
Production of Virus-Free Garlic Plants through Somatic Embryogenesis

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Culture Establishment, Induction of Somatic Embryogenesis and Plant Regeneration
2.3. Rooting and Acclimatisation
2.4. Virus Detection and Efficiency of Virus Elimination
2.4.1. Enzyme-Linked Immunosorbent Assay (ELISA)
2.4.2. Reverse Transcription Polymerase Chain Reaction (RT-PCR)
2.5. Statistical Analysis
3. Results and discussion
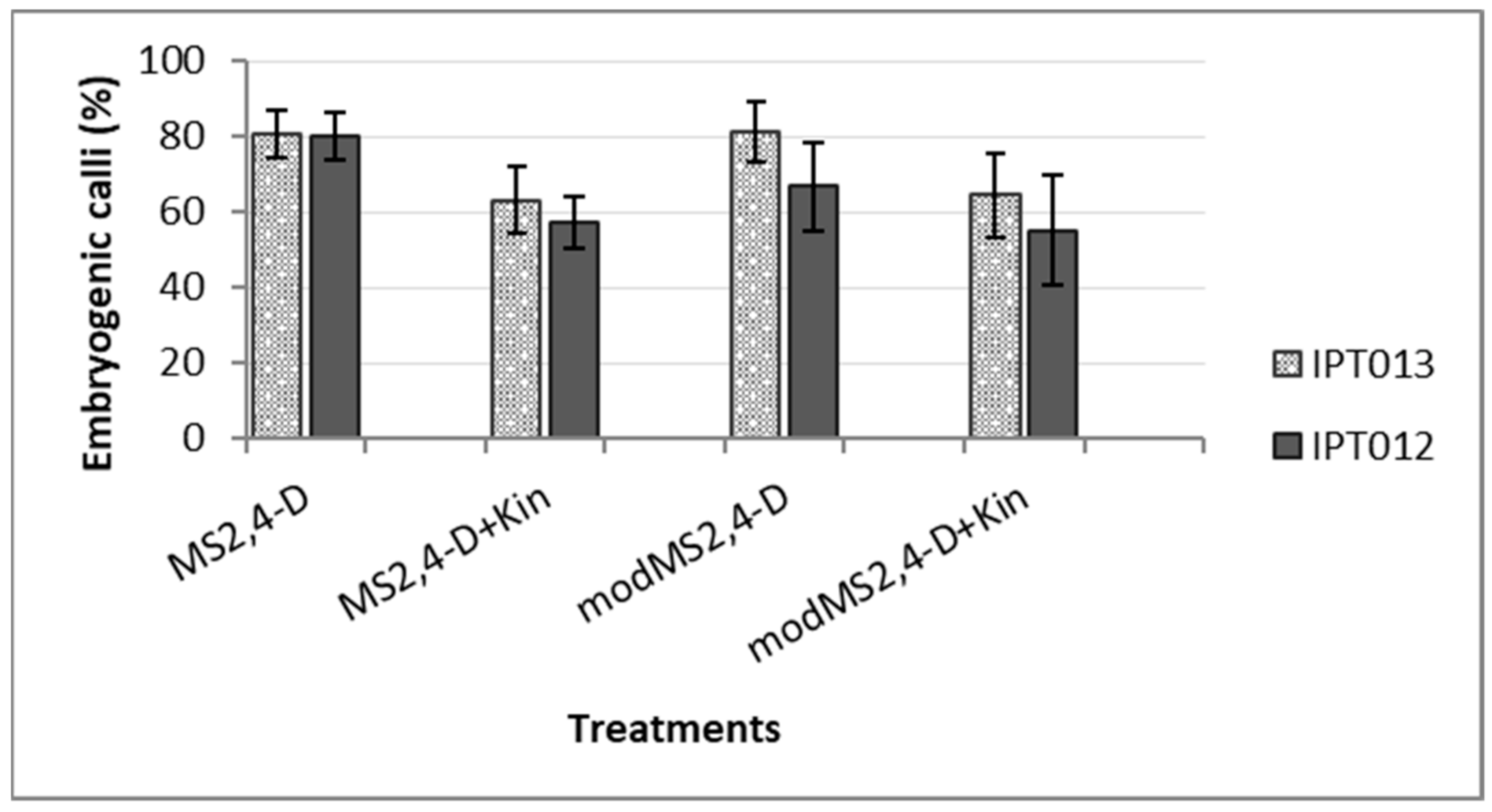
3.1. Culture Establishment, Callus Formation and Induction of Somatic Embryogenesis
3.2. Plant Regeneration
3.3. Rooting and Acclimatisation
3.4. Virus Detection and Efficiency of Virus Elimination
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Food and Agriculture Organization of the United Nations. FAOSTAT Statistical Database. 1997. Available online: http://www.fao.org/faostat/en/#data (accessed on 27 March 2021).
- González, R.E.; Soto, V.C.; Sance, M.M.; Camargo, A.B.; Galmarini, C.R. Variability of solids, organosulfur compounds, pungency and health-enhancing traits in garlic (Allium sativum L.) cultivars belonging to different ecophysiological groups. J. Agric. Food Chem. 2009, 57, 10282–10288. [Google Scholar] [CrossRef] [PubMed]
- Dumičić, G.; Miloš, B.; Žanić, K.; Urlić, B.; Jukić Špika, M.; Čagalj, M. Jadranski Češnjak; Institut za Jadranske Kulture i Melioraciju Krša: Split, Croatia, 2015; Available online: https://bib.irb.hr/datoteka/758246.luk_arci_zadnja.pdf (accessed on 25 February 2021). (In Croatian)
- Toth, N.; Fabek, S.; Vončina, D. Završno Izvješće: Revitalizacija Proizvodnje Češnjaka u Zadarskoj Županiji; Project Final Report; Agronomski Fakultet: Zagreb, Croatia, 2015. (In Croatian) [Google Scholar]
- Conci, V.C.; Canavelli, A.; Lunello, P. Yield losses associated with virus-infected garlic plants during five successive years. Plant Dis. 2003, 87, 1411–1415. [Google Scholar] [CrossRef] [Green Version]
- Vončina, D.; Ćurić, K.; Fabek, S.; Toth, N. First report of Onion yellow dwarf virus, Leek yellow stripe virus and Garlic common latent virus in Croatia. Plant Dis. 2016, 100, 656. [Google Scholar] [CrossRef]
- Vončina, D.; Ćurić, K.; Toth, N.; Uher, S.F. Viruses of some garlic ecotypes in Croatia. Special Issue: Proceedings/Tanaskovikj, Vjekoslav; Porchu, Kocho—Skopje: “Ss. Cyril and Methodius” University in Skopje, Faculty of Agricultural Sciences and Food-Skopje. J. Agric. Food Environ. Sci. 2017, 71, 87–91. [Google Scholar]
- Godena, S.; Ban, D.; Dumičić, G.; Ban, S.G. Incidence of viruses in cloves and bulbils of garlic ecotypes in Croatia. Acta Sci. Pol. Hortorum Cultus 2020, 5, 91–99. [Google Scholar] [CrossRef]
- Walkey, D.G.A.; Antill, D.N. Agronomic evaluation of virus-free and virus infected garlic (Allium sativum L.). J. Hortic. Sci. 1989, 64, 53–60. [Google Scholar] [CrossRef]
- Lot, H.; Chovelon, V.; Souche, S.; Delecolle, B. Effects of onion yellow dwarf and leek yellow stripe viruses on symptomatology and yield loss of three French garlic cultivars. Plant Dis. 1998, 82, 1381–1385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pramesh, D.; Baranwal, V.K. Production of virus-free garlic (Allium sativum L.) through meristem tip culture after solar or hot air treatment of cloves. J. Hortic. Sci. Biotechnol. 2015, 90, 180–186. [Google Scholar] [CrossRef]
- Moriconi, D.N.; Conci, V.C.; Nome, S.F. Rapid multiplication of garlic Allium sativum L. in vitro. Phyton 1990, 51, 145–151. [Google Scholar]
- Verbeek, M.; van Dijk, P.; van Well, P.M.A. Efficiency of eradication of four viruses from garlic (Allium sativum) by meristem-tip culture. Eur. J. Plant Pathol. 1995, 101, 231–239. [Google Scholar] [CrossRef]
- Torres, A.C.; Fajardo, T.V.; Dusi, A.N.; Resende, R.O.; Buso, J.A. Shoot tip culture and thermotherapy in recovering virus free plants of garlic. Hortic. Bras. 2000, 18, 192–195. [Google Scholar] [CrossRef] [Green Version]
- Conci, V.C.; Nome, S.F. Virus free garlic (Allium Sativum L.) plants obtained by thermotherapy and meristem tip culture. J. Phytopathol. 1991, 132, 186–192. [Google Scholar] [CrossRef]
- Kudělková, M.; Ondrušiková, E.; Sasková, H. Elimination of Garlic common latent virus by meristem culture and chemotherapy. Acta Hortic. 2016, 233–238. [Google Scholar] [CrossRef]
- Senula, A.; Keller, E.R.J.; Leseman, D.E. Elimination of viruses through meristem culture and thermotherapy for the establishment of an in–vitro collection of garlic (Allium sativum L.). Acta Hortic. 2000, 530, 121–128. [Google Scholar] [CrossRef]
- Danci, O.; Erdei, L.; Vidacs, L.; Danci, M.; Baciu, A.; David, I.; Berbentea, F. Influence of ribavirin on potato plants regeneration and virus. J. Hortic. For. Biotechnol. 2009, 13, 421–425. [Google Scholar]
- Haque, M.S.; Hattori, K.A.; Suzuki, A.; Tsuneyoshi, T. An efficient novel method of producing virus free plants from garlic root meristem. In Biotechnology and Sustainable Agriculture 2006 and Beyond; Xu, Z., Li, J., Xue, Y., Yang, W., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 107–110. [Google Scholar] [CrossRef]
- Goussard, P.G.; Wiid, J. The elimination of fanleaf virus from grapevines using in vitro somatic embryogenesis combined with heat therapy. S. Afr. J. Enol. Vitic. 1992, 13, 81–83. [Google Scholar] [CrossRef] [Green Version]
- Gambino, G.; Bondaz, J.; Gribaudo, I. Detection and elimination of viruses in callus, somatic embryos and regenerated plantlets of grapevine. Eur. J. Plant Pathol. 2006, 114, 397–404. [Google Scholar] [CrossRef]
- Malenica, N.; Jagić, M.; Pavletić, B.; Bauer, N.; Vončina, D.; Zdunić, G.; Leljak Levanić, D. Somatic embryogenesis as a tool for virus elimination in Croatian indigenous grapevine cultivars. Acta Bot. Croat. 2020, 79, 26–34. [Google Scholar] [CrossRef]
- Quainoo, A.K.; Wetten, A.C.; Allainguillaume, J. The effectiveness of somatic embryogenesis in eliminating the cocoa swollen shoot virus from infected cocoa trees. J. Virol. Methods 2008, 149, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Damba, Y.; Quainoo, A.K.; Sowley, E.N.K. Effectiveness of somatic embryogenesis in eliminating the cassava mosaic virus from infected cassava (Manihot esculenta Crantz) plant materials. Int. J. Sci. Technol. Res. 2013, 2, 282–287. [Google Scholar]
- Dewanti, P.; Widuri, L.I.; Ainiyati, C.; Okviandari, P.; Masairo; Sugiharto, B. Elimination of SCMV (sugarcane Mozaik virus) and rapid propagation of virus-free sugarcane (Saccharum officinarum L.) using somatic embryogenesis. Proc. Chem. 2016, 18, 96–102. [Google Scholar] [CrossRef] [Green Version]
- Keles, D.; Taskın, H.; Baktemur, G.; Yucel, N.K.; Buyukalaca, S. Somatic embryogenesis in garlic (Allium sativum L.). Acta Hortic. 2010, 923, 39–45. [Google Scholar] [CrossRef]
- Lee, S.Y.; Kim, H.H.; Kim, Y.K.; Park, N., II; Park, S.U. Plant regeneration of garlic (Allium sativum L.) via somatic embryogenesis. Sci. Res. Essays 2009, 4, 1569–1574. [Google Scholar]
- Hassan, M.N.; Haque, M.S.; Hassan, M.M.; Haque, M.S. An efficient protocol for somatic embryogenesis of garlic (Allium sativum L.) using root tip as explant. J. Bangladesh Agric. Univ. 2014, 12, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Nasim, S.A.; Mujib, A.; Kapoor, R.; Fatima, S.; Mahmooduzzafar, A.J. Somatic embryogenesis in Allium sativum L. (cv. Yamuna Safed 3): Improving embryo maturation and germination with PGRs and carbohydrates. Anales de Biología 2010, 32, 1–9. [Google Scholar]
- Khan, N.; Alam, M.S.; Nath, U.K. In vitro Regeneration of Garlic Through Callus Culture. J. Biol. Sci. 2004, 4, 189–191. [Google Scholar] [CrossRef] [Green Version]
- Fereol, L.; Chovelon, V.; Causse, S.; Michaux-Ferriere, N.; Kahane, R. Evidence of a somatic embryogenesis process for plant regeneration in garlic (Allium sativum L.). Plant Cell Rep. 2002, 21, 197–203. [Google Scholar] [CrossRef]
- Sata, S.J.; Bagatharia, S.B.; Thaker, V.S. Induction of direct somatic embryogenesis in garlic (Allium sativum). Meth. Cell. Sci. 2001, 22, 299–304. [Google Scholar] [CrossRef]
- Sudarmonowati, E.; Hartati, N.S.; Rahman, N. In vitro maintenance of embryogenic callus of garlic (Allium sativum) and its genetic stability after 6 year maintenance. Ann. Bogor. 2001, 7, 83–89. [Google Scholar]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassay with tobacco tissue culture. Physiol. Plant 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Fidan, H.; Baloğlu, S. First report of garlic common latent virus in garlic in Turkey. J. Plant Pathol. 2009, 91, 97–112. [Google Scholar]
- Dovas, C.I.; Hatziloukas, E.; Salomon, R.; Barg, E.; Shiboleth, Y.; Katis, N.I. Comparison of methods for virus detection in Allium spp. J. Phytopath. 2001, 149, 731–737. [Google Scholar] [CrossRef]
- SAS/STAT® Software, version [9.4] 2002–2012; SAS Institute Inc.: Cary, NC, USA, 2013.
- Luciani, G.F.; Mary, A.K.; Pellegrini, C.; Curvetto, N.R. Effects of explants and growth regulators in garlic callus formation and plant regeneration. Plant Cell Tissue Organ Cult. 2006, 87, 139–143. [Google Scholar] [CrossRef]
- Haque, M.S.; Wada, T.; Hattori, K. Efficient plant regeneration in garlic through somatic embryogenesis from root tip explants. Plant Prod. Sci. 1998, 1, 216–222. [Google Scholar] [CrossRef]
- White, P.R. The Cultivation of Animal and Plant Cells; Ronald Press: New York, NY, USA, 1963; Volume 2, p. 228. [Google Scholar]
- Lloyd, C.W.; Lowe, S.B.; Peace, G.W. The mode of action of 2,4-D in counter-acting the elongation of carrot cells grown in culture. J. Cell Sci. 1980, 45, 257–268. [Google Scholar] [CrossRef] [PubMed]
- Vondráková, Z.; Eliášová, K.; Fischerová, L.; Vágner, M. The role of auxins in somatic embryogenesis of Abies alba. Open Life Sci. 2011, 6, 587–596. [Google Scholar] [CrossRef]
- Khaleda, L.; Al-Forkan, M. Stimulatory effects of casein hydrolysate and proline in in vitro callus induction and plant regeneration from five deepwater rice (Oryza sativa L.). Biotechnology 2006, 5, 379–384. [Google Scholar] [CrossRef] [Green Version]
- Pasternak, T.P. The role of auxin, pH, and stress in the activation of embryogenic cell division in leaf protoplast-derived cells of alfalfa. Plant Physiol. 2002, 129, 1807–1819. [Google Scholar] [CrossRef] [Green Version]
- Zavattieri, M.A.; Frederico, A.M.; Lima, M.; Sabino, R.; Amhold-Schmidt, B. Induction of somatic embryogenesis as an example of stress-related plant reaction. Electron. J. Biotechnol. 2010, 13, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Nissen, P.; Minocha, S.C. Inhibition by 2,4-D of somatic embryogenesis in carrot as explored by its reversal by difluoromethylornithine. Physiol. Plant 1993, 89, 673–680. [Google Scholar] [CrossRef]
- Chée, R.P.; Cantliffe, D.J. Inhibition of somatic embryogenesis in response to 2, 3, 5-triiodobenzoic acid and 2, 4-dichlorophenoxyacetic acid in Ipomoea batatas (L.) Lam. cultured in vitro. J. Plant Physiol. 1989, 135, 398–403. [Google Scholar] [CrossRef]
- Venkatesh, K.; Rani, A.R.; Baburao, N.; Padmaja, G. Effect of auxins and auxin polar transport inhibitor (TIBA) on somatic embryogenesis in groundnut (Arachis hypogaea L.). Afr. J. Plant Sci. 2009, 3, 277–282. [Google Scholar]
- Garcia, C.; de Almeida, A.A.F.; Costa, M.; Britto, D.; Valle, R.; Royaert, S.; Marelli, J.P. Abnormalities in somatic embryogenesis caused by 2,4-D: An overview. Plant Cell Tissue Organ Cult. 2019, 137, 193–212. [Google Scholar] [CrossRef] [Green Version]
- George, E.F.; Sherrington, P.D. Plant Propagation by Tissue Culture—Handbook and Directory of Commercial Laboratories; Exegetics Limited: Edington, UK, 1984. [Google Scholar]
- Wang, J.; Tian, C.; Zhang, C.; Shi, B.; Cao, X.; Zhang, T.Q.; Zhao, Z.; Wang, J.W.; Jiao, Y. Cytokinin Signaling Activates WUSCHEL Expression during Axillary Meristem Initiation. Plant Cell 2017, 2, 1373–1387. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Qu, X.; Dong, Z.; Luo, L.; Shao, C.; Forner, J.; Lohmann, J.U.; Su, M.; Xu, M.; Liu, X.; et al. WUSCHEL Triggers Innate Antiviral Immunity in Plant Stem Cells. Science 2020, 370, 227–231. [Google Scholar] [CrossRef]
- Wang, P.J.; Hu, C.Y. Regeneration of virus-free plants through in vitro culture. In Advances in Biochemical Engineering, Plant Cell Culture II; Fiechter, A., Ed.; Springer: Berlin/Heidelberg, Germany, 1980; Volume 18, pp. 61–99. [Google Scholar] [CrossRef]
- Walkey, D.G.A. Applied Plant Virology; Springer: Dordrecht, The Netherlands, 1991. [Google Scholar] [CrossRef]
- Mitchell, J.W.; Schneider, I.R.; Gauch, H.G. Translocation of particles within plants. Science 1960, 131, 1863–1870. [Google Scholar] [CrossRef]
- Spencer, D.F.; Kimmins, W.L. Presence of plasmodesmata in callus cultures of tobacco and carrot. Can. J. Bot. 1969, 47, 2049–2050. [Google Scholar] [CrossRef]
- Brighigna, L.; Schiff, S.; Bennici, A. An ultrastructural study of Pittosporum tobira Ait. callus: The status of intercellular connections. Adv. Hortic. Sci. 1992, 6, 37–43. [Google Scholar] [CrossRef]
- Yulianingsih, R.; Hidayat, S.H.; Dinart, D. Elimination of Garlic common latent virus from garlic through meristem culture and thermotherapy. IOP Conf. Ser. Earth Environ. Sci. 2020, 468, 012028. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Treatments | Composition |
|---|---|
| MS2,4-D | MS salts and vitamins (Murashige and Skoog [34]), myoinositol 0.1 g L−1, sucrose 30 g L−1, proline 100 mg L−1, casein hydrolysate 100 mg L−1, 2,4-D 0.1 mg L−1, Plant agar (Duchefa) 6 g L−1; pH 5.8 |
| MS2,4-D + Kin | MS salts and vitamins, myoinositol 0.1 g L−1, sucrose 30 g L−1, proline 100 mg L−1, casein hydrolysate 100 mg L−1, 2,4-D 1 mg L−1, Kin 0.5 mg L−1, Plant agar (Duchefa) 6 g L−1; pH 5.8 |
| modMS2,4-D | Same as MS2,4-D but with ¼ of KNO3 and NH4NO3 and 2xMgSO4 as compared with full strength MS |
| modMS2,4-D + Kin | Same as MS2,4-D + Kin but with ¼ of KNO3 and NH4NO3 and 2xMgSO4 as compared with full strength MS |
| No. of Plants/Embryogenic Callus ± SE | |||
|---|---|---|---|
| Treatment | IPT013 | IPTO12 | Mean |
| MS2,4-D | 3.8 ± 0.9 | 3.0 ± 1.0 | 3.4 a |
| modMS2,4-D | 5.3 ± 1.6 | 2.3 ± 1.0 | 3.8 a |
| MS2,4 D+Kin→MS2,4-D | 0.4 ± 0.3 | 0.2 ± 0.1 | 0.3 c |
| modMS2,4 D+Kin→MS2,4-D | 2.6 ± 1.2 | 0.4 ± 0.2 | 1.5 b |
| Mean | 3.0 a | 1.5 b | |
| Previous Treatment | Rooted Plantlets ± SE (%) | Acclimatised Plants (%) | Acclimatised Plants (N) |
|---|---|---|---|
| MS2,4-D | 59.4 ± 3.3 b | 64.0 | 122 |
| MS2,4-D + Kin→MS2,4-D | 89.0 ± 7.3 a | 66.0 | 19 |
| Mother Plant | Viruses Confirmed in Mother Plants | Virus Elimination Success Rate Determined in Regenerants (No. of Virus Free Plants/No. of Tested Plants) | ||||
|---|---|---|---|---|---|---|
| OYDV | GCLV | LYSV | OYDV | GCLV | LYSV | |
| MS2,4-D treatment | ||||||
| IPT013-2 * | + | + | + | 1/11 | 2/11 | 2/11 |
| IPT013-3 | + | - | + | 0/7 | n/a | 2/7 |
| IPT013-6 | + | - | + | 4/16 | n/a | 7/16 |
| IPT013-7 | + | - | + | 3/9 | n/a | 3/9 |
| IPT013-8 | + | - | + | 3/13 | n/a | 3/13 |
| Ecotype IPT013 virus elimination rate | 11/56 19.6% | 2/11 18.2% | 17/56 30.4% | |||
| IPTO12-4 | + | + | + | 0/2 | 0/2 | 0/2 |
| IPTO12-6 | + | + | + | 1/7 | 1/7 | 1/7 |
| IPTO12-8 | + | + | + | 1/6 | 2/6 | 2/6 |
| Ecotype IPT012 virus elimination rate | 2/15 13.3% | 3/15 20% | 3/15 20% | |||
| Total virus elimination rate for MS2,4-D treatment | 13/71 18.3% | 5/26 19.2% | 20/71 28.2% | |||
| MS2,4-D + Kin→MS2,4-D treatment | ||||||
| IPT013-2 | + | + | + | 4/6 | 3/6 | 5/6 |
| IPT013-6 | + | - | + | 0/2 | n/a | 0/2 |
| Ecotype IPT013 virus elimination rate | 4/8 50% | 3/6 50% | 5/8 62.5% | |||
| IPTO12-8 | + | + | + | 0/3 | 0/3 | 0/3 |
| Ecotype IPT012 virus elimination rate | 0/3 0% | 0/3 0% | 0/3 0% | |||
| Total virus elimination rate for MS2,4-D + Kin→MS2,4-D treatment | 4/11 36.4% | 3/9 33.3% | 5/11 45.5% | |||
| Total virus elimination rate for both treatments | 17/82 20.7% | 8/35 22.9% | 25/82 30.5% | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kereša, S.; Kurtović, K.; Ban, S.G.; Vončina, D.; Jerčić, I.H.; Bolarić, S.; Lazarević, B.; Godena, S.; Ban, D.; Mihovilović, A.B. Production of Virus-Free Garlic Plants through Somatic Embryogenesis. Agronomy 2021, 11, 876. https://doi.org/10.3390/agronomy11050876
Kereša S, Kurtović K, Ban SG, Vončina D, Jerčić IH, Bolarić S, Lazarević B, Godena S, Ban D, Mihovilović AB. Production of Virus-Free Garlic Plants through Somatic Embryogenesis. Agronomy. 2021; 11(5):876. https://doi.org/10.3390/agronomy11050876
Chicago/Turabian StyleKereša, Snježana, Katarina Kurtović, Smiljana Goreta Ban, Darko Vončina, Ivanka Habuš Jerčić, Snježana Bolarić, Boris Lazarević, Sara Godena, Dean Ban, and Anita Bošnjak Mihovilović. 2021. "Production of Virus-Free Garlic Plants through Somatic Embryogenesis" Agronomy 11, no. 5: 876. https://doi.org/10.3390/agronomy11050876