High Soil Phosphorus Application Significantly Increased Grain Yield, Phosphorus Content but Not Zinc Content of Cowpea Grains

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Description of the Experiments
2.3. Determination of Phosphorus Concentration and Zinc Content from Plant Tissues
2.4. Statistical Analyses
2.4.1. Descriptive Analysis
2.4.2. Relationships among Yield, Phosphorus, and Zinc Content
2.4.3. Interaction of the Genotype Main Effect Plus Genotype by Environment of the Genotypes
3. Results
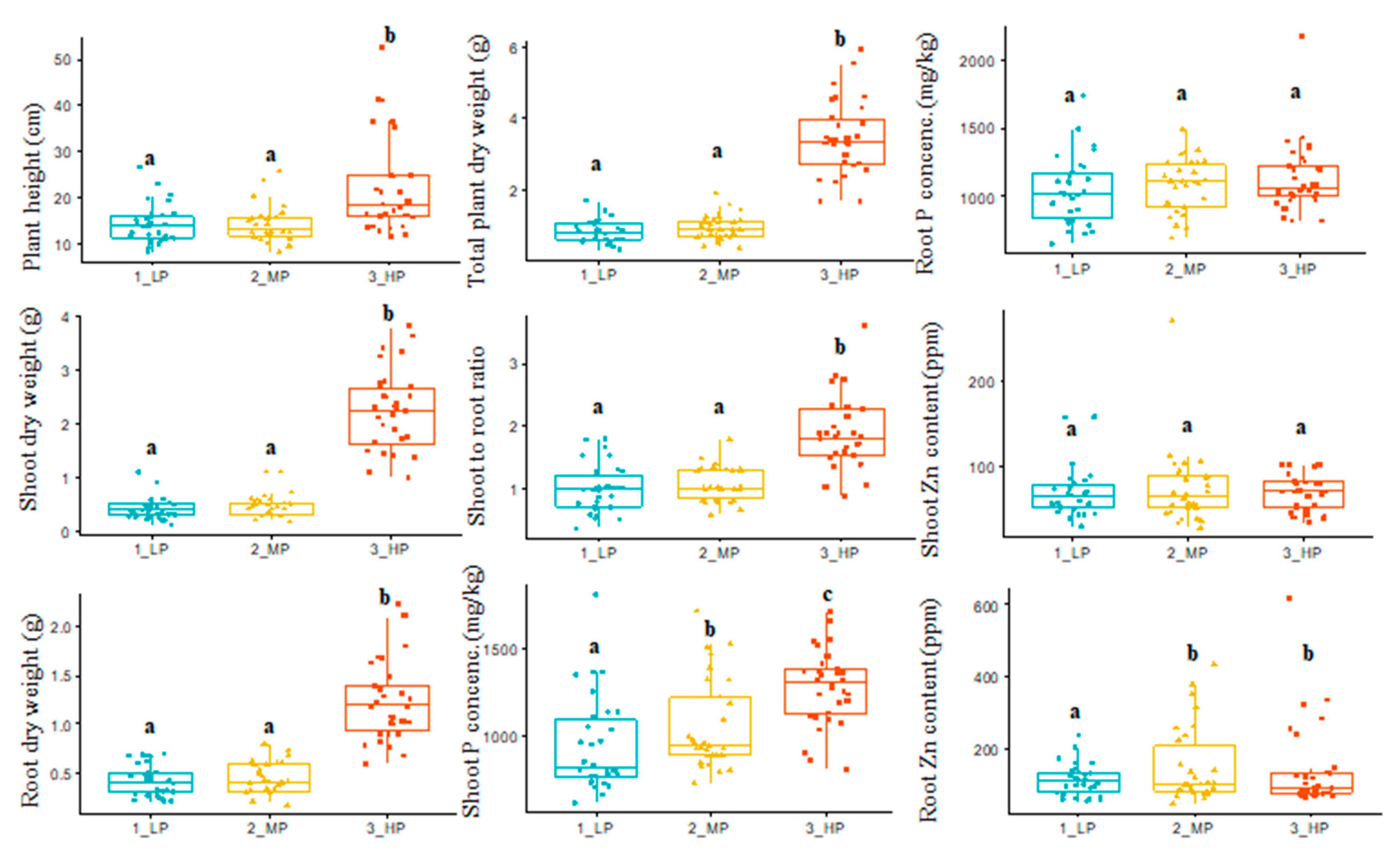
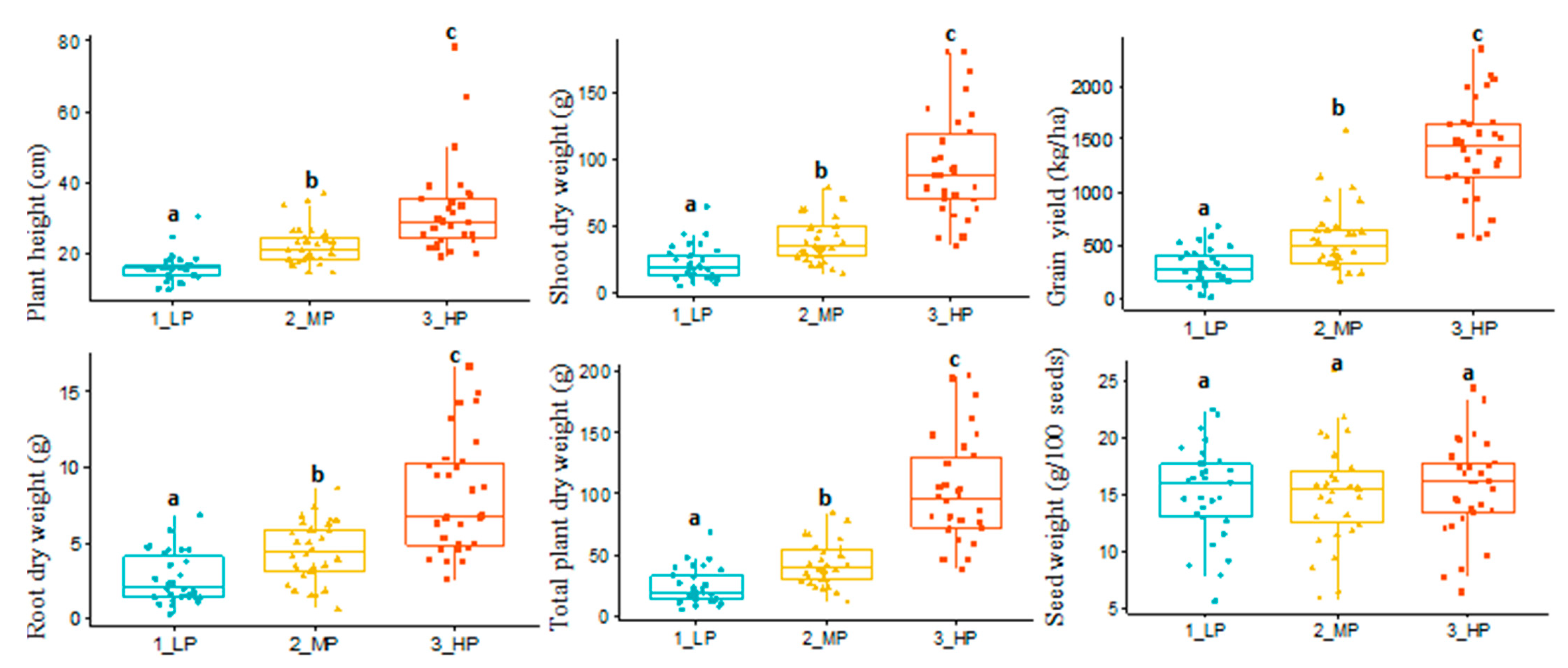
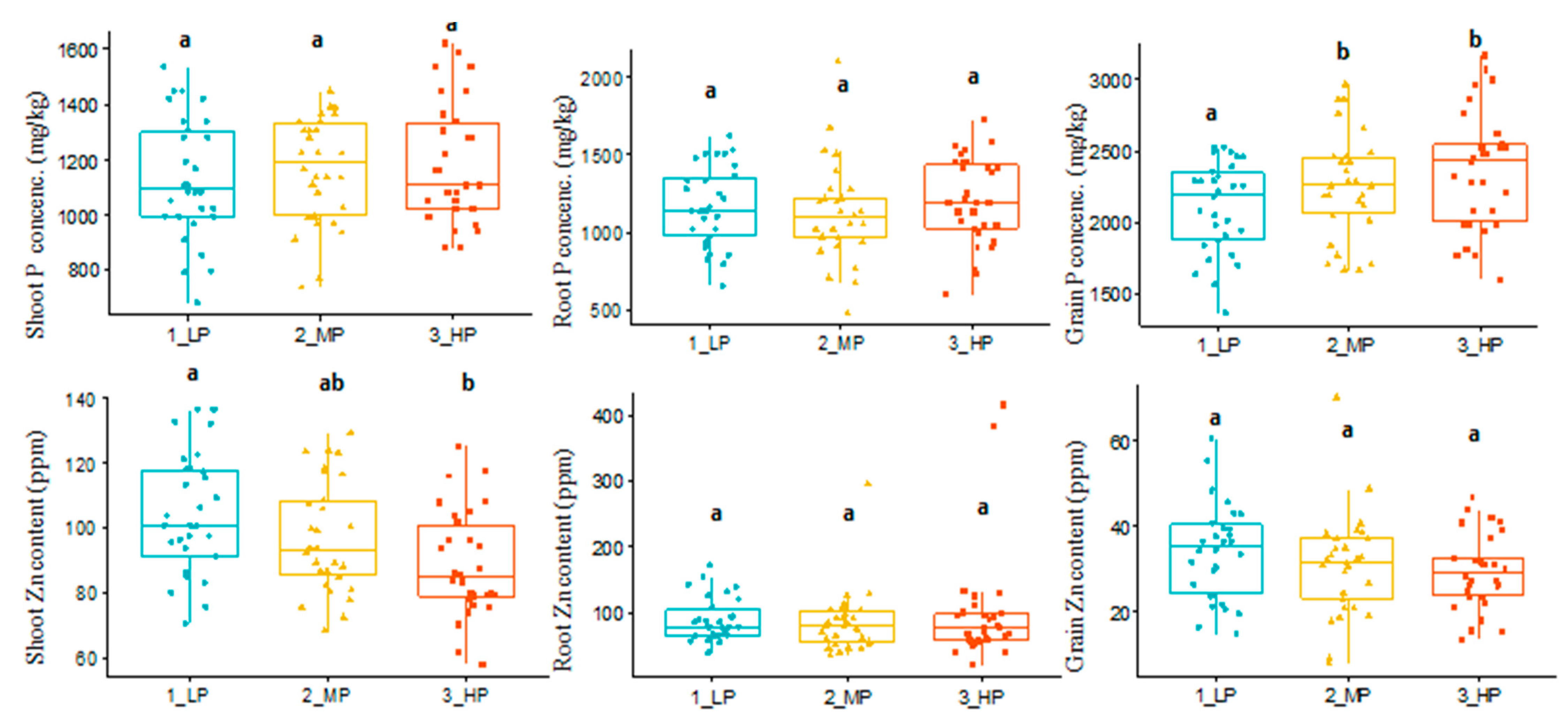
3.1. Variable Soil Phosphorus Effect on Shoot Dry Matter and Content of Phosphorus and Zinc
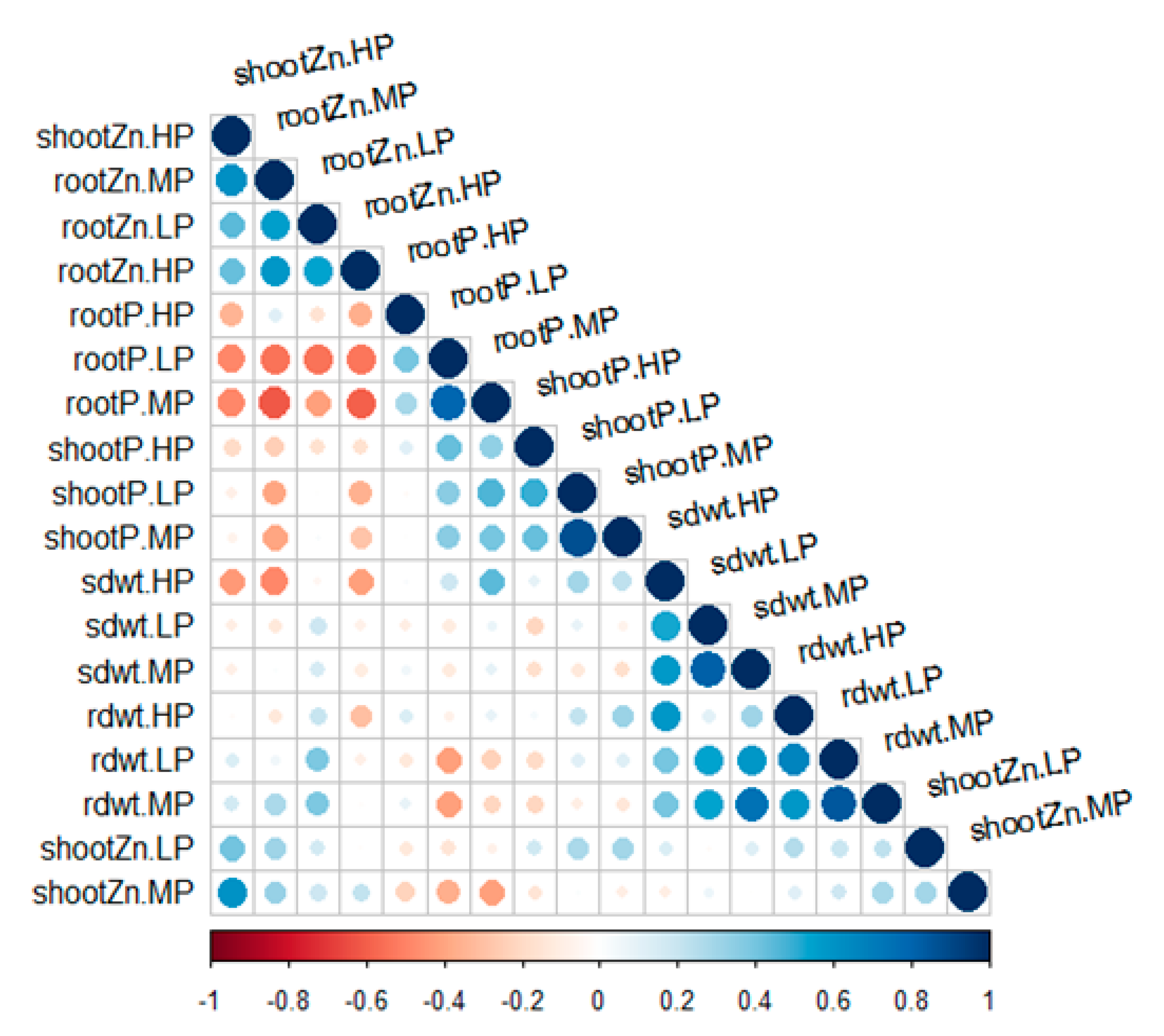
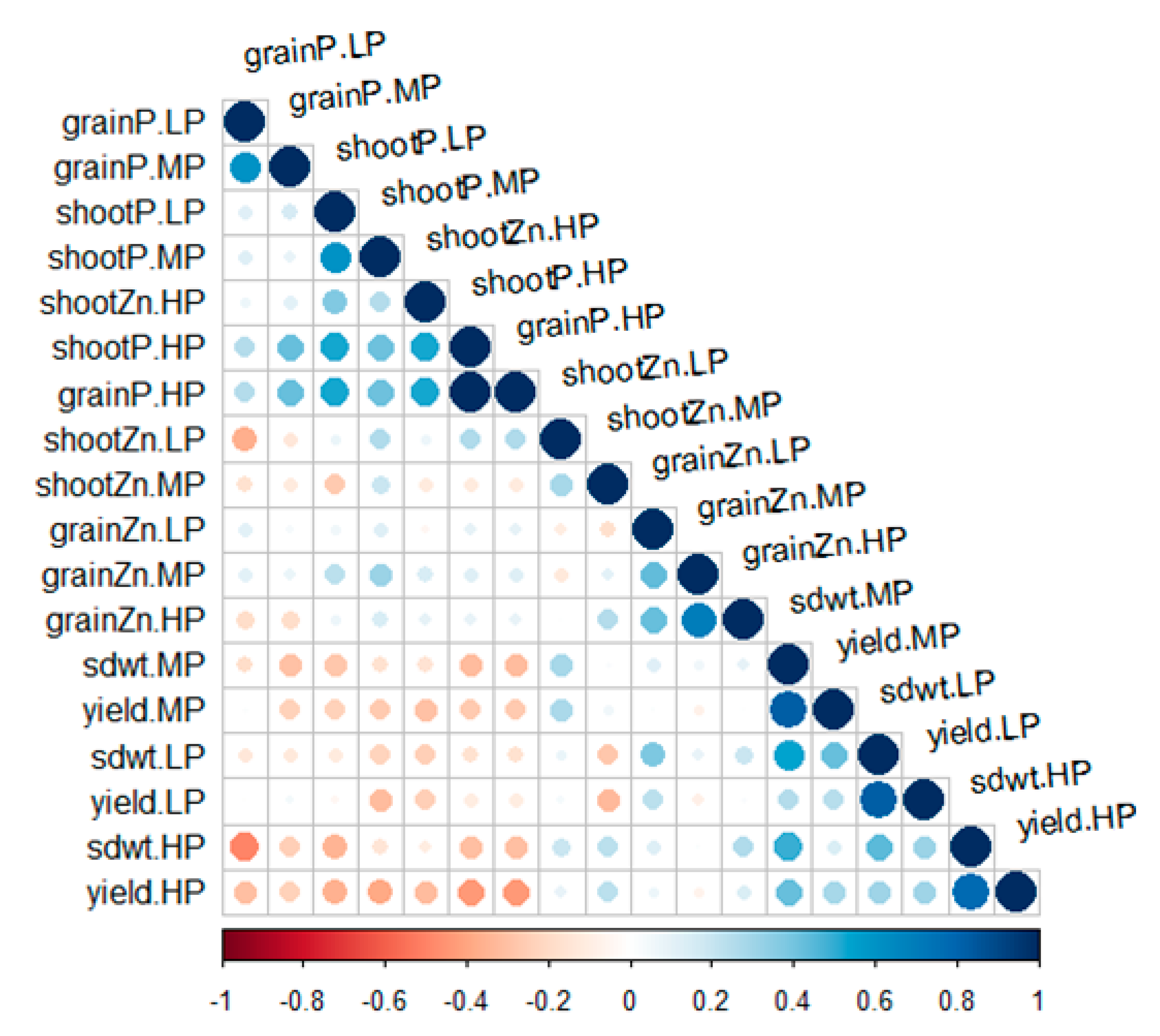
3.2. Phenotypic Association between Yield Components and Zinc Content in Cowpea
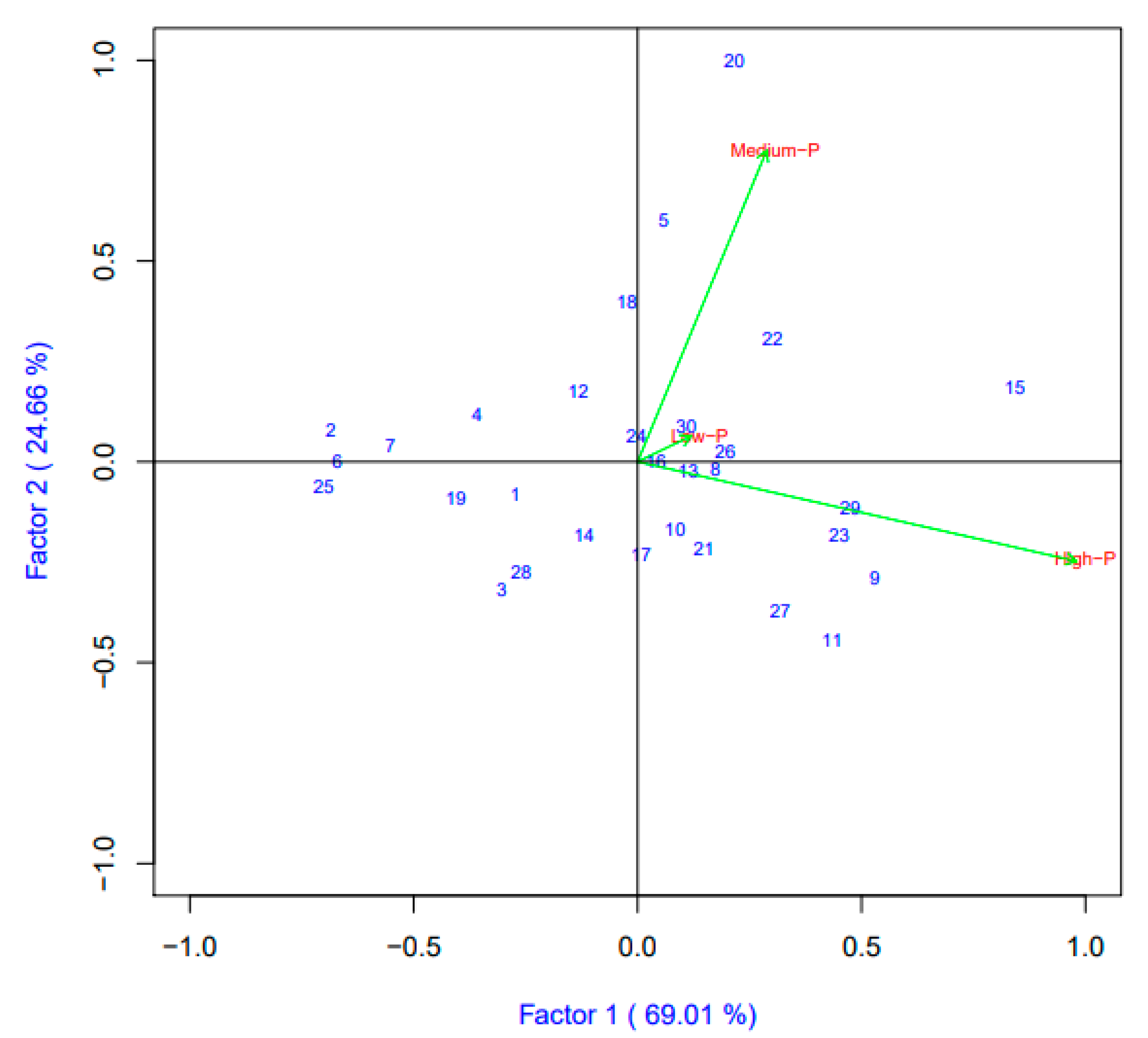
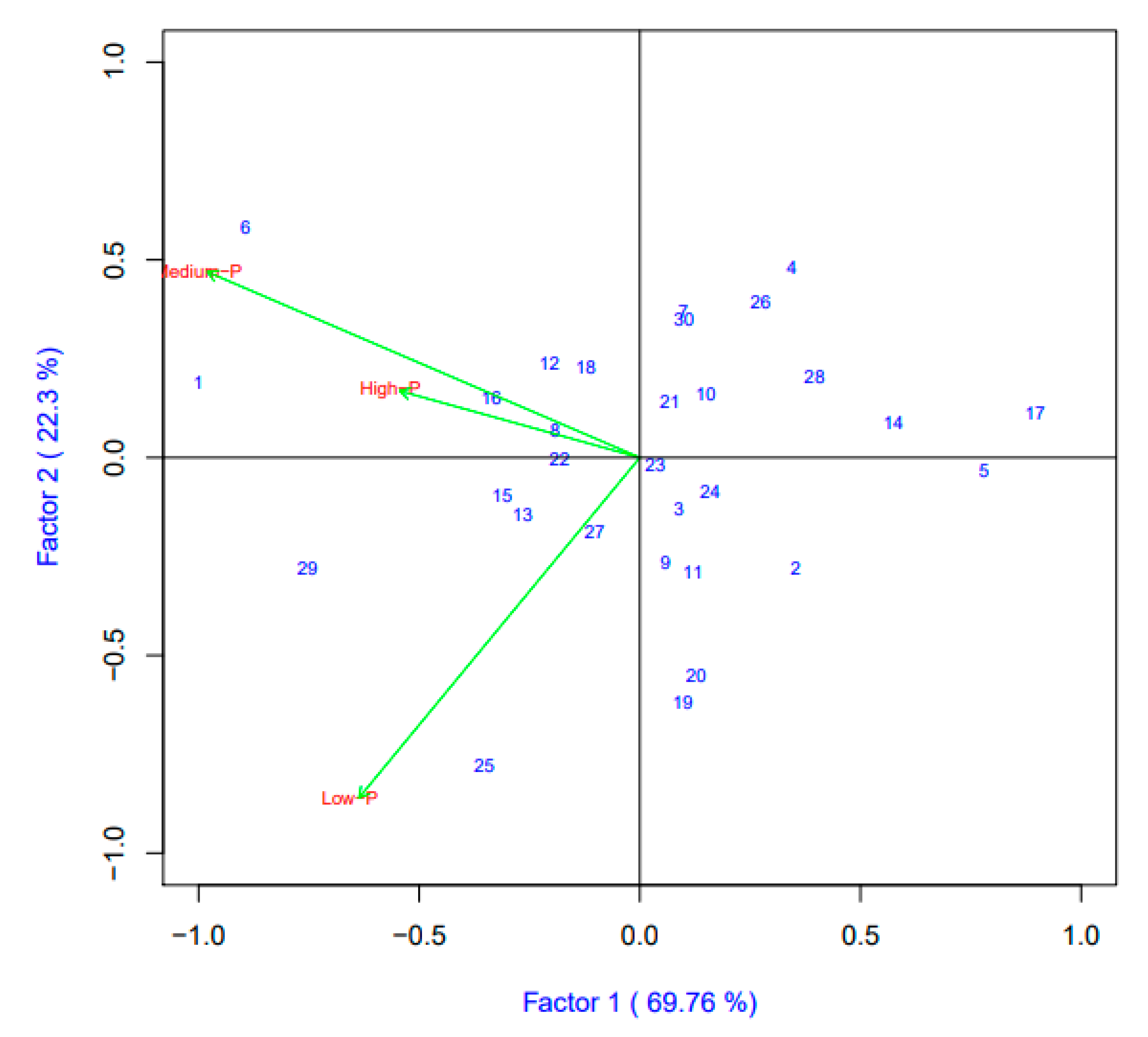
3.3. Interaction of Genotypes and Phosphorus Rates on Grain Yield and Zinc Content with Biplot Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- White, P.J.; Broadley, M.R. Biofortification of crops with seven mineral elements often lacking in human diets - iron, zinc, copper, calcium, magnesium, selenium and iodine. New Phytol. 2009, 182, 49–84. [Google Scholar] [CrossRef]
- Philipo, M.; Ndakidemi, P.A.; Mbega, E.R. Environmental and genotypes influence on seed iron and zinc levels of landraces and improved varieties of common bean (Phaseolus vulgaris L.) in Tanzania. Ecol. Genet. Genom. 2020, 15, 100056. [Google Scholar] [CrossRef]
- Hambidge, M. Human Zinc Deficiency. J. Nutr. 2000, 130, 1344S–1349S. [Google Scholar] [CrossRef]
- Brown, K.H.; Wuehler, S.E.; Peerson, J.M. The importance of zinc in human nutrition and estimation of the global prevalence of zinc defi ciency. Food Nutr. Bull. 2001, 22, 13. [Google Scholar] [CrossRef] [Green Version]
- Deshpande, J.D.; Joshi, M.M.; Giri, P.A. Zinc in Human Nutrition And Health 1. Int. J. Med. Sci. Public Health 2013, 2. [Google Scholar] [CrossRef] [Green Version]
- Abate, T.; Alene, A.D.; Bergvinson, D.; Shiferaw, B.; Silim, S.; Orr, A.; Asfaw, S. Tropical Grain Legumes in Africa and South Asia: Knowledge and Opportunities; International Crops Research Institute for the Semi-Arid Tropics; ICRISAT-Nairobi: Nairobi City, Kenya, 2011. [Google Scholar]
- Boukar, O.; Belko, N.; Chamarthi, S.; Togola, A.; Batieno, J.; Owusu, E.; Haruna, M.; Diallo, S.; Umar, M.L.; Olufajo, O.; et al. Cowpea (Vigna unguiculata): Genetics, genomics and breeding. Plant Breed. 2018, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Langyintuo, A.S.; Lowenberg-Deboer, J.; Faye, M.; Lambert, D.; Ibro, G.; Moussa, B.; Kergna, A.; Kushwaha, S.; Musa, S.; Ntoukam, G. Cowpea supply and demand in West and Central Africa. Field Crop. Res. 2003, 82, 215–231. [Google Scholar] [CrossRef]
- Phillips, R.D.; Mcwatters, K.H.; Chinnan, M.S.; Hung, Y.-C.; Beuchat, L.R.; Sefa-Dedeh, S.; Sakyi-Dawson, E.; Ngoddy, P.; Nnanyelugo, D.; Enwere, J.; et al. Utilization of cowpeas for human food. Field Crop. Res. 2003, 82, 193–213. [Google Scholar] [CrossRef]
- Samireddypalle, A.; Boukar, O.; Grings, E.; Fatokun, C.A.; Kodukula, P.; Devulapalli, R.; Okike, I.; Blümmel, M. Cowpea and Groundnut Haulms Fodder Trading and Its Lessons for Multidimensional Cowpea Improvement for Mixed Crop Livestock Systems in West Africa. Front. Plant Sci. 2017, 08, 30. [Google Scholar] [CrossRef] [Green Version]
- Snapp, S.; Rahmanian, M.; Batello, C. Pulse Crops for Sustainable Farms in Sub-Saharan Africa; FAO: Rome, Italy, 2018. [Google Scholar]
- Horn, L.; Shimelis, H.; Laing, M. Participatory appraisal of production constraints, preferred traits and farming system of cowpea in the northern Namibia: Implications for breeding. Legum. Res. 2014, 38, 691–700. [Google Scholar] [CrossRef] [Green Version]
- Magani, I.E.; Kuchinda, C. Effect of phosphorus fertilizer on growth, yield and crude protein content of cowpea (Vigna unguiculata [L.] Walp) in Nigeria. J. Appl. Biosci. 2009, 23, 1387–1393. [Google Scholar]
- Mohammed, S.B.; Mohammad, I.F.; Pangirayi, T.B.; Vernon, G.; Dzidzienyo, D.K.; Umar, M.L.; Umar, S. Farmers’ knowledge, perception, and use of phosphorus fertilization for cowpea production in Northern Guinea Savannah of Nigeria. Heliyon 2020, 6. [Google Scholar] [CrossRef]
- Genc, Y.; Humphries, J.M.; Lyons, G.H.; Graham, R.D. Exploiting genotypic variation in plant nutrient accumulation to alleviate micronutrient deficiency in populations. J. Trace Elem. Med. Biol. 2005, 18, 319–324. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, A.; Goufo, P.; Barros, A.; Domínguez-Perles, R.; Trindade, H.; Rosa, E.A.S.; Ferreira, L.; Rodrigues, M. Cowpea (Vigna unguiculata L. Walp), a renewed multipurpose crop for a more sustainable agri-food system: Nutritional advantages and constraints: Nutritional aptitude of cowpea. J. Sci. Food Agric. 2016, 96, 2941–2951. [Google Scholar] [CrossRef] [PubMed]
- Cichy, K.A.; Caldas, G.V.; Snapp, S.S.; Blair, M.W. QTL Analysis of Seed Iron, Zinc, and Phosphorus Levels in an Andean Bean Population. Crop Sci. 2009, 49. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.-Q.; Deng, Y.; Chen, R.-Y.; Cui, Z.-L.; Chen, X.-P.; Yost, R.; Zhang, F.-S.; Zou, C.-Q. The reduction in zinc concentration of wheat grain upon increased phosphorus-fertilization and its mitigation by foliar zinc application. Plant Soil 2012, 361, 143–152. [Google Scholar] [CrossRef]
- Blair, M.W.; Medina, J.I.; Astudillo, C.; Rengifo, J.; Beebe, S.E.; Machado, G.; Graham, R. QTL for seed iron and zinc concentration and content in a Mesoamerican common bean (Phaseolus vulgaris L.) population. Theor. Appl. Genet. 2010, 121, 1059–1070. [Google Scholar] [CrossRef]
- Manzeke, M.G.; Mtambanengwe, F.; Nezomba, H.; Watts, M.J.; Broadley, M.R.; Mapfumo, P. Zinc fertilization increases productivity and grain nutritional quality of cowpea (Vigna unguiculata [L.] Walp.) under integrated soil fertility management. Field Crop. Res. 2017, 213, 231–244. [Google Scholar] [CrossRef]
- White, P.J.; Broadley, M.R. Biofortifying crops with essential mineral elements. Trends Plant Sci. 2005, 10, 586–593. [Google Scholar] [CrossRef]
- Gaddameedi, A.; Ravikiran, K.T.; Mohammed, R.; Phuke, R.M.; Sadaiah, K.; Kavi Kishor, P.B.; Ashok Kumar, A. Inheritance studies on grain iron and zinc concentration and agronomic traits in sorghum [Sorghum bicolor (L.) Moench]. J. Cereal Sci. 2018, 83, 252–258. [Google Scholar] [CrossRef]
- ICRISAT. Enhancing Cowpea Productivity and Production in Drought-Prone Areas of Sub-Saharan Africa; ICRISAT: Patancheruvu, India, 2017. [Google Scholar]
- Olufajo, O.O.; Singh, B.B. Advances in cowpea cropping systems research. In Challenges and Opportunities for Enhancing Sustainable Cowpea Production; Fatokun, C.A., Tarawali, S.A., Singh, B.B., Kormawa, P.M., Tamò, M., Eds.; International Institute of Tropical Agriculture, IITA: Ibadan, Nigeria, 2002; pp. 267–277. [Google Scholar]
- Simpson, R.J.; Oberson, A.; Culvenor, R.A.; Ryan, M.H.; Veneklaas, E.J.; Lambers, H.; Lynch, J.P.; Ryan, P.R.; Delhaize, E.; Smith, F.A.; et al. Strategies and agronomic interventions to improve the phosphorus-use efficiency of farming systems. Plant Soil 2011, 349, 89–120. [Google Scholar] [CrossRef]
- Oladiran, O.; Olajire, F.; Abaidoo, R.C.; Nnenna, I. Phosphorus Response Efficiency in Cowpea Genotypes. J. Agric. Sci. 2012, 4, 81–90. [Google Scholar] [CrossRef]
- Sanginga, N.; Lyasse, O.; Singh, B.B. Phosphorus use efficiency and nitrogen balance of cowpea breeding lines in a low P soil of the derived savanna zone in West Africa. Plant Soil 2000, 220, 119–128. [Google Scholar] [CrossRef]
- Gyan-Ansah, S.; Adu-Dapaah, H.; Kumaga, F.; Gracen, V.; Nartey, F.K.K. Evaluation of cowpea (Vigna unguiculata L. Walp.) genotypes for phosphorus use efficiency. Acta Hortic. 2016, 373–380. [Google Scholar] [CrossRef]
- Adusei, G.; Gaiser, T.; Boukar, O.; Fatokun, C. Growth and yield responses of cowpea genotypes to soluble and rock P fertilizers on acid, highly weathered soil from humid tropical West Africa. Int. J. Biol. Chem. Sci. 2016, 10, 1493–1507. [Google Scholar] [CrossRef] [Green Version]
- Saidou, A.K.; Singh, B.B.; Abaidoo, R.C.; Iwuafor, E.N.O.; Sanginga, N. Response of cowpea lines to low Phosphorus tolerance and response to external application of P. Afr. J. Microbiol. Res. 2012, 6, 5479–5485. [Google Scholar] [CrossRef]
- Loneragan, J.F.; Webb, M.J. Interactions Between Zinc and Other Nutrients Affecting the Growth of Plants. In Zinc in Soils and Plants; Robson, A.D., Ed.; Springer Netherlands: Dordrecht, The Netherlands, 1993; pp. 119–134. [Google Scholar]
- Ayeni, D.F.; Ikwebe, J.; Onyezili, F.N. Iron and zinc levels in Vigna unguiculata (L.) Walp under varying phosphorus and fixed nitrogen treatment conditions. Food Energy Secur. 2018, 7, e00141. [Google Scholar] [CrossRef]
- Santos, C.A.F.; Boiteux, L.S. Breeding biofortifed cowpea lines for semi-arid tropical areas by combining higher seed protein and mineral levels. Genet. Mol. Res. 2013, 12, 6782–6789. [Google Scholar] [CrossRef] [PubMed]
- Ashok Kumar, A.; Reddy, B.V.S.; Ramaiah, B.; Sahrawat, K.L.; Pfeiffer, W.H. Gene effects and heterosis for grain iron and zinc concentration in sorghum [Sorghum bicolor (L.) Moench]. Field Crop. Res. 2013, 146, 86–95. [Google Scholar] [CrossRef] [Green Version]
- Huynh, B.-L.; Ehlers, J.D.; Close, T.J.; Cis, N.; Drabo, I.; Boukar, O.; Lucas, M.R.; Wanamaker, S.; Pottorff, M.; Roberts, P.A. Enabling Tools for Modern Breeding of Cowpea for Biotic Stress Resistance. In Translational Genomics for Crop Breeding, Volume I: Biotic Stress; Varshney, R.K., Tuberosa, R., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2013; pp. 183–199. [Google Scholar]
- Boukar, O.; Fatokun, C.A.; Huynh, B.-L.; Roberts, P.A.; Close, T.J. Genomic Tools in Cowpea Breeding Programs: Status and Perspectives. Front. Plant Sci. 2016, 7, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Muñoz-Amatriaín, M.; Mirebrahim, H.; Xu, P.; Wanamaker, S.I.; Luo, M.C.; Alhakami, H.; Alpert, M.; Atokple, I.; Batieno, B.J.; Boukar, O.; et al. Genome resources for climate-resilient cowpea, an essential crop for food security. Plant J. 2017, 89, 1042–1054. [Google Scholar] [CrossRef] [Green Version]
- Bationo, A.; Ntare, B.R.; Tarawali, S.A.; Tabo, R. Soil fertility management and cowpea production in the semiarid tropics. In Challenges and Opportunities for Enhancing Sustainable Cowpea Production; Fatokun, C.A., Tarawali, S.A., Singh, B.B., Kormawa, P.M., Tamo, M., Eds.; International Institute of Tropical Agriculture, IITA: Ibadan, Nigeria, 2002; pp. 301–318. [Google Scholar]
- Rothe, C.J. Breeding for Tolerance of Cowpea to Low Phosphorus Soil Conditions through Physiological and Genetic Studies. Ph.D. Thesis, Texas A&M University, College Station, TX, USA, 2014. [Google Scholar]
- Pacheco, Á.; Vargas, M.; Alvarado, G.; Rodríguez, F.; Crossa, J.; Burgueño, J. GEA-R (Genotype x Environment Analysis with R for Windows); Version 4.1; CIMMYT: Mexico City, Mexico, 2015. [Google Scholar]
- Kolawole, G.O.; Tian, G.; Singh, B.B. Differential response of cowpea lines to aluminum and phosphorus application. J. Plant Nutr. 2008, 23, 731–740. [Google Scholar] [CrossRef]
- Krasilnikoff, G.; Gahoonia, T.; Nielsen, N.E. Variation in phosphorus uptake efficiency by genotypes of cowpea (Vigna unguiculata) due to differences in root and root hair length and induced rhizosphere processes. Plant Soil 2003, 251, 83–91. [Google Scholar] [CrossRef]
- Safaya, N.M.; Singh, B. Differential susceptibility of two varieties of cowpea (Vigna unguiculata (L) Walp) to phosphorus-induced zinc deficiency. Plant Soil 1977, 48, 279–290. [Google Scholar] [CrossRef]
- Benvindo, R.N.; Prado, R.D.M.; Nóbrega, J.C.A.; Flores, R.A. Phosphorus Fertilization on the Nutrition and Yield of Cowpea Grown in an Arenosols. J. Agric. Environ. Sci 2014, 14, 434–439. [Google Scholar] [CrossRef]
- Nyoki, D.; Ndakidemi, P.A. Influence of Bradyrhizobium japonicum and Phosphorus on Micronutrient Uptake in Cowpea. A Case Study of Zinc (Zn), Iron (Fe), Copper (Cu) and Manganese (Mn). Am. J. Plant Sci. 2014, 5, 427–435. [Google Scholar] [CrossRef] [Green Version]
- Pongrac, P.; Fischer, S.; Thompson, J.A.; Wright, G.; White, P.J. Early Responses of Brassica oleracea Roots to Zinc Supply Under Sufficient and Sub-Optimal Phosphorus Supply. Front. Plant Sci. 2020, 10. [Google Scholar] [CrossRef] [PubMed]
- Pongrac, P.; Castillo-Michel, H.; Reyes-Herrera, J.; Hancock, R.D.; Fischer, S.; Kelemen, M.; Thompson, J.A.; Wright, G.; Likar, M.; Broadley, M.R.; et al. Effect of phosphorus supply on root traits of two Brassica oleracea L. genotypes. BMC Plant Biol. 2020, 20, 368. [Google Scholar] [CrossRef]
- Boawn, L.C.; Brown, J.C. Further Evidence for a P-Zn Imbalance in Plants. Soil Sci. Soc. Am. J. 1968, 32, 94–97. [Google Scholar] [CrossRef]
- Boawn, L.C.; Leggett, G.E. Phosphorus and Zinc Concentrations in Russet Burbank Potato Tissues in Relation to Development of Zinc Deficiency Symptoms. Soil Sci. Soc. Am. J. 1964, 28, 229–232. [Google Scholar] [CrossRef]
- Millikan, C. Effects of different levels of zinc and phosphorus on the growth of subterranean clover (Trifolium subterraneum L.). Aust. J. Agric. Res. 1963, 14, 180–205. [Google Scholar] [CrossRef]
- Ambler, J.E.; Brown, J.C. Cause of Differential Susceptibility to Zinc Deficiency in Two Varieties of Navy Beans (Phaseolus vulgaris L.). Agron. J. 1969, 61, 41–43. [Google Scholar] [CrossRef]
- Bogdanovic, D.; Ubavic, M.; Cuvardic, M. Effect of phosphorus fertilization on Zn and Cd contents in soil and corn plants. Nutr. Cycl. Agroecosyst. 1999, 54, 49–56. [Google Scholar] [CrossRef]
- Imran, M.; Rehim, A.; Sarwar, N.; Hussain, S. Zinc bioavailability in maize grains in response of phosphorous-zinc interaction. J. Plant Nutr. Soil Sci. 2016, 179, 60–66. [Google Scholar] [CrossRef]
- Ova, E.A.; Kutman, U.B.; Ozturk, L.; Cakmak, I. High phosphorus supply reduced zinc concentration of wheat in native soil but not in autoclaved soil or nutrient solution. Plant Soil 2015, 393, 147–162. [Google Scholar] [CrossRef]
- Ryan, M.H.; McInerney, J.K.; Record, I.R.; Angus, J.F. Zinc bioavailability in wheat grain in relation to phosphorus fertiliser, crop sequence and mycorrhizal fungi. J. Sci. Food Agric. 2008, 88, 1208–1216. [Google Scholar] [CrossRef]
- Singh, J.P.; Karamanos, R.E.; Stewart, J.W.B. The mechanism of phosphorus-induced zinc deficiency in bean (Phaseolus vulgaris L.). Can. J. Soil Sci. 1988, 68, 345–358. [Google Scholar] [CrossRef]
- Fageria, V.D. Nutrient interactions in crop plants. J. Plant Nutr. 2001, 24, 1269–1290. [Google Scholar] [CrossRef]
- Liu, D.-Y.; Zhang, W.; Liu, Y.-M.; Chen, X.-P.; Zou, C.-Q. Soil Application of Zinc Fertilizer Increases Maize Yield by Enhancing the Kernel Number and Kernel Weight of Inferior Grains. Front. Plant Sci. 2020, 11, 188. [Google Scholar] [CrossRef]
- Rose, T.J.; Impa, S.M.; Rose, M.T.; Pariasca-Tanaka, J.; Mori, A.; Heuer, S.; Johnson-Beebout, S.E.; Wissuwa, M. Enhancing phosphorus and zinc acquisition efficiency in rice: A critical review of root traits and their potential utility in rice breeding. Ann. Bot. 2013, 112, 331–345. [Google Scholar] [CrossRef] [Green Version]
- Qaswar, M.; Hussain, S.; Rengel, Z. Zinc fertilisation increases grain zinc and reduces grain lead and cadmium concentrations more in zinc-biofortified than standard wheat cultivar. Sci. Total Environ. 2017, 605–606, 454–460. [Google Scholar] [CrossRef]
- Oseni, T.O. Growth and zinc uptake of sorghum and cowpea in response to phosphorus and zinc fertilization. World J. Agric. Sci. 2009, 5, 670–674. [Google Scholar]
- Abdul Rahman, N.; Larbi, A.; Kotu, B.; Marthy Tetteh, F.; Hoeschle-Zeledon, I. Does Nitrogen Matter for Legumes? Starter Nitrogen Effects on Biological and Economic Benefits of Cowpea (Vigna unguiculata L.) in Guinea and Sudan Savanna of West Africa. Agronomy 2018, 8, 120. [Google Scholar] [CrossRef] [Green Version]
- Ngwene, B.; George, E.; Claussen, W.; Neumann, E. Phosphorus uptake by cowpea plants from sparingly available or soluble sources as affected by nitrogen form and arbuscular-mycorrhiza-fungal inoculation. J. Plant Nutr. Soil Sci. 2010, 173, 353–359. [Google Scholar] [CrossRef]
- Nishigaki, T.; Tsujimoto, Y.; Rinasoa, S.; Rakotoson, T.; Andriamananjara, A.; Razafimbelo, T. Phosphorus uptake of rice plants is affected by phosphorus forms and physicochemical properties of tropical weathered soils. Plant Soil 2019, 435, 27–38. [Google Scholar] [CrossRef] [Green Version]
- Amin, S.; Che, F.I.; Hanafi, M.M.; Hamed, Z.; Zaharah, A.R. Phosphorus and Zinc Uptake and Their Interaction Effect on Dry Matter and Chlorophyll Content of Sweet Corn (Zea mays var. Saccharat). J. Agron. 2013, 12, 187–192. [Google Scholar]
- Xue, Y.; Xia, H.; Christie, P.; Zhang, Z.; Li, L.; Tang, C. Crop acquisition of phosphorus, iron and zinc from soil in cereal/legume intercropping systems: A critical review. Ann. Bot. 2016, 117, 363–377. [Google Scholar] [CrossRef]
- Ukalski, K.; Klisz, M. Application of GGE biplot graphs in multi-environment trials on selection of forest trees. Folia For. Pol. Ser. A 2016, 58, 228–239. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Traits | Definition and Abbreviations |
|---|---|
| Grain yield, kg/ha | Grain yield in kg per hectare from the low, medium, and high phosphorus treatments (yield.LP, yield.MP, and yield.HP) |
| Grain phosphorus concentration, mg/kg | Phosphorus concentration determined from grain samples from the low, medium, and high phosphorus treatments (grainP.LP, grainP.MP, and grainP.HP) |
| Grain zinc content, ppm | The zinc content of grain samples from the low, medium, and high phosphorus treatments (grainZn.LP, grainZn.MP, and grainZn.HP) |
| Plant height, cm | The height of the plant measured in centimeters from the soil surface to the base of the top leaf on the main stem (average per genotype) (pht.LP, pht.MP, and pht.HP) |
| Root dry weight, gram | Weight of dry root, per plot of low, medium, and high phosphorus treatments (rdwt.LP, rdwt.MP, and rdwt.HP) |
| Root phosphorus concentration, mg/kg | Phosphorus concentration determined from root samples from the low, medium, and high phosphorus treatments (rootP.LP, rootP.MP, and rootP.HP) |
| Root zinc content, ppm | The zinc content of root samples from the low, medium, and high phosphorus treatments (rootZn.LP, rootZn.MP, and rootZn.HP) |
| Shoot dry weight, gram | Weight of dry shoot, per plot of low, medium, and high phosphorus treatments (sdwt.LP, sdwt.MP, and sdwt.HP) |
| Shoot phosphorus concentration, mg/kg | Phosphorus concentration determined from shoot samples from the low, medium, and high phosphorus treatments (shootP.LP, shootP.MP, and shootP.HP) |
| Shoot zinc content, ppm | The zinc content of shoot samples from the low, medium, and high phosphorus treatments (shootZn.LP, shootZn.MP, and shootZn.HP) |
| Soil phosphorus content, mg/kg | Plant available phosphorus from the low, medium, and high phosphorus plots (soilP.LP, soilP.MP, and soil.P.HP), determined using the Bray-I method |
| Screenhouse | Field | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Phosphorus Levels | Pht (cm) | Sdwt (g) | ShootP (mg/kg) | ShootZn (ppm) | Sdwt (g) | ShootP (mg/kg) | ShootZn (ppm) | Yield (kg/ha) | GrainP (mg/kg) | GrainZn (ppm) |
| LP | 14.5 | 0.4 | 942.4 | 70.7 | 20.6 | 1132.4 | 103.9 | 294.1 | 2106.8 | 34.3 |
| MP | 14.3 | 0.5 | 1054.1 | 73.4 | 38.9 | 1162.9 | 96.8 | 558.9 | 2286.4 | 31.5 |
| HP | 22.2 | 2.2 | 1275.9 | 68.3 | 94.5 | 1183.7 | 88.7 | 1406.6 | 2368.2 | 29.3 |
| RD (%) | 34.6 | 81.8 | 26.1 | −3.5 | 78.2 | 4.3 | −17.1 | 79.1 | 11.0 | −17.1 |
| Genotypes | Sdwt .HP | Sdwt .MP | Sdwt .LP | Yield .HP | Yield .MP | Yield .LP | GrainZn .HP | GrainZn .MP | GrainZn .LP |
|---|---|---|---|---|---|---|---|---|---|
| IT89KD-288 | 98.6 | 33.0 | 19.0 | 1475.0 | 599.2 | 99.9 | 43.7 | 39.2 | 36.2 |
| IAR-48 | 116.5 | 32.0 | 21.9 | 1547.1 | 421.9 | 388.7 | 41.1 | 22.6 | 26.3 |
| IT82E-18 | 90.8 | 49.0 | 21.3 | 1171.1 | 643.9 | 519.2 | 40.7 | 37.2 | 31.7 |
| IT97K-499-35 | 80.3 | 48.0 | 20.2 | 1261.1 | 925.1 | 289.3 | 39.5 | 34.6 | 30.3 |
| UCR779 | 93.6 | 13.0 | 19.5 | 1197.1 | 232.0 | 22.2 | 31.4 | 19.0 | 21.1 |
| DanMisra | 127.6 | 27.0 | 22.1 | 2105.5 | 459.9 | 571.9 | 31.1 | 24.2 | 39.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mohammed, S.B.; Dzidzienyo, D.K.; Yahaya, A.; L. Umar, M.; Ishiyaku, M.F.; Tongoona, P.B.; Gracen, V. High Soil Phosphorus Application Significantly Increased Grain Yield, Phosphorus Content but Not Zinc Content of Cowpea Grains. Agronomy 2021, 11, 802. https://doi.org/10.3390/agronomy11040802
Mohammed SB, Dzidzienyo DK, Yahaya A, L. Umar M, Ishiyaku MF, Tongoona PB, Gracen V. High Soil Phosphorus Application Significantly Increased Grain Yield, Phosphorus Content but Not Zinc Content of Cowpea Grains. Agronomy. 2021; 11(4):802. https://doi.org/10.3390/agronomy11040802
Chicago/Turabian StyleMohammed, Saba B., Daniel K. Dzidzienyo, Adama Yahaya, Muhammad L. Umar, Mohammad F. Ishiyaku, Pangirayi B. Tongoona, and Vernon Gracen. 2021. "High Soil Phosphorus Application Significantly Increased Grain Yield, Phosphorus Content but Not Zinc Content of Cowpea Grains" Agronomy 11, no. 4: 802. https://doi.org/10.3390/agronomy11040802