
The Epidemiology and Control of “Olive Quick Decline Syndrome” in Salento (Apulia, Italy)

Council for Agronomical Research and Economics (CREA)—Research Centre for Olive, Fruit and Citrus Crops, Via di Fioranello, 52, I-00134 Rome, Italy
Agronomy 2022, 12(10), 2475; https://doi.org/10.3390/agronomy12102475
Submission received: 8 September 2022
/
Revised: 30 September 2022
/
Accepted: 10 October 2022
/
Published: 11 October 2022
(This article belongs to the Special Issue Epidemiology of Plant Pathogenic Bacteria)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Xylella fastidiosa subsp. pauca sequence type 53 was found to be associated with olive trees showing extensive twig and branch dieback and plant death in the Gallipoli area of Salento and the common name of “olive quick decline syndrome” (OQDS) was given to the disease. Repeated interceptions in Europe and Italy of ornamental coffee plants originated from Central America, jointly in phylogenetic analyses of many strains of the pathogen, indicated this origin as the most probable source of its introduction in Salento. Drought events could have been conducive to the initial outbreaks of the disease. Subsequently, the pathogen largely spread over the territory according to a “stratified dispersal” model. The high susceptibility of the local cultivars Ogliarola salentina and Cellina di Nardò, a low soil content of zinc, copper, and manganese, improper pruning, and adverse climatic events could have further contributed to the spread of the pathogen. The polyphagous insect Philaenus spumarius L. is the main vector of the bacterium in the area. The adults were detected X. f. subsp. pauca-positive in early May, and their incidence was higher during spring and early autumn when they efficiently spread the bacterium among the olive trees. Many other host plant species can host the bacterium, and some of them can act as a “reservoir” for the disease spread. The aggressive fungus Neofusicoccum mediterraneum Crous, M.J. Wingf. And A.J.L. Philips, could also be involved in OQDS. A sustainable control strategy for reducing the incidence and severity of X. f. subsp. pauca in the olive groves of Salento that allows the trees to produce is presented and discussed. Resilient trees of Ogliarola salentina and Cellina di Nardò have been observed in the heavily infected areas of Salento.
1. Introduction
An olive (Olea europea L.) decline was observed in some areas of Salento (the province of Lecce, Apulia, Italy) during the end of the first decade of the new millennium [1]. Afterward, the decline spread northward and reached the provinces of Taranto and Brindisi and some municipalities of the province of Bari, causing extensive tree dieback and death. Initially, some phytopathogenic fungi, belonging to the genera Phaeoacremonium, Phoemoniella, Pleumostomophora, and Neofusicoccum, and insects, namely Zeuzera pyrina L., were retained among the causes of the decline that showed aspecific symptoms (i.e., leaf, twig, and branch dieback) [1]. In October 2013, Xylella fastidiosa Wells et al. was, for the first time, found to be associated with the declining olive trees [2] and, subsequently, isolated and characterized as subspecies pauca, also from symptomatic oleander (Nerium oleander L.) plants [3]. The discovery of this quarantine bacterium, never officially reported and validated before in Europe, prompted a series of studies aiming at elucidating some basic features of the decline, which was named “olive quick decline syndrome” (OQDS). Progress has been achieved in many areas of research, such as the molecular characterization and detection of the pathogen, the monitoring of disease spread and additional host plants, insect vector(s) identification and biology, and olive cultivar susceptibility and resistance [4]. In this article, the current knowledge about OQDS obtained so far is framed in an epidemiological context. We seek to tentatively provide a base for additional studies aimed at elucidating other aspects of the disease not yet completely clarified. In addition, a sustainable field control strategy, developed during recent years, aimed at reducing the incidence and severity of X. f. subsp. pauca in the olive groves of Salento is described and discussed.
2. Xylella fastidiosa: An Ancient Microbe of the Earth
X. fastidiosa is a strictly aerobic, non-motile bacterium that can survive and proliferate within the xylematic tissue of 560 plant species of 80 plant families [5]. It is displaced between the host plants through the xylem-sap feeding activity of some polyphagous insect vectors belonging to “Hemiptera”, suborder “Homoptera”; “Aphrophoridae”, “Cicadellidae”, and “Cercopidae” families [6,7]. Even though it is considered a benign commensal in most of its hosts, in some circumstances, this bacterium colonizes crop and ornamental plant species, causing relevant economic losses in North, Central, and South America as well as in southern Europe [8]. Generally, its main characteristic is the incapability to freely live outside the plant or the insect.
However, there is putative evidence that X. fastidiosa inhabited unexpected ecological niches of the Earth many millions of years before the appearance of plants and insects. With the aim of reconstructing the evolution of Prokaryotes, through a genomic timescale sequencing of different prokaryotic taxa strains, Battistuzzi et al. [9] estimated that X. fastidiosa originated about 600 million years ago, about 500 million years before the most closely related taxon (i.e., Xanthomonas) [9]. Concerning plant evolution, the transition from water to land and the origin of plant vascular tissue date back to around 470 million years ago [10] when the early terrestrial flora starts to diversify by colonizing different ecosystems [11], facilitated by symbiotic fungi [12]. Similarly, the early insects evolved about 479 million years ago, the flying insects about 407 million years ago [13], and the Hemiptera, to which the currently known X. fastidiosa vectors belong, about 160–100 million years ago [14]. If the genomic timescale reconstruction of Prokaryotes is correct, this microorganism apparently survived for more than 100 million years in a completely different environment than the current one and in the absence of land plants and insects. Moreover, its currently known insect vectors appeared after about 440–500 million years from its origin.
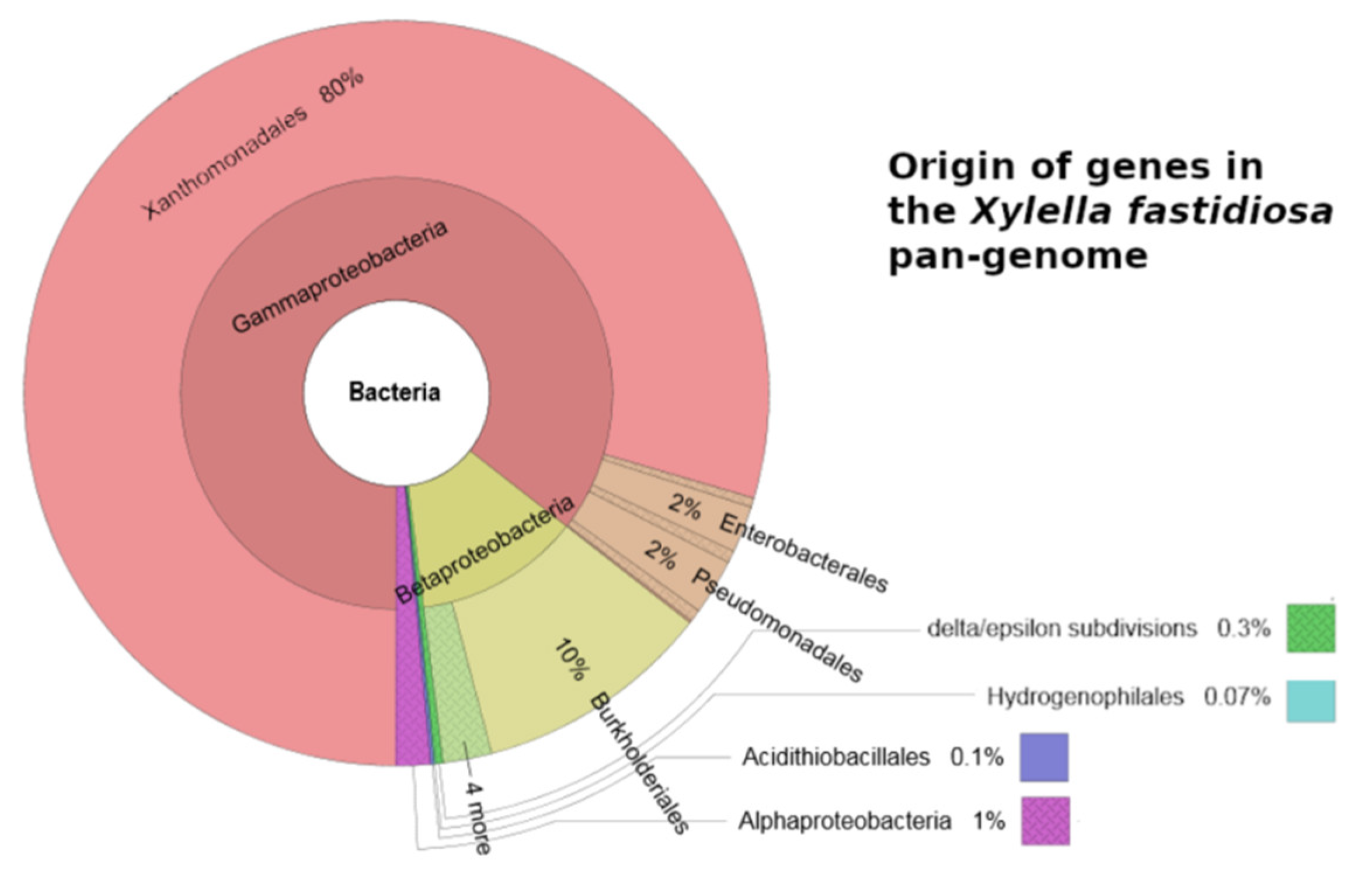
The most primitive plants were the green algae (i.e., photosynthetic eukaryiotic organisms) that originated about 1,500 million years ago [15,16]. For about 1000 million years these unicellular or pluricellular algae lived in marine environments as the sole representatives of the Plant Kingdom. It is known that green algae and bacteria live together in complex microbial communities by performing physiological activities that would not be possible in the absence of such partnerships [17]. Whether X. fastidiosa was able to establish relationships with such primitive plants in the absence of the vascular plants or insect vectors is not known. However, it should be said that this bacterium, through horizontal gene transfer, has obtained a relevant portion of its genome (i.e., about 20%) from distantly related bacteria [18] (Figure 1). Generally, this evolutionary “old age” could explain the recognized versatility of this microorganism in establishing relationships with so many plant species, together with its ability to rapidly colonize the xylem of the host plants with few cells without blocking the flow of the xylem sap [19] and to move against the xylem flow [20].
3. The Introduction in Salento of Xylella fastidiosa subsp. pauca through Coffee Plants
There is evidence that ornamental coffee (Coffea arabica L. and C. canephora Pierre ex Frohen) plants that originated from Central America and were shipped to Europe hosted cells of X. fastidiosa. After 2013, the subspecies fastidiosa was intercepted in France from C. canephora imported from Mexico, whereas the subspecies pauca was found in C. arabica plants from Ecuador [21]. Similarly, X. f. subsp. fastidiosa strains were intercepted both in the Netherlands [22] and in three different localities of northern Italy [23] from the same lot of C. arabica plants introduced from Costa Rica (Figure 2). Indirect but striking evidence for an introduction in Salento (Apulia, Italy) of X. f. subsp. pauca from Central America through ornamental coffee plants has been obtained through genomic sequence comparison. The molecular analyses and comparison of multiple strains of X. f. subsp. pauca isolated in Central and South America from different host plants with the strain isolated in Italy from an olive tree showing symptoms of OQDS revealed a very high similarity between this strain and strains isolated from coffee in Costa Rica, indicating a strict genetic relationship between them [24,25]. Sequence type (ST) 53 was always found to be associated with the olive and oleander decline, allowing for the further characterization of the pathogen [26].
These findings were also supported by additional analyses that found C. arabica plants imported from Costa Rica and Honduras as a reservoir for X. f. subsp. pauca strains [27]. The study also ascertained that the sequence type found in these plants very often belongs to the same ST associated with OQDS in Salento, namely ST53 [27]. The same ST was also found in other plant species such as Prunus dulcis, Prunus avium, Westringia fruticosa, Polygala myrtifolia, and Catharanthus roseus, which have grown close to the infected olive groves [26]. The genome sequencing of three ST53 strains isolated from infected olive trees revealed a very high similarity among these strains and with the related ones in Central America, suggesting that a single introduction of infected plant material took place and pointing to a “founder effect” for the X. f. subsp. pauca population [28].
Moreover, through a population genomic approach, based on the sequencing and comparison of multiple X. f. subsp. pauca strains isolated in Central America and in Salento from olive trees showing symptoms of OQDS, it has been assumed that the introduction of the bacterium, through infected coffee plants from Central America, most likely occurred in 2008 [29]. It should be added that X. f. subsp. pauca, including ST53, has been reported on coffee in Costa Rica during recent years [30,31] and that the export of plants from this country to different continents, including Europe, in recent decades is quite relevant [32], with about 43 million exported ornamental plants in 2012 alone (https://www.focus.it/ambiente/natura/xylella-il-batterio-killer-degli-olivi, accessed on 3 September 2022). Moreover, since 2000, a relevant import from Central America to Europe of potential ornamental host plants for the bacterium has been observed [33]. These data imply that X. f. subsp. pauca ST53 was most likely introduced from Central America, through already infected but not necessarily symptomatic ornamental coffee plants, into some nursery(ies) of Salento, where it subsequently infected the nearby olive trees.
Owing to the relevant import of coffee plants from Central America that occurred during the first decade of the new millennium in Italy and the frequent interception in Europe of X. f. subsp. pauca ST53 in coffee plants originating from that area [27], one corollary facet of this plausible reconstruction on the pathogen introduction in Salento concerns the absence of OQDS in other areas of Apulia, in nearby Mediterranean regions such as Basilicata, Calabria, and Sicily, and in other regions of Central Italy where olive is also cultivated. Considering that insect vectors are present in these areas [34,35,36], this situation could be due to the absence of the pathogen in the imported coffee plants or to intrinsic factors that characterize the olive groves of Salento, such as cultivar susceptibility and/or other predisposing factors [37], which will be taken into account in an epidemiological context.
4. Early Records of Initial Outbreaks of “Olive Quick Decline Syndrome” in Salento
The date of 2008, however, does not establish whether the pathogen was introduced in Salento in that year, whether it became adapted to the new host plant in that year, or whether the date represent the initial outbreak of OQDS [29]. Similarly, other studies, based on the records of monitoring data of the phytosanitary service of the Apulia region, together with logistic functions coupled with the best fitting models, have estimated that the initial OQDS spread occurred in 2008 with a margin of error of plus or minus five years [38]. Another analysis, based on the measurement of the land surface temperature index revealed that this index started to increase from 2006 to 2010, revealing a relevant increase in olive grove mortality during those years [39]. Within this scenario, there is other evidence, based on farmer observations, of symptoms of olive decline during the years 2004–2006 [40]. To these reconstructions, it should also be added that the first symptoms of OQDS (i.e., twig and branch dieback) can be visibly scored after 1–2 years from the initial inoculation of the pathogen within the leaf by the vector [41]. Altogether, these data could bring back the early phase of X. f. subsp. pauca ST53 olive colonization in Salento during 2002–2003.
In any case, at the time of the first official record in October 2013, a well-demarcated area, which involved some municipalities around Gallipoli, was identified as the place of the initial outbreak of OQDS in Salento [1]. However, the place where the infection started is not known. One striking piece of evidence of the OQDS outbreak is that, at the time of the official record, the disease was already present on about 8000–10,000 ha, which corresponds to about 1 million olive trees [1], which precluded any attempt of pathogen eradication in the area. One year after the first report, OQDS was reported on about 23,000 ha [42].
One prevailing aspect of the initial spread of OQDS was that, apart from the Gallipoli area where the disease had the time to expand between the continuum of olive groves due to the feeding activity of the insect vectors repeated over many years, additional initial foci of the disease were observed a few months after October 2013, several kilometers away and across from each other, as in the cases of Trepuzzi, Monteroni, Galatina, and Copertino (Lecce province) along with Oria and San Pietro Vernotico (Brindisi province). Owing to the possibility that infected vectors “hitchhiked” through mobile vehicles (i.e., cars, motorcycles, bikes, tractors, trucks, buses, trains), the long-distance dispersal events of OQDS have indeed remained highly probable and account for a “stratified dispersal” model of the pathogen in Salento that implies a relevant role for the non-olive host plants for the bacterium spreading [43]. Within this scenario, it seems useful to quote an experience that occurred during a monitoring survey carried out in the Campania region (Southern Italy) for ascertaining the possible presence of X. fastidiosa in that area. During the survey, an adult specimen of Philaenus spumarius, the main insect vector of X. f. subsp. pauca in Salento, adhered to a moving car for more than 40 km, remaining alive after displacement [44]. Moreover, during summer 2022, the insect vector was also found in the wheels of cars that moved from Salento to Bari through the freeway [45]. The introduction of infected ornamental coffee plants in other local nurseries or shopping centers of Salento is an alternative hypothesis.
5. Early Outbreaks and Drought Events
Upon arrival in the Gallipoli area, coffee plant(s) acted as a reservoir of X. f. subsp. pauca ST53. Ad hoc inoculation studies have ascertained that coffee plants are poor sources of this bacterium for the vector P. spumarius in comparison with olive plants [29]. The acquisition and transmission rates for the coffee plants, when used as a source for the vector, were indeed lower than those observed using olive plants [29]. This would mean that very few cells of the bacterium were initially taken by the insect vector from coffee plants and subsequently inoculated to the nearby olive trees, as also indicated by epidemiological studies aimed at assessing the risk potential of this bacterium in different crops [46]. The time that had passed from the arrival of the coffee plants in the nursery(ies) and the subsequent entering of the bacterium in the olive trees is unknown. However, if the reconstruction of the initial colonization and subsequent initial outbreaks described above is correct, it occurred a few years after the pathogen introduction in the area and after extensive tree dieback was observed, considering also the 1- to 2-year period during which visible symptoms and the very low initial inoculum dose of the bacterium in the area were observed. Some additional factors could have triggered a rapid expansion of the disease, drought being one of these.
Drought can be defined as a prolonged lack of groundwater, expressed in terms of recharge and storage in a certain area and over a particular period of time in comparison to normal conditions, and is usually caused by a lack of precipitation [47]. It is known that drought events can be greatly conducive to pathogen infection and tree mortality [48,49,50,51]. Drought per se causes changes and perturbations in fundamental metabolic processes, such as the formation of carbohydrates, proteins, lipids, amino acids, phytohormones level, and secondary metabolism compounds, with the main result being an alteration of resources among plant tissue [50]. A direct consequence of drought is altered plant access to nutrients in the soil and their consequent altered reallocation among plant tissues [52,53], causing a general weakening of the plant defense mechanisms needed to face phytopathogens [54], especially those that colonize the plant vascular system [55,56]. Severe water stress in olive trees caused a relevant reduction in photosynthetic activity [57].
Generally, repeated drought events can induce higher disease susceptibility in the plant through the evolutionary shift of certain genes related to plant defense mechanisms that work against fungal [58] or bacterial pathogens [59]. In particular, some Botryosphaeraceae, such as Neofusicoccum eucaliptorum on Eucalyptus globulus [60] and N. parvum on grapevine [61], are greatly enabled by drought events, and this can cause tree dieback. X. fastidiosa is also favored by water stress conditions in host plants, as observed for Parthenocissus quinquefolia where symptom progression was higher in low-water-infected plants [62]. An interaction that involved a redirection of gene transcription levels between water stress and X. fastidiosa in causing a higher symptom severity of Pierce’s disease in grapevines has also been verified [63]. Moreover, X. f. subsp. pauca possesses a thermostable protein, namely Xf-VapD, involved in the biofilm formation induced by the bacterium within the xylem tissue [64]. It is worth noting that the first outbreak of citrus variegated chlorosis in Brazil, caused by X. f. subsp. pauca, was first observed in an area characterized by a high deficit of water [65].
Within this context, it should be noted that Salento underwent repeated drought events in recent decades. In particular, in the years 2000, 2002, 2007, and 2008, severe droughts were recorded, as indicated by the standard precipitation index (SPI) in the Gallipoli, Nardò, Otranto, and Minervino di Lecce municipalities [66]. Moreover, these droughts had negative cascade events on the groundwater quality, resulting in increased salinity [66]. It is interesting to note that the drought events herein reported coincide with the scenario described above concerning the early records of OQDS, which reported the initial visible outbreaks around 2006 [39,40] or around 2008 [38]. Within this scenario, it has been found that the combined effect of water stress and the infection of X. f. subsp. pauca ST53 induced, on the cultivar Cellina di Nardò, a remarkable decrease in the relative water content of the tree and, consequently, an increase in the susceptibility toward the bacterium [67]. This cultivar, through the activation of specific genes related to drought response, would appear well adapted to the water stress conditions that are not rare in the Mediterranean area; such genes were not activated during the sole pathogen infection [67].
These relationships would suggest that repeated drought events could have played a significant role in accelerating plant colonization and enhancing the aggressiveness of X. f. subsp. pauca ST53 within water-stressed olive trees. Thus, starting from a few infected trees that hosted a low inoculum dose of the pathogen, the disease could have spread in the area over a few years.
6. Further Spread of “Olive Quick Decline Syndrome” in Salento
The wide and continued extension of the olive groves in the Salento areas where the OQDS outbreaks started were greatly conducive to the expansion of the disease, such that, only a few years after the first record, it was defined as an endemic [68]. The high abundance of alternative host plants (i.e., natural flora, oleander, and other ornamental shrubs) greatly augments the potential dispersal of the bacterium in the area [43,69]. However, there are different estimates about the advancement of the disease in the affected areas. Some data obtained from the Apulia regional services indicate an expansion front of about 20 km per year [41]. In another study, through logistic function analyses based on results provided by monitoring survey data, the shape and the rate of the movement of the disease front was assessed [38]. This study indicated that, by starting from Gallipoli, in the northwest direction, the disease had an estimated mean rate of movement of about 10 km per year, and an invasion front width that spans from about 100 to 150 km characterizes the OQDS spread in Salento [38]. By contrast, EFSA, by analyzing the disease dispersal, reported that 90% of the newly infected trees for one year were observed within a mean of 5.2 km of a previously infected area [46]. However, studies on P. spumarius (i.e., the main vector for X. f. subsp. pauca ST53 in Apulia) ascertained that the mean mobility of adults during their peak activity in May and June was about 200 m, with only 2% of the adults moving for about 400 m [70].
Based on the biological activity of the main insect vector in Salento, which indicates an adult dispersal of less than 0,5 km within a single olive grove during their peak activity in May and June, and jointly to its low possibility to be passively transported by wind [70], the wider estimates of the rate of OQDS spread in the infected area—calculated at 5 [46], 10 [38], and 20 km [41] per year—should be explained by additional factors. Such factors would have greatly augmented the velocity of the disease spread starting from the Gallipoli area in a northwest dispersal direction [38]. Among these factors, (a) the feeding activity and the dispersal of the vectors on the weeds and wild plants in autumn could be underestimated, (b) the observed ability to adhere to vehicles shown by the vectors could have highly enhanced the spread of the bacterium in the olive grove continuum, and (c) other phytopathogens (i.e., fungi and bacteria) could have contributed to wilting symptoms, such as those caused by X. f. subsp. pauca ST53.
Currently, the front of infection has reached many areas of the Taranto and Brindisi provinces and some municipalities south of Bari. It should also be noted that, in Salento, there is a relevant mobility of vehicles during the period when the feeding activity of the insect vectors is high (end of spring and early summer). During this time period, due to the commercial, working, and tourist activities, all kinds of motor and electric vehicles run across the heavily infected areas of Salento and reach northern areas, such as Basilicata and north of Bari, where olives are also cultivated to a great extent. The possibility that, during the last 15–20 years, no adult of infected P. spumarius, whose density can reach 40–100 nymphs per square meter of weeds [69], has reached these areas through vehicle “hitchhiking” seems quite low. Another reason for this is that, as stated above, contemporary to the outbreaks observed in the Gallipoli area, other foci of OQDS were observed in other locales of Salento quite far from each other and without a nursery nearby. Some intrinsic characteristics of these nearby still non-infected areas could have impeded the occurrence of OQDS.
7. Factors That Were Conducive to the Spread of “Olive Quick Decline Syndrome”
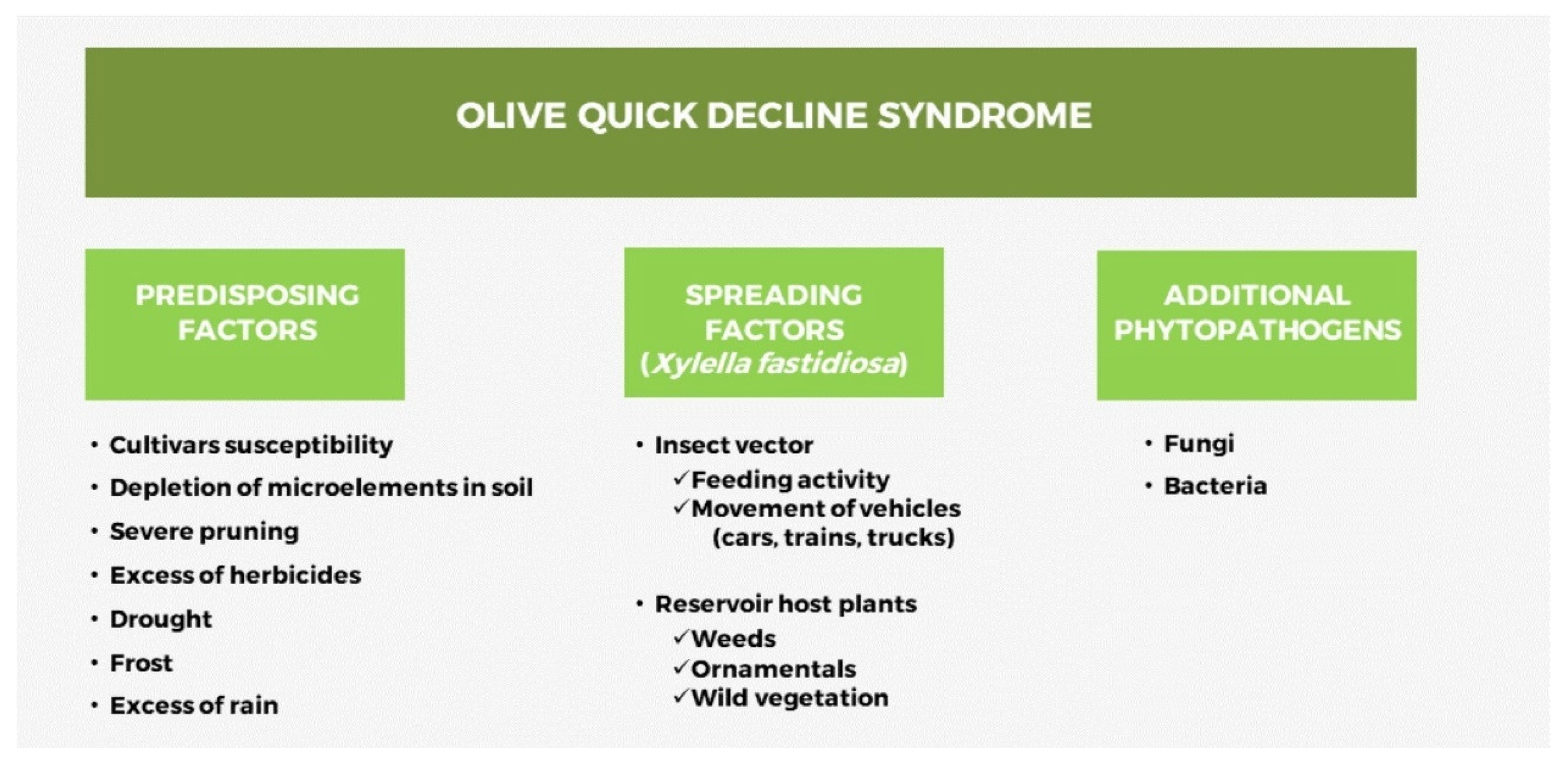
The non-immediate identification of the causal agent of OQDS was certainly among the main causes of the disease spread for many years in the Salento territory. It is evident that, given that about 1 million olive trees were judged as infected at the time of the official record of October 2013 [1], the pathogen had found favorable conditions for expansion since its introduction in the area from abroad. Within this scenario, given the consequent identification of the pathogen vector reported the subsequent year [71], there was a considerable time period during which the bacterium could exponentially spread for many years among the olive groves of Salento, which, in many cases, formed a continuum of trees for many kilometers [68,72]. During this time lapse, some predisposing factors jointly to the bacterium spread through the insect vectors activities and additional phytopathogens that occur in the area could have played a significant role either in augmenting the severity or in the further spread of OQDS in Salento (Figure 3).
The high susceptibility of the local olive cultivars used to produce high-quality oil [73,74], namely Ogliarola salentina and Cellina di Nardò, to X. f. subsp. pauca largely contributed to the spread of OQDS in Salento. These cultivars indeed show, upon infection, more xylem occlusions caused by tylose formation than the less susceptible cultivar Leccino [75]. There is also anatomical evidence that Cellina di Nardò has larger and fewer xylem vessels in comparison with Leccino, resulting in a higher vulnerability to droughts [76]. These large vessels are more prone to air embolism [76], tylose formation [77], and cavitation [78], possibly conducive to X. f. subsp. pauca infection [76] or the consequent twig wilting [77,78]. Moreover, both Ogliarola salentina and Cellina di Nardò have a lower level of hydroxytyrosol glucoside in comparison with Leccino, this phenolic compound having a high antioxidant activity directly involved in the defense mechanism against X. f. subsp. pauca [79]. In addition, when artificially inoculated, the development of visible symptoms occurs in Cellina di Nardò plants before it occurs in Leccino plants [80], and both Ogliarola salentina and Cellina di Nardò host a significantly higher number of X. f. subsp. pauca cells in the twigs than Leccino and other olive cultivars in naturally infected trees [81].
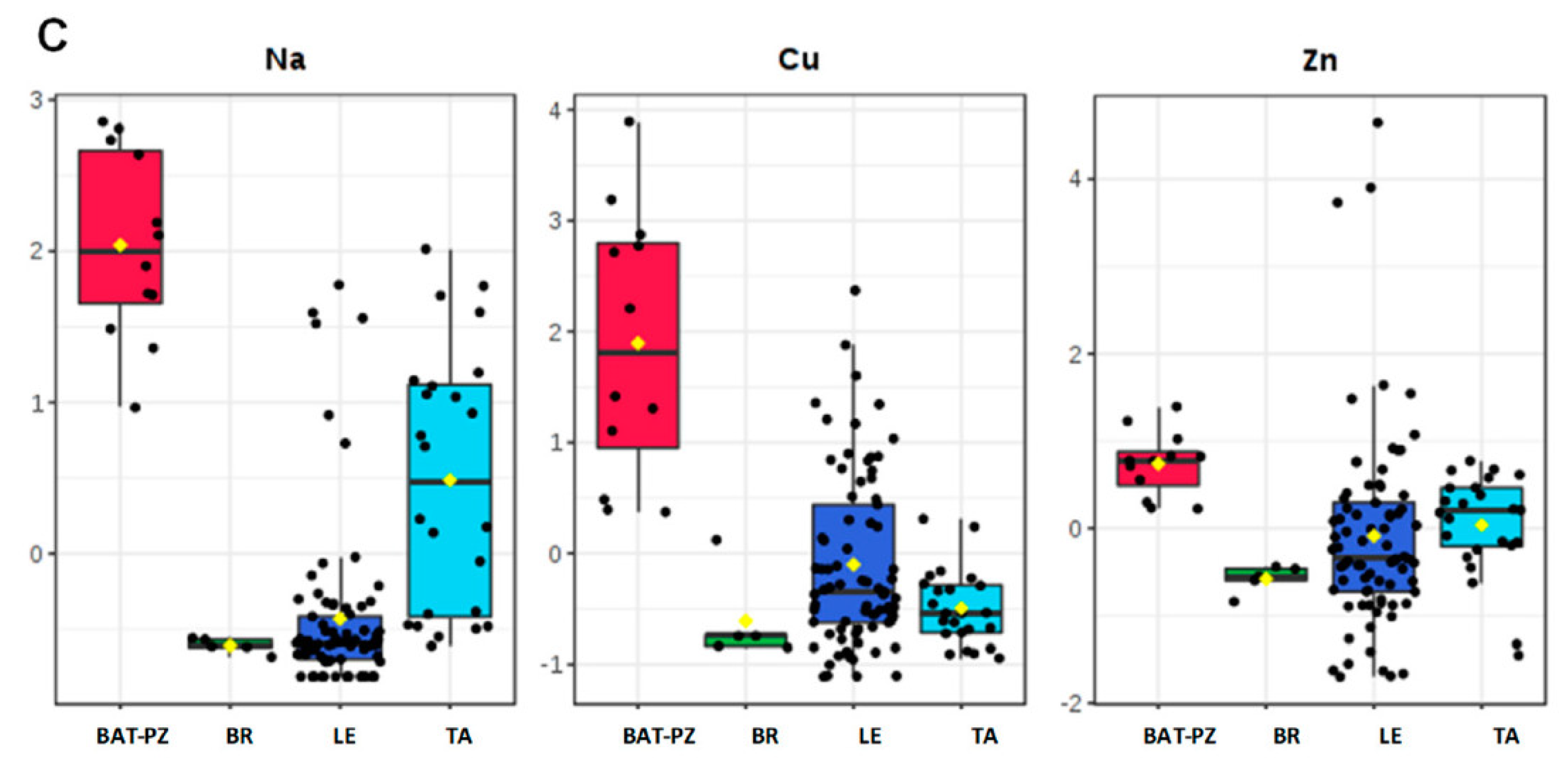
Soil characteristics of the areas where OQDS has spread in recent years could also have been conducive to the disease. A study aimed at assessing the ionome (i.e., elemental composition at ion level) content in the soil and leaves in the olive groves of the infected areas of Salento, in comparison with olive groves of Northern Apulia and Basilicata, where OQDS is not reported, ascertained a very low content of certain micronutrients in all of the sampled olive groves of the infected areas, where a significantly lower content of zinc, copper, and manganese was found in both soil and leaves [82] (Figure 4).
It should be noted that zinc is involved as a cofactor in many enzymes, such as alcohol dehydrogenase, RNA polymerase, and carbonic anhydrase [83], whereas copper is essential for the formation of chlorophyll [84], and manganese is involved in the photosynthetic machinery and in the detoxification of ROS [85]. Taken as a whole, these data point to trees that grow in highly depleted soil, and the leaf ionome composition reflects such a condition. Consequently, the combination of intrinsic cultivar susceptibility with a low amount of micronutrients in the soil that are important for overall plant physiology resulted in an environment that is highly conducive to OQDS spread.
Given the similar geological substrate (i.e., calcareous) of South and North Apulia, some factors could have determined the relevant decrease in micronutrients found in Salento. A survey on the utilization of herbicides used in Apulia from 2003 to 2014 verified that, in areas where OQDS appeared and spread (i.e., Lecce province), the utilization of glyphosate was used twice more than in the northern areas (i.e., Bari province) [86], with an average distribution in the olive groves of three times per year [87]. This relevant distribution of glyphosate that occurred for many years could have altered the chemical and biological equilibrium of the soil. It is known that such a distribution decreases the overall availability of nitrogen and phosphorus along with the organic matter content in the soil [88], and that there is a high reduction in beneficial pseudomonads that control soil-borne pathogenic fungi, with a consequent increase in agrobacteria that play a role in manganese oxidation [89,90,91]. In addition, the prolonged utilization of glyphosate can also induce a reduced assimilation of zinc and copper in the plant [92]. It seems evident that this agronomical practice, prolonged over many years, helped to weaken the olive trees, thus facilitating disease spread. It should be added that a direct link between glyphosate utilization and increases in plant disease severity has already been verified [93].
Additional factors, such as improper pruning and extreme climatic events (i.e., droughts, frost, and “water bombs”, an exceptionally large amount of rain falling in a few hours on a relatively small area), could have contributed to the increase in the tree susceptibility to OQDS. Hard pruning carried out in an attempt to reduce the X. f. subsp. pauca inoculum load within the tree has resulted in its subsequent further weakening and death [80,94]. It should be said indeed that in infected young olive trees, pruning has a limited effect in eliminating the bacterium from the tree [94]. It should also be added that, in Salento, in recent decades, tree pruning was very often performed every 4–5 years, resulting in damage to plant physiology [95]. In addition to the droughts previously reported that could have been conducive to the initial OQDS outbreaks, other drought events have been further recorded, such as that occurring during the summer of 2017, when 7.8 mm of total rainfall was recorded in Lecce province from 1 June to 30 August [95]. In the same year, in January, a prolonged (i.e., one week) frost with minimum temperatures that reached on some days from −4 to −5.1 °C was recorded [95]. Because of climate change, in recent years, the Salento area faced more extreme precipitation events such as the “water bombs” accompanied by strong wind and hail. Apart from the direct damage to vegetation, there are also indirect negative effects on tree physiology due to prolonged water permanence in soil (i.e., waterlogging) (Figure 5). When the soil is saturated with water on consecutive days, anaerobic conditions prevail, thus inducing root hypoxia in the olive trees [96,97]. It seems that an environment conducive to the rapid expansion of OQDS, given the combination of highly susceptible olive cultivars, improper agronomical practices, and repeated adverse climatic events, was present in Salento.
8. The Role of Vectors
One of the most intriguing aspects of OQDS outbreak and spread is the apparent ease with which an insect vector (i.e., P. spumarius) that had never encountered an alien bacterium such as X. fastidiosa in its evolutionary life so rapidly acquired the ability to efficiently spread it in a new host plant for the microbe (i.e., olive), starting its acquisition from a host plant (i.e., coffee) that is a poor source of pathogen acquisition for the insect [29]. By contrast, it is known that X. fastidiosa switches from a plant to an insect-colonized state only when it reaches a high cell density [98] and after a complex multistep process that involves, at the same time, plant, insect, and pathogen components (i.e., pectin, cutin, adhesins, and pili) [99]. Some difficulties have been observed during the probing and feeding of P. spumarius during the transmission of X. fastidiosa [100]. There might also be an additional possibility not usually considered for explaining the introduction of X. f. subsp. pauca from abroad: the direct introduction of an efficient vector from Central America that, upon its arrival in Salento, could have initially transmitted the pathogen to the olive trees and subsequently became extinct due to an inability to adapt to the new environment. Such a possibility was not completely discarded concerning X. fastidiosa outbreaks in the Balearic Islands, in Spain [101]. The live adults of Homalodisca vitripennis (“Hemiptera”: “Cicadellidae”), one of the main vectors of Pierce’s diseases caused by X. f. subsp. fastidiosa in California, was indeed intercepted in French Polynesia and Japan from cargo bins, hangars, and planes [46]. The presumptive fast adaptation of the bacterium to a new vector would require additional studies to determine the mechanisms of their interaction.
The polyphagous P. spumarius (“Hemiptera”: “Aphrophoridae”), also known as the “meadow spittlebug”, is the main vector of X. f. subsp. pauca ST53 in Salento [102,103]. Other less efficient vectors include Philaenus italosignus (“Hemiptera”: “Aphrophoridae”) and Neophilaenus campestris (“Hemiptera”: “Aphrophoridae”) [103,104] (Figure 6). P. italosignus is rarely found in olive groves, and the nymphs exclusively colonize Asphodelus spp., which also serves ovideposition, whereas N. campestris was positive for the bacterium only in May due to its non-preference for olive trees and its preference for Poaceae plant species [105].
In Apulia, the juvenile stages of P. spumarius (i.e., nymphs) for their feeding activity prefer plant species of families such as Asteraceae (Sonchus, Crepis, Picris), Fabaceae (Medicago, Vicia, Lathyrus), Rubiaceae (Galium), and Apiaceae (Daucus, Foeniculum), with G. album and F. vulgare as preferred species. Other species were found to be “non-preferred” or “avoided”, namely Poaceae (Avena, Lolium, Hordeum), Brassicaceae (Raphanus), Papaveraceae (Papaver), Fumariaceae (Fumaria), and Oxalidaceae (Oxalis) [69]. It should be noted that the juvenile stages of the vectors do not transmit the bacterium; rather, it is acquired and spread by the adults when an already infected plant is reached [69]. These findings are confirmed by detection analysis surveys performed on wild and cultivated plant species potentially infected by the bacterium that did not reveal its occurrence in the preferred plants for the nymphs of P. spumarius [4], even though it would seem that the very common weed Chenopodium album as well as Erigeron spp. could host the pathogen through the feeding activity of the adult vectors [69].
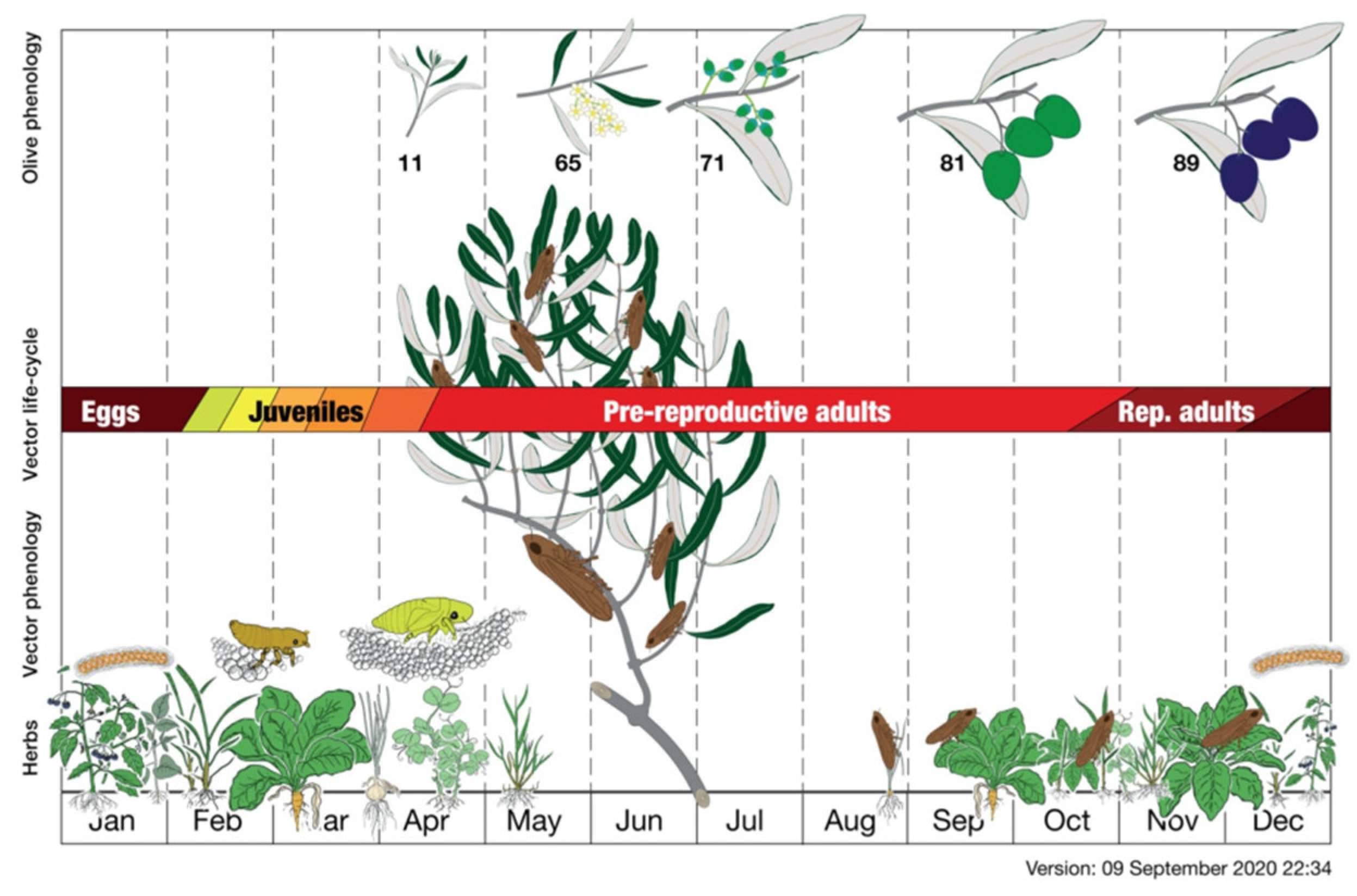
Egg hatching and naiad development occur within the temperature range between 2.8 and 10 °C, and nymphal development is observed at 27 °C [106]. The density of the nymphs in the olive groves is quite high, reaching 10–40 individuals per square meter [105], explaining the high potential of pathogen vector transmission when the adult stage has been reached [107]. In the olive groves, P. spumarius adults occur from late April to late autumn. They were detected as positive for X. f. subsp. pauca in early May, and their incidence was found to be higher throughout the spring season when they efficiently spread the bacterium among the olive trees [104,106]. The peak of occurrence in the olive crown was recorded in May and June, whereas the adults preferred afterward to move toward the surrounding wild shrubs and trees [105,106]. During the period of high abundance, the adult dispersal within the olive grove can range from 200 to 400 m [106], and the colonization of other plant species occurs mainly during the summer. In autumn, adults colonize herbaceous species for ovideposition [106] (Figure 7). It has been estimated that a single infected olive tree can spread the bacterium to 19 other susceptible hosts per year [108].
After artificial inoculation with the vector, the olive tree remains symptomless for one year, but the bacterium starts to colonize large parts of the plant xylem [80], thus potentially infectious during the first visible desiccation period [107,108]. Suckers that are stung by the infected vector become infectious as well [107]. Interestingly, a higher plant infestation rate by juvenile-stage P. spumarius has been found in olive groves of Lecce province when compared with olive groves of Bari province [105]. This implies that X. f. subsp. pauca should be considered an endemic in Southern Apulia. These findings reveal that X. f. subsp. pauca ST53 found an efficient vector for its spread in Salento. As observed above, this efficiency in acquiring and transmitting the bacterium from olive trees to other common plant species jointly with its relevant occurrence in Apulia, and the objective possibility of its passive displacement in the territory through motor vehicles for many years, contrasts with the absence of the pathogen in the northern areas of Apulia and the nearby regions.
9. The Role of “Reservoir” Host Plants
Symptomatic host plants for X. f. subsp. pauca ST53, other than olive plants, have been found in Salento [4]. Some cultivated fruit species, namely Prunus avium (sweet cherry) and Prunus dulcis (almond); some ornamental shrubs or trees, namely Acacia saligna, Grevillea juniperina, Laurus nobilis, Nerium oleander, Polygala myrtifolia, and Westringia fruticose; some typical Mediterranean wild species, such as Cistus criticus, Lavandula angustifolia, Lavandula stoechas, Myrtus communis, Phyllirea latifolia, Rosmarinus officinalis, and Spartium junceum; and other ornamental species, namely Hebe sp. and Pelargonium × fragrans, have been found to be infected. In addition, other plant species were found to be asymptomatic but were found positive upon analyses performed with ad hoc detection techniques, namely Euphorbia terracina, Myoporum insulare, Rhamnus alaternus, Vinca sp., and Westringia glabra [4]. Finally, other X. f. subsp. pauca-positive plant species have been found, but without any indication of a possible presence of symptoms: Asparagus acutifolius, Catharanthus sp., Chenopodium album, Dodonea viscosa, Eremophila maculata, Erigeron sumatrensis, Erigeron bonariensis, and Heliotropium europeum [4]. Wild oleasters were found to be infected as well [26]. ST53 was the sole sequence type found in the infected plants [4]. Other plant species commonly infected by X. f. subsp. pauca in South America and cultivated in Salento, such as Citrus spp., and grapevine were found to be consistently negative upon detection and artificial inoculation [4].
The wild Mediterranean plant species could play a relevant role as a “reservoir” enabling the spread of the pathogen among olive groves in Apulia. Such species are quite common in the areas close to the olive groves. It should also be noted that there is an apparent difference between the plant choice and colonization of juvenile-stage P. spumarius and its subsequent feeding activity as an adult and the linked pathogen transmission. As observed above, among the annual and biannual plant species, only Chenopodium album and Erigeron spp. should be considered as infected by the adults of the meadow spittlebug [105]. According to plant species reported as infected, it seems that the host range for adults is quite ample, posing an additional threat and difficulty for a reliable control of the vectors. Within this context, adults of the three vectors of X. f. subsp. pauca have also been found during the summer in pine (Pinus halepensis) and elm (Ulmus minor) trees located in wild habitats northward of the infected areas of Salento, which shows that natural and semi-natural areas located near olive groves can also potentially act as “reservoirs” for the further spread of the bacterium [109].
It appears that plant species other than olive trees can act as “reservoirs” for the additional spread of X. f. subsp. pauca ST53 in Apulia, and their wide occurrence in natural spaces poses an objective difficulty for the effective management of OQDS. For these plants, control measures planned for olive groves against insect vectors seem ineffective, representing a potential threat for future infections.
Within this scenario, it should be observed that, by following one of the first cases of eradication performed in Apulia at Pietro Vernotico (Brindisi province) in October 2015, after the removal of about 900 olive trees that surrounded eight X. fastidiosa-infected trees, the few remaining olive trees located near the culled ones were found to be infected by the bacterium in the early autumn of 2017, based on real-time PCR (Figure 8). Infected wild plants that had grown near the olive groves and that were not eliminated during the 2015 eradication or further movement of the vector could have reinfected the olive trees.
10. The Roles of Other Phytopathogenic Bacteria and Fungi
During surveys in the areas characterized by the occurrence of OQDS, it is quite common to observe in the olive groves additional specific disease symptoms caused by other phytopathogens. Olive knot, caused by Pseudomonas savastanoi pv. savastanoi, and peacock leaf spot, caused by Ventuaria (Spilocaea) oleaginea, are the most frequent. The bacterium can contribute to the overall decrease in plant defense substances. A reduction in phenolic compounds released from plant cell vacuoles during the induction of cavity formation incited by the pathogen’s pectolytic and hemicellulolytic enzymatic activity has been observed [110]. Such reduction also increases the plant’s susceptibility to the fungus [111]. These findings can be considered as additional factors that increase the susceptibility of olive groves to OQDS.
While the occurrence of the dangerous Verticillium wilt, caused by the soil-borne fungus Verticillium dahliae, and Phytophthora root and crown rot, caused by Phytophthora spp., seem to be excluded in an area of OQDS [1,112], other pathogenic fungi have been reported either in Salento or in Northern Apulia. Fungal genera such as Phaeoacremonium, Phoemoniella, Pleumostomorpha, and Neofusicoccum have been reported as common in Salento in the olive groves affected by OQDS [1,112], whereas Phaeoacremonium aleophilum, Pleurostomophora richardsiae, and Neofusicoccum parvum have been found to be associated with olive decline in Northern Apulia, with P. richardsiae being the most aggressive species [113]. In a subsequent analysis, Phoemoniella chlamydospora and species of Botryospheraceae were frequently found to be associated with olive cultivars that show OQDS symptoms in Salento [79]. However, these fungi are not retained involved in OQDS [1,79,112]. Fungi of the Botryospheraceae have also been found to be associated with an almond (Prunus dulcis) decline on Mallorca (Spain) [114]. Subsequently, the trees were found to be infected by X. fastidiosa [101].
However, in Salento, another aggressive fungal pathogen has recently been found, namely Neofusicoccum mediterraneum of the Botryospheraceae family [115]. Botryospheraceae spp. can rapidly colonize very young olive trees planted in the infected areas of Salento very close to the initial location of the OQDS outbreak [116], confirming the relevant presence of such fungi in the area. It is evident that the occurrence of N. mediterraneum should generally be regarded as an additional threat to the olive groves of Salento and Apulia, since this phytopathogen causes severe olive decline both in California and Spain [117,118,119]. This species is characterized by aggressiveness, as it is able to incite twig dieback in olive plants after a few weeks upon artificial inoculation and survives the mycelial stage at temperatures of 50 °C [115]. It can spread during pruning through infected scissors that allow the pathogen to colonize wounds [117,120]. In addition, visual symptoms can be easily confounded with those caused by X. f. subsp. pauca ST53 [115] (Figure 9), confounding both the aerial and ground monitoring surveys on OQDS. A differential symptomatology can be observed at early stages of infection when a reddish discoloration is visible at the edge of the leaf (Figure 10). The possible spread of fungal pathogens in olive groves would confirm a previous evaluation on a monitoring survey performed from September 2017 to March 2018 by the Agenzia Regionale per le attività Irrigue e Forestali (ARIF) of the Apulia Region on 190,000 ha of containment and buffer areas. Based on these surveys, which collected tree samples for additional laboratory analyses aimed at detecting olive trees infected by X. f. subsp. pauca ST53, it was evident that the bacterium was not found in 3300 samples taken from trees that showed symptoms of OQDS [121]. The possible occurrence of other phytopathogens, including N. mediterraneum, causing tree decline cannot be excluded.
Since the time at which this pathogen was present in Salento is difficult to establish, even though it was found in 2007 on olive drupes [122] and it had colonized olive wood, it could have helped to exacerbate the infection of X. f. subsp. pauca, and/or it might have been the main causal agent of some olive decline, confounded with that caused by the bacterium. Despite the common name OQDS, olive trees take from 2 to 5 years to completely collapse [41], and during this time period, a fungal pathogen can additionally contribute to tree decline.
These findings depict a more complicated scenario for OQDS than the simple and direct relationships between X. f. subsp. pauca ST53 and the olive tree decline. Generally, in declines of woody species, several phytopathogens (i.e., polymicrobial complex) have been found to be associated with the symptoms and with other microbes, each one of these species having a specific role in causing the disease [123,124,125]. These aspects have not been taken into account for OQDS and deserve additional consideration.
11. Cultivar Resistance and Resilience
The susceptibility to OQDS of the local cultivars Ogliarola salentina and Cellina di Nardò prompted the search for a resistant olive germplasm among other cultivars already cultivated in Salento and Apulia for possible utilization in new groves to face the disease. Upon surveys carried out in the entire infected area of Salento, two cultivars were judged as “resistant” to X. f. subsp. pauca ST53, namely the well-known Leccino, which is cultivated in many regions of Italy, and FS-17®, which is used either as clonal rootstock or a productive cultivar [126]. Olive groves planted with these cultivars appear to survive infection of the pathogen, whereas neighboring trees planted with susceptible ones were found to be wilted [126] The lower susceptibility of Leccino to X. f. subsp. pauca was also observed by transcriptomic profiling [127]. Moreover, both cultivars planted in the infected areas showed a lower pathogen occurrence in olive leaves in cell density assessments [126].
The lower susceptibility of Leccino is supposedly related to the amount of lignin found in the xylem vessels, which could reduce pathogen movement by slowing disease progression [128], to the amount of secondary metabolites such as the phenolic glicoside hydroxytyrosol glucoside [79], and to narrower xylem vessels that reduce the occurrence of air-embolism vulnerability [76]. Leccino appears to be less tolerant than the local cultivar Cellina di Nardò to drought, suggesting that its tolerance to the bacterium is mainly linked to lignin, phenolic compounds, and the activity of the antioxidant defense system [67].
However, further field surveys revealed that the cultivars regarded as “resistant” showed symptoms of OQDS, such that the term “tolerant” seems to be more appropriate for both Leccino and FS-17®, indicating “significantly reduced symptoms when compared with Ogliarola salentina and Cellina di Nardò, and a lower number of bacterial cells in infected tissue” [129,130]. A decline in adult Leccino trees indeed is frequently reported in infected areas (Figure 11).
In addition, for new plantations aimed at substituting old olive groves destroyed by OQDS, a failure in rooting during the first year has been observed, probably due to the water demand of young plants that, due to repeated droughts and a lack of irrigation, is not satisfied (Figure 12).
Usually, both Ogliarola salentina and Cellina di Nardò trees, when they start to show symptoms of OQDS, completely collapse over a period of months or years. In many cases, after some time, many suckers grow at the collar level; these suckers can usually grow and survive for a few years without any symptoms, but they show clear symptoms of OQDS later and suddenly wilt after a few weeks. This phenomenon is quite common, especially in areas where the first outbreaks of the diseases were observed. Thus, in the past, some farmers tried to train these suckers to form a new crown, but they failed due to their subsequent decline.
Resilience describes the capacity of a system to maintain its functions after the impact of an exogenous disturbance [131]. A case of resilience was described for Esca-diseased grapevine plants after curettage treatment performed in the autumn when all necrotic and dead tissue from the vine was removed [132]. The cured plants regularly sprouted in the following spring and yielded berries in the autumn, and they showed, in the midterm period, a low severity of the Esca disease [132].
The cases of olive trees that spontaneously recovered from OQDS might be considered cases of such resilience. Some Ogliarola salentina and Cellina di Nardò trees of different ages (i.e., either young or 70–80 years old), grown in areas where the first outbreaks of OQDS were observed (i.e., the Gallipoli area), survive and start to yield fruit after growth for four years without showing any symptoms of OQDS. The same trees were previously found to be completely wilted, and non-external products (i.e., fertilizers, pesticides, and hormones) were sprayed onto the canopy or released onto the soil. Some dead plant parts were pruned. In these cases, the tree started to sprout new twigs from the branches and trunk that, in 3–4 years, re-established the canopy and began to yield (Figure 13). Assessment on the occurrence of X. f. subsp. pauca within the foliage of such trees revealed the presence of the bacterium [133]. The causes that allow the olive trees to recover and to sustain the occurrence of the bacterium in the foliage still need to be investigated. The development of resilient plants is an important goal to achieve in the current global scenario of climate change [134].
12. Control
The field control of plant diseases (i.e., cure or therapy) is intended to reduce the incidence and the severity of pathogens on the crop through the field applications of different preventive and/or curative products or compounds, such as agrochemicals, biological control agents, fertilizers, and plant-growth-promoting microbes, for allowing the plant to vegetate and produce without eliminating the pathogen from the crop and/or from the area of cultivation (https://www.apsnet.org/edcenter/disimpactmngmnt/topc/EpidemiologyTemporal/Pages/ManagementStrategies.aspx, accessed on 3 September 2022) [135]. Eradication (i.e., elimination) entails a reduction to zero incidences of a specific disease in a defined geographic area through continued intervention measures [136]. Having in mind the basic principles of plant disease control, a protocol for reducing the incidence and severity of X. f. subsp. pauca ST53 has been studied and applied in olive groves of Salento [95]. The EFSA evaluated the preliminary study and suggested that the assessment of the protocol effectiveness be enlarged by studying the significant reduction in X. f. subsp. pauca cell density after treatment in a larger number of trees [130]. A further study based on a large number of trees was carried out, and the results confirmed and expanded previous analyses by indicating the efficacy of a zinc–copper–citric acid biocomplex in controlling field symptoms caused by the bacterium and reducing the density of the pathogen within the olive leaves [137]. This control strategy is based on some basic assumptions: (a) reduction in the cell density of the bacterium within the xylem tissue, (b) control of the insect vector P. spumarius, (c) maintenance of soil fertility, and (d) regular pruning.
- (A)
- Reduction in Pathogen Cell Density within the Xylem.
Since zinc and copper were found as the most effective ions in reducing the biofilm capability of X. fastidiosa [138,139], a patented biocomplex containing zinc (4%), copper (2%), and citric acid, namely Dentamet®, was preliminary tested for verifying its in vitro bactericidal activity, systemicity, ability to release zinc and copper ions in the xylem tissue, ability to reduce the cell density of X. f. subsp. pauca within the foliage, and capability to reduce field symptoms, together with assuring absence of its residues in the oil [95]. The study revealed bactericidal activity against X. fastidiosa, a relevant systemicity of the biocomplex when sprayed on the olive canopy, a significant reduction in the pathogen cell density due to the effective release of ions into the xylem, and the absence of residues in the oil. At the end of the study, the olive leaves still contained an elevated amount of zinc and copper. The protocol consists in spraying the olive canopy with an atomizer once per month during early spring and early autumn, at a rate of 3.9 L/ha, when each adult tree receives at least 20 L of the spray solution. The olive trees can also receive traditional treatment for controlling common pest and pathogens (i.e., olive fruit flies, olive knot, and olive peacock spot).
In a further study, the bactericidal activity of the biocomplex was also verified toward the subspecies fastidiosa and multiplex. In addition, in a midterm period (i.e., from three to four years of continuously applying of the control strategy), the efficacy of the protocol to allow coexistence with the bacterium and to restore the productivity of the trees was also evaluated [137]. It was observed that the canopy spray reduced the pathogen concentration to a cell density highly similar to, or even lower than, that found in the tolerant cultivars Leccino and FS-17® (i.e., 104 colony-forming units (CFU)/mL), allowing an average yield of about 3–4 tons/ha per year [137] (Figure 14). Moreover, when the control strategy was applied to Leccino, a pathogen mean cell density of about 102 CFU/mL was found. The amount of copper released after treatment for one year was about 500 g/ha, much less than 4 kg/ha, which is the current limit for copper amount allowed for organic agriculture. Over the years, this control strategy has been successfully applied in some olive groves of Salento in small farms (i.e., 1–2 ha) and in farms with a relevant surface (i.e., 10–500 ha) (Figure 15), and professionalism that helped them win national and international awards for their oil quality. This sustainable strategy falls within the possibility of disease control through the appropriate management of plant nutrition, once the effectiveness of the tested compound in the field is established [140,141].
- (B)
- Control of the Insect Vector Philaenus spumarius
Another fundamental part of the success of X. f. subsp. pauca ST53 field control concerns the ability to apply a satisfactory reduction in egg- and juvenile-stage P. spumarius vectors through tillage and the mechanical removal of weeds grown in olive groves and nearby environments. Since the protocol does not include chemical treatments for the reduction in the adult vector population, care should be taken in eliminating the eggs and the juvenile-stage vectors. The first action should be taken during early to mid-winter to eliminate eggs deposited by adults in ground slits. Light tillage, repeated possibly every month, should be applied in the grove to disrupt these eggs [103]. Subsequently, from early February to early May, spontaneous herbs can be mowed to eliminate juvenile-stage specimens (i.e., naiades and nymphs) [103]. It should be emphasized that the abandoned olive groves host the highest population of vectors when compared with cultivated groves [103]. Consequently, given that a considerable portion of territories previously cultivated with olives have been abandoned, these practices should be considered of paramount importance for managers of olive groves in use today.
- (C)
- Maintenance of Soil Fertility
Soil fertility can largely influence plants when facing disease, so soil management is fundamental to properly apply effective pathogen control strategies [142,143]. Consequently, all agronomical measures that can increase the overall soil content of macroelements, microelements, and beneficial soil microorganisms should be applied to olive groves in conjunction with the control measures discussed above. As previously discussed, we have observed that the soil of Salento where OQDS is prevalent lacks basic microelements such as zinc, copper, and manganese. In addition, in some cases, an overall disequilibrium in the content of macro- and microelements has been revealed; consequently, soil analyses that additionally consider the status of soil microorganisms are recommended so that the optimal amount of such elements can be applied.
- (D)
- Regular Pruning
The pruning of olive trees influences both the tree physiology and the quality of the oil [144]. In adult trees, pruning is also performed to potentially maintain the equilibrium between vegetative and reproductive functions, which can limit the tree’s alternate bearing, which facilitates pest and disease control as well as the harvest [145,146]. In recent decades, in many areas of Salento, olive pruning has been performed on a 4- to 5-year basis, and very large cuttings of the branches are usually performed. This practice is not considered suitable for correct management of tree physiology or productivity [146]. In addition, attempts to remove wilted parts after an OQDS outbreak through large cuts of the main branches with the aim of eliminating the pathogen(s) have always resulted in further weakening of the tree, followed by its death, especially in centennial trees. An appropriate time frame for pruning is one or two years. This also allows the zinc–copper–citric acid biocomplex to more effectively enter the foliage through nebulization [95] and mitigates the relevant zinc and copper depletion observed in the tree leaves infected by X. f. subsp. pauca [147].
The success of the control strategy, as for other plant diseases, is strongly due to the consistency of protocol applications over several months, along with the performance of other measures for vector control, management ensuring soil fertility, and appropriate timing of pruning. Spraying the biocomplex two or three times occasionally does not solve the problem. A proactive and professional farmer mentality is the basis for OQDS field control. An additional input for OQDS control will be provided by the compound(s) under study for N. mediterraneum. The introduction of a dangerous phytopathogen from abroad has changed the traditional olive agroecosystem of Salento, and in facing this unwanted plague, a modern and appropriate approach by farmers is fundamental [72].
Funding
This review was funded by the MIPAAF project “Salvaguardia e valorizzazione del patrimonio olivicolo italiano con azioni di ricerca nel settore della difesa fitosanitaria (SALVAOLIVI)” and by the agreement with Regione Puglia: “Strategie di controllo integrato per il contenimento di Xylella fastidiosa in oliveti pugliesi ed analisi epidemiologiche del complesso del disseccamento rapido dell’olivo (CoDiRO).
Conflicts of Interest
The author declares no conflict of interest.
References
- Martelli, G.P. The current status of the quick decline syndrome of olive in southern Italy. Phytoparasitica 2016, 44, 1–10. [Google Scholar] [CrossRef]
- Saponari, M.; Boscia, D.; Nigro, F.; Martelli, G.P. Identification of DNA sequences related to Xylella fastidiosa in oleander, almond and olive trees exhibiting leaf scorch symptoms in Apulia (Southern Italy). J. Plant Pathol. 2013, 94, 688. [Google Scholar]
- Cariddi, C.; Saponari, M.; Boscia, D.; De Stradis, A.; Loconsole, G.; Nigro, F.; Porcelli, F.; Potere, O.; Martelli, G.P. Isolation of a Xylella fastidiosa strain infecting olive and oleander in Apulia, Italy. J. Plant Pathol. 2014, 96, 425–429. [Google Scholar]
- Saponari, M.; Giampetruzzi, A.; Loconsole, G.; Boscia, D.; Saldarelli, P. Xylella fastidiosa in olive in Apulia: Where we stand. Phytopathology 2019, 109, 176–186. [Google Scholar] [CrossRef] [Green Version]
- Delbianco, A.; Czwienczek, E.; Pautasso, M.; Kozelska, S.; Monguidi, M.; Stancanelli, G. A new resource for research and risk analysis: The updated European Food Safety Authority database of Xylella spp. host plant species. Phytopathology 2019, 109, 213–215. [Google Scholar] [CrossRef] [Green Version]
- Chatterjee, S.; Almeida, R.P.P.; Lindow, S. Living in two worlds: The plant and insect lifestyles of Xylella fastidiosa. Annu. Rev. Phytopathol. 2008, 46, 243–271. [Google Scholar] [CrossRef] [Green Version]
- Cornara, D.; Morente, M.; Markheiser, A.; Bodino, N.; Tsai, C.-W.; Fereres, A.; Redak, R.A.; Perring, T.M.; Spotti Lopes, J.R. An overview of the worldwide vectors of Xylella fastidiosa. Entomol. Gen. 2019, 39, 157–181. [Google Scholar] [CrossRef]
- Castro, C.; Di Salvo, B.; Roper, M.C. Xylella fastidiosa: A re-emerging plant pathogen that threatens crops globally. PLoS Pathog. 2021, 17, e1009813. [Google Scholar] [CrossRef]
- Battistuzzi, F.U.; Feijao, A.; Hedges, S.B. A genomic timescale of prokaryote evolution: Insights into the origin of methanogenesis, phototrophy, and the colonization of land. BMC Evol. Biol. 2004, 4, 44. [Google Scholar] [CrossRef] [Green Version]
- Morris, J.L.; Puttick, M.N.; Clark, J.W.; Edwards, D.; Kenrick, P.; Pressel, S.; Wellman, C.H.; Yang, Z.; Schneider, H.; Donoghue, P.H.J. The timescale of early plant evolution. Proc. Natl. Acad. Sci. USA 2018, 115, E2274–E2283. [Google Scholar] [CrossRef] [Green Version]
- Bowles, A.M.C.; Bechtold, U.; Paps, J. The origin of land plants is rooted in two bursts of genomic novelty. Curr. Biol. 2020, 30, 530–536. [Google Scholar] [CrossRef] [Green Version]
- Field, K.J.; Pressel, S.; Duckett, J.G.; Rimington, W.R.; Bidartondo, M.I. Symbiotic options for the conquest of land. Trends Ecol. Evol. 2015, 30, 477–486. [Google Scholar] [CrossRef]
- Misof, B.; Liu, S.; Meusemann, K.; Peters, R.S.; Donath, A.; Mayer, C.; Frandsen, P.B.; Ware, J.; Flouri, T.; Beutel, R.G.; et al. Phylogenomics resolves the timing and pattern of insect evolution. Science 2014, 346, 763–767. [Google Scholar] [CrossRef]
- Morris, C.E.; Moury, B. Revisiting the concept of host range of plant pathogens. Annu. Rev. Phytopathol. 2019, 57, 63–90. [Google Scholar] [CrossRef]
- Lewis, L.A.; McCourt, R.M. Green algae and the origin of land plants. Botany 2004, 91, 1535–1556. [Google Scholar] [CrossRef] [Green Version]
- Yoon, H.S.; Hackett, J.; Ciniglia, C.; Pinto, G.; Bhattacharya, D. A molecular timeline for the origin of photosynthetic eukaryotes. Mol. Biol. Evol. 2004, 21, 809–818. [Google Scholar] [CrossRef] [Green Version]
- Donohue, T.J.; Codjell, R.J. Microorganisms and clean energy. Nat. Rev. Microbiol. 2006, 4, 800. [Google Scholar] [CrossRef] [Green Version]
- Firrao, G.; Scortichini, M.; Pagliari, L. Orthology-based estimate of the contribution of horizontal gene transfer from distantly related bacteria to the intraspecific diversity and differentiation of Xylella fastidiosa. Pathogens 2021, 10, 46. [Google Scholar] [CrossRef]
- Newman, K.L.; Almeida, R.P.P.; Porcell, A.K.; Lindow, S.E. Use of a green fluorescence strain for analysis of Xylella fastidiosa colonization of Vitis vinifera. Appl. Environ. Microbiol. 2003, 69, 7319–7327. [Google Scholar] [CrossRef] [Green Version]
- Pereira, W.E.L.; Ferreira, C.B.; Caserta, R.; Melotto, M.; De Souza, A.A. Xylella fastidiosa subsp. pauca and fastidiosa colonize Arabidopsis systemically and induce anthocyanin accumulation in infected leaves. Phytopathology 2019, 109, 225–232. [Google Scholar] [CrossRef] [Green Version]
- Jacques, M.-A.; Denancé, N.; Legendre, B.; Morel, E.; Briand, M.; Mississipi, S.; Durand, K.; Olivier, V.; Portier, P.; Poliakoff, F.; et al. New coffee plant-infecting Xylella fastidiosa variants derived via homologous recombination. Appl. Environ. Microbiol. 2016, 82, 1556–1568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergsma-Vlami, M.; Van de Bilt, J.L.J.; Tjou-Tam-Sin, N.N.A.; Van de Vossenberg, B.T.L.H.; Westenberg, M. Xylella fastidiosa in Coffea arabica ornamental plants imported from Costa Rica and Honduras in The Netherlands. J. Plant Pathol. 2015, 97, 395. [Google Scholar]
- Loconsole, G.; Saponari, M.; Boscia, D.; D’Attoma, G.; Morelli, M.; Martelli, G.P.; Almeida, R.P.P. Intercepted isolates of Xylella fastidiosa in Europe reveal novel genetic diversity. Eur. J. Plant Pathol. 2016, 146, 85–94. [Google Scholar] [CrossRef]
- Marcelletti, S.; Scortichini, M. Xylella fastidiosa CoDiRO strain associated with the olive quick decline syndrome in southern Italy belongs to a clonal complex of the subspecies pauca that evolved in Central America. Microbiology 2016, 162, 2087–2098. [Google Scholar] [CrossRef]
- Giampetruzzi, A.; Saponari, M.; Loconsole, G.; Boscia, D.; Savino, V.N.; Almeida, R.P.P.; Zicca, S.; Landa, B.B.; Chacon-Diaz, C.; Saldarelli, P. Genome-wide analysis provides evidence on the genetic relatedness of the emergent Xylella fastidiosa genotype in Italy to isolates from Central America. Phytopathology 2017, 107, 816–827. [Google Scholar] [CrossRef] [Green Version]
- EFSA. Susceptibility of Olea europea L. varieties to Xylella fastidiosa subsp. pauca ST53. EFSA J. 2015, 15, 4772. [Google Scholar]
- Bergsma-Vlami, M.; Van de Bilt, J.L.J.; Tjoi-Tam-Sin, N.N.A.; Helderman, C.M.; Gorkink-Smits, P.P.N.A.; Landman, N.M.; Van Nieuwburg, J.G.W.; Van Veen, E.J.; Westenberg, M. Assessment of the genetic diversity of Xylella fastidiosa in imported ornamental Coffea arabica plants. Plant Pathol. 2017, 66, 1065–1074. [Google Scholar] [CrossRef]
- Ramazzotti, M.; Cimaglia, F.; Gallo, A.; Ranaldi, F.; Surico, G.; Mita, G.; Bleve, G.; Marchi, G. Insights on a founder effect: The case of Xylella fastidiosa in the Salento area of Apulia, Italy. Phytopathol. Mediterr. 2018, 57, 8–25. [Google Scholar]
- Sicard, A.; Saponari, M.; Vanhove, M.; Castillo, A.I.; Giampetruzzi, A.; Loconsole, G.; Saldarelli, P.; Boscia, D.; Neema, C.; Almeida, R.P.P. Introduction and adaptation of an emerging pathogen to olive trees in Italy. Microb. Genom. 2021, 7, 000735. [Google Scholar] [CrossRef]
- Montero-Astúa, M.; Chacon-Diaz, C.; Aguilar, E.; Rodriguez, C.M.; Garita, L.; Villalobos, W.; Moreira, L.; Hartung, J.S.; Rivera, C. Isolation and molecular characterization of Xylella fastidiosa from coffee plants in Costa Rica. J. Microbiol. 2008, 46, 482–490. [Google Scholar] [CrossRef]
- Nunney, L.; Ortiz, B.; Russall, S.A.; Ruiz Sanchez, R.; Stouthamer, R. The complex biogeography of the plant pathogen Xylella fastidiosa: Genetic evidence of introduction and subspecific introgression in Central America. PLoS ONE 2014, 9, e112463. [Google Scholar] [CrossRef] [Green Version]
- Mathias, M.C. Costa Rica: A giant in ornamentals. FlowerTech 2006, 9, 18–21. [Google Scholar]
- EFSA Panel on Plant Health (PLH). Scientific Opinion on the risks to plant health posed by Xylella fastidiosa in the EU territory, with the identification and evaluation of risk reduction options. EFSA J. 2015, 13, 3989. [Google Scholar] [CrossRef]
- Lombardo, L.; Rizzo, P.; Novellis, C.; Vizzarri, V. Preliminary molecular survey of the possible presence of Xylella fastidiosa in the upper ionian coasts of Calabria, Italy, through the capture and analysis of its main vector insects. Insects 2021, 12, 446. [Google Scholar] [CrossRef]
- Trotta, V.; Forlano, P.; Caccavo, V.; Fanti, P.; Battaglia, D. A survey of potential vectors of the plant pathogenic bacterium Xylella fastidiosa in the Basilicata region, Italy. Bull. Insect. 2021, 74, 273–283. [Google Scholar]
- Cantini, C.; Poggioni, L.; Nencioni, A.; Sacchetti, P. Evaluation of the population of the meadow spittlebug Philaenus spumarius in Tuscan olive groves. In Proceedings of the 2nd Conference on Xylella fastidiosa: How Research Can Support Solutions, Ajaccio, France, 29–30 October 2019; pp. 91–92. [Google Scholar]
- Scortichini, M. Predisposing factors for “olive quick decline syndrome” in Salento (Apulia, Italy). Agronomy 2020, 10, 1445. [Google Scholar] [CrossRef]
- Kottelenberg, D.; Hemerick, L.; Saponari, M.; Van der Werf, W. Shape and rate of movement of the invasion front of Xylella fastidiosa spp. pauca in Puglia. Sci. Rep. 2021, 11, 1061. [Google Scholar] [CrossRef]
- Semeraro, T.; Buccolieri, R.; Vergine, M.; De Bellis, L.; Luvisi, A.; Emmanuel, R.; Marwan, R. Analysis of olive grove destruction by Xylella fastidiosa bacterium on the land surface temperature in Salento detected using satellite images. Forests 2021, 12, 1266. [Google Scholar] [CrossRef]
- Ciervo, M. II Disseccamento Degli Olivi in Puglia, Evidenze, Contraddizioni, Anomalie, Scenari; Un punto di vista geografico; Società Geografica Italiana: Roma, Italy, 2020; 74p. [Google Scholar]
- Fierro, A.; Liccardo, A.; Porcelli, F. A lattice model to manage the vector and infection of the Xylella fastidiosa in olive trees. Sci. Rep. 2019, 9, 8723. [Google Scholar] [CrossRef] [Green Version]
- Stokstad, E. Italy’s olives under siege. Science 2015, 348, 620. [Google Scholar] [CrossRef] [PubMed]
- White, S.M.; Bullock, J.M.; Hooftman, D.A.P.; Chapman, D.S. Modelling the spread and control of Xylella fastidiosa in the early stages of invasion in Apulia, Italy. Biol. Invasions 2017, 19, 1825–1837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zoina, A. (University “Federico II”, Naples, Italy). Personal communication, 2018. [Google Scholar]
- Totorizzo, G. II Viaggio Della Xylella Sulla Statale Lecce-Bari. “Vettore Sulle Gomme” La Mappa Lo Dimostra; La Repubblica-Bari: Bari, Italy, 2022; p. 2. [Google Scholar]
- EFSA Panel on plant health (PLH). Update of the scientific opinion on the risks to plant health posed by Xylella fastidiosa in the EU territory. EFSA J. 2019, 17, 5665. [Google Scholar]
- Van Loon, A.F. Hydrological drought explained. Wiley Interdiscip. Rev. Water 2015, 2, 359–392. [Google Scholar]
- Desprez-Loustau, M.-L.; Marcais, B.; Nageleisen, L.-M.; Piou, D.; Vannini, A. Interactive effects of drought and pathogens in forest trees. Ann. For. Sci. 2006, 63, 597–612. [Google Scholar] [CrossRef] [Green Version]
- Sturrock, R.N.; Frankel, S.J.; Brown, A.V.; Hennon, P.E.; Kliejunas, J.T.; Lewis, K.J.; Worrall, J.J.; Woods, A.J. Climate change and forest diseases. Plant Pathol. 2011, 60, 133–149. [Google Scholar] [CrossRef]
- Wakelin, S.A.; Gomez-Gallego, M.; Jones, E.; Smaill, S.; Lear, G.; Lambie, S. Climate change induced drought impacts on plant disease in New Zealand. Austral. Plant Pathol. 2018, 47, 101–114. [Google Scholar] [CrossRef]
- Caldeira, M.C. The timing of drought coupled with pathogens may boost tree mortality. Tree Physiol. 2019, 39, 1–5. [Google Scholar] [CrossRef]
- Gargallo-Garriga, A.; Sardans, J.; Pérez-Trujillo, M.; Rivas-Ubach, A.; Oravec, M.; Vecerova, K.; Urban, O.; Jentsch, A.; Kreyling, J.; Beierkuhnlein, C.; et al. Opposite metabolic responses of shoots and roots to drought. Sci. Rep. 2014, 4, 6829. [Google Scholar] [CrossRef] [Green Version]
- He, M.; Dijkstra, F.A. Drought effect on plant nutrition and phosphorus: A meta-analysis. New Phytol. 2014, 204, 924–931. [Google Scholar] [CrossRef]
- Oliva, J.; Stenlid, J.; Martínez-Vilalta, J. The effect of fungal pathogens on the water and carbon economy of trees: Implications for drought induced mortality. New Phytol. 2014, 203, 1028–1035. [Google Scholar] [CrossRef] [Green Version]
- Fukuda, K.; Utsuzawa, S.; Sakaue, D. Correlation between acoustic emission, water status and xylem embolism in pine wilt disease. Tree Physiol. 2007, 6, 969–976. [Google Scholar] [CrossRef] [Green Version]
- Yadeta, K.A.; Thomma, B.P.H.J. The xylem as battleground for plant hosts and vascular wilt pathogens. Front. Plant Sci. 2013, 4, 97. [Google Scholar] [CrossRef] [Green Version]
- Angelopoulos, K.; Dichio, B.; Xiloyannis, C. Inhibition of photosynthesis in olive trees (Olea europea L.) during water stress and rewatering. J. Exp. Bot. 1996, 47, 1093–1100. [Google Scholar] [CrossRef] [Green Version]
- O’Hara, N.B.; Franks, S.J.; Kane, N.C.; Tittes, S.; Rest, J.S. Evolution of pathogen response genes associated with increased disease susceptibility during adaptation to an extreme drought in a Brassica rapa plant population. BMC Ecol. Evol. 2021, 21, 61. [Google Scholar] [CrossRef]
- Choudhary, A.; Senthil-Kumar, M. Drought attenuates plant defense against bacterial pathogens by suppressing the expression of CBP60g/SARD1 during combined stress. Plant Cell Environ. 2022, 45, 1127–1145. [Google Scholar] [CrossRef]
- Barradas, C.; Pinto, G.; Correia, B.; Castro, B.B.; Phillips, A.J.L.; Alves, A. Drought × disease interaction in Eucalyptus globulus under Neofusicoccum eucalyptorum infection. Plant Pathol. 2018, 67, 87–96. [Google Scholar] [CrossRef]
- Galarneanu, E.R.A.; Lawrence, D.P.; Travadon, R.; Baumgartner, K. Drought exacerbates Botryosphaeria dieback symptoms in grapevines and confounds host-based molecular markers of infection by Neofusicoccum parvum. Plant Dis. 2019, 103, 1738–1745. [Google Scholar] [CrossRef]
- McElrone, A.J.; Sherald, J.L.; Forseth, I.N. Effects of water stress on symptomatology and growth of Parthenocissus quinquefolia infected by Xylella fastidiosa. Plant Dis. 2001, 85, 1160–1164. [Google Scholar] [CrossRef] [Green Version]
- Choi, H.-K.; Iandolino, A.; Goes da Silva, F.; Cook, D.R. Water deficit modulates the response of Vitis vinifera to the Pierce’s disease pathogen Xylella fastidiosa. Mol. Plant-Microb. Interact. 2013, 26, 643–657. [Google Scholar] [CrossRef]
- Mendes, J.S.; Da, S.; Santiago, A.; Toledo, M.A.S.; Rosselli-Murai, L.K.; Favaro, M.T.P.; Santos, C.A.; Horta, M.A.C.; Crucello, A.; Beloti, L.L.; et al. VapD in Xylella fastidiosa is a thermostable protein with ribonuclease activity. PLoS ONE 2015, 10, e0145765. [Google Scholar] [CrossRef]
- Laranjeira, F.F.; Bergamin Filho, A.; Amorim, L.; Aguilar-Vildoso, C.I.; Coletta Filho, H. Fungos, Procariotos e Doenças Abióticas; Mattos Júnior, D., De Negri, J.D., Pio, R.M., Pompeu Júnior, J., Eds.; Citros; Instituto Agronômico/Fundag: Campinas, Brazil, 2005; pp. 509–566. [Google Scholar]
- Alfio, M.R.; Balacco, G.; Parisi, A.; Totaro, V.; Fidelibus, M.D. Drought index as indicator of salinization of the Salento aquifer (Southern Italy). Water 2020, 12, 1927. [Google Scholar] [CrossRef]
- De Pascali, M.; Vergine, M.; Sabella, E.; Aprile, A.; Nutricati, E.; Nicoli, F.; Buja, I.; Negro, C.; Miceli, A.; Rampino, P.; et al. Molecular effects of Xylella fastidiosa and drought combine stress in olive trees. Plants 2019, 8, 437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strona, G.; Carstens, C.J.; Beck, P.S.A. Network analysis reveal why Xylella fastidiosa will persist in Europe. Sci. Rep. 2017, 7, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dongiovanni, C.; Cavalieri, V.; Bodino, N.; Tauro, T.; Di Carolo, M.; Fumarola, G.; Altamura, G.; Lasorella, C.; Bosco, D. Plant selection and population trend of spittlebug immatures (Hemiptera: Aphrophoridae) in olive groves of the Apulia region of Italy. J. Econ. Entomol. 2018, 112, 67–74. [Google Scholar] [CrossRef] [Green Version]
- Bodino, N.; Cavalieri, V.; Dongiovanni, C.; Simonetto, A.; Saladini, M.A.; Plazio, E.; Gilioli, G.; Molinatto, G.; Saponari, M.; Bosco, D. Dispersal of Philaenus spumarius (Hemiptera: Aphrophoridae), a vector of Xylella fastidiosa, in olive grove and meadow agroecosystems. Behav. Ecol. 2021, 50, 267–279. [Google Scholar] [CrossRef]
- Saponari, M.; Loconsole, G.; Cornara, D.; Yokomi, R.K.; De Stradis, R.A.; Boscia, D.; Bosco, D.; Martelli, G.P.; Krugner, R.; Porcelli, F. Infectivity and transmission of Xylella fastidiosa by Philaenus spumarius (Hemiptera: Aphrophoridae) in Apulia, Italy. J. Econom. Entomol. 2014, 107, 1316–1319. [Google Scholar] [CrossRef] [Green Version]
- Scortichini, M. The multi-millenial olive agroecosystem of Salento (Apulia, Italy) threatened by Xylella fastidiosa subsp. pauca: A working possibility of restoration. Sustainability 2020, 12, 6700. [Google Scholar] [CrossRef]
- Del Coco, L.; De Pascali, S.A.; Fanizzi, F.P. NMR-metabolomic study on monovarietal and blend Salento EVOOs including some from secular olive trees. Food Nutr. Sci. 2014, 5, 89–95. [Google Scholar]
- Negro, C.; Aprile, A.; Luvisi, A.; Nicolì, F.; Nutricati, E.; Vergine, M.; Miceli, A.; Blando, F.; Sabella, E.; De Bellis, L. Phenolic profile and antioxidant activity of Italian monovarietal extravirgin olive oil. Antioxidants 2019, 8, 161. [Google Scholar] [CrossRef] [Green Version]
- De Benedictis, M.; De Caroli, M.; Baccelli, I.; Marchi, G.; Bleve, G.; Gallo, A.; Ranaldi, F.; Falco, V.; Pasquali, V.; Piro, G.; et al. Vessel occlusion in three cultivars of Olea europaea naturally exposed to Xylella fastidiosa in open field. J. Phytopathol. 2017, 165, 589–594. [Google Scholar] [CrossRef]
- Petit, G.; Bleve, G.; Gallo, A.; Mita, G.; Montanaro, G.; Nuzzo, V.; Zambonini, D.; Pitacco, A. Susceptibility to Xylella fastidiosa and functional xylem anatomy in Olea europea: Revisiting a tale of plant-pathogen interaction. AoB Plants 2021, 13, 4. [Google Scholar] [CrossRef]
- Cardinale, M.; Luvisi, A.; Meyer, J.B.; Sabella, E.; De Bellis, L.; Cruz, A.C.; Ampatzidis, Y.; Cherubini, P. Specific fluorescent in situ hybridization (FISH) test to highlight colonization of xylem vessels by Xylella fastidiosa in naturally infected olive trees (Olea europea L.). Front. Plant Sci. 2018, 9, 431. [Google Scholar] [CrossRef] [Green Version]
- Sabella, E.; Aprile, A.; Genga, A.; Siciliano, T.; Nutricati, E.; Nicoli, F.; Vergine, M.; Negro, C.; De Bellis, L.; Luvisi, A. Xylem cavitation susceptibility and refilling mechanisms in olive trees infected by Xylella fastidiosa. Sci. Rep. 2019, 9, 9602. [Google Scholar] [CrossRef] [Green Version]
- Luvisi, A.; Aprile, A.; Sabella, E.; Vergine, M.; Nicolì, F.; Nutricati, E.; Miceli, A.; Negro, C.; De Bellis, L. Xylella fastidiosa subsp. pauca (CoDiRO strain) in four olive (Olea europea L.) cultivars: Profile of phenolic compounds in leaves and progression of leaf scorch symptoms. Phytopathol. Mediterr. 2017, 56, 259–273. [Google Scholar]
- Saponari, M.; Boscia, D.; Altamura, G.; Loconsole, G.; Zicca, S.; D’Attoma, G.; Morelli, M.; Palmisano, F.; Saponari, A.; Tavano, D.; et al. Isolation and pathogenicity of Xylella fastidiosa associated to the olive quick decline syndrome in southern Italy. Sci. Rep. 2017, 7, 17723. [Google Scholar] [CrossRef] [Green Version]
- Pavan, S.; Vergine, M.; Nicoli, F.; Sabella, E.; Aprile, A.; Negro, C.; Fanelli, V.; Savoia, M.A.; Montilon, V.; Susca, L.; et al. Screening of olive biodiversity defines genotypes potentially resistant to Xylella fastidiosa. Front. Plant Sci. 2021, 12, 723879. [Google Scholar] [CrossRef]
- Del Coco, L.; Migoni, D.; Girelli, C.R.; Angilè, F.; Scortichini, M.; Fanizzi, F.P. Soil and leaf ionome heterogeneity in Xylella fastidiosa subsp. pauca-infected, non-infected and treated olive groves in Apulia, Italy. Plants 2020, 9, 760. [Google Scholar] [CrossRef]
- Eide, D.J. The oxidative stress of zinc deficiency. Metallomics 2011, 3, 1124–1129. [Google Scholar] [CrossRef]
- Caspi, V.; Droppa, M.; Horvath, G.; Malkin, S.; Marder, J.B.; Raskin, V.I. The effect of copper on chlorophyll organization during greening of barley leaves. Photosynth. Res. 1999, 62, 165–174. [Google Scholar] [CrossRef]
- Alejandro, S.; Höller, S.; Meier, B.; Peiter, E. Manganese in plants: From acquisition to subcellular allocation. Front. Plant Sci. 2020, 11, 300. [Google Scholar] [CrossRef] [Green Version]
- Ciervo, M. The olive quick decline syndrome (OQDS) diffusion in Apulia region: An apparent contradiction according to the agricultural model. Belgeo 2016, 4. [Google Scholar] [CrossRef]
- Collavo, A. Resistenza al Glifosate nelle Colture Arboree; Veneto Agricoltura, Forum Fitoiatrico 19 Gennaio; Legnaro (PD), Italy. 2012. Available online: https://www.venetoagricoltura.org/upload/pubblicazioni/Forum%20Fito%20023/COLLAVO.pdf (accessed on 17 July 2022).
- Ferreira, I.Q.; Arrobas, M.; Claro, A.M.; Rodrigues, M.A. Soil management in rainfed olive orchards may result in conflicting effects on olive production and soil fertility. Span. J. Agric. Res. 2013, 2, 472–480. [Google Scholar] [CrossRef]
- Kremer, R.J.; Means, N.E.; Kim, S. Glyphosate affects soybean root exudation and rhizosphere microorganisms. Intern. J. Anal. Chem. 2005, 85, 1165–1174. [Google Scholar] [CrossRef] [Green Version]
- Kremer, R.J.; Means, N.E. Glyphosate and glyphosate-resistant crop interactions with rhizosphere microorganisms. Europ. J. Agron. 2009, 31, 153–161. [Google Scholar] [CrossRef]
- Aristide, L.; Reed, R.L.; Wilkes, R.A.; Youngster, T.; Kukurugya, M.A.; Katz, V.; Sasaki, C.R.S. Glyphosate-induced specific and widespread perturbations in the metabolome of soil Pseudomonas species. Front. Environ. Sci. 2017, 5, 34. [Google Scholar] [CrossRef] [Green Version]
- Serra, A.P.; Marchetti, M.E.; Da Silva Candido, A.C.; Ribero Dias, A.C.; Christoffoleti, P.J. Glyphosate influence on nitrogen, manganese, iron, copper and zinc nutritional efficiency in glyphosate resistant soybean. Cienc. Rural 2011, 41, 77–84. [Google Scholar] [CrossRef] [Green Version]
- Martinez, D.A.; Loaning, U.E.; Graham, M.C. Impacts of glyphosate-based herbicides on disease resistance and health of crops: A review. Environ. Sci. Eur. 2018, 30, 2. [Google Scholar] [CrossRef] [Green Version]
- De Nadal Gasparini dos Santos, B.; Anguita-Maeso, M.; Della Coletta Filho, H. Transmission and distribution of Xylella fastidiosa subsp. pauca in olive trees as a parameter for managing olive quick decline syndrome. Plant Pathol. 2022. [Google Scholar] [CrossRef]
- Scortichini, M.; Chen, J.; De Caroli, M.; Dalessandro, G.; Pucci, N.; Modesti, V.; L’Aurora, A.; Petriccione, M.; Zampella, L.; Mastrobuoni, F.; et al. A zinc-copper-citric acid biocomplex shows promise for control of Xylella fastidiosa subsp. pauca in olive trees in Apulia region (southern Italy). Phytopathol. Mediterr. 2018, 57, 48–72. [Google Scholar]
- Parent, C.; Capelli, N.; Berger, A.; Crèvecoeur, M. An overview of plant responses to waterlogging. Plant Stress 2008, 2, 20–27. [Google Scholar]
- Navarro, C.; Parra, M.A. Plantacíon. In El Cultivo del Olivo; Barranco, D., Fernández-Escobar, R., Rallo, L., Eds.; Junta de Andalucía y Mundi-Prensa: Madrid, Spain, 2008; pp. 163–195. [Google Scholar]
- Hill, B.L.; Purcell, A.H. Populations of Xylella fastidiosa in plants required for transmission by an efficient vector. Phytopathology 1997, 87, 1197–1201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Killiny, N.; Almeida, R.P.P. Factors affecting the initial adhesion of and retention of the plant pathogen Xylella fastidiosa in the foregut of an insect vector. Appl. Environ. Microbiol. 2014, 80, 420–426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cornara, D.; Marra, M.; Morente, M.; Garzo, E.; Moreno, A.; Saponari, M.; Fereres, A. Feeding behavior in relation to spittlebug transmission of Xylella fastidiosa. J. Pest Sci. 2020, 93, 1197–1213. [Google Scholar] [CrossRef]
- Olmo, D.; Nieto, A.; Borràs, D.; Montesinos, M.; Adrover, F.; Pascual, A.; Gost, P.A.; Quetglas, B.; Urbano, A.; De Dios Garcia, A.; et al. Landscape epidemiology of Xylella fastidiosa in the Balearic Islands. Agronomy 2021, 11, 473. [Google Scholar] [CrossRef]
- Cornara, D.; Cavalieri, V.; Dongiovanni, C.; Altamura, G.; Palmisano, F.; Bosco, D.; Porcelli, F.; Almeida, R.P.P.; Saponari, M. Transmission of Xylella fastidiosa by naturally infected Philaenus spumarius (Hemiptera, Aphrophoridae) to different host plants. J. Appl. Entomol. 2017, 141, 80–87. [Google Scholar] [CrossRef]
- Picciotti, U.; Lahbib, N.; Sefa, V.; Porcelli, F.; Garganese, F. Aphrophoridae role in Xylella fastidiosa subsp. pauca ST53 invasion in Southern Italy. Pathogens 2021, 10, 1035. [Google Scholar] [CrossRef]
- Cavalieri, V.; Altamura, G.; Fumarola, G.; Di Carolo, M.; Saponari, M.; Cornara, D.; Bosco, D.; Dongiovanni, C. Transmission of Xylella fastidiosa subspecies pauca sequence type 53 by different insect species. Insects 2019, 10, 324. [Google Scholar] [CrossRef] [Green Version]
- Cornara, D.; Bosco, D.; Fereres, A. Philaenus spumarius: When an old acquaintance becomes a new threat to European agriculture. J. Pest. Sci. 2018, 91, 957–972. [Google Scholar] [CrossRef]
- Bodino, N.; Cavalieri, V.; Dongiovanni, C.; Plazio, E.; Saladini, M.A.; Volani, S.; Simonetto, A.; Fumarola, G.; Di Carolo, M.; Porcelli, F.; et al. Phenology, seasonal abundance, and stage-structure of spittlebug (Hemiptera: Aphrophoridae) populations in olive groves of Italy. Sci. Rep. 2019, 9, 17725. [Google Scholar] [CrossRef] [Green Version]
- White, S.M.; Navas-Cortes, J.A.; Bullock, J.M.; Boscia, D.; Chapman, D.S. Estimating the epidemiology of emerging Xylella fastidiosa outbreaks in olives. Plant Pathol. 2020, 69, 1403–1413. [Google Scholar] [CrossRef]
- Sicard, A.; Zeilinger, A.R.; Vanhove, M.; Schartel, T.E.; Beal, D.J.; Daugherty, M.P.; Almeida, R.P.P. Xylella fastidiosa: Insights into an emerging plant pathogen. Annu. Rev. Phytopathol. 2018, 56, 181–202. [Google Scholar] [CrossRef] [Green Version]
- Cornara, D.; Panzarino, O.; Santoiemma, G.; Bodino, N.; Loverre, P.; Mastronardi, M.G.; Mattia, C.; De Lillo, E.; Addante, R. Natural areas as reservoir of candidate vectors of Xylella fastidiosa. Bull. Insect. 2021, 74, 173–180. [Google Scholar]
- Buonaurio, R.; Moretti, C.; Passos da Silva, D.; Cortese, C.; Ramos, C.; Venturi, V. The olive knot disease as a model to study the role of interspecies bacterial communities in plant disease. Front. Plant Sci. 2015, 6, 434. [Google Scholar] [CrossRef] [Green Version]
- Zine el Aabidine, A.; Baissac, Y.; Moukhli, A.; Jay-Allemand, C.; Khadari, B.; El Modafar, C. Resistance of olive tree to Spilocaea oleaginea is mediated by the synthesis of phenolic compounds. Int. J. Agr. Biol. 2010, 12, 61–67. [Google Scholar]
- Nigro, F.; Boscia, D.; Antelmi, I.; Ippolito, A. Fungal species associated with a severe decline of olive in southern Italy. J. Plant Pathol. 2013, 95, 668. [Google Scholar]
- Carlucci, A.; Raimondo, M.L.; Cibelli, F.; Phillips, A.J.L.; Lops, F. Pleurostomophora richardsiae, Neofusicoccum parvum and Phaeoacremonium aleophilum associated with decline of olive in southern Italy. Phytopathol. Mediterr. 2013, 52, 517–527. [Google Scholar]
- Gramaje, D.; Agusti-Brisach, C.; Pèrez-Sierra, A.; Moralejo, E.; Olmo, D.; Mostert, L.; Damm, U.; Armengal, J. Fungal trunk pathogens associated with wood decay of almond trees on Mallorca (Spain). Persoonia 2012, 28, 1–13. [Google Scholar] [CrossRef]
- Brunetti, A.; Matere, A.; Lumia, V.; Pasciuta, V.; Fusco, V.; Sansone, D.; Marangi, P.; Cristella, N.; Faggioli, F.; Scortichini, M.; et al. Neofusicoccum mediterraneum is involved in a twig and branch dieback of olive trees observed in Salento (Apulia, Italy). Pathogens 2022, 11, 53. [Google Scholar] [CrossRef]
- Pilotti, M. (CREA-Research Centre for Plant Protection and Certification, Roma, Italy). Personal communication, 2022. [Google Scholar]
- Moral, J.; Muñoz-Díez, C.; González, N.; Trapero, A.; Michailides, T.J. Characterization and pathogenicity of Botryosphaeriaceae species collected from olive and other hosts in Spain and California. Phytopathology 2010, 100, 1340–1351. [Google Scholar] [CrossRef] [Green Version]
- Úrbez-Torres, J.R.; Peduto, F.; Vossen, P.M.; Krueger, W.H.; Gubler, W.D. Olive twig and branch dieback: Etiology, incidence, and distribution in California. Plant Dis. 2013, 97, 231–244. [Google Scholar] [CrossRef] [Green Version]
- Moral, J.; Agustí-Brisach, C.; Pérez-Rodríguez, M.; Xaviér, C.; Raya, M.C.; Rhouma, A.; Trapero, A. Identification of fungal species associated with branch dieback of olive and resistance of table cultivars to Neofusicoccum mediterraneum and Botryosphaeria dothidea. Plant Dis. 2017, 101, 306–316. [Google Scholar] [CrossRef] [Green Version]
- Romero, M.A.; Sánchez, M.E.; Trapero, A. First report of Botryosphaeria ribis as a branch dieback pathogen of olive trees in Spain. Plant Dis. 2007, 89, 20. [Google Scholar] [CrossRef]
- Scortichini, M.; Cesari, G. An evaluation of the monitoring surveys of the quarantine bacterium Xylella fastidiosa performed in containment and buffer areas of Apulia, southern Italy. Appl. Biosafety 2019, 24, 96–99. [Google Scholar] [CrossRef]
- Lazzizera, C.; Frisullo, S.; Alves, A.; Phillips, A.J.L. Morphology, phylogeny and pathogenicity of Botryosphaeria and Neofusicoccum species associated with drupe rot of olives in southern Italy. Plant Pathol. 2008, 57, 948–956. [Google Scholar] [CrossRef]
- Denman, S.; Doonan, J.; Ramson-Jones, E.; Broberg, M.; Plummer, S.; Kirk, S.; Scarlett, K.; Griffiths, A.R.; Kaczmarek, M.; Forster, J.; et al. Microbiome and infectivity studies reveal complex polyspecies tree disease in acute oak decline. ISME J. 2018, 12, 386–399. [Google Scholar] [CrossRef] [Green Version]
- Griffiths, S.M.; Galambao, M.; Rowntree, J.; Goodhead, I.; Hall, J.; O’Brien, D.; Atkinson, N.; Antwis, R.E. Complex associations between cross-kingdom microbial endophytes and host genotype in ash dieback disease dynamics. J. Ecol. 2020, 108, 291–309. [Google Scholar] [CrossRef] [Green Version]
- Hrycan, J.; Hart, M.; Bowen, P.; Forge, T.; Urbez-Torres, J.R. Grapevine trunk disease fungi: Their roles as latent pathogens and stress factors that favour disease development and symptom expression. Phytopathol. Mediterr. 2020, 59, 395–424. [Google Scholar]
- Boscia, D.; Altamura, D.; Ciniero, A.; Di Carolo, M.; Dongiovanni, C.; Fumarola, G.; Giampetruzzi, A.; Greco, P.; La Notte, P.; Loconsole, G.; et al. Resistenza a Xylella fastidiosa in diverse cultivar di olivo. Inf. Agrar. 2017, 11, 59–63. [Google Scholar]
- Giampetruzzi, A.; Morelli, M.; Saponari, M.; Loconsole, G.; Chiumenti, M.; Boscia, D.; Savino, V.; Martelli, G.P.; Saldarelli, P. Transcriptome profiling of two olive cultivars in response to infection by the CoDiRO strain of Xylella fastidiosa subsp. pauca. BMC Genom. 2016, 17, 475. [Google Scholar] [CrossRef] [PubMed]
- Sabella, E.; Luvisi, A.; Aprile, A.; Negro, C.; Vergine, M.; Nicolì, F.; Miceli, A.; De Bellis, L. Xylella fastidiosa induces differential expression of lignification related-genes and lignin accumulation in tolerant olive trees cv. Leccino. J. Plant Physiol. 2018, 220, 60–68. [Google Scholar] [CrossRef] [PubMed]
- EFSA PLH Panel. Scientific opinion on the updated pest categorization of Xylella fastidiosa. EFSA J. 2018, 16, 5357. [Google Scholar]
- EFSA PLH Panel. Scientific opinion on the effectiveness of in planta control measures for Xylella fastidiosa. EFSA J. 2019, 17, 5666. [Google Scholar]
- Holling, C.S. Resilience and stability of ecological systems. Annu. Rev. Ecol. Syst. 1973, 4, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Cholet, C.; Bruez, E.; Lecomte, P.; Barsacq, A.; Martignon, T.; Giudici, M.; Simonit, M.; Rey, P.; Dubourdieu, D.; Gény, L. Plant resilience and physiological modifications induced by curettage of Esca-diseased grapevines. OENO One 2021, 1, 153–169. [Google Scholar] [CrossRef]
- Loreti, S. (CREA-Research Centre for Plant Protection and Certification, Roma, Italy). Personal communication, 2022. [Google Scholar]
- Kim, J.H.; Hilleary, L.; Seroka, A.; He, S.H. Crops of the future: Building a climate-resilient plant immune system. Curr. Opin. Plant Biol. 2021, 60, 101997. [Google Scholar] [CrossRef]
- Agrios, G.N. Plant Pathology, 5th ed.; Academic Press Books; Elsevier: Amsterdam, The Netherlands, 2004. [Google Scholar]
- Dowdle, W.R. The principles of disease elimination and eradication. Bull. World Health Organ. 1998, 76 (Suppl. S2), 22–25. [Google Scholar]
- Tatulli, G.; Modesti, V.; Pucci, N.; Scala, V.; L’Aurora, A.; Lucchesi, S.; Salustri, M.; Scortichini, M.; Loreti, S. Further in vitro assessment and mid-term evaluation of control strategy of Xylella fastidiosa subsp. pauca in olive groves of Salento (Apulia, Italy). Pathogens 2021, 10, 85. [Google Scholar] [CrossRef]
- Cobine, P.A.; Cruz, L.F.; Navarrete, F.; Duncan, D.; Tygart, M.; De La Fuente, L. Xylella fastidiosa differentially accumulates mineral elements in biofilm and planktonic cells. PLoS ONE 2013, 8, e54936. [Google Scholar] [CrossRef]
- Navarrete, F.; De La Fuente, L. Response of Xylella fastidiosa to zinc: Decreased culturability, increased exopolysaccharide production, and formation of resilient biofilms under flow conditions. Appl. Env. Microbiol. 2014, 80, 1097–1107. [Google Scholar] [CrossRef] [Green Version]
- Navarrete, F.; De La Fuente, L. Zinc detoxification is required for full virulence and modification of the host leaf ionome by Xylella fastidiosa. Mol. Plant Microbe Interact. 2015, 28, 497–507. [Google Scholar] [CrossRef] [Green Version]
- Scortichini, M. Sustainable management of diseases in horticulture: Conventional and new options. Horticulturae 2022, 8, 517. [Google Scholar] [CrossRef]
- Keane, P.J.; Kerr, A. Factors affecting disease development. In Plant Pathogens and Plant Disease; Brown, J.F., Ogle, H.G., Eds.; Rockvale Publications: Armidale, Australia, 1997; pp. 287–298. [Google Scholar]
- Ghorbani, R.; Wilcockson, S.; Koochekl, A.; Leifert, C. Soil management for sustainable crop disease control: A review. In Organic Farming, Pest Control and Remediation of Soil Pollutants; Lichtfouse, E., Ed.; Springer: San Francisco, CA, USA, 2009; pp. 177–227. [Google Scholar]
- Morales, A.; Hurtado, M.I.; Fischer, T. Influence of pruning intensity of olive trees on the oil characteristics (“Coratina”). Acta Hortic. 2014, 1057, 717–723. [Google Scholar] [CrossRef]
- Sibbett, G.S. Pruning mature bearing olive trees. In Olive Production Manual, 2nd ed.; Sibbett, G.S., Ferguson, L., Eds.; University of California, Division of Agriculture and Natural Resource: Oakland, CA, USA, 2005; pp. 55–59. [Google Scholar]
- Rodrigues, M.A.; Lopes, J.I.; Ferreira, I.Q.; Arrobas, M. Olive tree response to the severity of pruning. Turk. J. Agric. For. 2018, 42, 103–113. [Google Scholar] [CrossRef]
- Scortichini, M.; Migoni, D.; Angilè, F.; Del Coco, L.; Girelli, C.R.; Zampella, L.; Mastrobuoni, F.; Fanizzi, F.P. Xylella fastidiosa subsp. pauca on olive in Salento (Southern Italy): Infected trees have low in planta micronutrient content. Phytopathol. Mediterr. 2019, 58, 39–48. [Google Scholar]
Figure 1.
Taxonomic subdivision of the nearest ortholog assignment of all genes in the Xylella fastidiosa pan-genome. It appears that about 20% of the total genes of Xylella fastidiosa have been acquired from distantly related bacterial taxa. (Data from [18]).
Figure 1.
Taxonomic subdivision of the nearest ortholog assignment of all genes in the Xylella fastidiosa pan-genome. It appears that about 20% of the total genes of Xylella fastidiosa have been acquired from distantly related bacterial taxa. (Data from [18]).

Figure 2.
Symptomatic ornamental coffee (Coffea arabica L.) plant introduced in 2015 in Italy from Costa Rica, intercepted by the regional phytosanitary service of the Regione Veneto, and resulted positive for Xylella fastidiosa upon detection. The plant was destroyed immediately after the detection. (Reproduced with permission of the regional phytosanitary service of Regione Veneto).
Figure 2.
Symptomatic ornamental coffee (Coffea arabica L.) plant introduced in 2015 in Italy from Costa Rica, intercepted by the regional phytosanitary service of the Regione Veneto, and resulted positive for Xylella fastidiosa upon detection. The plant was destroyed immediately after the detection. (Reproduced with permission of the regional phytosanitary service of Regione Veneto).

Figure 3.
Synoptic panel that illustrates the predisposing factors, spreading factors, and occurrence of additional phytopathogens all involved in possibly causing the “olive quick decline syndrome” in Salento (Apulia, Italy).
Figure 3.
Synoptic panel that illustrates the predisposing factors, spreading factors, and occurrence of additional phytopathogens all involved in possibly causing the “olive quick decline syndrome” in Salento (Apulia, Italy).

Figure 4.
Box plot for the discriminant microelements sodium (Na), copper (Cu), and zinc (Zn) as obtained for olive leaf samples obtained from olive groves of the non-infected area BAT - PZ (BAT: Barletta—Andria—Trani and PZ: Potenza provinces) and from olive groves of the infected areas: BR (province of Brindisi), LE (province of Lecce), and TA (province of Taranto). C: panel C from the figure reproduced from [82].
Figure 4.
Box plot for the discriminant microelements sodium (Na), copper (Cu), and zinc (Zn) as obtained for olive leaf samples obtained from olive groves of the non-infected area BAT - PZ (BAT: Barletta—Andria—Trani and PZ: Potenza provinces) and from olive groves of the infected areas: BR (province of Brindisi), LE (province of Lecce), and TA (province of Taranto). C: panel C from the figure reproduced from [82].

Figure 5.
Waterlogging in an olive grove of Salento due to a relative excess of precipitation occurring over a short time. This situation occurs quite frequently in recent years due to climate change. Prolonged anaerobic conditions in the soil can occur, causing deleterious effects to the root system of the trees.
Figure 5.
Waterlogging in an olive grove of Salento due to a relative excess of precipitation occurring over a short time. This situation occurs quite frequently in recent years due to climate change. Prolonged anaerobic conditions in the soil can occur, causing deleterious effects to the root system of the trees.

Figure 6.
Adults of (A) Philaenus spumarius, (B) Philaenus italosignus, and (C) Neophilaenus campestris. P. spumarius is the main vector for Xylella fastidiosa subsp. pauca in Salento. (Reproduced from [103]).
Figure 6.
Adults of (A) Philaenus spumarius, (B) Philaenus italosignus, and (C) Neophilaenus campestris. P. spumarius is the main vector for Xylella fastidiosa subsp. pauca in Salento. (Reproduced from [103]).

Figure 7.
Philaenus spumarius life cycle related to herb and olive tree phenology. Olive phenology numbers refer to the phenological phases of olive: 11: first leaves completely separated; 65: full flowering, at least 50% of flowers open; 71: fruit size about 10% of the final size; 81: beginning of fruit coloring. 89: harvest maturity. (Reproduced from [103]).
Figure 7.
Philaenus spumarius life cycle related to herb and olive tree phenology. Olive phenology numbers refer to the phenological phases of olive: 11: first leaves completely separated; 65: full flowering, at least 50% of flowers open; 71: fruit size about 10% of the final size; 81: beginning of fruit coloring. 89: harvest maturity. (Reproduced from [103]).

Figure 8.
Situation in early autumn 2017 at San Pietro Vernotico (Brindisi province) after the eradication of about 900 olive trees that surrounded eight olive trees infected by Xylella fastidiosa subsp. pauca, carried out during autumn 2015. The trees in the figure were left in place at the border of the eradication area (in the background) since they were more than 100 m away from the infection front. In early autumn 2017, all the remaining trees resulted symptomatic and positive for X. f. subsp. pauca. The spontaneous plants not eliminated during the eradication could have acted jointly as a “reservoir” for the possible vector activity in the area for the further pathogen spread in the area.
Figure 8.
Situation in early autumn 2017 at San Pietro Vernotico (Brindisi province) after the eradication of about 900 olive trees that surrounded eight olive trees infected by Xylella fastidiosa subsp. pauca, carried out during autumn 2015. The trees in the figure were left in place at the border of the eradication area (in the background) since they were more than 100 m away from the infection front. In early autumn 2017, all the remaining trees resulted symptomatic and positive for X. f. subsp. pauca. The spontaneous plants not eliminated during the eradication could have acted jointly as a “reservoir” for the possible vector activity in the area for the further pathogen spread in the area.

Figure 9.
Olive cultivar Ogliarola salentina at Mesagne (Brindisi province) showing late symptoms caused by Neofusicoccum mediterraneum. At a first glance the symptoms can be easily confounded with those induced by Xylella fastidiosa subsp. pauca.
Figure 9.
Olive cultivar Ogliarola salentina at Mesagne (Brindisi province) showing late symptoms caused by Neofusicoccum mediterraneum. At a first glance the symptoms can be easily confounded with those induced by Xylella fastidiosa subsp. pauca.

Figure 10.
Early symptoms induced by Neofusicoccum mediterraneum (A) and Xylella fastidiosa subsp. pauca (B) on olive leaves.
Figure 10.
Early symptoms induced by Neofusicoccum mediterraneum (A) and Xylella fastidiosa subsp. pauca (B) on olive leaves.

Figure 11.
The olive cultivar Leccino is retained resistant to Xylella fastidiosa subsp. pauca, but there are examples of severe OQDS of Leccino trees in the entire infected area of Salento. Two Leccino olive groves nearby Otranto that show clear symptoms of OQDS (Lecce province).
Figure 11.
The olive cultivar Leccino is retained resistant to Xylella fastidiosa subsp. pauca, but there are examples of severe OQDS of Leccino trees in the entire infected area of Salento. Two Leccino olive groves nearby Otranto that show clear symptoms of OQDS (Lecce province).

Figure 12.
Attempts to replant olive groves in Salento with Leccino in between declining trees (A) or in place of a previous olive grove (B). The failure of the young trees is evident. Note that plowing soil along the row facilitates deeper infiltration of rainfall (B).
Figure 12.
Attempts to replant olive groves in Salento with Leccino in between declining trees (A) or in place of a previous olive grove (B). The failure of the young trees is evident. Note that plowing soil along the row facilitates deeper infiltration of rainfall (B).

Figure 13.
Resilient centennial olive trees in Gallipoli area where OQDS was first observed. Both Cellina di Nardò (A) and Ogliarola salentina (B) trees do not show any apparent symptom even though both trees resulted positive for the occurrence of Xylella fastidiosa subsp. pauca. These trees did not receive any treatment upon the infection and began to yield fruits.
Figure 13.
Resilient centennial olive trees in Gallipoli area where OQDS was first observed. Both Cellina di Nardò (A) and Ogliarola salentina (B) trees do not show any apparent symptom even though both trees resulted positive for the occurrence of Xylella fastidiosa subsp. pauca. These trees did not receive any treatment upon the infection and began to yield fruits.

Figure 14.
Olive grove trained with the local cultivars Ogliarola salentina and Cellina di Nardò at Cannole (Lecce province) (in the middle) that applies the control strategy toward Xylella fastidiosa subsp. pauca. Note the “drift effect” of the treatments on the bordering withered trees.
Figure 14.
Olive grove trained with the local cultivars Ogliarola salentina and Cellina di Nardò at Cannole (Lecce province) (in the middle) that applies the control strategy toward Xylella fastidiosa subsp. pauca. Note the “drift effect” of the treatments on the bordering withered trees.

Figure 15.
Neighboring olive groves at Nardò (Lecce province). In panel (A), during June 2019, the olive grove in the lower part of the image had the control strategy applied and the trees appear healthy, whereas the olive grove in upper part of the image had cultivation abandoned and the trees appear completely withered. Panel (B) shows the situation in June 2022. The olive grove that has continued to apply the control strategy (lower part of the image) still continues to produce oil with the local cultivars, whereas the neighboring farm replaced olive trees with tomato (on the left). In the background, other olive groves appear completely withered.
Figure 15.
Neighboring olive groves at Nardò (Lecce province). In panel (A), during June 2019, the olive grove in the lower part of the image had the control strategy applied and the trees appear healthy, whereas the olive grove in upper part of the image had cultivation abandoned and the trees appear completely withered. Panel (B) shows the situation in June 2022. The olive grove that has continued to apply the control strategy (lower part of the image) still continues to produce oil with the local cultivars, whereas the neighboring farm replaced olive trees with tomato (on the left). In the background, other olive groves appear completely withered.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Scortichini, M. The Epidemiology and Control of “Olive Quick Decline Syndrome” in Salento (Apulia, Italy). Agronomy 2022, 12, 2475. https://doi.org/10.3390/agronomy12102475
AMA Style
Scortichini M. The Epidemiology and Control of “Olive Quick Decline Syndrome” in Salento (Apulia, Italy). Agronomy. 2022; 12(10):2475. https://doi.org/10.3390/agronomy12102475
Chicago/Turabian StyleScortichini, Marco. 2022. "The Epidemiology and Control of “Olive Quick Decline Syndrome” in Salento (Apulia, Italy)" Agronomy 12, no. 10: 2475. https://doi.org/10.3390/agronomy12102475
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.