y, Hussain T, Eksomtramage T, Onthong J. Beneficial Effects of Silicon Fertilizer on Growth and Physiological Responses in Oil Palm. Agronomy. 2022; 12(2):413. https://doi.org/10.3390/agronomy12020413
Chicago/Turabian StyleDuangpan, Saowapa, Yanipha Tongchu, Tajamul Hussain, Theera Eksomtramage, and Jumpen Onthong. 2022. "Beneficial Effects of Silicon Fertilizer on Growth and Physiological Responses in Oil Palm" Agronomy 12, no. 2: 413. https://doi.org/10.3390/agronomy12020413