
UAV- and Random-Forest-AdaBoost (RFA)-Based Estimation of Rice Plant Traits

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Sites and Design
2.2. Field Data Collection
2.2.1. Ground Data
2.2.2. Spectral Data
2.3. Image Pre-Processing
2.4. Data Processing
2.4.1. Raw and Indices Calculation and Extraction
2.4.2. Machine Learning Algorithm
Random Forest-AdaBoost (RFA) Regressor
2.5. Performance Evaluation
3. Results
3.1. Descriptive Statistics of Plant Traits
3.2. Relationships between Multispectral Images and Plant Traits
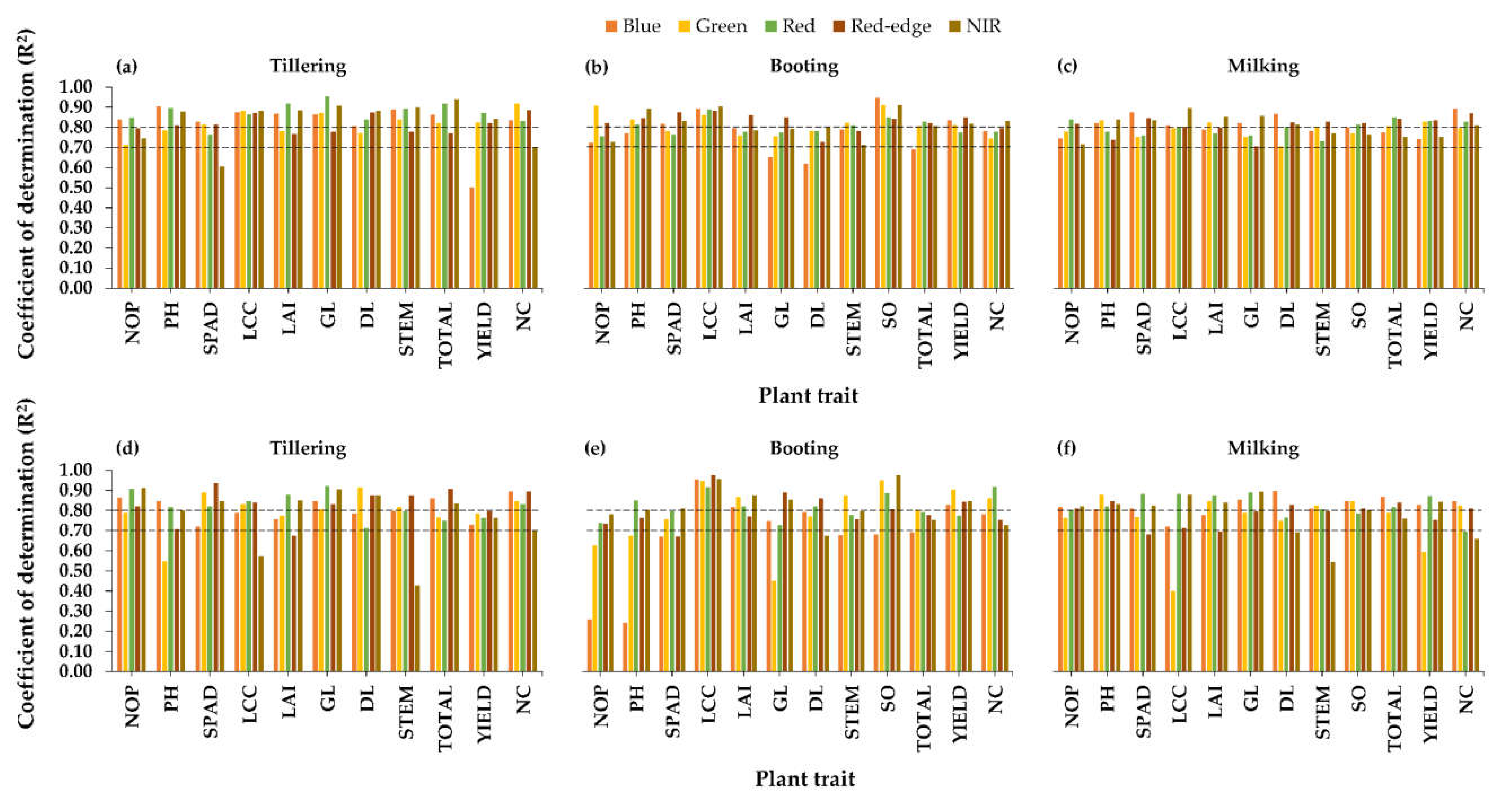
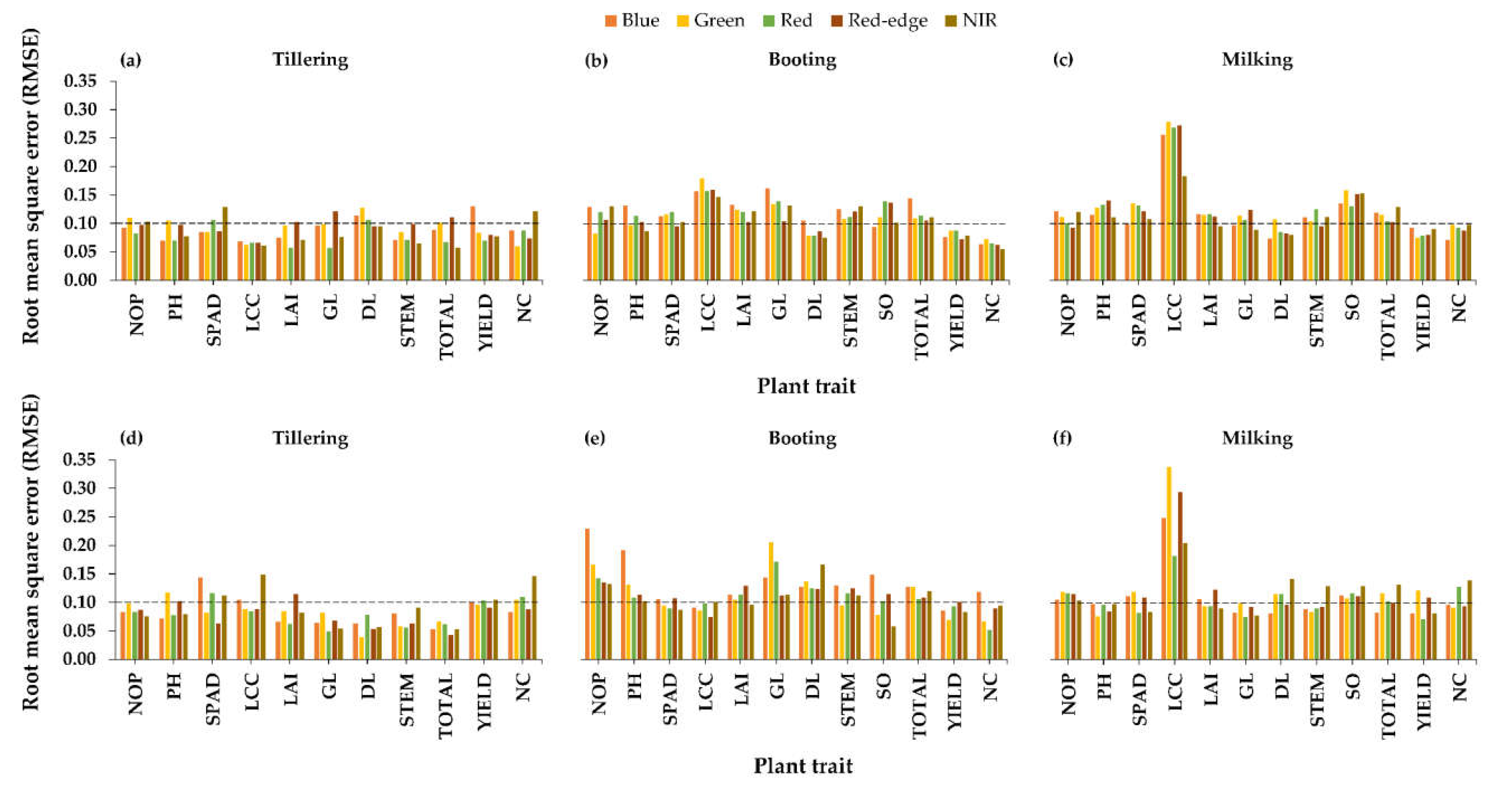
3.2.1. Relationships between Raw Bands and Plant Traits
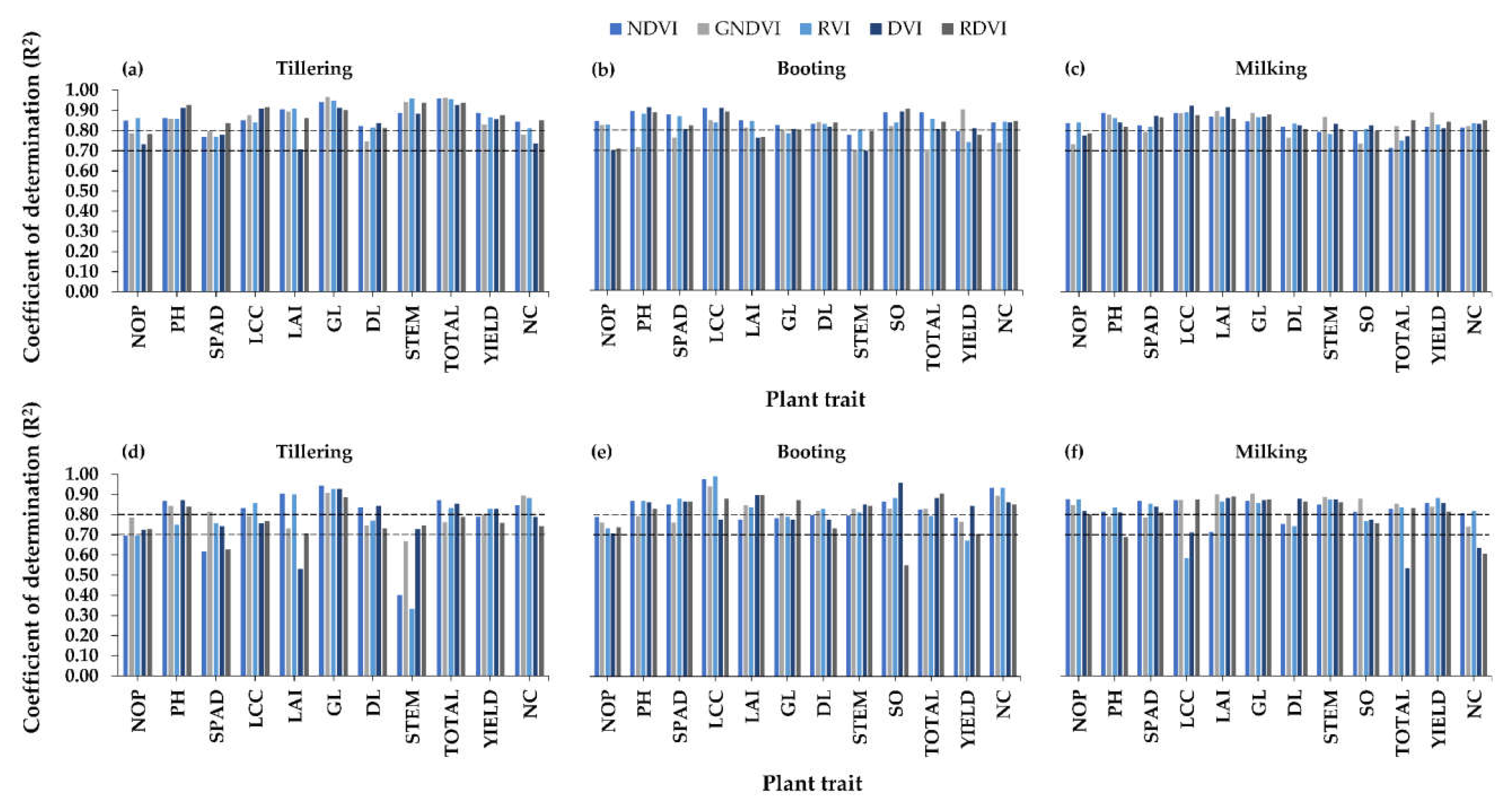
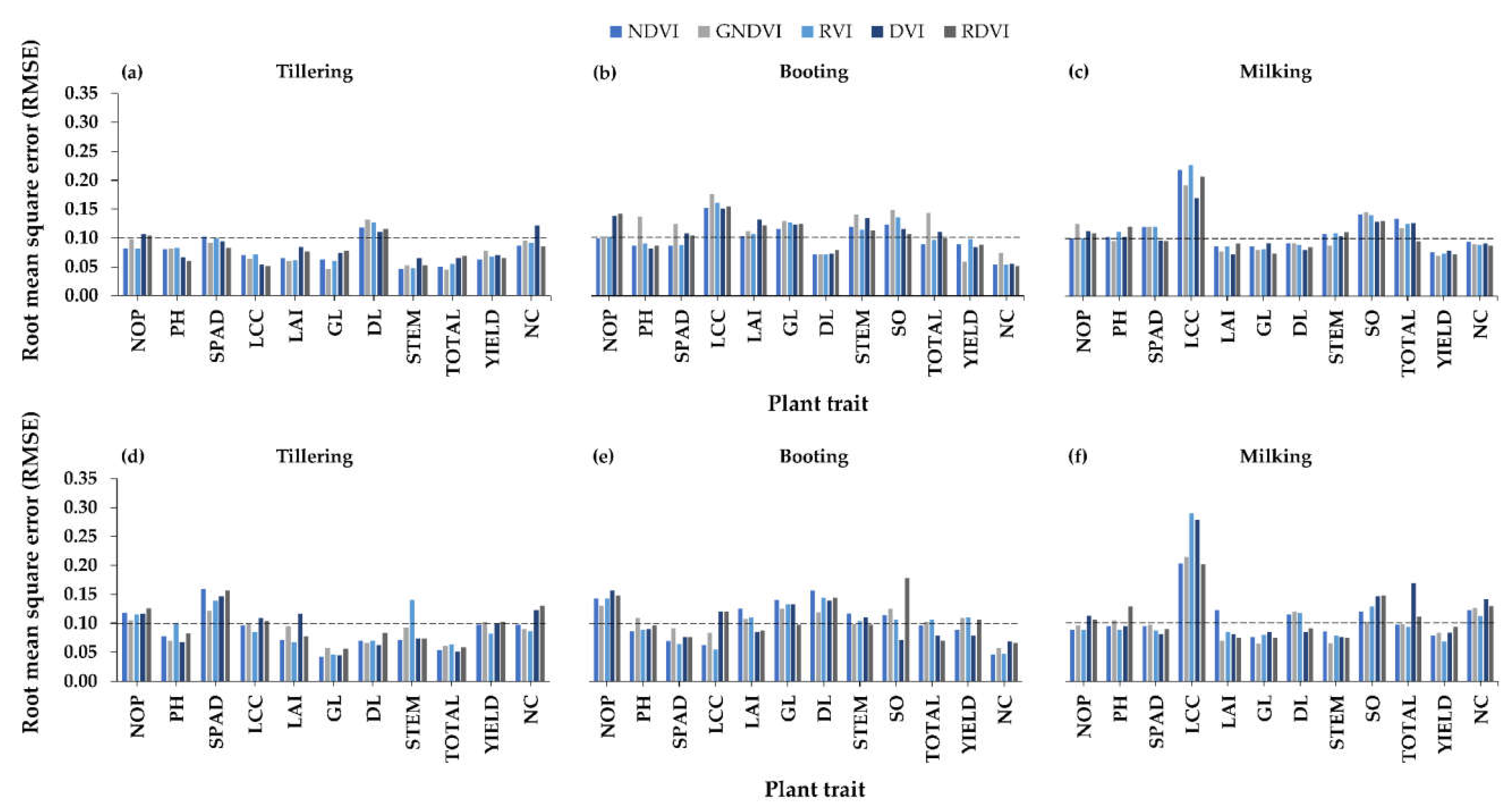
3.2.2. Relationships between Visible-NIR Indices and Plant Traits
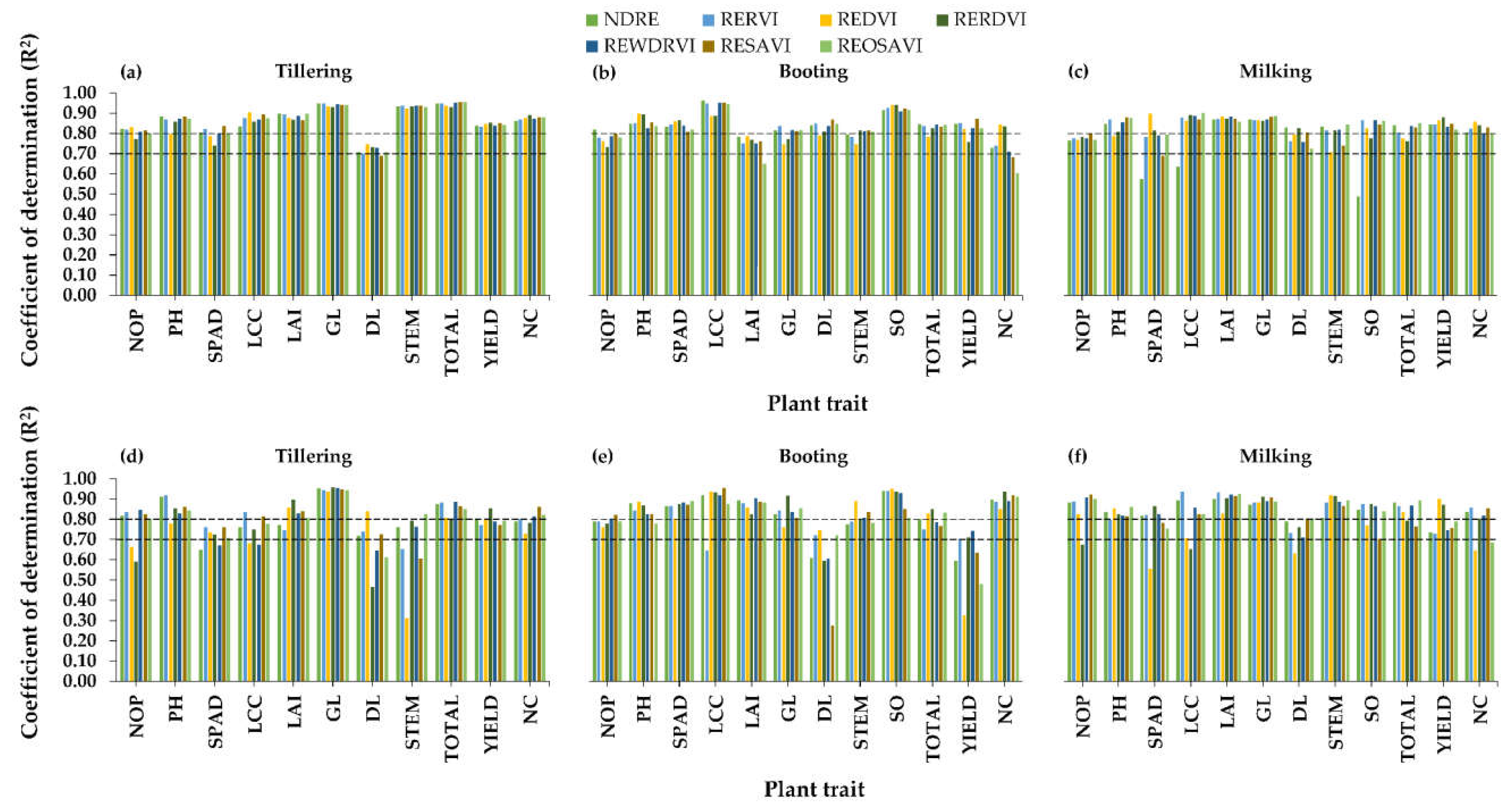
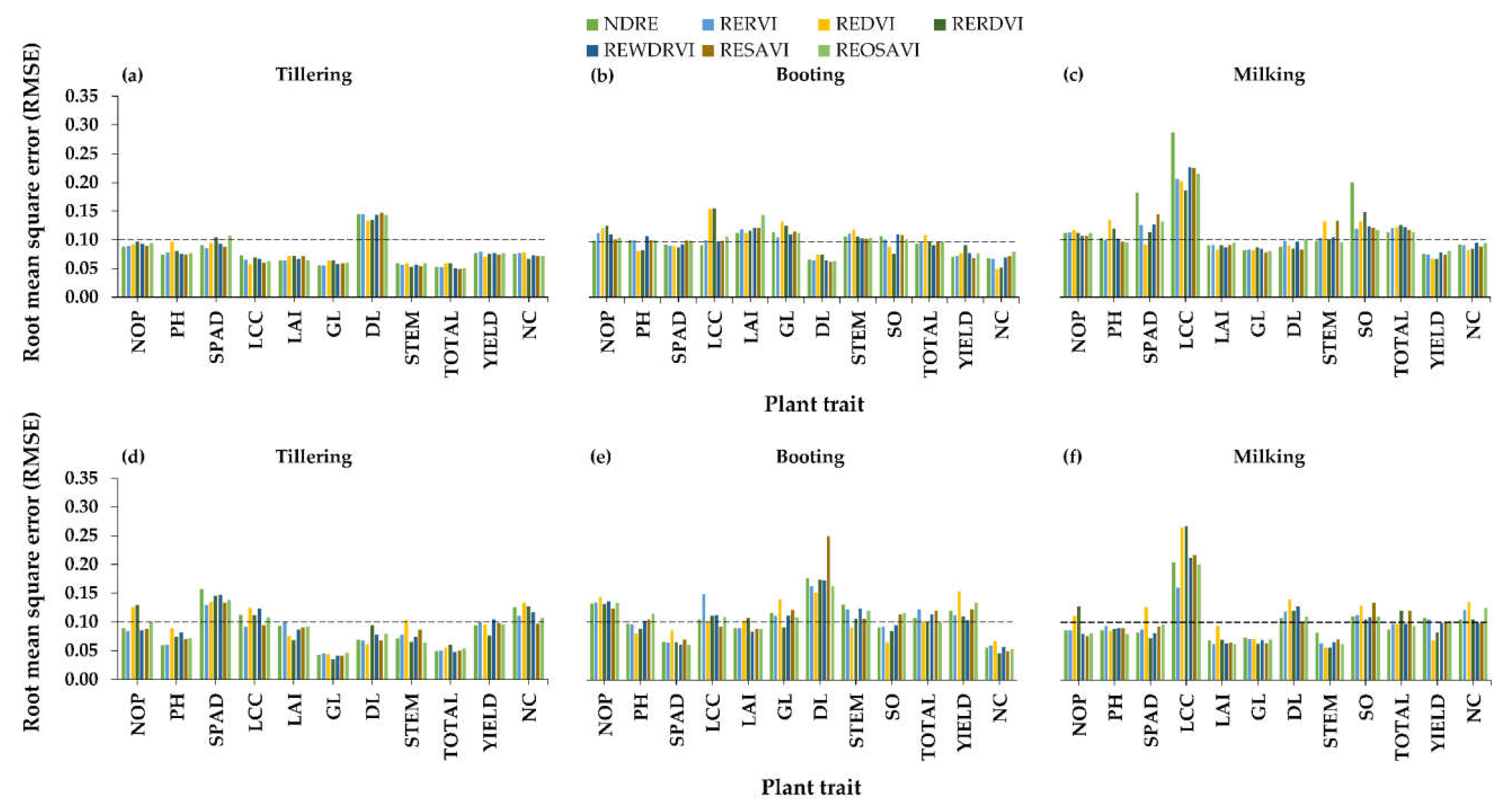
3.2.3. Relationships between Red-Edge-NIR Indices and Plant Traits
3.3. Performance of the Random Forest-AdaBoost Algorithms
4. Discussion
4.1. Performance of Plant Traits Estimations
4.2. Best Vegetation Indices for Plant Traits Estimations
4.3. Best Rice Growth Stage for Plant Trait Estimations
4.4. Performance of the Random Forest-AdaBoost Algorithms
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kefauver, S.C.; Vicente, R.; Vergara-Díaz, O.; Fernandez-Gallego, J.A.; Kerfal, S.; Lopez, A.; Melichar, J.P.; Serret Molins, M.D.; Araus, J.L. Comparative UAV and field phenotyping to assess yield and nitrogen use efficiency in hybrid and conventional barley. Front. Plant Sci. 2017, 8, 1733. [Google Scholar] [CrossRef] [PubMed]
- Tattaris, M.; Reynolds, M.P.; Chapman, S.C. A direct comparison of remote sensing approaches for high-throughput phenotyping in plant breeding. Front. Plant Sci. 2017, 7, 1131. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Liu, J.; Zhao, C.; Li, Z.; Huang, Y.; Yu, H.; Xu, B.; Yang, X.; Zhu, D.; Zhang, X.; et al. Unmanned aerial vehicle remote sensing for field-based crop phenotyping: Current status and perspectives. Front. Plant Sci. 2017, 8, 1111. [Google Scholar] [CrossRef]
- Watanabe, K.; Guo, W.; Arai, K.; Takanashi, H.; Kajiya-Kanegae, H.; Kobayashi, M.; Yano, K.; Tokunaga, T.; Fujiwara, T.; Tsutsumi, N.; et al. High-throughput phenotyping of sorghum plant height using an unmanned aerial vehicle and its application to genomic prediction modeling. Front. Plant Sci. 2017, 8, 421. [Google Scholar] [CrossRef] [Green Version]
- Chawade, A.; van Ham, J.; Blomquist, H.; Bagge, O.; Alexandersson, E.; Ortiz, R. High-throughput field-phynotyping tools for plant breeding and precision agriculture. Agronomy 2019, 9, 258. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.; Miao, Y.; Huang, Y.; Shi, W.; Hu, X.; Wang, X.; Wan, J. Evaluating an Unmanned Aerial Vehicle-Based Remote Sensing System for Estimation of Rice Nitrogen Status. In Proceedings of the 4th International Conference on Agro-Geoinformatics (Agro-geoinformatics), Istanbul, Turkey, 20–24 July 2015; pp. 198–203. [Google Scholar] [CrossRef]
- Zheng, H.; Cheng, T.; Li, D.; Zhou, X.; Yao, X.; Tian, Y.; Cao, W.; Zhu, Y. Evaluation of RGB, color-infrared and multispectral images acquired from unmanned aerial systems for the estimation of nitrogen accumulation in rice. Remote Sens. 2018, 10, 824. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Ding, X.; Kuang, Q.; Ata-UI-Karim, S.T.; Cheng, T.; Liu, X.; Tian, Y.; Zhu, Y.; Cao, W.; Cao, Q. Potential of UAV-based active sensing for monitoring rice leaf nitrogen status. Front. Plant Sci. 2018, 14, 1834. [Google Scholar] [CrossRef] [Green Version]
- Zheng, H.; Cheng, T.; Zhou, M.; Li, D.; Yao, X.; Tian, Y.; Cao, W.; Zhu, Y. Improved estimation of rice aboveground biomass combining textural and spectral analysis of UAV imagery. Precis. Agric. 2019, 20, 611. [Google Scholar] [CrossRef]
- Cen, H.; Wan, L.; Zhu, J.; Li, Y.; Li, X.; Zhu, Y.; Weng, H.; Wu, W.; Yin, W.; Xu, C.; et al. Dynamic monitoring of biomass of rice under different nitrogen treatments using a lightweight UAV with dual image-frame snapshot cameras. Plant Methods 2019, 15, 32. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Zheng, H.B.; Xu, X.Q.; He, J.Y.; Ge, X.K.; Yao, X.; Cheng, T.; Zhu, Y.; Cao, W.X.; Tian, Y.C. Predicting grain yield in rice using multi-temporal vegetation indices from UAV-based multispectral and digital imagery. ISPRS J. Photogramm. Remote Sens. 2017, 130, 246. [Google Scholar] [CrossRef]
- Duan, B.; Fang, S.; Zhu, R.; Wu, X.; Wang, S.; Gong, Y.; Peng, Y. Remote estimation of rice yield with unmanned aerial vehicle (UAV) data and spectral mixture analysis. Front. Plant Sci. 2019, 10, 204. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Q.; Fang, S.; Peng, Y.; Gong, Y.; Zhu, R.; Wu, X.; Ma, Y.; Duan, B.; Liu, J. UAV-based biomass estimation for rice-combining spectral, TIN-based structural and meteorological features. Remote Sens. 2019, 11, 890. [Google Scholar] [CrossRef] [Green Version]
- Kawamura, K.; Asai, H.; Yasuda, T.; Khanthavong, P.; Soisouvanh, P.; Phongchanmixay, S. Field phenotyping of plant height in an upland rice field in Laos using low-cost small unmanned aerial vehicles (UAVs). Plant Prod. Sci. 2020, 23, 452. [Google Scholar] [CrossRef]
- Wu, J.; Yang, G.; Yang, X.; Xu, B.; Han, L.; Zhu, Y. Automatic counting of in situ rice seedlings from UAV images based on a deep fully Convolutional Neural Network. Remote Sens. 2019, 11, 691. [Google Scholar] [CrossRef] [Green Version]
- Saberioon, M.M.; Gholizadeh, A. Novel approach for estimating nitrogen content in paddy fields using low altitude remote sensing system. Int. Arch. Photogramm. Remote Sens. Spat. Inf. Sci. Isprs Arch. 2016, 1011–1015. [Google Scholar] [CrossRef] [Green Version]
- Colorado, J.D.; Cera-Bornacelli, N.; Caldas, J.S.; Petro, E.; Rebolledo, M.C.; Cuellar, D.; Calderon, F.; Mondragon, I.F.; Jaramillo-Botero, A. Estimation of nitrogen in rice crops from UAV-captured images. Remote Sens. 2020, 12, 3396. [Google Scholar] [CrossRef]
- Yuan, S.; Peng, S.; Li, T. Evaluation and application of the ORYZA rice model under different crop managements with high-yielding rice cultivars in central China. Field Crop. Res. 2017, 212, 115. [Google Scholar] [CrossRef]
- Liu, X.J.; Qiang, C.A.O.; Yuan, Z.F.; Xia, L.I.U.; Wang, X.L.; TIan, Y.C.; Cao, W.X.; Yan, Z.H.U. Leaf area index based nitrogen diagnosis in irrigated lowland rice. J. Integr. Agric. 2018, 17, 111. [Google Scholar] [CrossRef] [Green Version]
- Tian, G.; Gao, L.; Kong, Y.; Hu, X.; Xie, K.; Zhang, R.; Ling, N.; Shen, Q.; Guo, S. Improving rice population productivity by reducing nitrogen rate and increasing plant density. PLoS ONE 2017, 12, e0182310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Chang, Q.; Yang, J.; Zhang, X.; Li, F. Estimation of paddy rice leaf area index using machine learning methods based on hyperspectral data from multi-year experiments. PLoS ONE 2018, 13, e0207624. [Google Scholar] [CrossRef] [Green Version]
- Ndikumana, E.; Ho Tong Minh, D.; Nguyen, D.; Thu, H.; Baghdadi, N.; Courault, D.; Hossard, L.; El Moussawi, I. Estimation of rice height and biomass using multitemporal SAR Sentinel-1 for Camargue, Southern France. Remote Sens. 2018, 10, 1394. [Google Scholar] [CrossRef] [Green Version]
- Zha, H.; Miao, Y.; Wang, T.; Li, Y.; Zhang, J.; Sun, W.; Feng, Z.; Kusnierek, K. Improving unmanned aerial vehicle remote sensing-based rice nitrogen nutrition index prediction with machine learning. Remote Sens. 2020, 12, 215. [Google Scholar] [CrossRef] [Green Version]
- Liang, L.; Di, L.; Huang, T.; Wang, J.; Lin, L.; Wang, L.; Yang, M. Estimation of leaf nitrogen content in wheat using new hyperspectral indices and a Random Forest regression algorithm. Remote Sens. 2018, 10, 1940. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Yang, J.; Shi, S.; Chen, B.; Du, L.; Gong, W.; Song, S. Estimating rice leaf nitrogen concentration: Influence of regression algorithms based on passive and active leaf reflectance. Remote Sens. 2017, 9, 951. [Google Scholar] [CrossRef] [Green Version]
- Yuan, H.; Yang, G.; Li, C.; Wang, Y.; Liu, J.; Yu, H.; Feng, H.; Xu, B.; Zhao, X.; Yang, X. Retrieving soybean leaf area index from unmanned aerial vehicle hyperspectral remote sensing: Analysis of RF, ANN, and SVM regression models. Remote Sens. 2017, 9, 309. [Google Scholar] [CrossRef] [Green Version]
- Apolo-Apolo, O.E.; Pérez-Ruiz, M.; Martínez-Guanter, J.; Egea, G. A mixed data-based deep neural network to estimate leaf area index in wheat breeding trials. Agronomy 2020, 10, 175. [Google Scholar] [CrossRef] [Green Version]
- Jeong, J.H.; Resop, J.P.; Mueller, N.D.; Fleisher, D.H.; Yun, K.; Butler, E.E.; Timlin, D.J.; Shim, K.M.; Gerber, J.S.; Reddy, V.R.; et al. Random Forests for global and regional crop yield predictions. PLoS ONE 2016, 11. [Google Scholar] [CrossRef]
- Mishra, S.; Mishra, D.; Santra, G.H. Adaptive Boosting of weak regressors for forecasting of crop production considering climatic variability: An empirical assessment. J. King Saud Univ. Comp. Info. Sci. 2017, 32, 949. [Google Scholar] [CrossRef]
- Sun, C.; Feng, L.; Zhang, Z.; Ma, Y.; Crosby, T.; Naber, M.; Wang, Y. Prediction of end-of-season tuber yield and tuber set in potatoes using in-season UAV-based hyperspectral imagery and machine learning. Sensors 2020, 20, 5293. [Google Scholar] [CrossRef]
- El Bilali, A.; Taleb, A. Prediction of irrigation water quality parameters using machine learning models in a semi-arid environment. J. Saudi Soc. Agric. Sci. 2020, 19, 439. [Google Scholar] [CrossRef]
- Department of Agriculture (DOA). Paddy Statistics of Malaysia 2014; Statistics Unit, Planning, Information Technology and Communications Division, DOA: Putrajaya, Malaysia, 2015. Available online: http://www.doa.gov.my/index/resources/aktiviti_sumber/sumber_awam/maklumat_pertanian/perangkaan_tanaman/perangkaan_padi_2015.pdf (accessed on 15 January 2021).
- Meier, U. Growth Stages of Mono- and Dicotyledonous Plants: BBCH Monograph, 2nd ed.; Federal Biological Research Centre for Agriculture and Forestry: Braunschweigh, Germany, 2001; pp. 66–70. [Google Scholar]
- IRRI-CREMNET (Crop and Resource Management Network). Use of Leaf Colour Chart (LCC) for N Management in Rice; International Rice Research Institute (IRRI) Network Technology: Los Baños, Philippines, 1996. [Google Scholar]
- Miller, G.L.; Miller, E.E. Determination of nitrogen in biological materials. Anal. Chem. 1948, 20, 481–488. [Google Scholar] [CrossRef]
- Cao, Q.; Miao, Y.; Wang, H.; Huang, S.; Cheng, S.; Khosla, R.; Jiang, R. Non-destructive estimation of rice plant nitrogen status with Crop Circle multispectral active canopy sensor. Field Crop. Res. 2013, 154, 133. [Google Scholar] [CrossRef]
- Tian, Y.C.; Gu, K.J.; Chu, X.; Yao, X.; Cao, W.X.; Zhu, Y. Comparison of different hyperspectral vegetation indices for canopy leaf nitrogen concentration estimation in rice. Plant Soil 2014, 376, 193. [Google Scholar] [CrossRef]
- Kanke, Y.; Tubana, B.; Dalen, M.; Harrell, D. Evaluation of red and red-edge reflectance-based vegetation indices for rice biomass and grain yield prediction models in paddy fields. Precis. Agric. 2016, 17, 507. [Google Scholar] [CrossRef]
- Cheng, T.; Song, R.; Li, D.; Zhou, K.; Zheng, H.; Yao, X.; Tian, Y.; Cao, W.; Zhu, Y. Spectroscopic estimation of biomass in canopy components of paddy rice using dry matter and chlorophyll indices. Remote Sens. 2017, 9, 319. [Google Scholar] [CrossRef] [Green Version]
- Zheng, H.; Ma, J.; Zhou, M.; Li, D.; Yao, X.; Cao, W.; Cheng, T. Enhancing the nitrogen signals of rice canopies across critical growth stages through the integration of textural and spectral information from unmanned aerial vehicle (UAV) multispectral imagery. Remote Sens. 2020, 12, 957. [Google Scholar] [CrossRef] [Green Version]
- Rouse, J.W.; Haas, R.H.; Schell, J.A.; Deering, D.W. Monitoring the Vernal Advancement and Retrogradation (Green Wave Effect) of Natural Vegetation; NASA/GSFC Type III Final Report; Goddard Space Flight Center: Greenbelt, MD, USA, 1973; p. 371. [Google Scholar]
- Gitelson, A.A.; Kaufman, Y.J.; Merzlyak, M.N. Use of a green channel in remote sensing of global vegetation from EOS-MODIS. Remote Sens. Environ. 1996, 58, 289. [Google Scholar] [CrossRef]
- Pearson, R.L.; Miller, L.D. Remote Mapping of Standing Crop Biomass for Estimation of the Productivity of the Short-Grass Prairie, Pawnee National Grasslands, Colorado. In Proceedings of the 8th International Symposium on Remote Sensing of Environment, Ann Arbor, MI, USA, 2–6 October 1972; p. 1355. Available online: https://ui.adsabs.harvard.edu/abs/1972rse.conf.1355P/abstract (accessed on 15 January 2021).
- Birth, G.S.; McVey, G.R. Measuring the color of growing turf with a reflectance spectrophotometer. Agron. J. 1968, 60, 640. [Google Scholar] [CrossRef]
- Rougean, J.L.; Breon, F.M. Estimating PAR absorbed by vegetation from bidirectional reflectance measurements. Remote Sens. Environ. 1995, 51, 375. [Google Scholar] [CrossRef]
- Barnes, E.; Clarke, T.; Richard, S.; Colaizzi, P.; Harberland, J.; Kostrzewski, M.; Waller, P.; Choi, C.; Riley, E.; Thompson, T.; et al. Coincident Detection of Crop Water Stress, Nitrogen Status and Canopy Density Using Ground Based Multispectral Data. In Proceedings of the 5th International Conference on Precision Agriculture, Bloomington, IN, USA, 16–19 July 2000. [Google Scholar]
- Gitelson, A.A.; Merzlyak, M.N.; Lichtenthaler, H.K. Detection of red edge position and chlorophyll content by reflectance measurement near 700 nm. J. Plant Physiol. 1996, 148, 501. [Google Scholar] [CrossRef]
- Pedregosa, F.; Varoquaux, G.; Gramfort, A.; Michel, V.; Thirion, B.; Grisel, O.; Blondel, M.; Prettenhofer, P.; Weiss, R.; Dubourg, V.; et al. Scikit-Learn: Machine Learning in Python. J. Mach. Learn. Res. 2011, 12, 2825. [Google Scholar]
- Zhou, C.; Ye, H.; Hu, J.; Shi, X.; Hua, S.; Yue, J.; Xu, Z.; Yang, G. Automated counting of rice panicle by applying deep learning model to images from unmanned aerial vehicle platform. Sensors 2019, 19, 3106. [Google Scholar] [CrossRef] [Green Version]
- Mao, H.; Meng, J.; Ji, F.; Zhang, Q.; Fang, H. Comparison of machine learning regression algorithms for cotton leaf area index retrieval using Sentinel-2 spectral bands. Appl. Sci. 2019, 9, 1459. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Song, Y.; Jiang, B.; Chen, B.; Shan, G. Two-stage bagging pruning for reducing the ensemble size and improving the classification performance. Math. Probl. Eng. 2019, 2019, 8906034. [Google Scholar] [CrossRef]
- Bühlmann, P. Bagging, boosting and ensemble methods. In Handbook of Computational Statistics; Springer: Berlin/Heidelberg, Germany, 2012; pp. 985–1022. [Google Scholar] [CrossRef]
- Strobl, C.; Boulesteix, A.L.; Zeileis, A.; Hothorn, T. Bias in Random Forest variable importance measures: Illustrations, sources and a solution. Bmc Bioinform. 2007, 8, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breiman, L. Random Forests. Mach Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef] [Green Version]
- Song, J. Bias corrections for Random Forest in regression using residual rotation. J. Korean Stat. Soc. 2015, 44, 321. [Google Scholar] [CrossRef]
- Freund, Y.; Schapire, R.E. Experiments with a New Boosting Algorithm. In Proceedings of the Thirteenth International Conference on International Conference on Machine Learning, Bari, Italy, 3–6 July 1996; Morgan Kaufmann Publishers Inc.: San Francisco, CA, USA, 1996; p. 148. [Google Scholar]
- Chin, W.W. The Partial Least Squares approach for structural equation modeling. In Methodology for Business and Management. Modern Methods for Business Research; Marcoulides, G.A., Ed.; Lawrence Erlbaum Associates Publishers: Mahwah, NJ, USA, 1998; pp. 295–336. [Google Scholar]
- Furuya, S. Growth diagnosis of rice plants by means of leaf color. JPM. Agric. Res. Q. 1987, 20, 147. [Google Scholar]
- Lin, F.F.; Deng, J.S.; Shi, Y.Y.; Chen, L.S.; Wang, K. Investigation of SPAD meter-based indices for estimating rice nitrogen status. Comput. Electron. Agric. 2010, 71 (Supp. 1), S60–S65. [Google Scholar] [CrossRef]
- Islam, M.R.; Haque, K.S.; Akter, N.; Karim, M.A. Leaf chlorophyll dynamics in wheat based on SPAD meter reading and its relationship with grain yield. Sci. Agric. 2014, 8, 13. [Google Scholar] [CrossRef]
- Muharam, F.M.; Maas, S.J.; Bronson, K.F.; Delahunty, T. Estimating cotton nitrogen nutrition status using leaf greenness and ground cover information. Remote Sens. 2015, 7, 7007. [Google Scholar] [CrossRef] [Green Version]
- Din, M.; Zheng, W.; Rashid, M.; Wang, S.; Shi, Z. Evaluating hyperspectral vegetation indices for leaf area index estimation of Oryza sativa L. at diverse phenological stages. Front. Plant Sci. 2017, 8, 820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vecchio, Y.; Agnusdei, G.P.; Miglietta, P.P.; Capitanio, F. Adoption of precision farming tools: The case of Italian farmers. Int. J. Environ. Res. Public Health 2020, 17, 869. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Index | Equation | Reference |
|---|---|---|
| Raw bands | ||
| Blue | ||
| Green | ||
| Red | ||
| Red-Edge | ||
| NIR | ||
| Visible-NIR indices | ||
| Normalized Difference Vegetation Index (NDVI) | Rouse et al. [41] | |
| Green Normalized Difference Vegetation Index (GNDVI) | Gitelson et al. [42] | |
| Ratio Vegetative Index (RVI) | Pearson and Miller [43] | |
| Difference Vegetation Index (DVI) | Birth and McVey [44] | |
| Renormalized Difference Vegetation Index (RDVI) | Roujean and Breon [45] | |
| Red-edge-NIR indices | ||
| Normalized Difference Red-Edge Index (NDRE) | Barnes et al. [46] | |
| Red-Edge Ratio Vegetation Index (RERVI) | Gitelson et al. [47] | |
| Red-Edge Difference Vegetation Index (REDVI) | Cao et al. [36] | |
| Red-Edge Renormalized Different Vegetation Index (RERDVI) | Cao et al. [36] | |
| Red-Edge Wide Dynamic Range Vegetation Index (REWDRVI) | Cao et al. [36] | |
| Red-Edge Soil Adjusted Vegetation Index (RESAVI) | Cao et al. [36] | |
| Red-Edge Optimized Soil Adjusted Vegetation Index (REOSAVI) | Cao et al. [36] | |
| Parameter | Description | Randomize Search Value |
|---|---|---|
| n_estimators | The number of regression trees in the forest. The higher the n_estimator parameter, the larger is the number of trees created. | 10–200 |
| max_features | The maximum number of features for splitting the node. | Auto, sqrt, log2 |
| max_depth | The maximum depth of each regression tree. | 10–200 |
| min_samples_split | The minimum number of samples for splitting an internal node. | 2–10 |
| min_samples_leaf | The minimum number of samples needed to be at a leaf node. | 1–4 |
| Growth Stage | Plant Trait | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| DA | NOP | PH | SPAD | LCC | LAI | GL | DL | STEM | SO | TOTAL | NC | YIELD | |
| Tillering | MEAN | 38.70 | 0.50 | 32.80 | 3.00 | 3.20 | 33.90 | 2.60 | 37.30 | - | 74.20 | 2.30 | - |
| MIN | 21.00 | 0.40 | 27.60 | 2.00 | 1.10 | 21.20 | 0.70 | 11.70 | - | 39.80 | 1.70 | - | |
| MAX | 64.50 | 0.70 | 36.80 | 3.50 | 6.00 | 53.50 | 5.80 | 58.60 | - | 117.80 | 2.90 | - | |
| SD | 8.90 | 0.10 | 2.00 | 0.30 | 1.00 | 7.10 | 1.10 | 8.40 | - | 15.40 | 0.20 | - | |
| Booting | MEAN | 39.80 | 0.70 | 35.70 | 3.20 | 6.60 | 72.80 | 13.90 | 145.30 | 6.10 | 238.30 | 2.10 | - |
| MIN | 27.50 | 0.60 | 31.30 | 3.00 | 3.70 | 44.60 | 5.30 | 101.00 | 0.00 | 168.40 | 0.80 | - | |
| MAX | 53.50 | 0.90 | 39.90 | 4.00 | 11.30 | 102.10 | 28.90 | 174.50 | 34.60 | 297.60 | 2.60 | - | |
| SD | 6.20 | 0.10 | 2.00 | 0.30 | 1.80 | 15.00 | 5.10 | 17.30 | 9.20 | 30.30 | 0.30 | - | |
| Milking | MEAN | 31.10 | 0.90 | 35.60 | 3.60 | 5.20 | 51.30 | 26.30 | 115.60 | 112.30 | 306.30 | 1.90 | - |
| MIN | 17.50 | 0.80 | 29.30 | 3.00 | 1.90 | 22.50 | 13.60 | 74.60 | 60.00 | 215.90 | 1.30 | - | |
| MAX | 43.00 | 1.00 | 40.60 | 4.00 | 8.70 | 83.90 | 52.10 | 179.10 | 180.20 | 422.10 | 2.40 | - | |
| SD | 5.50 | 0.10 | 2.60 | 0.40 | 1.50 | 12.80 | 7.70 | 22.20 | 31.50 | 48.80 | 0.20 | - | |
| Maturing | MEAN | - | - | - | - | - | - | - | - | - | - | - | 128.40 |
| MIN | - | - | - | - | - | - | - | - | - | - | - | 80.00 | |
| MAX | - | - | - | - | - | - | - | - | - | - | - | 173.50 | |
| SD | - | - | - | - | - | - | - | - | - | - | - | 16.90 | |
| Growth Stage | Plant Trait | Raw Band | Visible-NIR Indices | Red-Edge-NIR Indices | Trait Score | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Blue | Green | Red | Red-edge | NIR | NDVI | GNDVI | RVI | DVI | RDVI | NDRE | RERVI | REDVI | RERDVI | REWDRVI | RESAVI | REOSAVI | |||
| Tillering | NOP | 1 | 0 | 1 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 1 | 0 | 8 |
| PH | 1 | 0 | 1 | 0 | 0 | 1 | 1 | 0 | 1 | 1 | 1 | 1 | 0 | 1 | 1 | 1 | 1 | 12 | |
| SPAD | 0 | 1 | 1 | 1 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 5 | |
| LCC | 0 | 1 | 1 | 1 | 0 | 1 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 7 | |
| LAI | 0 | 0 | 1 | 0 | 1 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 9 | |
| GL | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 17 | |
| DL | 0 | 1 | 0 | 1 | 1 | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 6 | |
| STEM | 0 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 3 | |
| TOTAL | 1 | 0 | 0 | 1 | 1 | 1 | 0 | 1 | 1 | 0 | 1 | 1 | 1 | 0 | 1 | 1 | 1 | 12 | |
| YIELD | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 0 | 1 | 0 | 1 | 1 | 0 | 0 | 0 | 6 | |
| NC | 1 | 1 | 1 | 1 | 0 | 1 | 1 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 1 | 1 | 1 | 11 | |
| Index score | 5 | 6 | 7 | 8 | 6 | 7 | 5 | 6 | 5 | 2 | 5 | 6 | 5 | 4 | 6 | 7 | 6 | ||
| Growth Stage | Plant Trait | Raw Band | Visible-NIR Indices | Red-Edge-NIR Indices | Trait Score | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Blue | Green | Red | Red-edge | NIR | NDVI | GNDVI | RVI | DVI | RDVI | NDRE | RERVI | REDVI | RERDVI | REWDRVI | RESAVI | REOSAVI | |||
| Booting | NOP | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 2 |
| PH | 0 | 0 | 1 | 0 | 1 | 1 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 0 | 12 | |
| SPAD | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 12 | |
| LCC | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 0 | 1 | 1 | 0 | 1 | 1 | 1 | 1 | 1 | 15 | |
| LAI | 1 | 1 | 1 | 0 | 1 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 15 | |
| GL | 0 | 0 | 0 | 1 | 1 | 0 | 1 | 0 | 0 | 1 | 1 | 1 | 0 | 1 | 1 | 1 | 1 | 10 | |
| DL | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 4 | |
| STEM | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 1 | 0 | 0 | 1 | 1 | 1 | 1 | 0 | 9 | |
| SO | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 15 | |
| TOTAL | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 1 | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 7 | |
| YIELD | 1 | 1 | 0 | 1 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 5 | |
| NC | 0 | 1 | 1 | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 14 | |
| Index score | 3 | 6 | 6 | 5 | 7 | 6 | 8 | 8 | 8 | 8 | 7 | 6 | 8 | 9 | 8 | 8 | 7 | ||
| Growth Stage | Plant Trait | Raw Band | Visible Indices | Red-Edge-NIR Indices | Trait Score | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Blue | Green | Red | Red-edge | NIR | NDVI | GNDVI | RVI | DVI | RDVI | NDRE | RERVI | REDVI | RERDVI | REWDRVI | RESAVI | REOSAVI | |||
| Milking | NOP | 1 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 0 | 1 | 1 | 1 | 0 | 1 | 1 | 1 | 14 |
| PH | 1 | 1 | 1 | 1 | 1 | 1 | 0 | 1 | 1 | 0 | 1 | 0 | 1 | 1 | 1 | 1 | 1 | 14 | |
| SPAD | 1 | 0 | 1 | 0 | 1 | 1 | 0 | 1 | 1 | 1 | 1 | 1 | 0 | 1 | 1 | 0 | 0 | 11 | |
| LCC | 0 | 0 | 1 | 0 | 1 | 1 | 1 | 0 | 0 | 1 | 1 | 1 | 0 | 0 | 1 | 1 | 1 | 10 | |
| LAI | 0 | 1 | 1 | 0 | 1 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 14 | |
| GL | 1 | 0 | 1 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 15 | |
| DL | 1 | 0 | 0 | 1 | 0 | 0 | 1 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 7 | |
| STEM | 1 | 1 | 1 | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 15 | |
| SO | 1 | 1 | 0 | 1 | 0 | 1 | 1 | 0 | 0 | 0 | 1 | 1 | 0 | 1 | 1 | 0 | 1 | 10 | |
| TOTAL | 1 | 0 | 1 | 1 | 0 | 1 | 1 | 1 | 0 | 1 | 1 | 1 | 1 | 0 | 1 | 0 | 1 | 12 | |
| YIELD | 1 | 0 | 1 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 10 | |
| NC | 1 | 1 | 0 | 1 | 0 | 1 | 0 | 1 | 0 | 0 | 1 | 1 | 0 | 1 | 1 | 1 | 0 | 10 | |
| Index score | 10 | 5 | 9 | 6 | 7 | 10 | 9 | 9 | 8 | 8 | 10 | 9 | 7 | 8 | 10 | 8 | 9 | ||
| TOTAL SCORE | 18 | 17 | 22 | 19 | 20 | 23 | 22 | 23 | 21 | 18 | 22 | 21 | 20 | 21 | 24 | 23 | 22 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muharam, F.M.; Nurulhuda, K.; Zulkafli, Z.; Tarmizi, M.A.; Abdullah, A.N.H.; Che Hashim, M.F.; Mohd Zad, S.N.; Radhwane, D.; Ismail, M.R. UAV- and Random-Forest-AdaBoost (RFA)-Based Estimation of Rice Plant Traits. Agronomy 2021, 11, 915. https://doi.org/10.3390/agronomy11050915
Muharam FM, Nurulhuda K, Zulkafli Z, Tarmizi MA, Abdullah ANH, Che Hashim MF, Mohd Zad SN, Radhwane D, Ismail MR. UAV- and Random-Forest-AdaBoost (RFA)-Based Estimation of Rice Plant Traits. Agronomy. 2021; 11(5):915. https://doi.org/10.3390/agronomy11050915
Chicago/Turabian StyleMuharam, Farrah Melissa, Khairudin Nurulhuda, Zed Zulkafli, Mohamad Arif Tarmizi, Asniyani Nur Haidar Abdullah, Muhamad Faiz Che Hashim, Siti Najja Mohd Zad, Derraz Radhwane, and Mohd Razi Ismail. 2021. "UAV- and Random-Forest-AdaBoost (RFA)-Based Estimation of Rice Plant Traits" Agronomy 11, no. 5: 915. https://doi.org/10.3390/agronomy11050915