
Fusarium Head Blight and Associated Mycotoxins in Grains and Straw of Barley: Influence of Agricultural Practices

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling Procedure, Sample Origin, and Questionnaire
2.2. Identification of Fusarium Head Blight Causing Species with Seed Health Tests
2.3. Extraction of Fusarium graminearum DNA and Quantitative PCR
2.4. Analysis of Fusarium mycotoxins with LC-MS/MS
2.5. Data Analysis
3. Results
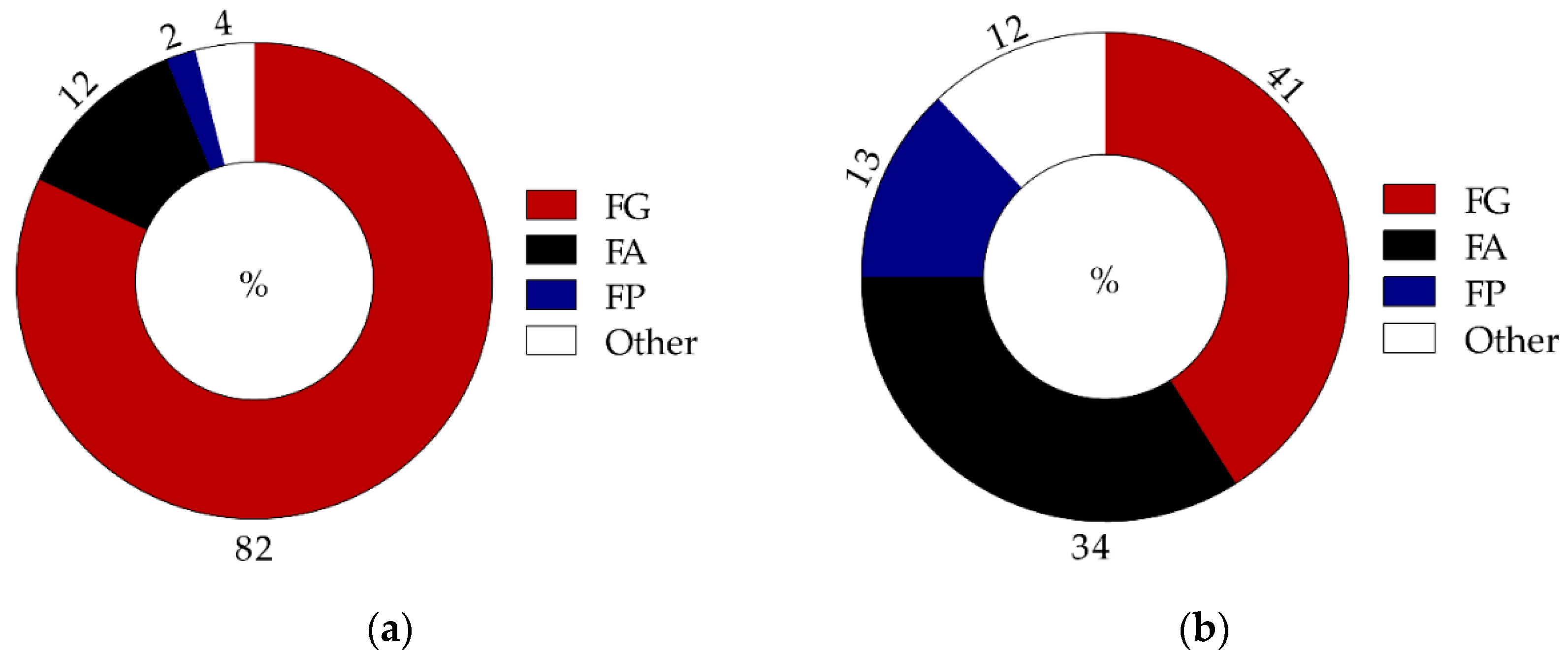
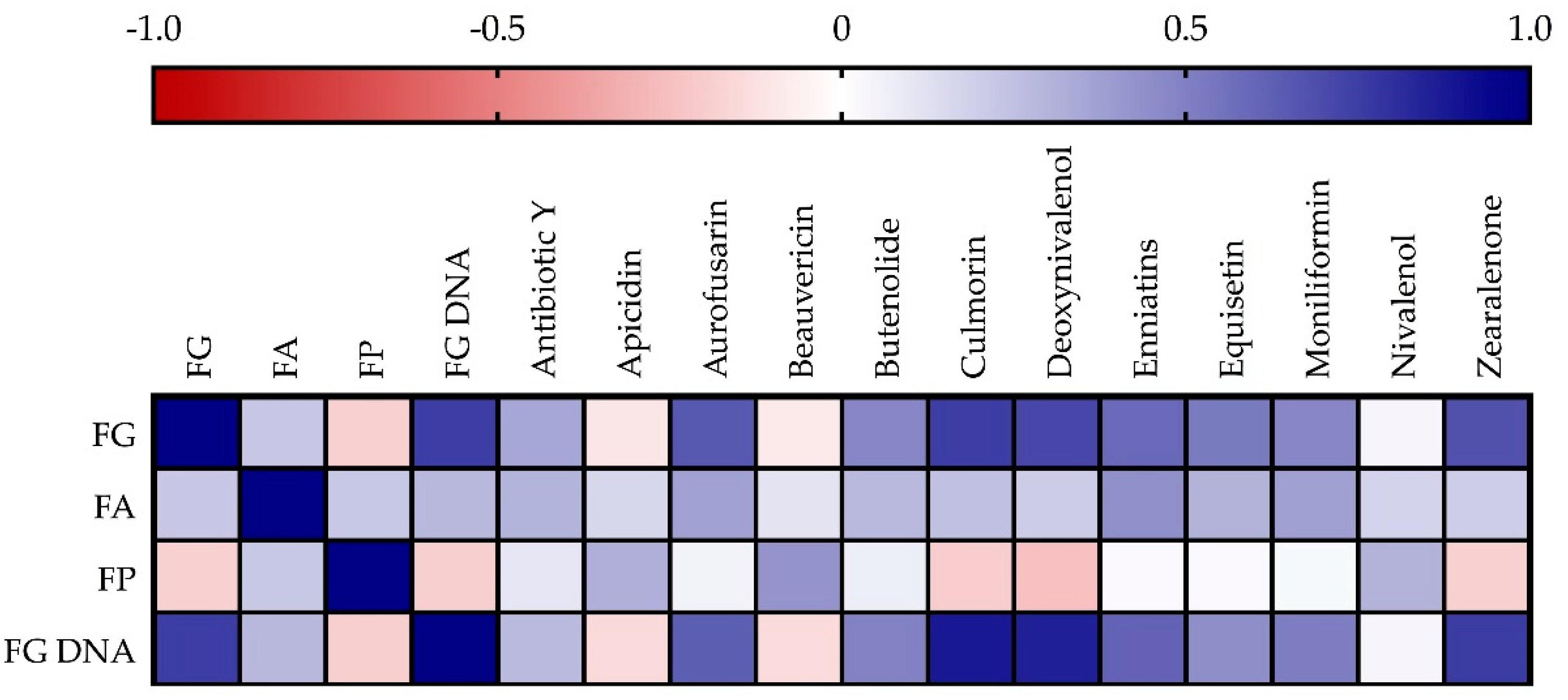
3.1. Incidence of Fusarium Head Blight Causing Species in Grains and Correlations
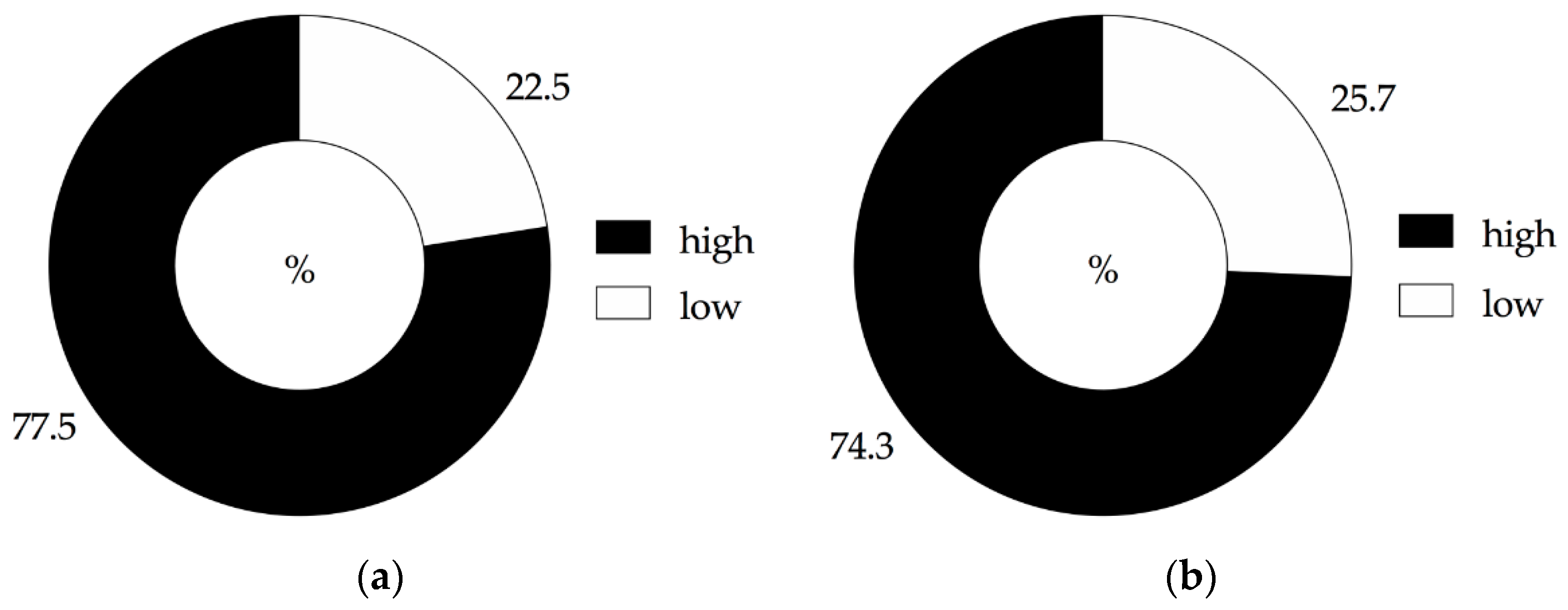
3.2. Influence of Agricultural Practices on Fusarium Head Blight Species Incidence and Fusarium graminearum DNA Amount in Grains
3.3. Influence of Agricultural Practices on Fusarium mycotoxins Content in Grains
3.4. Influence of Agricultural Practices on Fusarium graminearum DNA Amount and Fusarium mycotoxins Content in Straw
4. Discussion
4.1. Incidence of Fusarium Head Blight Causing Species in Grains and Correlations with Mycotoxins
4.2. Influence of Agricultural Practices Performed before and during Barley Production
4.2.1. Before Barley Production
4.2.2. During Barley Production
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAOSTAT. Database. Available online: http://www.fao.org/faostat/en/#data (accessed on 8 January 2021).
- Agriopoulou, S.; Stamatelopoulou, E.; Varzakas, T. Advances in occurrence, importance, and mycotoxin control strategies: Prevention and detoxification in foods. Foods 2020, 9, 137. [Google Scholar] [CrossRef] [PubMed]
- Alshannaq, A.; Yu, J.H. Occurrence, toxicity, and analysis of major mycotoxins in food. Int. J. Environ. Res. Public Health 2017, 14, 632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marin, S.; Ramos, A.J.; Cano-Sancho, G.; Sanchis, V. Mycotoxins: Occurrence, toxicology, and exposure assessment. Food Chem. Toxicol. 2013, 60, 218–237. [Google Scholar] [CrossRef]
- EFSA. Deoxynivalenol in food and feed: Occurrence and exposure. EFSA J. 2013, 11, 3379–3435. [Google Scholar]
- D’Mello, J.P.F.; Placinta, C.M.; Macdonald, A.M.C. Fusarium mycotoxins: A review of global implications for animal health, welfare and productivity. Anim. Feed Sci. Technol. 1999, 80, 183–205. [Google Scholar] [CrossRef] [Green Version]
- Drakopoulos, D.; Sulyok, M.; Krska, R.; Logrieco, A.F.; Vogelgsang, S. Raised concerns about the safety of barley grains and straw: A Swiss survey reveals a high diversity of mycotoxins and other fungal metabolites. Food Control 2021, 125, 107919. [Google Scholar] [CrossRef]
- Jestoi, M. Emerging Fusarium-mycotoxins fusaproliferin, beauvericin, enniatins, and moniliformin—A review. Crit. Rev. Food Sci. Nutr. 2008, 48, 21–49. [Google Scholar] [CrossRef] [PubMed]
- Beccari, G.; Caproni, L.; Tini, F.; Uhlig, S.; Covarelli, L. Presence of Fusarium species and other toxigenic fungi in malting barley and multi-mycotoxin analysis by liquid chromatography–high-resolution mass spectrometry. J. Agric. Food Chem. 2016, 64, 4390–4399. [Google Scholar] [CrossRef]
- Schöneberg, T.; Martin, C.; Wettstein, F.E.; Bucheli, T.D.; Mascher, F.; Bertossa, M.; Musa, T.; Keller, B.; Vogelgsang, S. Fusarium and mycotoxin spectra in Swiss barley are affected by various cropping techniques. Food Addit. Contam. Part A 2016, 33, 1608–1619. [Google Scholar] [CrossRef] [Green Version]
- Shah, L.; Ali, A.; Yahya, M.; Zhu, Y.; Wang, S.; Si, H.; Rahman, H.; Ma, C. Integrated control of Fusarium head blight and deoxynivalenol mycotoxin in wheat. Plant Pathol. 2018, 67, 532–548. [Google Scholar] [CrossRef]
- Vogelgsang, S.; Beyer, M.; Pasquali, M.; Jenny, E.; Musa, T.; Bucheli, T.D.; Wettstein, F.E.; Forrer, H.-R. An eight-year survey of wheat shows distinctive effects of cropping factors on different Fusarium species and associated mycotoxins. Eur. J. Agron. 2019, 105, 62–77. [Google Scholar] [CrossRef]
- Xu, X.M.; Nicholson, P. Community ecology of fungal pathogens causing wheat head blight. Annu. Rev. Phytopathol. 2009, 47, 83–103. [Google Scholar] [CrossRef]
- Wegulo, S.N.; Baenziger, P.S.; Hernandez Nopsa, J.; Bockus, W.W.; Hallen-Adams, H. Management of Fusarium head blight of wheat and barley. Crop Protect. 2015, 73, 100–107. [Google Scholar] [CrossRef]
- Paul, P.A.; Lipps, P.E.; Hershman, D.E.; McMullen, M.P.; Draper, M.A.; Madden, L.V. Efficacy of triazole-based fungicides for Fusarium Head Blight and deoxynivalenol control in wheat: A multivariate meta-analysis. Phytopathology 2008, 98, 999–1011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simpson, D.R.; Weston, G.E.; Turner, J.A.; Jennings, P.; Nicholson, P. Differential control of head blight pathogens of wheat by fungicides and consequences for mycotoxin contamination of grain. Eur. J. Plant Pathol. 2001, 107, 421–431. [Google Scholar] [CrossRef]
- Anonymous. Swiss Agricultural Policies—Objectives, Tools, Prospects. Available online: https://www.cbd.int/financial/pes/swiss-pesagriculturalpolicy.pdf (accessed on 14 December 2020).
- Jäggi, E. Support schemes and agriculture in Switzerland. In Proceedings of the Concerted Action Seminar: Potential for Environmental Cross-Compliance Matters, Roskilde, Denmark, 24–25 November 2003. [Google Scholar]
- Anonymous. Bio Suisse. Available online: https://www.bio-suisse.ch/en/home.php (accessed on 14 December 2020).
- Vogelgsang, S.; Sulyok, M.; Hecker, A.; Jenny, E.; Krska, R.; Schuhmacher, R.; Forrer, H.R. Toxigenicity and pathogenicity of Fusarium poae and Fusarium avenaceum on wheat. Eur. J. Plant Pathol. 2008, 122, 265–276. [Google Scholar] [CrossRef]
- Leslie, J.F.; Summerell, B.A. The Fusarium Laboratory Manual; Blackwell Publishing: Oxford, UK, 2006; 388p. [Google Scholar]
- Drakopoulos, D.; Kägi, A.; Gimeno, A.; Six, J.; Jenny, E.; Forrer, H.-R.; Musa, T.; Meca, G.; Vogelgsang, S. Prevention of Fusarium head blight infection and mycotoxins in wheat with cut-and-carry biofumigation and botanicals. Field Crop Res. 2020, 246, 107681. [Google Scholar] [CrossRef]
- Brandfass, C.; Karlovsky, P. Simultaneous detection of Fusarium culmorum and F. graminearum in plant material by duplex PCR with melting curve analysis. BMC Microbiol. 2006, 6, 4. [Google Scholar] [CrossRef] [Green Version]
- Sulyok, M.; Stadler, D.; Steiner, D.; Krska, R. Validation of an LC-MS/MS-based dilute-and-shoot approach for the quantification of > 500 mycotoxins and other secondary metabolites in food crops: Challenges and solutions. Anal. Bioanal. Chem. 2020, 412, 2607–2620. [Google Scholar] [CrossRef] [Green Version]
- Asuero, A.G.; Sayago, A.; González, A.G. The correlation coefficient: An overview. Crit. Rev. Anal. Chem. 2006, 36, 41–59. [Google Scholar] [CrossRef]
- Osborne, L.E.; Stein, J.M. Epidemiology of Fusarium head blight on small-grain cereals. Int. J. Food Microbiol. 2007, 119, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Schöneberg, T.; Jenny, E.; Wettstein, F.E.; Bucheli, T.D.; Mascher, F.; Bertossa, M.; Musa, T.; Seifert, K.; Gräfenhan, T.; Keller, B.; et al. Occurrence of Fusarium species and mycotoxins in Swiss oats—Impact of cropping factors. Eur. J. Agron. 2018, 92, 123–132. [Google Scholar] [CrossRef]
- Nielsen, L.K.; Cook, D.J.; Edwards, S.G.; Ray, R.V. The prevalence and impact of Fusarium head blight pathogens and mycotoxins on malting barley quality in UK. Int. J. Food Microbiol. 2014, 179, 38–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drakopoulos, D.; Meca, G.; Torrijos, R.; Marty, A.; Kägi, A.; Jenny, E.; Forrer, H.-R.; Six, J.; Vogelgsang, S. Control of Fusarium graminearum in wheat with mustard-based botanicals: From in vitro to in planta. Front. Microbiol. 2020, 11, 1595. [Google Scholar] [CrossRef]
- Langseth, W.; Ghebremeskel, M.; Kosiak, B.; Kolsaker, P.; Miller, D. Production of culmorin compounds and other secondary metabolites by Fusarium culmorum and F. graminearum strains isolated from Norwegian cereals. Mycopathologia 2001, 152, 23–34. [Google Scholar] [CrossRef]
- Boedi, S.; Berger, H.; Sieber, C.; Münsterkötter, M.; Maloku, I.; Warth, B.; Sulyok, M.; Lemmens, M.; Schuhmacher, R.; Güldener, U.; et al. Comparison of Fusarium graminearum transcriptomes on living or dead wheat differentiates substrate-responsive and defense-responsive genes. Front. Microbiol. 2016, 7, 1113. [Google Scholar] [CrossRef]
- Beccari, G.; Prodi, A.; Tini, F.; Bonciarelli, U.; Onofri, A.; Oueslati, S.; Limayma, M.; Covarelli, L. Changes in the Fusarium head blight complex of malting barley in a three-year field experiment in Italy. Toxins 2017, 9, 120. [Google Scholar] [CrossRef] [Green Version]
- Mielniczuk, E.; Skwaryło-Bednarz, B. Fusarium Head Blight, mycotoxins and strategies for their reduction. Agronomy 2020, 10, 509. [Google Scholar] [CrossRef] [Green Version]
- Parry, D.W.; Jenkinson, P.; Mcleod, L. Fusarium ear blight (scab) in small-grain cereals—A review. Plant Pathol. 1995, 44, 207–238. [Google Scholar] [CrossRef]
- Janssen, E.M.; Mourits, M.C.M.; van der Fels-Klerx, H.J.; Lansink, A.G.J.M.O. Pre-harvest measures against Fusarium spp. infection and related mycotoxins implemented by Dutch wheat farmers. Crop Protect. 2019, 122, 9–18. [Google Scholar] [CrossRef]
- Dill-Macky, R.; Jones, R.K. The effect of previous crop residues and tillage on Fusarium Head Blight of wheat. Plant Dis. 2000, 84, 71–76. [Google Scholar] [CrossRef] [Green Version]
- Nichea, M.; Palacios, S.; Chiacchiera, S.; Sulyok, M.; Krska, R.; Chulze, S.; Torres, A.; Ramirez, M. Presence of multiple mycotoxins and other fungal metabolites in native grasses from a wetland ecosystem in Argentina intended for grazing cattle. Toxins 2015, 7, 3309–3329. [Google Scholar] [CrossRef] [Green Version]
- Twarużek, M.; Dembek, R.; Pańka, D.; Soszczyńska, E.; Zastempowska, E.; Grajewski, J. Evaluation of cytotoxicity and mould contamination of selected plants from meadows covered by the agri-environmental program. Toxins 2019, 11, 228. [Google Scholar] [CrossRef] [Green Version]
- Steinkellner, S.; Langer, I. Impact of tillage on the incidence of Fusarium spp. in soil. Plant Soil 2004, 267, 13–22. [Google Scholar] [CrossRef]
- Bottalico, A.; Perrone, G. Toxigenic Fusarium species and mycotoxins associated with head blight in small-grain cereals in Europe. Eur. J. Plant Pathol. 2002, 108, 611–624. [Google Scholar] [CrossRef]
- McMullen, M.; Bergstrom, G.; de Wolf, E.; Dill-Macky, R.; Hershman, D.; Shaner, G.; Van Sanford, D. A unified effort to fight an enemy of wheat and barley: Fusarium head blight. Plant Dis. 2012, 96, 1712–1728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Mello, J.P.F.; Macdonald, A.M.C.; Postel, D.; Dijksma, W.T.P.; Dujardin, A.; Placinta, C.M. Pesticide use and mycotoxin production in Fusarium and Aspergillus phytopathogens. Eur. J. Plant Pathol. 1998, 104, 741–751. [Google Scholar]
- Linkmeyer, A.; Hofer, K.; Rychlik, M.; Herz, M.; Hausladen, H.; Hückelhoven, R.; Hess, M. Influence of inoculum and climatic factors on the severity of Fusarium head blight in German spring and winter barley. Food Addit. Contam. Part A 2016, 33, 489–499. [Google Scholar] [CrossRef]
- Edwards, S.G.; Pirgozliev, S.R.; Hare, M.C.; Jenkinson, P. Quantification of trichothecene-producing Fusarium species in harvested grain by competitive PCR to determine efficacies of fungicides against Fusarium head blight of winter wheat. Appl. Environ. Microbiol. 2001, 67, 1575–1580. [Google Scholar] [CrossRef] [Green Version]
- Paul, P.A.; McMullen, M.P.; Hershman, D.E.; Madden, L.V. Meta-analysis of the effects of triazole-based fungicides on wheat yield and test weight as influenced by Fusarium head blight intensity. Phytopathol. 2010, 100, 160–171. [Google Scholar] [CrossRef] [Green Version]
- Cowger, C.; Arellano, C.; Marshall, D.; Fitzgerald, J. Managing Fusarium head blight in winter barley with cultivar resistance and fungicide. Plant Dis. 2019, 103, 1858–1864. [Google Scholar] [CrossRef] [Green Version]
- Edwards, S.G. Influence of agricultural practices on Fusarium infection of cereals and subsequent contamination of grain by trichothecene mycotoxins. Toxicol. Lett. 2004, 153, 29–35. [Google Scholar] [CrossRef]
- Zhang, Y.J.; Fan, P.S.; Zhang, X.; Chen, C.J.; Zhou, M.G. Quantification of Fusarium graminearum in harvested grain by real-time polymerase chain reaction to assess efficacies of fungicides on Fusarium head blight, deoxynivalenol contamination, and yield of winter wheat. Phytopathology 2009, 99, 95–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mesterházy, Á.; Bartok, T.; Lamper, C. Influence of wheat cultivar, species of Fusarium, and isolate aggressiveness on the efficacy of fungicides for control of Fusarium Head Blight. Plant Dis. 2003, 87, 1107–1115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forrer, H.-R.; Hecker, A.; Külling, C.; Kessler, P.; Jenny, E.; Krebs, H. Effect of fungicides on fusaria of wheat (in German). Agrarforschung 2000, 7, 258–263. [Google Scholar]
- Jenkinson, P.; Parry, D.W. Splash dispersal of conidia of Fusarium culmorum and Fusarium avenaceum. Mycol. Res. 1994, 98, 506–510. [Google Scholar] [CrossRef]
- Mesterházy, Á. Types and components of resistance to Fusarium head blight of wheat. Plant Breed. 1995, 114, 377–386. [Google Scholar] [CrossRef]
- Fauzi, M.; Paulitz, T. The effect of plant growth regulators and nitrogen on Fusarium head blight of the spring wheat cultivar Max. Plant Dis. 1994, 78, 289–292. [Google Scholar] [CrossRef]
- Vogelgsang, S.; Musa, T.; Bänziger, I.; Kägi, A.; Bucheli, T.D.; Wettstein, F.E.; Pasquali, M.; Forrer, H.R. Fusarium mycotoxins in Swiss wheat: A survey of growers’ samples between 2007 and 2014 shows strong year and minor geographic effects. Toxins 2017, 9, 246. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Explanatory Variable | Level and Number of Observations |
|---|---|
| Production system 1 | ÖLN (138, 129), extenso (94, 89), organic (21, 19) |
| Sowing season | autumn (235, 221), spring (18, 16) |
| Barley variety 2 | Meridian (60, 56), Cassia (39, 37), Tonic (33, 32), Semper (33, 31), Hobbit (28, 28), Etincel (13, 9), Eunova (11, 9), Wootan (9, 9), Caravan (8, 8), other (17, 17) |
| Previous crop 3 | small-grain cereals (156, 145), maize (40, 38), canola (26, 26), pasture (8, 8), other (23, 20) |
| Pre-previous crop 3 | maize (118, 109), pasture (40, 39), canola (33, 31), small-grain cereals (24, 23), other (37, 34) |
| Tillage 4 | conventional tillage (130, 120), reduced tillage (123, 117) |
| Plant height at harvest | short: x ≤ median; tall: x > median |
| Grain yield | low: x ≤ median; high: x > median |
| Fungicide type | triazole (74, 66), triazole + strobilurin (37, 36), strobilurin (13, 13) |
| Growth regulator application | yes (129, 120), no (124, 117) |
| Fertilizer type | mineral (134, 128), mineral + manure (85, 78), manure (31, 29) |
| Harvest year | 2017 (130, 122), 2016 (123, 115) |
| Incidence (%) of Fusarium graminearum, F. avenaceum, F. poae, and Microdochium spp. in barley grains |
| Amount of F. graminearum (FG) DNA (copies of FG DNA per ng total DNA) in grains and straw of barley |
| Fusarium mycotoxin content (µg kg−1) in grains and straw of barley: antibiotic Y, apicidin, aurofusarin, beauvericin, butenolide, culmorin, deoxynivalenol, enniatins 1, equisetin, moniliformin, nivalenol, and zearalenone |
| 2016 | 2017 | |||
|---|---|---|---|---|
| Fusarium Head Blight Causing Species | Mean | Maximum | Mean | Maximum |
| Fusarium graminearum | 15 | 81 | 3 | 31 |
| Fusarium avenaceum | 2 | 15 | 2 | 42 |
| Fusarium poae | 0.4 | 8 | 1 | 21 |
| Microdochium spp. | 34 | 96 | 2 | 22 |
| Response Variable | Incidence of FG (n = 207) | Incidence of FA (n = 143) | Incidence of FP (n = 64) | Incidence of M (n = 200) | Amount of FG DNA (n = 227) |
|---|---|---|---|---|---|
| Explanatory variable1 | |||||
| Production system | * | ||||
| Sowing season | * | *** | ** | * | |
| Barley variety | |||||
| Previous crop | * | *** | |||
| Pre-previous crop | ** | ||||
| Tillage | ** | ** | |||
| Plant height at harvest | * | * | |||
| Grain yield | * | ||||
| Fungicide type | |||||
| Growth regulator application | |||||
| Fertilizer type | *** | * | |||
| Harvest year | *** | *** | *** |
| Incidence of Fusarium graminearum, n = 207, median: 6% | |
| Tillage ** | conventional tillage (−1.6), reduced tillage (+1.6) |
| Harvest year *** | 2017 (−4.5), 2016 (+3.8) |
| Incidence of Fusarium avenaceum, n = 143, median: 2% | |
| Sowing season * | autumn (−0.6), spring (+1.7) |
| Incidence of Fusarium poae, n = 64, median: 2% | |
| Production system * | ÖLN (−1.2), extenso (−0.1), organic (+2.0) |
| Sowing season *** | autumn (−1.8), spring (+3.2) |
| Previous crop species * | small-grain cereals (−0.9), maize (−0.3), canola (−0.3), other (+0.2), pasture (+2.4) |
| Pre−previous crop species ** | canola (−1.4), other (−1.0), maize (−0.9), small-grain cereals (+0.2), pasture (+2.5) |
| Tillage ** | reduced tillage (−1.7), conventional tillage (+1.3) |
| Plant height at harvest * | tall (−1.5), short (+1.2) |
| Fertilizer type *** | mineral + manure (−1.8), mineral (−0.6), manure (+2.4) |
| Incidence of Microdochium species, n = 200, median: 14.5% | |
| Sowing season ** | spring (−2.2), autumn (+0.6) |
| Plant height at harvest * | short (−1.0), tall (+1.1) |
| Grain yield * | high (−1.2), low (+1.1) |
| Harvest year *** | 2017 (−5.9), 2016 (+4.7) |
| Amount of Fusarium graminearum DNA, n = 227, median: 5.1 DNA copies per ng total DNA | |
| Sowing season * | spring (−1.4), autumn (+0.3) |
| Previous crop species *** | other (−1.4), pasture (−0.9), small-grain cereals (−0.7), canola (+0.3), maize (+2.5) |
| Fertilizer type * | manure (−1.7), mineral + manure (+0.4), mineral (+0.5) |
| Harvest year *** | 2017 (−3.6), 2016 (+3.5) |
| Enniatins content, n = 253, median: 157 µg kg−1 | |
| Production system * | extenso (−1.6), ÖLN (+0.9), organic (+1.1) |
| Sowing season ** | autumn (−0.6), spring (+2.0) |
| Barley variety * | Semper (−1.9), Cassia (−0.6), Caravan (−0.5), Hobbit (−0.3), other (−0.2), Wootan (−0.2), Meridian (+0.5), Tonic (+0.8), Etincel (+1.0), Eunova (+1.9) |
| Previous crop species * | other (−0.7), small-grain cereals (−0.5), canola (−0.3), maize (+0.9), pasture (+2.0) |
| Harvest year *** | 2017 (−4.6), 2016 (+4.7) |
| Aurofusarin content, n = 200, median: 200 µg kg−1 | |
| Sowing season *** | autumn (−0.8), spring (+2.7) |
| Barley variety * | Semper (−1.6), Hobbit (−0.9), Tonic (−0.4), Etincel (−0.4), Cassia (−0.1), Caravan (+0.3), Wootan (+0.3), Meridian (+0.4), other (+1.0), Eunova (+2.3) |
| Previous crop species * | other (−0.7), canola (−0.4), small-grain cereals (−0.1), maize (0), pasture (+2.0) |
| Harvest year *** | 2017 (−3.8), 2016 (+3.2) |
| Deoxynivalenol content, n = 197, median: 105 µg kg−1 | |
| Previous crop species ** | canola (−1.0), pasture (−0.7), other (−0.6), small-grain cereals (−0.5), maize (+2.3) |
| Tillage ** | conventional tillage (−1.5), reduced tillage (+1.4) |
| Fungicide type * | triazole (−0.6), triazole + strobilurin (0), strobilurin (+1.5) |
| Harvest year *** | 2017 (−3.0), 2016 (+2.8) |
| Equisetin content, n = 192, median: 14 µg kg−1 | |
| Growth regulator * | yes (−1.2), no (+1.3) |
| Harvest year *** | 2017 (−1.9), 2016 (+1.4) |
| Moniliformin content, n = 191, median: 9.6 µg kg−1 | |
| Sowing season ** | autumn (−0.6), spring (+1.9) |
| Growth regulator * | no (−1.0), yes (+0.9) |
| Harvest year *** | 2017 (−2.3), 2016 (+1.9) |
| Culmorin content, n = 158, median: 141 µg kg−1 | |
| Production system * | organic (−1.6), extenso (−0.9), ÖLN (+1.1) |
| Sowing season * | spring (−1.7), autumn (+0.3) |
| Growth regulator * | no (−1.3), yes (+1.0) |
| Fertilizer type ** | manure (−1.9), mineral (−0.3), mineral + manure (+1.2) |
| Harvest year *** | 2017 (−2.9), 2016 (+2.2) |
| Butenolide content, n = 124, median: 65 µg kg−1 | |
| Production system ** | extenso (−1.7), ÖLN (+0.9), organic (+1.1) |
| Sowing season *** | autumn (−1.0), spring (+2.3) |
| Barley variety *** | Wootan (−1.7), Semper (−1.6), Caravan (−1.4), Etincel (−0.9), Hobbit (−0.4), other (0), Cassia (+0.2), Meridian (+0.3), Tonic (+1.8), Eunova (+2.3) |
| Growth regulator * | no (−1.2), yes (+1.2) |
| Antibiotic Y content, n = 98, median: 122 µg kg−1 | |
| Sowing season ** | autumn (−0.8), spring (+1.8) |
| Grain yield * | high (−1.2), low (+1.0) |
| Zearalenone content, n = 95, median: 9.9 µg kg−1 | |
| Plant height at harvest * | short (−1.0), tall (+1.0) |
| Harvest year *** | 2017 (−2.2), 2016 (+1.0) |
| Beauvericin content, n = 85, median: 0.6 µg kg−1 | |
| Production system *** | ÖLN (−2.1), extenso (+1.0), organic (+2.3) |
| Sowing season *** | autumn (−1.6), spring (+3.1) |
| Barley variety *** | Tonic (−1.7), Meridian (−1.4), Wootan (−0.7), Hobbit (−0.2), Cassia (0), Etincel (0), Semper (+0.4), other (+1.2), Eunova (+2.4) |
| Previous crop species * | canola (−1.7), other (−0.2), maize (0), small-grain cereals (+0.1), pasture (+1.5) |
| Pre-previous crop species *** | other (−1.5), maize (−1.3), canola (−0.2), small-grain cereals (+0.3), pasture (+2.6) |
| Plant height at harvest *** | tall (−2.0), short (+1.3) |
| Grain yield ** | high (−1.4), low (+1.3) |
| Fungicide type * | triazole (−1.2), triazole + strobilurin (+0.1), strobilurin (+1.9) |
| Growth regulator *** | yes (−2.2), no (+2.1) |
| Fertilizer type *** | mineral + manure (−1.3), mineral (−1.1), manure (+3.0) |
| Nivalenol content, n = 83, median: 23 µg kg−1 | |
| Production system *** | ÖLN (−2.0), extenso (+0.8), organic (+2.1) |
| Sowing season *** | autumn (−1.5), spring (+3.0) |
| Barley variety ** | Meridian (−1.3), Caravan (−1.0), Tonic (−0.9), Semper (−0.8), Cassia (−0.4), Hobbit (0), Etincel (+0.4), Wootan (+0.4), other (+1.4), Eunova (+2.4) |
| Pre-previous crop species * | maize (−1.6), small-grain cereals (0), canola (+0.3), other (+0.3), pasture (+1.6) |
| Plant height at harvest *** | tall (−2.2), short (+1.5) |
| Growth regulator ** | yes (−1.7), no (+1.4) |
| Fertilizer type *** | mineral + manure (−1.7), mineral (−0.5), manure (+2.5) |
| Apicidin content, n = 73, median: 6 µg kg−1 | |
| Sowing season *** | autumn (−1.0), spring (+2.1) |
| Barley variety ** | Hobbit (−2.1), Etincel (−1.0), Semper (−0.7), Meridian (−0.5), Cassia (−0.2), Wootan (0), Caravan (+0.7), other (+0.7), Tonic (+1.4), Eunova (+1.5) |
| Previous crop species * | canola (−1.2), other (−1.1), maize (−0.9), small-grain cereals (+0.7), pasture (+1.6) |
| Fertilizer type * | mineral + manure (−1.0), mineral (−0.3), manure (+1.6) |
| Response Variable | Enniatins 2 (n = 253) | Aurofusarin (n = 200) | Deoxynivalenol (n = 197) | Equisetin (n = 192) | Moniliformin (n = 191) | Culmorin (n = 158) | Butenolide (n = 124) | Antibiotic Y (n = 98) | Zearalenone (n = 95) | Beau-vericin (n = 85) | Nivalenol (n = 83) | Apicidin (n = 73) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Explanatory variable1 | ||||||||||||
| Production system | * | * | ** | *** | *** | |||||||
| Sowing season | ** | *** | ** | * | *** | ** | *** | *** | *** | |||
| Barley variety | * | * | *** | *** | ** | ** | ||||||
| Previous crop | * | * | ** | * | * | |||||||
| Pre-previous crop | *** | * | ||||||||||
| Tillage | ** | |||||||||||
| Plant height at harvest | * | *** | *** | |||||||||
| Grain yield | * | ** | ||||||||||
| Fungicide type | * | * | ||||||||||
| Growth regulator application | * | * | * | * | *** | ** | ||||||
| Fertilizer type | ** | *** | *** | * | ||||||||
| Harvest year | *** | *** | *** | *** | *** | *** | *** |
| Response Variable | Amount of FG DNA(n = 207) | Enniatins 2 (n = 237) | Beauvericin (n = 187) | Deoxynivalenol (n = 163) | Equisetin (n = 161) | Moniliformin (n = 154) | Aurofusarin (n = 146) | Culmorin (n = 141) | Apicidin (n = 131) | Antibiotic Y (n = 77) | Nivalenol (n = 68) | Zearalenone (n = 53) | Butenolide (n = 23) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Explanatory variable1 | |||||||||||||
| Production system | * | *** | *** | * | |||||||||
| Sowing season | ** | ** | *** | * | *** | *** | |||||||
| Barley variety | |||||||||||||
| Previous crop | *** | * | |||||||||||
| Pre-previous crop | * | ** | * | ||||||||||
| Tillage | * | ||||||||||||
| Plant height at harvest | * | * | |||||||||||
| Fungicide type | |||||||||||||
| Growth regulator application | *** | *** | |||||||||||
| Fertilizer type | ** | *** | ** | *** | *** | ||||||||
| Harvest year | *** | *** | *** | * | * | *** | ** |
| Amount of Fusarium graminearum DNA, n = 207, median: 26 DNA copies per ng total DNA | |
| Sowing season ** | spring (−1.8), autumn (+0.4) |
| Previous crop species *** | other (−1.7), pasture (−1.1), small-grain cereals (−0.5), canola (+0.6), maize (+2.1) |
| Plant height at harvest * | short (−1.0), tall (+1.1) |
| Fertilizer type ** | manure (−1.6), mineral (−0.5), mineral + manure (+1.5) |
| Harvest year *** | 2017 (−5.1), 2016 (+4.7) |
| Enniatins content, n = 237, median: 148 µg kg−1 | |
| Sowing season ** | autumn (−0.6), spring (+2.1) |
| Harvest year *** | 2017 (−2.5), 2016 (+2.6) |
| Beauvericin content, n = 187, median: 2.3 µg kg−1 | |
| Production system * | extenso (−0.7), ÖLN (−0.2), organic (+1.8) |
| Sowing season *** | autumn (−0.9), spring (+2.9) |
| Pre-previous crop species * | canola (−1.1), other (−0.7), small-grain cereals (−0.6), maize (+0.1), pasture (+2.0) |
| Fertilizer type *** | mineral (−1.3), mineral + manure (−0.2), manure (+2.8) |
| Harvest year *** | 2016 (−4.0), 2017 (+3.7) |
| Deoxynivalenol content, n = 163, median: 188 µg kg−1 | |
| Production system *** | extenso (−1.7), organic (−1.6), ÖLN (+1.8) |
| Sowing season * | spring (−1.5), autumn (+0.3) |
| Tillage * | conventional tillage (−1.0), reduced tillage (+1.0) |
| Plant height at harvest * | short (−1.1), tall (+1.1) |
| Growth regulator *** | no (−2.0), yes (+1.8) |
| Fertilizer type ** | manure (−2.1), mineral (+0.1), mineral + manure (+1.0) |
| Equisetin content, n = 161, median: 29 µg kg−1 | |
| Production system *** | ÖLN (−2.5), extenso (+1.1), organic (+2.7) |
| Sowing season *** | autumn(−0.7), spring (+2.1) |
| Pre-previous crop species ** | other (−1.1), canola (−0.7), maize (−0.6), small-grain cereals (0), pasture (+2.3) |
| Growth regulator *** | yes (−2.7), no (+2.2) |
| Fertilizer type *** | mineral (−1.5), mineral + manure (−0.3), manure (+3.0) |
| Moniliformin content, n = 154, median: 10 µg kg−1 | |
| Sowing season *** | autumn (−1.1), spring (+3.3) |
| Previous crop species * | canola (−0.9), small−grain cereals (−0.4), other (−0.2), maize (+0.1), pasture (+2.5) |
| Aurofusarin content, n = 146, median: 232 µg kg−1 | |
| Harvest year * | 2017 (−1.3), 2016 (+1.1) |
| Culmorin content, n = 141, median: 575 µg kg−1 | |
| Production system * | organic (−1.9), extenso (0), ÖLN (+0.5) |
| Pre-previous crop species * | other (−1.6), small-grain cereals (−1.1), maize (+0.3), pasture (+0.3), canola (+1.6) |
| Fertilizer type *** | manure (−2.2), mineral + manure (−0.5), mineral (+1.3) |
| Harvest year * | 2017 (−1.4), 2016 (+1.1) |
| Apicidin content, n = 131, median: 24 µg kg−1 | |
| Harvest year *** | 2016 (−1.7), 2017 (+1.6) |
| Antibiotic Y content, n = 77, median: 199 µg kg−1 | |
| No significant associations with explanatory variables | |
| Nivalenol content, n = 68, median: 109 µg kg−1 | |
| Harvest year** | 2017 (−1.1), 2016 (+1.8) |
| Zearalenone content, n = 53, median: 20 µg kg−1 | |
| No significant associations with explanatory variables | |
| Butenolide content, n = 23, median: 264 µg kg−1 | |
| No significant associations with explanatory variables | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Drakopoulos, D.; Sulyok, M.; Jenny, E.; Kägi, A.; Bänziger, I.; Logrieco, A.F.; Krska, R.; Vogelgsang, S. Fusarium Head Blight and Associated Mycotoxins in Grains and Straw of Barley: Influence of Agricultural Practices. Agronomy 2021, 11, 801. https://doi.org/10.3390/agronomy11040801
Drakopoulos D, Sulyok M, Jenny E, Kägi A, Bänziger I, Logrieco AF, Krska R, Vogelgsang S. Fusarium Head Blight and Associated Mycotoxins in Grains and Straw of Barley: Influence of Agricultural Practices. Agronomy. 2021; 11(4):801. https://doi.org/10.3390/agronomy11040801
Chicago/Turabian StyleDrakopoulos, Dimitrios, Michael Sulyok, Eveline Jenny, Andreas Kägi, Irene Bänziger, Antonio F. Logrieco, Rudolf Krska, and Susanne Vogelgsang. 2021. "Fusarium Head Blight and Associated Mycotoxins in Grains and Straw of Barley: Influence of Agricultural Practices" Agronomy 11, no. 4: 801. https://doi.org/10.3390/agronomy11040801