
Melatonin Pretreatment Alleviated Inhibitory Effects of Drought Stress by Enhancing Anti-Oxidant Activities and Accumulation of Higher Proline and Plant Pigments and Improving Maize Productivity

 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Experimental Treatments
2.3. Sampling and Measurement
2.3.1. Determination of Plant Leaf Water Relations
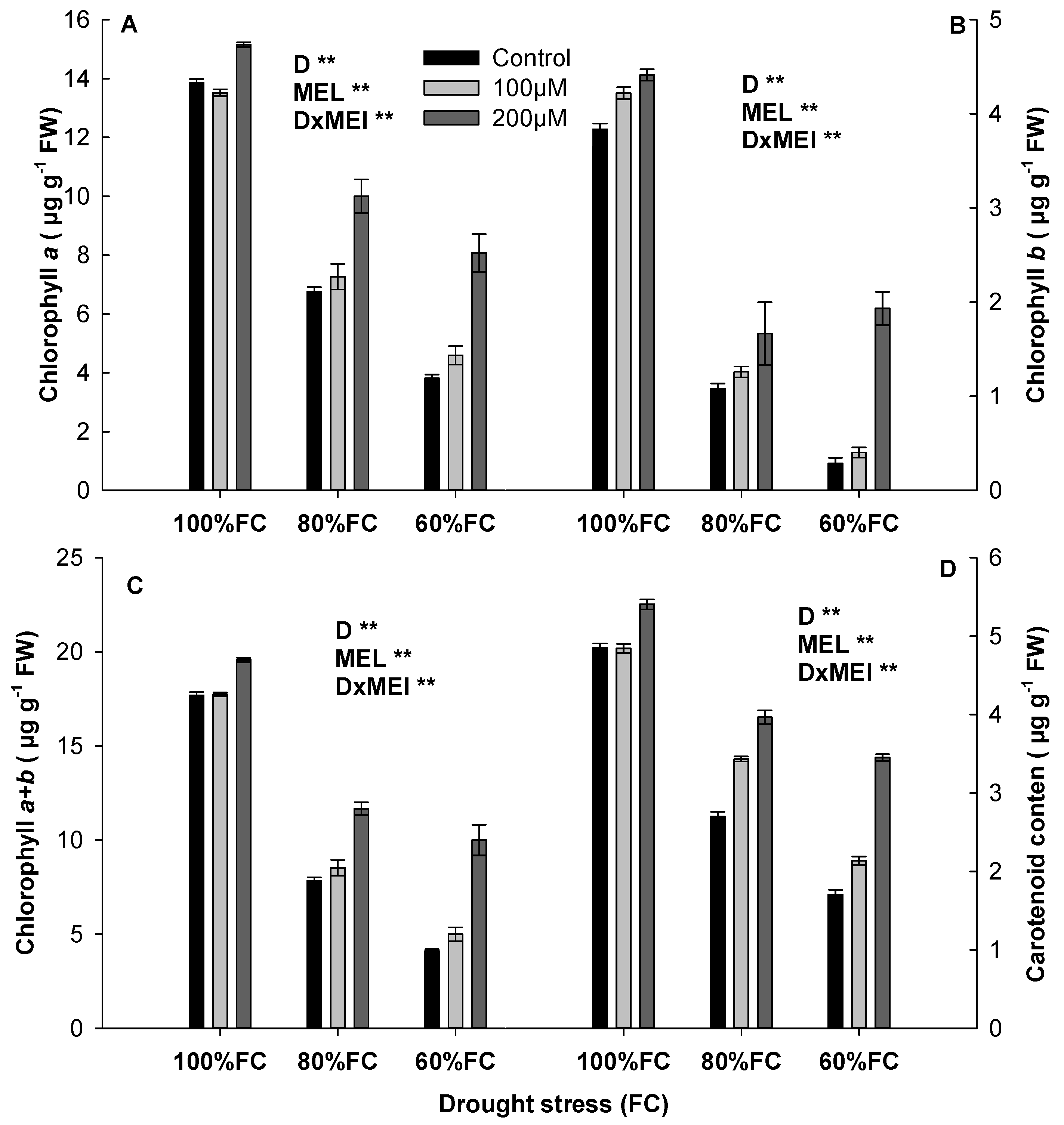
2.3.2. Chlorophyll and Carotenoids Determination
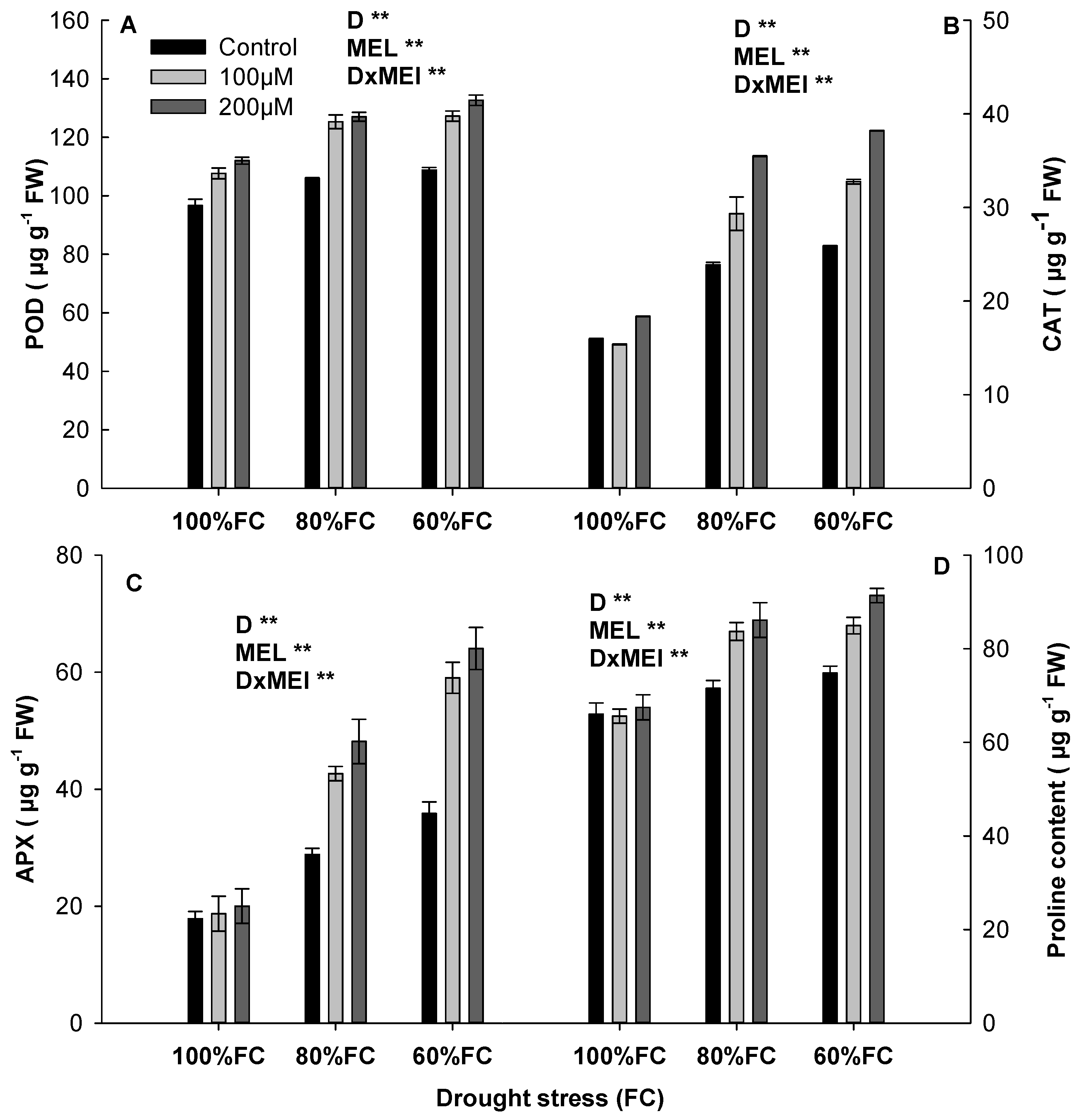
2.3.3. Determination of Anti-Oxidant Enzyme Activity
2.3.4. Proline Measurement
2.3.5. Plant Growth Attributes
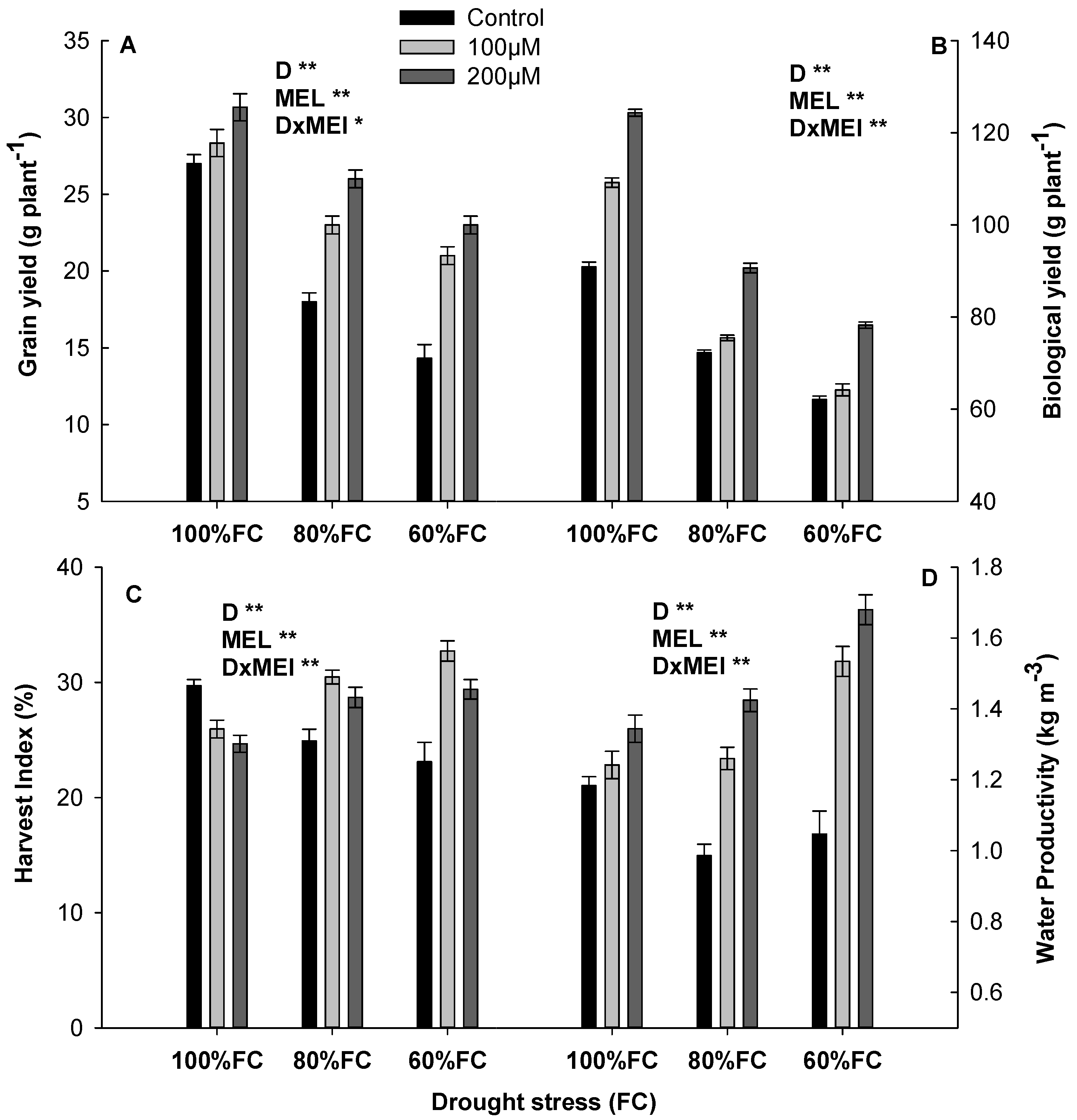
2.3.6. Yield and Yield Components
2.4. Statistical Analysis
3. Results
3.1. Effect of Melatonin on Plant Leaf Water Relations
3.2. Effect of Melatonin on Plant Pigment
3.3. Response of MEL to Anti-Oxidant Activities and Proline Content
3.4. Effect of MEL on Maize Growth Traits during Drought Stress
3.5. Yield Components with MEL Application under Drought Stress
3.6. Maize Yields and Water Productivity with MEL under Drought Stress
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liang, G.; Liu, J.; Zhang, J.; Guo, J. Effects of Drought Stress on Photosynthetic and Physiological Parameters of Tomato. J. Am. Soc. Hortic. Sci. 2020, 145, 12–17. [Google Scholar] [CrossRef] [Green Version]
- Cui, G.; Zhao, X.; Liu, S.; Sun, F.; Zhang, C.; Xi, Y. Beneficial Effects of Melatonin in Overcoming Drought Stress in Wheat Seedlings. Plant Physiol. Biochem. 2017, 118, 138–149. [Google Scholar] [CrossRef]
- Naeem, M.; Naeem, M.S.; Ahmad, R.; Ahmad, R.; Ashraf, M.Y.; Ihsan, M.Z.; Nawaz, F.; Athar, H.u.R.; Ashraf, M.; Abbas, H.T.; et al. Improving Drought Tolerance in Maize by Foliar Application of Boron: Water Status, Antioxidative Defense and Photosynthetic Capacity. Arch. Agron. Soil Sci. 2018, 64, 626–639. [Google Scholar] [CrossRef]
- Zhao, Z.-Y.; Wang, P.-Y.; Xiong, X.-B.; Wang, Y.-B.; Zhou, R.; Tao, H.-Y.; Grace, U.A.; Wang, N.; Xiong, Y.-C. Environmental risk of multi-year polythene film mulching and its green solution in arid irrigation region. J. Hazard. Mater. 2022, 435, 128981. [Google Scholar] [CrossRef]
- Sunaina, B.; Kumar, J.R.; Rupak, K.; Mahesh, R. A Case Study on Soil Fertility Status and Maize Productivity in Dang District, Nepal. Malays. J. Sustain. Agric. 2019, 3, 56–59. [Google Scholar] [CrossRef]
- Ma, S.; Wang, Z.; Guo, X.; Wang, F.; Huang, J.; Sun, B.; Wang, X. Sourdough improves the quality of whole-wheat flour products: Mechanisms and challenges—A review. Food Chem. 2021, 360, 130038. [Google Scholar] [CrossRef]
- Vijayaraghavareddy, P.; Lekshmy, S.V.; Struik, P.C.; Makarla, U.; Yin, X.; Sreeman, S. Production and scavenging of reactive oxygen species confer to differential sensitivity of rice and wheat to drought stress. Crop Environ. 2022, 1, 15–23. [Google Scholar] [CrossRef]
- Dawood, M.G.; El-Awadi, M.E. Disminución Del Estrés Salino En Plantas de Vicia Faba L. a Través de La Activación de Las Semillas Con Melatonina. Acta Biol. Colomb. 2015, 20, 223–235. [Google Scholar] [CrossRef]
- Shi, H.; Chen, K.; Wei, Y.; He, C. Fundamental Issues of Melatonin-Mediated Stress Signaling in Plants. Front. Plant Sci. 2016, 7, 1124. [Google Scholar] [CrossRef] [Green Version]
- Imran, M.; Latif Khan, A.; Shahzad, R.; Aaqil Khan, M.; Bilal, S.; Khan, A.; Kang, S.M.; Lee, I.J. Exogenous Melatonin Induces Drought Stress Tolerance by Promoting Plant Growth and Antioxidant Defence System of Soybean Plants. AoB Plants 2021, 13, plab026. [Google Scholar] [CrossRef]
- Normah, H.; Hanapi, M.J. Antioxidant Capacity of The Green Leafy Vegetables Using Oxygen Radical Antioxidant Capacity (Orac), 2,2’-Azino-Bis (3-Ethylbenzothiazoline-6-Sulphonic Acid(Abts) And 2,2-Diphenyl-1-Picrylhydrazyl (Dpph) Assays. Sci. Herit. J. 2019, 3, 1–7. [Google Scholar]
- Shams ul Hassan, S.; Jin, H.z.; Abu-Izneid, T.; Rauf, A.; Ishaq, M.; Suleria, H.A.R. Stress-driven discovery in the natural products: A gateway towards new drugs. Biomed. Pharmacother. 2019, 109, 459–467. [Google Scholar] [CrossRef] [PubMed]
- Ayliffe, M.; Periyannan, S.K.; Feechan, A.; Dry, I.; Schumann, U.; Wang, M.B.; Pryor, A.; and Lagudah, E. A simple method for comparing fungal biomass in infected plant tissues. Mol. Plant-Microbe Interact. 2013, 26, 658–667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarropoulou, V.; Dimassi-Theriou, K.; Therios, I.; Koukourikou-Petridou, M. Melatonin Enhances Root Regeneration, Photosynthetic Pigments, Biomass, Total Carbohydrates and Proline Content in the Cherry Rootstock PHL-C (Prunus Avium × Prunus Cerasus). Plant Physiol. Biochem. 2012, 61, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Tan, D.-X.; Sharma, R. Historical Perspective and Evaluation of the Mechanisms by Which Melatonin Mediates Seasonal Reproduction in Mammals. Melatonin Res. 2018, 1, 59–77. [Google Scholar] [CrossRef]
- Martinez, V.; Nieves-Cordones, M.; Lopez-Delacalle, M.; Rodenas, R.; Mestre, T.C.; Garcia-Sanchez, F.; Rubio, F.; Nortes, P.A.; Mittler, R.; Rivero, R.M. Tolerance to Stress Combination in Tomato Plants: New Insights in the Protective Role of Melatonin. Molecules 2018, 23, 535. [Google Scholar] [CrossRef] [Green Version]
- Jiang, D.; Lu, B.; Liu, L.; Duan, W.; Chen, L.; Li, J.; Zhang, K.; Sun, H.; Zhang, Y.; Dong, H.; et al. Exogenous Melatonin Improves Salt Stress Adaptation of Cotton Seedlings by Regulating Active Oxygen Metabolism. PeerJ 2020, 8, e10486. [Google Scholar] [CrossRef]
- Tan, D.-X.; Manchester, L.C.; Esteban-Zubero, E.; Zhou, Z.; Reiter, R.J. Melatonin as a Potent and Inducible Endogenous Anti-oxidant: Synthesis and Metabolism. Molecules 2015, 20, 18886–18906. [Google Scholar] [CrossRef] [Green Version]
- Florido, J.; Rodriguez-Santana, C.; Martinez-Ruiz, L.; López-Rodríguez, A.; Acuña-Castroviejo, D.; Rusanova, I.; Escames, G. Understanding the Mechanism of Action of Melatonin, Which Induces ROS Production in Cancer Cells. Antioxidants 2022, 11, 1621. [Google Scholar] [CrossRef]
- Ahmad, S.; Kamran, M.; Ding, R.; Meng, X.; Wang, H.; Ahmad, I.; Fahad, S.; Han, Q. Exogenous Melatonin Confers Drought Stress by Promoting Plant Growth, Photosynthetic Capacity and Antioxidant Defense System of Maize Seedlings. PeerJ 2019, 7, e7793. [Google Scholar] [CrossRef] [Green Version]
- Fleta-Soriano, E.; Díaz, L.; Bonet, E.; Munné-Bosch, S. Melatonin May Exert a Protective Role against Drought Stress in Maize. J. Agron. Crop Sci. 2017, 203, 286–294. [Google Scholar] [CrossRef]
- Nawaz, K.; Chaudhary, R.; Sarwar, A.; Ahmad, B.; Gul, A.; Hano, C.; Abbasi, B.H.; Anjum, S. Melatonin as Master Regulator in Plant Growth, Development and Stress Alleviator for Sustainable Agricultural Production: Current Status and Future Perspectives. Sustainability 2021, 13, 294. [Google Scholar] [CrossRef]
- Li, H.; Chang, J.; Chen, H.; Wang, Z.; Gu, X.; Wei, C.; Zhang, Y.; Ma, J.; Yang, J.; Zhang, X. Exogenous Melatonin Confers Salt Stress Tolerance to Watermelon by Improving Photosynthesis and Redox Homeostasis. Front. Plant Sci. 2017, 8, 295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, D.; Ni, Z.; Xia, H.; Xie, Y.; Lv, X.; Wang, J.; Lin, L.; Deng, Q.; Luo, X. Exogenous Melatonin Promotes Biomass Accumulation and Photosynthesis of Kiwifruit Seedlings under Drought Stress. Sci. Hortic. 2019, 246, 34–43. [Google Scholar] [CrossRef]
- Lee, H.Y.; Back, K. Melatonin Plays a Pivotal Role in Conferring Tolerance against Endoplasmic Reticulum Stress via Mitogen-Activated Protein Kinases and BZIP60 in Arabidopsis Thaliana. Melatonin Res. 2018, 1, 94–108. [Google Scholar] [CrossRef] [Green Version]
- Turk, H.; Erdal, S.; Genisel, M.; Atici, O.; Demir, Y.; Yanmis, D. The Regulatory Effect of Melatonin on Physiological, Biochemical and Molecular Parameters in Cold-Stressed Wheat Seedlings. Plant Growth Regul. 2014, 74, 139–152. [Google Scholar] [CrossRef]
- Arnon, D.I. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Cakmak, I.; Marschner, H. Magnesium Deficiency and High Light Intensity Enhance Activities of Superoxide Dismutase, Ascorbate Peroxidase, and Glutathione Reductase in Bean Leaves. Plant Physiol. 1992, 98, 1222–1227. [Google Scholar] [CrossRef] [Green Version]
- Nakano, Y.; Asada, K. Hydrogen Peroxide Is Scavenged by Ascorbate-Specific Peroxidase in Spinach Chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar] [CrossRef]
- Havir, E.A.; McHale, N.A. Biochemical and Developmental Characterization of Multiple Forms of Catalase in Tobacco Leaves. Plant Physiol. 1987, 84, 450–455. [Google Scholar] [CrossRef] [Green Version]
- Tiwari, J.K.; Munshi, A.D.; Kumar, R.; Pandey, R.N.; Arora, A.; Bhat, J.S.; Sureja, A.K. Effect of Salt Stress on Cucumber: Na+-K+ Ratio, Osmolyte Concentration, Phenols and Chlorophyll Content. Acta Physiol. Plant. 2010, 32, 103–114. [Google Scholar] [CrossRef]
- Hossain, M.A.; Bhattacharjee, S.; Armin, S.M.; Qian, P.; Xin, W.; Li, H.Y.; Burritt, D.J.; Fujita, M.; Tran, L.S.P. Hydrogen Peroxide Priming Modulates Abiotic Oxidative Stress Tolerance: Insights from ROS Detoxification and Scavenging. Front. Plant Sci. 2015, 6, 420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Webber, H.; Ewert, F.; Olesen, J.E.; Müller, C.; Fronzek, S.; Ruane, A.C.; Bourgault, M.; Martre, P.; Ababaei, B.; Bindi, M.; et al. Diverging Importance of Drought Stress for Maize and Winter Wheat in Europe. Nat. Commun. 2018, 9, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, G.; Liu, X.; Cui, Z. Achieving global food security by focusing on nitrogen efficiency potentials and local production. Glob. Food Secur. 2021, 29, 100536. [Google Scholar] [CrossRef]
- Li, W.; Shi, Y.; Zhu, D.; Wang, W.; Liu, H.; Li, J.; Shi, N.; Ma, L.; Fu, S. Fine root biomass and morphology in a temperate forest are influenced more by the nitrogen treatment approach than the rate. Ecol. Indic. 2021, 130, 108031. [Google Scholar] [CrossRef]
- Hassan, S.S.; Shah, S.A.A.; Pan, C.; Fu, L.; Cao, X.; Shi, Y.; Wu, X.; Wang, K.; Wu, B. Production of an antibiotic enterocin from a marine actinobacteria strain H1003 by metal-stress technique with enhanced enrichment using response surface methodology. Pak. J. Pharm. Sci. 2017, 30, 313–324. [Google Scholar]
- Samuel, A.D.; Bungau, S.; Fodor, I.K.; Tit, D.M.; Blidar, C.F.; David, A.T.; Melinte (Frunzulica), C.E. Effects of liming and fertilization on the dehydrogenase and catalase activities. Rev. Chim. 2019, 70, 3464–3468. [Google Scholar] [CrossRef]
- Samuel, A.D.; Brejea, R.; Domuta, C.; Bungau, S.; Cenusa, N.; Tit, D.M. Enzymatic indicators of soil quality. J. Environ. Prot. Ecol. 2017, 18, 871–878. [Google Scholar]
- Ul Hassan, S.S.; Muhammad, I.; Abbas, S.Q.; Hassan, M.; Majid, M.; Jin, H.Z.; Bungau, S. Stress driven discovery of natural products from actinobacteria with anti-oxidant and cytotoxic activities including docking and admet properties. Int. J. Mol. Sci. 2021, 22, 11432. [Google Scholar] [CrossRef]
- Yamauchi, Y. Chapter 7: Integrated Chemical Control of Abiotic Stress Tolerance Using Biostimulants. In Plant, Abiotic Stress Responses to Climate Change; Intech Open: London, UK, 2018. [Google Scholar] [CrossRef] [Green Version]
- Emiliani, J.; D’Andrea, L.; Falcone Ferreyra, M.L.; Maulión, E.; Rodriguez, E.; Rodriguez-Concepción, M.; Casati, P. A Role for β,β-Xanthophylls in Arabidopsis UV-B Photoprotection. J. Exp. Bot. 2018, 69, 4921–4933. [Google Scholar] [CrossRef]
- Choudhury, F.K.; Rivero, R.M.; Blumwald, E.; Mittler, R. Reactive oxygen species, abiotic stress and stress combination. Plant J. 2017, 90, 856–867. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; He, C. Regulation of Plant Reactive Oxygen Species (ROS) in Stress Responses: Learning from AtRBOHD. Plant Cell Rep. 2016, 35, 995–1007. [Google Scholar] [CrossRef]
- Singh, A.; Singh, N.B.; Hussain, I.; Singh, H. Effect of biologically synthesized copper oxide nanoparticles on metabolism and antioxidant activity to the crop plants Solanum lycopersicum and Brassica oleracea var. botrytis. J. Biotechnol. 2017, 262, 11–27. [Google Scholar] [CrossRef] [PubMed]
- Regier, N.; Cosio, C.; von Moos, N.; Slaveykova, V.I. Effects of Copper-Oxide Nanoparticles, Dissolved Copper and Ultraviolet Radiation on Copper Bioaccumulation, Photosynthesis and Oxidative Stress in the Aquatic Macrophyte Elodea Nuttallii. Chemosphere 2015, 128, 56–61. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Su, X.; Chen, Y.; Fan, X.; He, L.; Guo, J.; Wang, Y.; Yang, Q. Melatonin Improves Drought Resistance in Maize Seedlings by Enhancing the Anti-oxidant System and Regulating Abscisic Acid Metabolism to Maintain Stomatal Opening Under PEG-Induced Drought. J. Plant Biol. 2021, 64, 299–312. [Google Scholar] [CrossRef]
- Samuel, A.D.; Tit, D.M.; Melinte (Frunzulica), C.E.; Iovan, C.; Purza, L.; Gitea, M.; Bungau, S. Enzymological and physicochemical evaluation of the effects of soil management practices. Rev. Chim. 2017, 68, 2243–2247. [Google Scholar] [CrossRef]
- Bungau, S.; Behl, T.; Aleya, l.; Bourgeade, P.; Aloui-Sosse, B.; Purza, A.L.; Abid, A.; Samuel, A.D. Expatiating the impact of anthropogenic aspects and climatic factors on long term soil monitoring and management. Environ Sci. Pollut. Res. 2021, 202, 30528–30550. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Drought Stress | MEL | RWC (%) | WUC | WSD(%) | RSD(%) | LWC(%) |
|---|---|---|---|---|---|---|
| 100% FC (No Stress) | Control | 88.6 a | 0.24d | 11.4 f | 5.02 e | 41.7 d |
| 100 µM | 87.6 a | 0.19d | 12.4f | 7.53 d | 57.3 c | |
| 200 µM | 89.0 a | 0.09d | 11.0 f | 7.55 d | 65.8 b | |
| 80% FC (Mild stress) | Control | 80.6 bc | 0.66c | 19.4 de | 7.99 d | 36.5 d |
| 100 µM | 79.0 c | 0.32d | 21.0 d | 12.57 c | 54.2c | |
| 200 µM | 82.1 b | 0.14d | 17.9 e | 13.68 c | 73.2 b | |
| 60% FC (severe stress) | Control | 62.3 f | 1.71a | 37.7 a | 17.60 b | 35.2 d |
| 100 µM | 67.8 e | 1.11b | 32.2 b | 24.90 a | 69.8 b | |
| 200 µM | 73.0 d | 0.34d | 27.0 c | 23.25 a | 82.0 a | |
| ‡SOV | DS | ** | ** | ** | ** | ** |
| MEL | ** | ** | ** | ** | ** | |
| DS × MEL | ** | ** | ** | * | ** |
| Drought Stress | Mel | Leaves Number | Stem Diameter | Plant Height | Leaf Area |
|---|---|---|---|---|---|
| 100%FC | Control | 10.2 ab(±0.6) | 1.1b–d (±0.06) | 52.5 a (±1) | 146.9 c (±7.9) |
| 100µM | 10.5 ab (±0.3) | 1.2a–c (±0.06) | 48.3 ab (±1.5) | 149.2 bc (±3.1) | |
| 200µM | 10.5 ab(± 0.3) | 1.2ab (±0.07) | 47.0 a–c (±2.2) | 163.6 ab (±4.6) | |
| 80%FC | Control | 10.0 ab (±0.3) | 1.1b–d (±0.04) | 43.2 b–d(±0.0) | 128.7 de(±2.2) |
| 100µM | 11.0 a (±0.6) | 1.2abc (±0.04) | 45.1 ac(±1.8) | 159.9 bc (±0.5) | |
| 200µM | 11.0 a (±0.6) | 1.3a (±0.03) | 52.1 a(±2.2) | 175.4 a (±7.1) | |
| 60%FC | Control | 8.0 d (±0.6) | 1.0d (±0.03) | 36.8 e(±0.7) | 102.3 f (±6.3) |
| 100µM | 9.0 c(±0.01) | 1.1cd (±0.03) | 41.9 c–e(±3.7) | 114.8 ef (±5.5) | |
| 200µM | 9.5 bc(±0.3) | 1.1cd (±0.0 3) | 38.1 de(±2.9) | 129.7 d (±1.0) | |
| ‡SOV | DS | ** | ** | ** | ** |
| MEL | ** | ** | ** | ** | |
| DS×MEL | ** | ** | ** | * | |
| Drought stress | Mel | Root Weight | Stem Weight | Leaf Weight | dry Matter (g plant−1) |
| 100% FC | Control | 7.5 c (± 0.3) | 21.9 ab (±4.6) | 21.0 bcd(±0.1) | 50.4 a (±8.7) |
| 100 µM | 8.0 c (±0.2) | 23.5 ab (±1.6) | 24.8 ab (±2.1) | 56.3 a (±7.0) | |
| 200 µM | 7.2 c (± 0.2) | 19.5 abc (±2.5) | 22.5 bc (±1.9) | 49.2 a (±1.3) | |
| 80% FC | Control | 5.9 d (±0.1) | 17.9 bcd (±1.2) | 19.3 cd (±1.1) | 43.1 b (±4.0) |
| 100 µM | 9.6 b(±0.3) | 22.3 ab (±4.4) | 25.6 ab (±3.1) | 57.5 a (±10.3) | |
| 200 µM | 7.2 c (± 0.6) | 26.6 a (±2.0) | 26.5 a (±2.3) | 60.3 a (±5.8) | |
| 60% FC | Control | 6.8 c (± 0.3) | 11.4 d (±1.6) | 13.4 e (±0.1) | 31.6 b (±5.3) |
| 100 µM | 7.2 c (±0.1) | 13.9 cd (±1.8) | 15.7 d (±1.2) | 36.8 b (±5.1) | |
| 200 µM | 7.6 c (±0.3) | 12.5 cd (±0.1) | 15.9 d (±1.7) | 36.0 b (±2.2) | |
| ‡SOV | DS | ** | ** | ** | ** |
| MEL | ** | ** | NS | ** | |
| DS × MEL | ** | ** | ** | * |
| Drought stress | MEL | Grains Ear−1 | Ear Weight | Thousand-Grain Weight (g) |
|---|---|---|---|---|
| 100% FC | Control | 75.0 (±1.16) | 57.7 (±0.48) | 241.9 (±4.2) |
| 100 µM | 99.7 (±0.89) | 58.3 (±0.53) | 245.5 (±4.8) | |
| 200 µM | 144.7 (0.33) | 57.2 (±65) | 249.3 (±6.2) | |
| 80% FC | Control | 80.3 (±0.43) | 50.5 (±0.32) | 217.2 (±4.8) |
| 100 µM | 109.3 (±0.83) | 52.7 (±0.91) | 223.0 (±1.3) | |
| 200 µM | 127.3 (±1.02) | 53.4 (±0.55) | 252.1 (±5.6) | |
| 60% FC | Control | 23.7 (±1.86) | 44.5 (±0.52) | 203.4 (±4.9) |
| 100 µM | 35.0 (±0.90) | 42.6 (±0.49) | 212.7 (±3.8) | |
| 200 µM | 61.7 (±0.53) | 40.3 (±0.71) | 252.0 (±4.4) | |
| ‡SOV | DS | ** | ** | ** |
| MEL | ** | ns | ** | |
| DS × MEL | ** | ns | ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gul, N.; Haq, Z.U.; Ali, H.; Munsif, F.; Hassan, S.S.u.; Bungau, S. Melatonin Pretreatment Alleviated Inhibitory Effects of Drought Stress by Enhancing Anti-Oxidant Activities and Accumulation of Higher Proline and Plant Pigments and Improving Maize Productivity. Agronomy 2022, 12, 2398. https://doi.org/10.3390/agronomy12102398
Gul N, Haq ZU, Ali H, Munsif F, Hassan SSu, Bungau S. Melatonin Pretreatment Alleviated Inhibitory Effects of Drought Stress by Enhancing Anti-Oxidant Activities and Accumulation of Higher Proline and Plant Pigments and Improving Maize Productivity. Agronomy. 2022; 12(10):2398. https://doi.org/10.3390/agronomy12102398
Chicago/Turabian StyleGul, Nasib, Zia Ul Haq, Hina Ali, Fazal Munsif, Syed Shams ul Hassan, and Simona Bungau. 2022. "Melatonin Pretreatment Alleviated Inhibitory Effects of Drought Stress by Enhancing Anti-Oxidant Activities and Accumulation of Higher Proline and Plant Pigments and Improving Maize Productivity" Agronomy 12, no. 10: 2398. https://doi.org/10.3390/agronomy12102398