
Arbuscular Mycorrhiza Symbiosis as a Factor of Asteraceae Species Invasion

Abstract
:1. Introduction
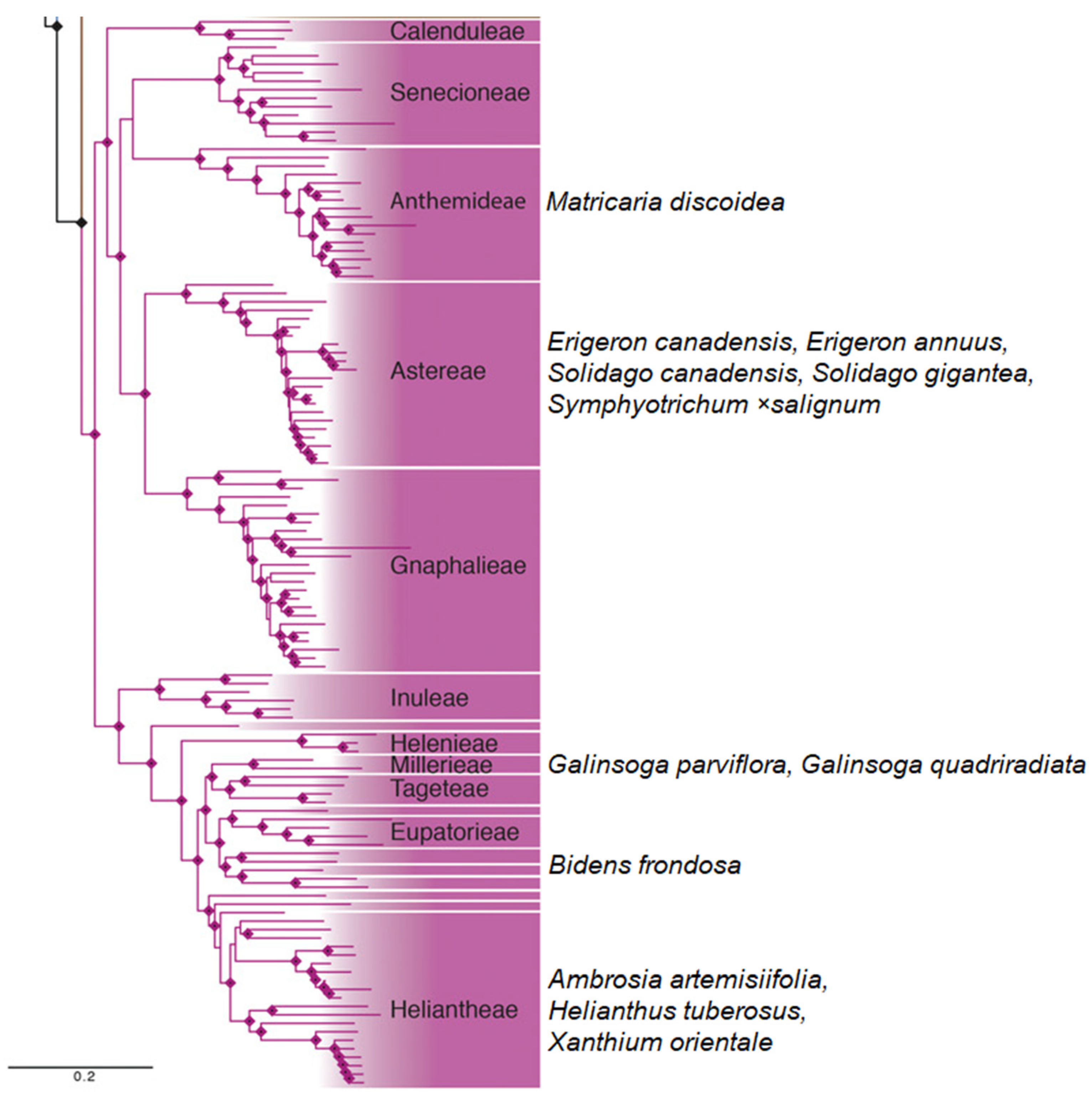
2. Invasive Asteraceae Species Associated with AMF
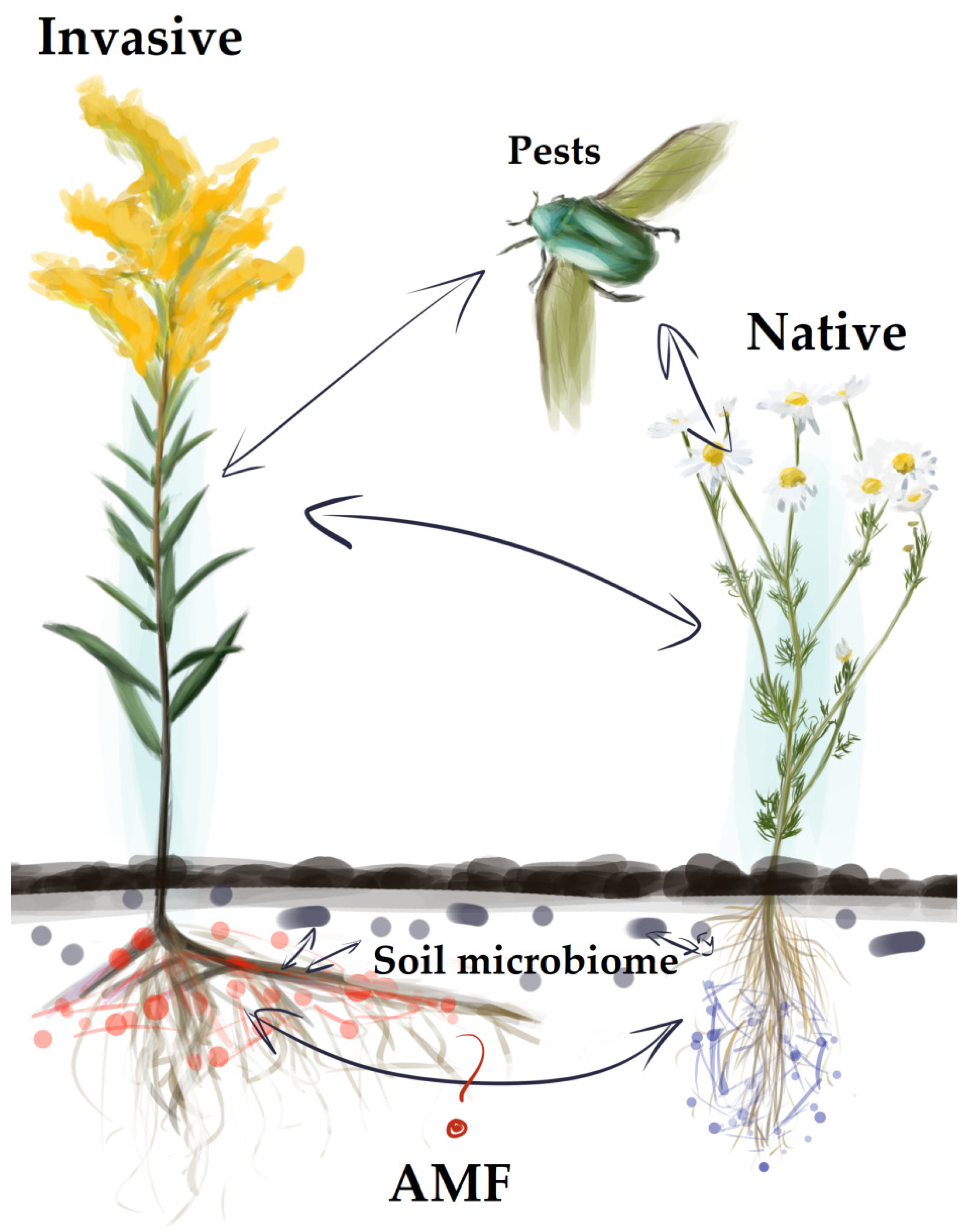
2.1. The Nature of Symbiotic Relationship between Invasive/Native Plants and AMF Communities
2.2. Arbuscular Mycorrhiza and Soil Microbiota
2.3. Common Mycorrhizal Networks
3. Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Medve, R. The Mycorrhizae of Pioneer Species in Disturbed Ecosystems in Western Pennsylvania. Am. J. Bot. 1984, 71, 787–794. [Google Scholar] [CrossRef]
- Mehraj, G.; Khuroo, A.; Hamid, M.; Muzafar, I.; Rashid, I.; Malik, A. Floristic Diversity and Correlates of Naturalization of Alien Flora in Urban Green Spaces of Srinagar City. Urban Ecosyst. 2021, 24, 1231–1244. [Google Scholar] [CrossRef]
- Day, N.; Dunfield, K.; Antunes, P. Fungi from a Non-Native Invasive Plant Increase Its Growth but Have Different Growth Effects on Native Plants. Biol. Invasions 2016, 18, 231–243. [Google Scholar] [CrossRef]
- Bongard, C.; Butler, K.; Fulthorpe, R. Investigation of Fungal Root Colonizers of the Invasive Plant Vincetoxicum rossicum and Co-Occurring Local Native Plants in a Field and Woodland Area in Southern Ontario. Nat. Conserv. 2013, 4, 55–76. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Y.; Tang, J.; Leng, D.; Hu, S.; Yong, J.W.H.; Chen, X. An Invasive Plant Promotes Its Arbuscular Mycorrhizal Symbioses and Competitiveness through Its Secondary Metabolites: Indirect Evidence from Activated Carbon. PLoS ONE 2014, 9, e97163. [Google Scholar] [CrossRef]
- Li, Y.-P.; Feng, Y.-L.; Kang, Z.-L.; Zhang, J.-L.; Yajun, C. Changes in Soil Microbial Communities Due to Biological Invasions Can Reduce Allelopathic Effects. J. Appl. Ecol. 2017, 54, 1281–1290. [Google Scholar] [CrossRef] [Green Version]
- Chagnon, P.; Bradley, R.L.; Klironomos, J.N. Mycorrhizal Network Assembly in a Community Context: The Presence of Neighbours Matters. J. Ecol. 2020, 108, 366–377. [Google Scholar] [CrossRef]
- Qin, F.; Yu, S. Arbuscular Mycorrhizal Fungi Protect Native Woody Species from Novel Weapons. Plant Soil 2019, 440, 39–52. [Google Scholar] [CrossRef]
- Rydlová, J.; Vosátka, M. Associations of Dominant Plant Species with Arbuscular Mycorrhizal Fungi during Vegetation Development on Coal Mine Spoil Banks. Folia Geobot. 2001, 36, 85–97. [Google Scholar] [CrossRef]
- Torrecillas, E.; Alguacil, M.M.; Roldán, A. Host Preferences of Arbuscular Mycorrhizal Fungi Colonizing Annual Herbaceous Plant Species in Semiarid Mediterranean Prairies. Appl. Environ. Microbiol. 2012, 78, 6180–6186. [Google Scholar] [CrossRef]
- Gucwa-Przepióra, E.; Błaszkowski, J. Arbuscular Mycorrhiza of Plants Spontaneously Colonizing the Soda Heap in Jaworzno (Southern Poland). Acta Soc. Bot. Pol. 2011, 76, 69–74. [Google Scholar] [CrossRef] [Green Version]
- Mummey, D.L.; Rillig, M.C. The Invasive Plant Species Centaurea maculosa Alters Arbuscular Mycorrhizal Fungal Communities in the Field. Plant Soil 2006, 288, 81–90. [Google Scholar] [CrossRef]
- Selvaraj, T.; Murugan, R.; Bhaskaran, C. Arbuscular Mycorrhizal Association of Kashini (Cichorium intybus L.) in Relationto Physicochemical Characters. Mycorrhiza News 2001, 13, 14–16. [Google Scholar]
- Oliveira, R.S.; Vosátka, M.; Dodd, J.C.; Castro, P.M.L. Studies on the Diversity of Arbuscular Mycorrhizal Fungi and the Efficacy of Two Native Isolates in a Highly Alkaline Anthropogenic Sediment. Mycorrhiza 2005, 16, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Dhar, P.P.; Al-Qarawi, A.A.; Mridha, M.A.U. Arbuscular Mycorrhizal Fungal Association in Asteraceae Plants Growing in the Arid Lands of Saudi Arabia. J. Arid Land 2015, 7, 676–686. [Google Scholar] [CrossRef]
- Allen, N.; Nordlander, M.; McGonigle, T.; Basinger, J.; Kaminskyj, S. Arbuscular Mycorrhizae on Axel Heiberg Island (80° N) and at Saskatoon (52° N) Canada. Can. J. Bot. 2006, 84, 1094–1100. [Google Scholar] [CrossRef]
- Renker, C.; Blanke, V.; Buscot, F. Diversity of Arbuscular Mycorrhizal Fungi in Grassland Spontaneously Developed on Area Polluted by a Fertilizer Plant. Environ. Pollut. 2005, 135, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Gorbunova, A.O.; Sumina, O.I. Dynamics of mycorrization in some plant species during progressive succession on sand quarries (Leningrad region). Bot. Zhurnal 2021, 106, 22–42. [Google Scholar] [CrossRef]
- Sinegani, A.; Yeganeh, M. The occurrence of arbuscular mycorrhizal fungi in soil and root of medicinal plants in Bu-Ali Sinagarden in Hamadan, Iran. Biol. J. Microorg. 2017, 5, 43–59. [Google Scholar] [CrossRef]
- Štajerová, K.; Smilauerova, M.; Šmilauer, P. Arbuscular Mycorrhizal Symbiosis of Herbaceous Invasive Neophytes in the Czech Republic. Preslia 2009, 81, 341–355. [Google Scholar]
- Crişan, I.; Vidican, R. Root Colonization by Micromycetes in Ten Asteraceae Species from Cluj County. J. Hortic. For. Biotechnol. 2019, 23, 15–57. [Google Scholar]
- Paudel, S.; Baer, S.G.; Battaglia, L.L. Arbuscular Mycorrhizal Fungi (AMF) and Success of Triadica sebifera Invasion in Coastal Transition Ecosystems along the Northern Gulf of Mexico. Plant Soil 2014, 378, 337–349. [Google Scholar] [CrossRef]
- Majewska, M.L.; Błaszkowski, J.; Nobis, M.; Rola, K.; Nobis, A.; Łakomiec, D.; Czachura, P.; Zubek, S. Root-Inhabiting Fungi in Alien Plant Species in Relation to Invasion Status and Soil Chemical Properties. Symbiosis 2015, 65, 101–115. [Google Scholar] [CrossRef]
- Zubek, S.; Majewska, M.L.; Błaszkowski, J.; Stefanowicz, A.M.; Nobis, M.; Kapusta, P. Invasive Plants Affect Arbuscular Mycorrhizal Fungi Abundance and Species Richness as Well as the Performance of Native Plants Grown in Invaded Soils. Biol. Fertil. Soils 2016, 52, 879–893. [Google Scholar] [CrossRef] [Green Version]
- Cumming, J.R.; Kelly, C.N. Pinus virginiana Invasion Influences Soils and Arbuscular Mycorrhizae of a Serpentine Grassland. J. Torrey Bot. Soc. 2007, 134, 63–73. [Google Scholar] [CrossRef]
- Liu, G.; Liu, R.-L.; Zhang, W.-G.; Yang, Y.-B.; Bi, X.-Q.; Li, M.-Z.; Chen, X.-Y.; Nie, H.; Zhu, Z.-H. Arbuscular Mycorrhizal Colonization Rate of an Exotic Plant, Galinsoga quadriradiata, in Mountain Ranges Changes with Altitude. Mycorrhiza 2021, 31, 161–171. [Google Scholar] [CrossRef]
- Fracchia, S.; Aranda, A.; Gopar, A.; Silvani, V.; Fernandez, L.; Godeas, A. Mycorrhizal Status of Plant Species in the Chaco Serrano Woodland from Central Argentina. Mycorrhiza 2009, 19, 205–214. [Google Scholar] [CrossRef]
- Li, L.-Q.; Zhang, M.-S.; Liang, Z.-P.; Xiao, B.; Wan, F.; Liu, W.-X. Arbuscular Mycorrhizal Fungi Enhance Invasive Plant, Ageratina adenophora Growth and Competition with Native Plants. Chin. J. Ecol. 2016, 35, 79–86. [Google Scholar] [CrossRef]
- Stevens, K.J.; Wall, C.B.; Janssen, J.A. Effects of Arbuscular Mycorrhizal Fungi on Seedling Growth and Development of Two Wetland Plants, Bidens frondosa L., and Eclipta prostrata (L.) L., Grown under Three Levels of Water Availability. Mycorrhiza 2011, 21, 279–288. [Google Scholar] [CrossRef]
- Sun, Y.; Umer, M.; Wu, P.; Guo, Y.; Ren, W.; Han, X.; Li, Q.; Wu, B.; Shen, K.; Xia, T.; et al. Indigenous Microorganisms Offset the Benefits of Growth and Nutrition Regulated by Inoculated Arbuscular Mycorrhizal Fungi for Four Pioneer Herbs in Karst Soil. PLoS ONE 2022, 17, e0266526. [Google Scholar] [CrossRef]
- Zhang, F.; Li, Q.; Yerger, E.; Chen, X.; Shi, Q.; Wan, F. AM Fungi Facilitate the Competitive Growth of Two Invasive Plant Species, Ambrosia artemisiifolia and Bidens pilosa. Mycorrhiza 2018, 28, 703–715. [Google Scholar] [CrossRef] [PubMed]
- Nacoon, S.; Jogloy, S.; Riddech, N.; Mongkolthanaruk, W.; Ekprasert, J.; Cooper, J.; Boonlue, S. Combination of Arbuscular Mycorrhizal Fungi and Phosphate Solubilizing Bacteria on Growth and Production of Helianthus tuberosus under Field Condition. Sci. Rep. 2021, 11, 6501. [Google Scholar] [CrossRef]
- Spatafora, J.; Chang, Y.; Benny, G.; Lazarus, K.; Smith, M.; Berbee, M.; Bonito, G.; Corradi, N.; Grigoriev, I.; Gryganskyi, A.; et al. A Phylum-Level Phylogenetic Classification of Zygomycete Fungi Based on Genome-Scale Data. Mycologia 2016, 108, 1028–1046. [Google Scholar] [CrossRef] [PubMed]
- Hibbett, D.S.; Binder, M.; Bischoff, J.F.; Blackwell, M.; Cannon, P.F.; Eriksson, O.E.; Huhndorf, S.; James, T.; Kirk, P.M.; Lücking, R.; et al. A Higher-Level Phylogenetic Classification of the Fungi. Mycol. Res. 2007, 111, 509–547. [Google Scholar] [CrossRef] [PubMed]
- James, T.Y.; Stajich, J.E.; Hittinger, C.T.; Rokas, A. Toward a Fully Resolved Fungal Tree of Life. Annu. Rev. Microbiol. 2020, 74, 291–313. [Google Scholar] [CrossRef]
- Karandashov, V.; Bucher, M. Symbiotic Phosphate Transport in Arbuscular Mycorrhizas. Trends Plant Sci. 2005, 10, 22–29. [Google Scholar] [CrossRef]
- Govindarajulu, M.; Jin, H.; Abubaker, J.; Douds, D.; Allen, J.; Bücking, H.; Lammers, P.; Shachar-Hill, Y. Nitrogen Transfer in the Arbuscular Mycorrhizal Symbiosis. Nature 2005, 435, 819–823. [Google Scholar] [CrossRef]
- Cavagnaro, T.R. The Role of Arbuscular Mycorrhizas in Improving Plant Zinc Nutrition under Low Soil Zinc Concentrations: A Review. Plant Soil 2008, 304, 315–325. [Google Scholar] [CrossRef]
- Allen, J.W.; Shachar-Hill, Y. Sulfur Transfer through an Arbuscular Mycorrhiza. Plant Physiol. 2009, 149, 549–560. [Google Scholar] [CrossRef] [Green Version]
- Lehmann, A.; Rillig, M.C. Understanding Mechanisms of Soil Biota Involvement in Soil Aggregation: A Way Forward with Saprobic Fungi? Soil Biol. Biochem. 2015, 88, 298–302. [Google Scholar] [CrossRef]
- Kobae, Y.; Ohtomo, R.; Morimoto, S.; Sato, D.; Nakagawa, T.; Oka, N.; Sato, S. Isolation of Native Arbuscular Mycorrhizal Fungi within Young Thalli of the Liverwort Marchantia paleacea. Plants 2019, 8, 142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harrier, L.A.; Watson, C.A. The Potential Role of Arbuscular Mycorrhizal (AM) Fungi in the Bioprotection of Plants against Soil-Borne Pathogens in Organic and/or Other Sustainable Farming Systems: Role of Arbuscular Mycorrhizal Fungi in Bioprotection of Plants. Pest Manag. Sci. 2004, 60, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Jin, Y.; Tang, J.; Chen, X. The Invasive Plant Solidago canadensis L. Suppresses Local Soil Pathogens through Allelopathy. Appl. Soil Ecol. 2009, 41, 215–222. [Google Scholar] [CrossRef]
- Zhang, S.; Zhu, W.; Wang, B.; Tang, J.; Chen, X. Secondary Metabolites from the Invasive Solidago canadensis L. Accumulation in Soil and Contribution to Inhibition of Soil Pathogen Pythium ultimum. Appl. Soil Ecol. 2011, 48, 280–286. [Google Scholar] [CrossRef]
- Qu, L.; Wang, M.; Biere, A. Interactive Effects of Mycorrhizae, Soil Phosphorus, and Light on Growth and Induction and Priming of Defense in Plantago lanceolata. Front. Plant Sci. 2021, 12. [Google Scholar] [CrossRef]
- Hoeksema, J.; Chaudhary, B.; Gehring, C.; Johnson, N.; Karst, J.; Koide, R.; Pringle, A.; Zabinski, C.; Bever, J.; Moore, J.; et al. A Meta-Analysis of Context-Dependency in Plant Response to Inoculation with Mycorrhizal Fungi. Ecol. Lett. 2010, 13, 394–407. [Google Scholar] [CrossRef]
- Sandrini, M.; Nerva, L.; Sillo, F.; Balestrini, R.; Chitarra, W.; Zampieri, E. Abiotic Stress and Belowground Microbiome: The Potential of Omics Approaches. Int. J. Mol. Sci. 2022, 23, 1091. [Google Scholar] [CrossRef]
- Jin, L.; Gu, Y.; Xiao, M.; Chen, J.; Li, B. The History of Solidago canadensis Invasion and the Development of Its Mycorrhizal Associations in Newly-Reclaimed Land. Funct. Plant Biol. 2004, 31, 979. [Google Scholar] [CrossRef]
- Höpfner, I.; Beyschlag, W.; Bartelheimer, M.; Werner, C.; Unger, S. Role of Mycorrhization and Nutrient Availability in Competitive Interactions between the Grassland Species Plantago lanceolata and Hieracium pilosella. Plant Ecol. 2015, 216, 887–899. [Google Scholar] [CrossRef]
- Jentschke, G.; Godbold, D. Metal Toxicity and Ectomycorrhizas. Physiol. Plant. 2000, 109, 107–116. [Google Scholar] [CrossRef] [Green Version]
- Whipps, J.M. Prospects and Limitations for Mycorrhizas in Biocontrol of Root Pathogens. Can. J. Bot. 2004, 82, 1198–1227. [Google Scholar] [CrossRef]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis, 3rd ed.; Academic Press: Amsterdam, The Netherlands; Boston, MA, USA, 2008. [Google Scholar]
- Adolfsson, L.; Solymosi, K.; Andersson, M.X.; Keresztes, Á.; Uddling, J.; Schoefs, B.; Spetea, C. Mycorrhiza Symbiosis Increases the Surface for Sunlight Capture in Medicago truncatula for Better Photosynthetic Production. PLoS ONE 2015, 10, e0115314. [Google Scholar] [CrossRef] [PubMed]
- Fumanal, B.; Plenchette, C.; Chauvel, B.; Bretagnolle, F. Which Role Can Arbuscular Mycorrhizal Fungi Play in the Facilitation of Ambrosia artemisiifolia L. Invasion in France? Mycorrhiza 2006, 17, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Kong, L.; Chen, X.; Yerger, E.H.; Li, Q.; Chen, F.; Xu, H.; Zhang, F. Arbuscular Mycorrhizal Fungi Enhance the Growth of the Exotic Species Ambrosia artemisiifolia. J. Plant Ecol. 2022, 15, 581–595. [Google Scholar] [CrossRef]
- Řezáčová, V.; Řezáč, M.; Gryndler, M.; Hršelová, H.; Gryndlerová, H.; Michalová, T. Plant Invasion Alters Community Structure and Decreases Diversity of Arbuscular Mycorrhizal Fungal Communities. Appl. Soil Ecol. 2021, 167, 104039. [Google Scholar] [CrossRef]
- Bunn, R.; Ramsey, P.; Lekberg, Y. Do Native and Invasive Plants Differ in Their Interactions with, Arbuscular Mycorrhizal Fungi? A Meta-Analysis. J. Ecol. 2015, 103, 1547–1556. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.-J.; Li, Q.; Chen, F.-X.; Xu, H.-Y.; Inderjit; Wan, F.-H. Arbuscular Mycorrhizal Fungi Facilitate Growth and Competitive Ability of an Exotic Species Flaveria bidentis. Soil Biol. Biochem. 2017, 115, 275–284. [Google Scholar] [CrossRef]
- Cheng, J.-K.; Yue, M.-F.; Yang, H.-R.; Chen, B.-M.; Xin, G.-R. Do Arbuscular Mycorrhizal Fungi Help the Native Species Bidens biternata Resist the Invasion of Bidens Alba? Plant Soil 2019, 444, 443–455. [Google Scholar] [CrossRef]
- Klironomos, J. Feedback with Soil Biota Contributes to Plant Rarity and Invasiveness in Communities. Nature 2002, 417, 67–70. [Google Scholar] [CrossRef]
- Ashton, I.; Hyatt, L.A.; Howe, K. Invasive Species Accelerate Decomposition and Litter Nitrogen Loss in a Mixed Deciduous Forest. Ecol. Appl. 2005, 15, 1263–1272. [Google Scholar] [CrossRef]
- Stinson, K.; Campbell, S.; Powell, J.; Wolfe, B.; Callaway, R.; Thelen, G.; Hallett, S.; Prati, D.; Klironomos, J. Invasive Plant Suppresses the Growth of Native Tree Seedlings by Disrupting Belowground Mutualisms. PLoS Biol. 2006, 4, e140. [Google Scholar] [CrossRef] [PubMed]
- Vogelsang, K.; Bever, J. Mycorrhizal Densities Decline in Association with Nonnative Plants and Contribute to Plant Invasion. Ecology 2009, 90, 399–407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, X.; Chen, W.; Ivanov, S.; MacLean, A.; Wight, H.; Ramaraj, T.; Mudge, J.; Harrison, M.; Fei, Z. Genome and Evolution of the Arbuscular Mycorrhizal Fungus Diversispora epigaea (Formerly Glomus versiforme) and Its Bacterial Endosymbionts. New Phytol. 2018, 221, 1556–1573. [Google Scholar] [CrossRef] [PubMed]
- Xing, Y.; Yang, Y.; Xu, L.; Hao, N.; Zhao, R.; Wang, J.; Li, S.; Zhang, D.; Zhang, T.; Kang, T. The Diversity of Associated Microorganisms in Different Organs and Rhizospheric Soil of Arctium lappa L. Curr. Microbiol. 2020, 77, 746–754. [Google Scholar] [CrossRef]
- Emmett, B.; Lévesque-Tremblay, V.; Harrison, M. Conserved and Reproducible Bacterial Communities Associate with Extraradical Hyphae of Arbuscular Mycorrhizal Fungi. ISME J. 2021, 15, 2276–2288. [Google Scholar] [CrossRef]
- Rozmoš, M.; Bukovská, P.; Hrselova, H.; Kotianová, M.; Dudáš, M.; Gančarčíková, K.; Jansa, J. Organic Nitrogen Utilisation by an Arbuscular Mycorrhizal Fungus Is Mediated by Specific Soil Bacteria and a Protist. ISME J. 2021, 16, 676–685. [Google Scholar] [CrossRef]
- Zhang, Q.; Yao, L.; Yang, R.; Yang, X.; Tang, J.; Chen, X. Potential Allelopathic Effects of an Invasive Species Solidago canadensis on the Mycorrhizae of Native Plant Species. Allelopathy J. 2007, 20, 71–77. [Google Scholar]
- Thorpe, A.S.; Thelen, G.C.; Diaconu, A.; Callaway, R.M. Root Exudate Is Allelopathic in Invaded Community but Not in Native Community: Field Evidence for the Novel Weapons Hypothesis. J. Ecol. 2009, 97, 641–645. [Google Scholar] [CrossRef]
- Kato-Noguchi, H.; Kato, M. Allelopathy and Allelochemicals of Solidago canadensis L. and S. altissima L. for Their Naturalization. Plants 2022, 11, 3235. [Google Scholar] [CrossRef]
- Dong, L.-J.; Yu, H.-W.; He, W.-M. What Determines Positive, Neutral and Negative Impacts of Solidago canadensis Invasion on Native Plant Species Richness? Sci. Rep. 2015, 5, 16804. [Google Scholar] [CrossRef] [Green Version]
- Awaydul, A.; Zhu, W.; Yuan, Y.; Xiao, J.; Hu, H.; Chen, X.; Koide, R.T.; Cheng, L. Common Mycorrhizal Networks Influence the Distribution of Mineral Nutrients between an Invasive Plant, Solidago canadensis, and a Native Plant, Kummerowa striata. Mycorrhiza 2019, 29, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Langeroudi, A.R. To What Extent Arbuscular Mycorrhiza Can Protect Chicory (Cichorium intybus L.) against Drought Stress. Sci. Hortic. 2019, 263, 109109. [Google Scholar] [CrossRef]
- Řezáčová, V.; Rezác, M.; Gryndlerová, H.; Wilson, G.; Michalová, T. Arbuscular Mycorrhizal Fungi Favor Invasive Echinops sphaerocephalus When Grown in Competition with Native Inula Conyzae. Sci. Rep. 2020, 10, 20287. [Google Scholar] [CrossRef] [PubMed]
- Dong, L.-J.; Ma, L.-N.; He, W.-M. Arbuscular Mycorrhizal Fungi Help Explain Invasion Success of Solidago canadensis. Appl. Soil Ecol. 2021, 157, 103763. [Google Scholar] [CrossRef]
- Stahlhut, K.; Dowell, J.; Temme, A.; Burke, J.; Goolsby, E.; Mason, C. Genetic Control of Arbuscular Mycorrhizal Colonization by Rhizophagus intraradices in Helianthus annuus (L.). Mycorrhiza 2021, 31, 723–734. [Google Scholar] [CrossRef] [PubMed]
- Mandel, J.R.; Dikow, R.B.; Siniscalchi, C.M.; Thapa, R.; Watson, L.E.; Funk, V.A. A Fully Resolved Backbone Phylogeny Reveals Numerous Dispersals and Explosive Diversifications throughout the History of Asteraceae. Proc. Natl. Acad. Sci. USA 2019, 116, 14083–14088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaur, S.; Campbell, B.J.; Suseela, V. Root Metabolome of Plant–Arbuscular Mycorrhizal Symbiosis Mirrors the Mutualistic or Parasitic Mycorrhizal Phenotype. New Phytol. 2022, 234, 672–687. [Google Scholar] [CrossRef]
- Kaur, S.; Suseela, V. Unraveling Arbuscular Mycorrhiza-Induced Changes in Plant Primary and Secondary Metabolome. Metabolites 2020, 10, 335. [Google Scholar] [CrossRef]
- Malygin, D.; Mandryk-Litvinkovich, M.; Sokornova, S. Does Arbuscular Mycorrhiza Favor Invasion of Some Asteraceae Tribes? Plant Prot. News 2021, 104, 144–152. [Google Scholar] [CrossRef]
- Davison, J.; Moora, M.; Öpik, M.; Adholeya, A.; Ainsaar, L.; Bâ, A.; Burla, S.; Diedhiou, A.G.; Hiiesalu, I.; Jairus, T.; et al. Global Assessment of Arbuscular Mycorrhizal Fungus Diversity Reveals Very Low Endemism. Science 2015, 349, 970–973. [Google Scholar] [CrossRef] [Green Version]
- Wu, B.; Hogetsu, T.; Isobe, K.; Ishii, R. Community Structure of Arbuscular Mycorrhizal Fungi in a Primary Successional Volcanic Desert on the Southeast Slope of Mount Fuji. Mycorrhiza 2007, 17, 495–506. [Google Scholar] [CrossRef] [PubMed]
- Shah, M.; Callaway, R.; Shah, T.; Houseman, G.; Pál, R.; Xiao, S.; Luo, W.; Rosche, C.; Reshi, Z.; Khasa, D.; et al. Conyza canadensis Suppresses Plant Diversity in Its Nonnative Ranges but Not at Home: A Transcontinental Comparison. New Phytol. 2014, 202, 1286–1296. [Google Scholar] [CrossRef] [PubMed]
- Shah, M.; Beaulieu, M.-E.; Reshi, Z.; Qureshi, S.; Khasa, D. A Cross-City Molecular Biogeographic Investigation of Arbuscular Mycorrhizas in Conyza canadensis Rhizosphere across Native and Non-Native Regions. Ecol. Process. 2015, 4, 7. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Gao, C.; Guo, L.-D. Changes in Arbuscular Mycorrhizal Fungus Community along an Exotic Plant Eupatorium adenophorum Invasion in a Chinese Secondary Forest. J. Microbiol. 2013, 51, 295–300. [Google Scholar] [CrossRef]
- Shen, K.; Cornelissen, J.H.C.; Wang, Y.; Wu, C.; He, Y.; Ou, J.; Tan, Q.; Xia, T.; Kang, L.; Guo, Y.; et al. AM Fungi Alleviate Phosphorus Limitation and Enhance Nutrient Competitiveness of Invasive Plants via Mycorrhizal Networks in Karst Areas. Front. Ecol. Evol. 2020, 8, 125. [Google Scholar] [CrossRef]
- Tang, J.S.; Zhao, Z.L.; Ma, M. Glomus mosseae Promotes Xanthium italicum Invasion. Sains Malays. 2020, 49, 2425–2432. [Google Scholar] [CrossRef]
- Martín, J.; Morcillo, R.; Vierheilig, H.; Ocampo, J.; Ludwig-Müller, J.; Garrido, J. Ethylene-dependent/Ethylene-independent ABA Regulation of Tomato Plants Colonized by Arbuscular Mycorrhiza Fungi. New Phytol. 2011, 190, 193–205. [Google Scholar] [CrossRef]
- Bitterlich, M.; Krügel, U.; Boldt-Burisch, K.; Franken, P.; Kühn, C. The Sucrose Transporter SlSUT2 from Tomato Interacts with Brassinosteroid Functioning and Affects Arbuscular Mycorrhiza Formation. Plant J. 2014, 78, 877–889. [Google Scholar] [CrossRef]
- Etemadi, M.; Gutjahr, C.; Couzigou, J.-M.; Zouine, M.; Audran, C.; Timmers, A.; Bouzayen, M.; Guillaume, B.; Combier, J.-P. Auxin Perception Is Required for Arbuscule Development in Arbuscular Mycorrhizal Symbiosis. Plant Physiol. 2014, 166, 281–292. [Google Scholar] [CrossRef] [Green Version]
- Dehua, L.; Wang, S.; Cui, M.; Liu, J.; Chen, A.; Xu, G. Phytohormones Regulate the Development of Arbuscular Mycorrhizal Symbiosis. Int. J. Mol. Sci. 2018, 19, 3146. [Google Scholar] [CrossRef] [Green Version]
- Wu, F.; Gao, Y.; Yang, W.; Sui, N.; Zhu, J. Biological Functions of Strigolactones and Their Crosstalk With Other Phytohormones. Front. Plant Sci. 2022, 13. [Google Scholar] [CrossRef] [PubMed]
- Koricheva, J.; Gange, A.C.; Jones, T. Effects of Mycorrhizal Fungi on Insect Herbivores: A Meta-Analysis. Ecology 2009, 90, 2088–2097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, D.; Tan, M.; Wu, S.; Zheng, L.; Wang, Q.; Wang, G.; Yan, S. Defense Responses of Arbuscular Mycorrhizal Fungus-Colonized Poplar Seedlings against Gypsy Moth Larvae: A Multiomics Study. Hortic. Res. 2021, 8, 245. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Jiachao, Z.; George, T.; Limpens, E.; Feng, G. Arbuscular Mycorrhizal Fungi Conducting the Hyphosphere Bacterial Orchestra. Trends Plant Sci. 2021, 27, 402–411. [Google Scholar] [CrossRef]
- Gahan, J.; Schmalenberger, A. Arbuscular Mycorrhizal Hyphae in Grassland Select for a Diverse and Abundant Hyphospheric Bacterial Community Involved in Sulfonate Desulfurization. Appl. Soil Ecol. 2015, 89, 113–121. [Google Scholar] [CrossRef] [Green Version]
- Long, L.; Zhu, H.; Yao, Q.; Ai, Y. Analysis of Bacterial Communities Associated with Spores of Gigaspora margarita and Gigaspora rosea. Plant Soil 2008, 310, 1–9. [Google Scholar] [CrossRef]
- Agnolucci, M.; Battini, F.; Cristani, C.; Giovannetti, M. Diverse Bacterial Communities Are Recruited on Spores of Different Arbuscular Mycorrhizal Fungal Isolates. Biol. Fertil. Soils 2015, 51, 379–389. [Google Scholar] [CrossRef]
- Frey-Klett, P.; Garbaye, J.; Tarkka, M. The Mycorrhiza Helper Bacteria Revisited. New Phytol. 2007, 176, 22–36. [Google Scholar] [CrossRef]
- Pivato, B.; Offre, P.; Marchelli, S.; Barbonaglia, B.; Mougel, C.; Lemanceau, P.; Berta, G. Bacterial Effects on Arbuscular Mycorrhizal Fungi and Mycorrhiza Development as Influenced by the Bacteria, Fungi, and Host Plant. Mycorrhiza 2008, 19, 81–90. [Google Scholar] [CrossRef]
- Battini, F.; Grønlund, M.; Agnolucci, M.; Giovannetti, M.; Jakobsen, I. Facilitation of Phosphorus Uptake in Maize Plants by Mycorrhizosphere Bacteria. Sci. Rep. 2017, 7, 4686. [Google Scholar] [CrossRef] [Green Version]
- Gopal, S.; Shagol, C.; Kim, K.; Han, S.; Sa, T. Spore Associated Bacteria Regulates Maize Root K+/Na+ Ion Homeostasis to Promote Salinity Tolerance during Arbuscular Mycorrhizal Symbiosis. BMC Plant Biol. 2018, 18, 109. [Google Scholar] [CrossRef] [Green Version]
- Turrini, A.; Avio, L.; Giovannetti, M.; Agnolucci, M. Functional Complementarity of Arbuscular Mycorrhizal Fungi and Associated Microbiota: The Challenge of Translational Research. Front. Plant Sci. 2018, 9, 1407. [Google Scholar] [CrossRef]
- Diagne, N.; Ngom, M.; Djighaly, P.I.; Fall, D.; Hocher, V.; Svistoonoff, S. Roles of Arbuscular Mycorrhizal Fungi on Plant Growth and Performance: Importance in Biotic and Abiotic Stressed Regulation. Diversity 2020, 12, 370. [Google Scholar] [CrossRef]
- Ujvári, G.; Turrini, A.; Avio, L.; Agnolucci, M. Possible Role of Arbuscular Mycorrhizal Fungi and Associated Bacteria in the Recruitment of Endophytic Bacterial Communities by Plant Roots. Mycorrhiza 2021, 31, 527–544. [Google Scholar] [CrossRef] [PubMed]
- Tisserant, E.; Malbreil, M.; Kuo, A.; Kohler, A.; Symeonidi, A.; Balestrini, R.; Charron, P.; Duensing, N.; Frei Dit Frey, N.; Gianinazzi-Pearson, V.; et al. Genome of an Arbuscular Mycorrhizal Fungus Provides Insight into the Oldest Plant Symbiosis. Proc. Natl. Acad. Sci. USA 2013, 111, 20117–20122. [Google Scholar] [CrossRef] [Green Version]
- Iffis, B.; St-Arnaud, M.; Hijri, M. Bacteria Associated with Arbuscular Mycorrhizal Fungi within Roots of Plants Growing in a Soil Highly Contaminated with Aliphatic and Aromatic Petroleum Hydrocarbons. FEMS Microbiol. Lett. 2014, 358, 44–54. [Google Scholar] [CrossRef] [Green Version]
- Khaekhum, S.; Ekprasert, J.; Suebrasri, T.; Seemakram, W.; Mongkolthanaruk, W.; Riddech, N.; Jogloy, S.; Boonlue, S. Co-Inoculation of an Endophytic and Arbuscular Mycorrhizal Fungus Improve Growth and Yield of Helianthus tuberosus L. under Field Condition. J. Fungi 2021, 7, 976. [Google Scholar] [CrossRef]
- Svenningsen, N.; Watts-Williams, S.; Joner, E.; Battini, F.; Efthymiou (Katerina), A.; Cruz-Paredes, C.; Nybroe, O.; Jakobsen, I. Suppression of the Activity of Arbuscular Mycorrhizal Fungi by the Soil Microbiota. ISME J. 2018, 12, 1296–1307. [Google Scholar] [CrossRef] [Green Version]
- Martinez, A.; Obertello, M.; Pardo, A.; Ocampo, J.A.; Godeas, A. Interactions between Trichoderma pseudokoningii Strains and the Arbuscular Mycorrhizal Fungi Glomus mosseae and Gigaspora rosea. Mycorrhiza 2004, 14, 79–84. [Google Scholar] [CrossRef]
- Gamalero, E.; Berta, G.; Massa, N.; Glick, B.R.; Lingua, G. Interactions between Pseudomonas putida UW4 and Gigaspora rosea BEG9 and Their Consequences for the Growth of Cucumber under Salt-Stress Conditions: Salt Affects Bacteria–AM Fungi Interactions. J. Appl. Microbiol. 2010, 108, 236–245. [Google Scholar] [CrossRef]
- Lewandowski, T.; Dunfield, K.; Antunes, P. Isolate Identity Determines Plant Tolerance to Pathogen Attack in Assembled Mycorrhizal Communities. PLoS ONE 2013, 8, e61329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eschen, R.; Hunt, S.; Mykura, C.; Gange, A.; Sutton, B. The Foliar Endophytic Fungal Community Composition in Cirsium arvense Is Affected by Mycorrhizal Colonization and Soil Nutrient Content. Fungal Biol. 2010, 114, 991–998. [Google Scholar] [CrossRef] [PubMed]
- Hanlon, M.T.; Coenen, C. Genetic Evidence for Auxin Involvement in Arbuscular Mycorrhiza Initiation. New Phytol. 2011, 189, 701–709. [Google Scholar] [CrossRef]
- Cabral, C.; Wollenweber, B.; Antonio, C.; Ravnskov, S. Activity in the Arbuscular Mycorrhizal Hyphosphere Warning Neighbouring Plants. Front. Plant Sci. 2019, 10, 511. [Google Scholar] [CrossRef]
- Selosse, M.; Richard, F.; Simard, S. Mycorrhizal Networks: Des Liaisons Dangereuses? Trends Ecol. Evol. 2006, 21, 621–628. [Google Scholar] [CrossRef]
- Horton, T.; van der Heijden, M.G.A. The Role of Symbioses in Seedling Establishment and Survival. In Seedling Ecology and Evolution; Leck, M.A., Parker, V.T., Simpson, R.L., Eds.; Cambridge University Press: Cambridge, UK, 2008; pp. 189–213. [Google Scholar]
- van der Heijden, M.G.A.; Horton, T. Socialism in Soil? The Importance of Mycorrhizal Fungal Networks for Facilitation in Natural Ecosystems. J. Ecol. 2009, 97, 1139–1150. [Google Scholar] [CrossRef]
- Mycorrhizal Networks, 1st ed.; Horton, T.R. Springer: Dordrecht, The Netherlands, 2015. [Google Scholar]
- Barto, E.; Antunes, P.; Stinson, K.; Koch, A.; Klironomos, J.; Cipollini, D. Differences in Arbuscular Mycorrhizal Fungal Communities Associated with Sugar Maple Seedlings in and Outside of Invaded Garlic Mustard Forest Patches. Biol. Invasions 2011, 13, 2755–2762. [Google Scholar] [CrossRef]
- Walder, F.; Niemann, H.; Mathimaran, N.; Lehmann, M.; Boller, T.; Wiemken, A. Mycorrhizal Networks: Common Goods of Plants Shared under Unequal Terms of Trade. Plant Physiol. 2012, 159, 789–797. [Google Scholar] [CrossRef] [Green Version]
- Babikova, Z.; Gilbert, L.; Bruce, T.; Birkett, M.; Caulfield, J.; Woodcock, C.; Pickett, J.; Johnson, D. Underground Signals Carried through Neighbouring Plants of Aphid Attack. Ecol. Lett. 2013, 16, 835–843. [Google Scholar] [CrossRef]
- Merrild, M.; Ambus, P.; Rosendahl, S.; Jakobsen, I. Common Arbuscular Mycorrhizal Networks Amplify Competition for Phosphorus between Seedlings and Established Plants. New Phytol. 2013, 200, 229–240. [Google Scholar] [CrossRef]
- Weremijewicz, J.; Janos, D. Common Mycorrhizal Networks Amplify Size Inequality in Andropogon gerardii Monocultures. New Phytol. 2013, 198, 203–213. [Google Scholar] [CrossRef] [PubMed]
- Fellbaum, C.; Mensah, J.; Cloos, A.; Strahan, G.; Kiers, T.; Bücking, H. Fungal Nutrient Allocation in Common Mycelia Networks Is Regulated by the Carbon Source Strength of Individual Host Plants. New Phytol. 2014, 203, 646–656. [Google Scholar] [CrossRef] [PubMed]
- Johnson, D.; Gilbert, L. Interplant Signalling through Hyphal Networks. New Phytol. 2014, 205, 1448–1453. [Google Scholar] [CrossRef] [PubMed]
- Jakobsen, I.; Hammer, E. Nutrient Dynamics in Arbuscular Mycorrhizal Networks. Mycorrhizal Netw. 2015, 20, 91–131. [Google Scholar] [CrossRef]
- Walder, F.; van der Heijden, M.G.A. Regulation of Resource Exchange in the Arbuscular Mycorrhizal Symbiosis. Nat. Plants 2015, 1, 15159. [Google Scholar] [CrossRef]
- Weremijewicz, J.; Sternberg, L.; Janos, D. Common Mycorrhizal Networks Amplify Competition by Preferential Mineral Nutrient Allocation to Large Host Plants. New Phytol. 2016, 212, 461–471. [Google Scholar] [CrossRef] [Green Version]
- Weremijewicz, J.; Sternberg, L.; Janos, D. Arbuscular Common Mycorrhizal Networks Mediate Intra- and Interspecific Interactions of Two Prairie Grasses. Mycorrhiza 2018, 28, 71–83. [Google Scholar] [CrossRef]
- Song, Y.; Wang, M.; Zeng, R.; Groten, K.; Baldwin, I.T. Priming and Filtering of Antiherbivore Defences among Nicotiana attenuata Plants Connected by Mycorrhizal Networks. Plant Cell Environ. 2019, 42, 2945–2961. [Google Scholar] [CrossRef]
- Horn, S.; Hempel, S.; Verbruggen, E.; Rillig, M.C.; Caruso, T. Linking the Community Structure of Arbuscular Mycorrhizal Fungi and Plants: A Story of Interdependence? ISME J. 2017, 11, 1400–1411. [Google Scholar] [CrossRef] [Green Version]
- Malbreil, M.; Tisserant, E.; Martin, F.; Roux, C. Genomics of Arbuscular Mycorrhizal Fungi: Out of the Shadows. Adv. Botan. Res. 2014, 70, 259–290. [Google Scholar]
- Delavaux, C.; Ramos, R.; Stürmer, S.; Bever, J. Environmental Identification of Arbuscular Mycorrhizal Fungi Using the LSU RDNA Gene Region: An Expanded Database and Improved Pipeline. Mycorrhiza 2022, 32, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Krüger, M.; Stockinger, H.; Krüger, C.; Schüßler, A. DNA-based Species Level Detection of Glomeromycota: One PCR Primer Set for All Arbuscular Mycorrhizal Fungi. New Phytol. 2009, 183, 212–223. [Google Scholar] [CrossRef] [PubMed]
- Stefani, F.; Bencherif, K.; Sabourin, S.; Lounès-Hadj Sahraoui, A.; Banchini, C.; Séguin, S.; Dalpé, Y. Taxonomic Assignment of Arbuscular Mycorrhizal Fungi in an 18S Metagenomic Dataset: A Case Study with Saltcedar (Tamarix aphylla). Mycorrhiza 2020, 30, 243–255. [Google Scholar] [CrossRef] [PubMed]
- Kolaříková, Z.; Slavíková, R.; Belz, C.; Krüger, M.; Kohout, P. PacBio Sequencing of Glomeromycota RDNA: A Novel Amplicon Covering All Widely Used Ribosomal Barcoding Regions and Its Applicability in Taxonomy and Ecology of Arbuscular Mycorrhizal Fungi. New Phytol. 2021, 231, 490–499. [Google Scholar] [CrossRef] [PubMed]
- Řezáčová, V.; Řezáč, M.; Líblová, Z.; Michalová, T.; Heneberg, P. Stable Colonization of Native Plants and Early Invaders by Arbuscular Mycorrhizal Fungi after Exposure to Recent Invaders from the Asteraceae Family. Invasive Plant Sci. Manag. 2021, 14, 147–155. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Asteraceae Species | Life Cycle | AMF Colonization Level * | Invasive/Native | References |
|---|---|---|---|---|
| Cardueae | ||||
| Carduus acanthoides L. | biennial/perennial | medium | native | [9] |
| Carduus tenuiflorus Curtis | annual/biennial | medium | native | [10] |
| Centaurea scabiosa L. | perennial | medium | introduced | [11] |
| Centaurea stoebe (=Centaurea maculosa) L. | biennial/perennial | medium | invasive | [12] |
| Cichorieae | ||||
| Cichorium intybus L. | perennial | high | native/introduced | [13] |
| Lactuca serriola L. | annual/biennial | medium | native | [14] |
| Sonchus arvensis L. | perennial | low | pres | [15] |
| Taraxacum officinale (L.) Weber ex F.H.Wigg. | perennial | high | pres | [16] |
| Senecioneae | ||||
| Senecio vernalis Waldst. and Kit. | biennial | low | native | [17] |
| Tussilago farfara L. | perennial | medium | native | [18] |
| Anthemideae | ||||
| Achillea millefolium L. | perennial | medium | native | [19] |
| Artemisia vulgaris L. | perennial | medium | native | [18] |
| Matricaria discoidea (=Matricaria matricarioides) DC. | annual | medium | native/introduced | [20] |
| Tanacetum vulgare L. | perennial | high | native | [21] |
| Astereae | ||||
| Aster squamatus (Spreng.) Hieron. | annual | medium | introduced | [14] |
| Baccharis halimifolia L. | perennial | low | native | [22] |
| Erigeron annuus (L.) Desf. | annual | high | invasive | [23] |
| Erigeron bonariensis (=Conyza bonariensis) L. | annual | medium | introduced | [15] |
| Erigeron canadensis (=Conyza canadensis) L. | annual | medium | invasive | [23] |
| Solidago canadensis | perennial | high | invasive | [23] |
| Solidago gigantea | perennial | high | invasive | [24] |
| Solidago nemoralis Aiton | perennial | medium | native | [25] |
| Millerieae | ||||
| Galinsoga quadriradiata (=Galinsoga ciliata) Ruiz and Pav. | annual | medium | invasive | [26] |
| Galinsoga parviflora Cav. | annual | low | invasive | [27] |
| Eupatorieae | ||||
| Ageratina adenophora (Spreng.) R.M.King and H.Rob. | perennial | medium | invasive | [28] |
| Coreopsideae | ||||
| Bidens frondosa | annual | low | native | [29] |
| high | invasive | [23] | ||
| Bidens tripartite L. | annual | medium | introduced | [30] |
| Heliantheae | ||||
| Ambrosia artemisiifolia | annual | medium | invasive | [31] |
| Helianthus annuus L. | annual | high | invasive | [21] |
| Helianthus tuberosus | perennial | medium | introduced | [32] |
| high | invasive | [23] | ||
| Parthenium hysterophorus L. | annual | medium | native | [27] |
| Rudbeckia laciniata L. | perennial | high | invasive | [24] |
| Xanthium albinum (Widder) Scholz and Sukopp | annual | high | invasive | [23] |
| Host-Plant Species | Associated AMF Community | References |
|---|---|---|
| Brown knapweed Centaurea jacea L. | Glomus sp. | [81] |
| Claroideoglomus sp. | ||
| Diversispora sp. ** | ||
| Acaulospora sp. ** | ||
| Archaeospora sp. | ||
| Thistle Cirsium purpuratum (Maxim) Matsuma. | Acaulospora sp. ** | [82] |
| Rhizoglomus sp. | ||
| Rhizophagus sp. | ||
| Diversispora sp. ** | ||
| Canadian fleabane Erigeron canadensis (=Conyza canadensis) | Glomus versiforme | [23,83,84] |
| Funneliformis caledonius | ||
| F. mosseae | ||
| Rhizophagus intraradices | ||
| Septoglomus constrictum | ||
| Claroideoglomus claroideum | ||
| C. etunicatum | ||
| Diversisporaeburnean ** | ||
| Diversispora sp. ** | ||
| Canadian goldenrod Solidago canadensis | Septoglomus constrictum | [23,75] |
| Funneliformis mosseae | ||
| F. geosporus | ||
| Claroideoglomus claroideum | ||
| C. etunicatum | ||
| Oehlia diaphana | ||
| Glomus versiforme | ||
| Acaulospora excavate ** | ||
| A. mellea | ||
| Diversispora sp. | ||
| Shaggy soldier Galinsoga quadriradiata | Glomus spp. | [23] |
| Claroideoglomus claroideum | ||
| Funneliformis mosseae | ||
| Septoglomus constrictum * | ||
| Crofton weed Ageratina adenophora (=Eupatorium adenophorum) | Claroideoglomus etunicatum | [85,86] |
| Funneliformis geosporus | ||
| Rhizophagus aggregatum | ||
| Diversispora arenaria ** | ||
| Annual forb Bidens pilosa | Septoglomus viscosum | [31] |
| Septoglomus constrictum | ||
| Glomus perpusillum | ||
| Devil’s beggarticks Bidens frondosa | Claroideoglomus claroideum | [23] |
| Claroideoglomus drummondii | ||
| Septoglomus constrictum | ||
| Diversispora sp. ** | ||
| Common ragweed Ambrosia artemisiifolia | Funneliformis mosseae | [31,55] |
| Glomus reticulatum * | ||
| Glomus perpusillum | ||
| Septoglomus constrictum * | ||
| Septoglomus viscosum | ||
| Unundentidied spores | ||
| Jerusalem artichoke Helianthus tuberosus | Funneliformis mosseae | [23] |
| Claroideoglomus claroideum | ||
| Septoglomus constrictum | ||
| Diversispora sp. ** | ||
| Cutleaf coneflower Rudbeckia laciniata | Glomus macrocarpum | [24] |
| Acaulospora cavernata** | ||
| Claroideoglomus claroideum | ||
| Funneliformis mosseae | ||
| Pacispora franciscana ** | ||
| Rhizoglomus fasciculatum | ||
| Scutellospora dipurpurescens ** | ||
| Septoglomus constrictum | ||
| Cocklebur Xanthium albinum | Funneliformis mosseae | [23,87] |
| Claroideoglomus claroideum | ||
| Septoglomus constrictum | ||
| Rhizoglomus microaggregatum |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sokornova, S.; Malygin, D.; Terentev, A.; Dolzhenko, V. Arbuscular Mycorrhiza Symbiosis as a Factor of Asteraceae Species Invasion. Agronomy 2022, 12, 3214. https://doi.org/10.3390/agronomy12123214
Sokornova S, Malygin D, Terentev A, Dolzhenko V. Arbuscular Mycorrhiza Symbiosis as a Factor of Asteraceae Species Invasion. Agronomy. 2022; 12(12):3214. https://doi.org/10.3390/agronomy12123214
Chicago/Turabian StyleSokornova, Sonya, Daniil Malygin, Anton Terentev, and Viktor Dolzhenko. 2022. "Arbuscular Mycorrhiza Symbiosis as a Factor of Asteraceae Species Invasion" Agronomy 12, no. 12: 3214. https://doi.org/10.3390/agronomy12123214