
Allometric Individual Leaf Area Estimation in Chrysanthemum


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
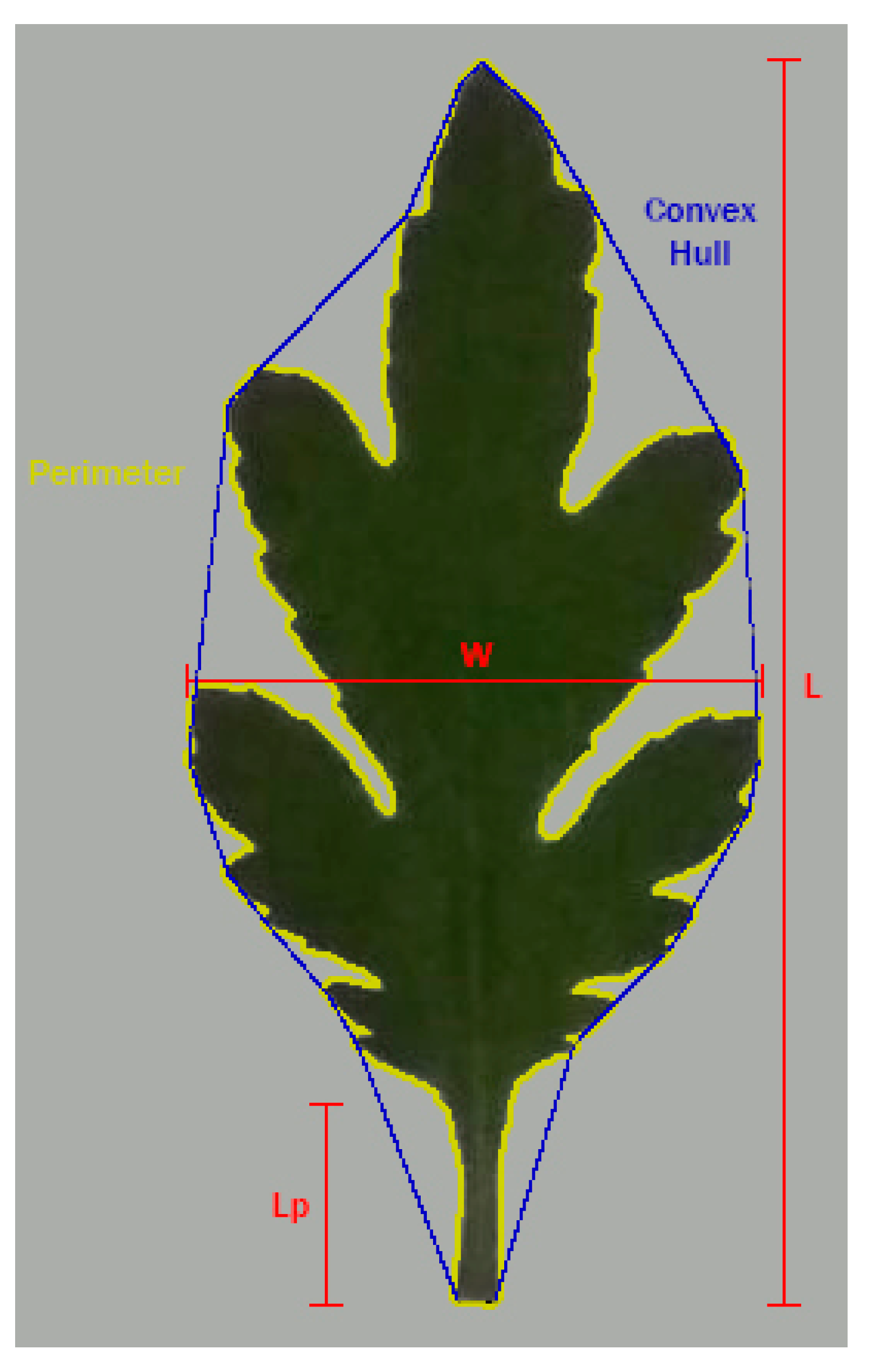
2.2. Morphometric Analysis
2.3. Statistical Analysis
3. Results
3.1. Genetic Variation in Leaf Morphological Traits
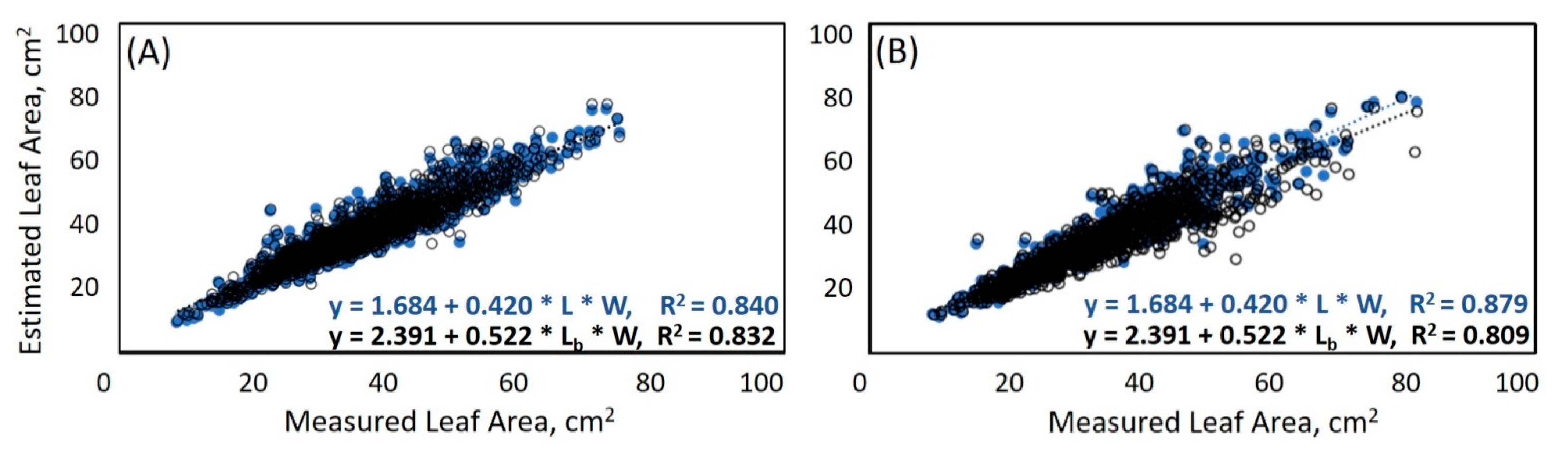
3.2. Calculation of LA Based on Leaf Morphological Traits
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| L | leaf length |
| LA | leaf area |
| Lb | blade length |
| Lp | petiole length |
| T | tolerance |
| VIF | variance inflation factor |
| W | leaf width |
References
- Fanourakis, D.; Hyldgaard, B.; Giday, H.; Bouranis, D.; Körner, O.; Nielsen, K.L.; Ottosen, C.O. Differential effects of elevated air humidity on stomatal closing ability of Kalanchoë blossfeldiana between the C3 and CAM states. Environ. Exp. Bot. 2017, 143, 115–124. [Google Scholar] [CrossRef]
- Fauset, S.; Freitas, H.C.; Galbraith, D.R.; Sullivan, M.J.P.; Aidar, P.M.; Joly, C.A.; Gloor, M.U. Differences in leaf thermoregulation and water use strategies between three co-occurring Atlantic forest tree species. Plant Cell Environ. 2018, 41, 1618–1631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, J.; Zhang, Y.; Zhang, Y.; Bian, Z.; Fanourakis, D.; Yang, Q.; Li, T. Morphological and physiological properties of indoor cultivated lettuce in response to additional far-red light. Sci. Hortic. 2019, 257, 108725. [Google Scholar] [CrossRef]
- Keramatlou, I.; Sharifani, M.; Sabouri, H.; Alizadeh, M.; Kamkar, B. A simple linear model for leaf area estimation in persian walnut (Juglans Regia L.). Sci. Hortic. 2015, 184, 36–39. [Google Scholar] [CrossRef]
- Li, S.; van der Werf, W.; Zhu, J.; Guo, Y.; Li, B.; Ma, Y.; Evers, J.B. Estimating the contribution of plant traits to light partitioning in simultaneous maize/soybean intercropping. J. Exp. Bot. 2021, erab077. [Google Scholar] [CrossRef]
- Honnaiah, P.A.; Sridhara, S.; Gopakkali, P.; Ramesh, N.; Mahmoud, E.A.; Alkallas, F.H.; Abdelmohsen, S.A.M.; El-Ansary, D.O.; Elansary, H.O. Influence of sowing windows and genotypes on growth, radiation interception, conversion efficiency and yield of guar. Saudi J. Biol. Sci. 2021. [Google Scholar] [CrossRef]
- Jo, W.J.; Shin, J.H. Effect of leaf-area management on tomato plant growth in greenhouses. Hortic. Environ. Biotechnol. 2020, 61, 981–988. [Google Scholar] [CrossRef]
- Fiorani, F.; Schurr, U. Future scenarios for plant phenotyping. Annu. Rev. Plant Biol. 2012, 64, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Fanourakis, D.; Giday, H.; Hyldgaard, B.; Bouranis, D.; Körner, O.; Ottosen, C.O. Low air humidity during growth promotes stomatal closure ability in roses. Eur. J. Hortic. Sci. 2019, 84, 245–252. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Tian, L.; Liang, S.; Becker-Reshef, I.; Su, W.; Zhang, X.; Zhu, D.; Wu, W. Improving winter wheat yield estimation by assimilation of the leaf area index from Landsat TM and MODIS data into the WOFOST model. Agric. For. Meteorol. 2015, 204, 106–121. [Google Scholar] [CrossRef] [Green Version]
- Fanourakis, D.; Bouranis, D.; Tsaniklidis, G.; Rezaei Nejad, A.; Ottosen, C.O.; Woltering, E.J. Genotypic and phenotypic differences in fresh weight partitioning of cut rose stems: Implications for water loss. Acta Physiol. Plant. 2020, 42, 48. [Google Scholar] [CrossRef]
- Fanourakis, D.; Briese, C.; Max, J.F.J.; Kleinen, S.; Putz, A.; Fiorani, F.; Ulbrich, A.; Schurr, U. Rapid determination of leaf area and plant height by using light curtain arrays in four species with contrasting shoot architecture. Plant Methods 2014, 10, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das Choudhury, S.; Bashyam, S.; Qiu, Y.; Samal, A.; Awada, T. Holistic and component plant phenotyping using temporal image sequence. Plant Methods 2018, 14, 35. [Google Scholar] [CrossRef] [PubMed]
- Antunes, W.C.; Pompelli, M.F.; Carretero, D.M.; Da Matta, F.M. Allometric models for non-destructive leaf area estimation in coffee (Coffea arabica and Coffea canephora). Ann. Appl. Biol. 2008, 153, 33–40. [Google Scholar] [CrossRef] [Green Version]
- Pompelli, M.F.; Antunes, W.C.; Ferreira, D.T.R.G.; Cavalcante, P.G.S.; Wanderley-Filho, H.C.L.; Endres, L. Allometric models for non-destructive leaf area estimation of Jatropha curcas. Biomass Bioenergy 2012, 36, 77–85. [Google Scholar] [CrossRef]
- Verwijst, T.; Wen, D.Z. Leaf allometry of Salix viminalis during the first growing season. Tree Physiol. 1996, 16, 655–660. [Google Scholar] [CrossRef]
- Cornelissen, J.H.C.; Lavorel, S.; Garnier, E.; Díaz, S.; Buchmann, N.; Gurvich, D.E.; Reich, P.B.; Ter Steege, H.; Morgan, H.D.; Van Der Heijden, M.G.A.; et al. A handbook of protocols for standardised and easy measurement of plant functional traits worldwide. Austr. J. Bot. 2003, 51, 335–380. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Zhang, L. Leaf shape alters the coefficients of leaf area estimation models for Saussurea stoliczkai in central Tibet. Photosynthetica 2012, 50, 337–342. [Google Scholar] [CrossRef]
- Yu, X.J.; Shi, P.J.; Schrader, J.; Niklas, K.J. Nondestructive estimation of leaf area for 15 species of vines with different leaf shapes. Am. J. Bot. 2020, 107, 1481–1490. [Google Scholar] [CrossRef]
- Fallovo, C.; Cristofori, V.; Mendoza-de Gyves, E.; Rivera, C.M.; Fanasca, S.; Bignami, C.; Sassine, Y.; Rouphael, Y. Leaf area estimation model for small fruits from linear measurements. HortScience 2008, 43, 2263–2267. [Google Scholar] [CrossRef] [Green Version]
- Sala, F.; Arsene, G.G.; Iordănescu, O.; Boldea, M. Leaf area constant model in optimizing foliar area measurement in plants: A case study in apple tree. Sci. Hortic. 2015, 193, 218–224. [Google Scholar] [CrossRef]
- Perez-Perez, J.M.; Serrano-Cartagena, J.; Micol, J.L. Genetic analysis of natural variations in the architecture of Arabidopsis thaliana vegetative leaves. Genetics 2002, 162, 893–915. [Google Scholar]
- Niinemets, Ü.; Al Afas, N.; Cescatti, A.; Pellis, A.; Ceulemans, R. Petiole length and biomass investment in support modify light interception efficiency in dense poplar plantations. Tree Physiol. 2004, 24, 141–154. [Google Scholar] [CrossRef] [PubMed]
- Gao, K.; Song, X.; Kong, D.; Dai, S. Genetic analysis of leaf traits in small-flower chrysanthemum (Chrysanthemum × morifolium Ramat.). Agronomy 2020, 10, 697. [Google Scholar] [CrossRef]
- Seif, M.; Aliniaeifard, S.; Arab, M.; Mehrjerdi, M.Z.; Shomali, A.; Fanourakis, D.; Li, T.; Woltering, E. Monochromatic red light during plant growth decreases the size and improves the functionality of stomata in chrysanthemum. Funct. Plant Biol. 2021, 48, 515–528. [Google Scholar] [CrossRef]
- Chitwood, D.H.; Kumar, R.; Ranjan, A.; Pelletier, J.M.; Townsley, B.; Ichihashi, Y.; Martinez, C.C.; Zumstein, K.; Harada, J.J.; Maloof, J.N.; et al. Light-induced indeterminacy alters shade avoiding tomato leaf morphology. Plant Physiol. 2015, 169, 2030–2047. [Google Scholar] [CrossRef] [Green Version]
- Gupta, S.; Rosenthal, D.M.; Stinchcombe, J.R.; Baucom, R.S. The remarkable morphological diversity of leaf shape in sweetpotato (Ipomoea batatas): The influence of genetics, environment, and G9E. New Phytol. 2020, 225, 2183–2195. [Google Scholar] [CrossRef]
- Marquaridt, D.W. Generalized inverses, ridge regression, biased linear estimation, and nonlinear estimation. Technometrics 1970, 12, 591–612. [Google Scholar] [CrossRef]
- Gill, J. Outliers, residuals, and influence in multiple regression. J. Anim. Breed. Genet. 1986, 103, 161–175. [Google Scholar] [CrossRef]
- Hassanvand, F.; Rezaei Nejad, A.; Fanourakis, D. Morphological and physiological components mediating the silicon-induced enhancement of geranium essential oil yield under saline conditions. Ind. Crops Prod. 2019, 134, 19–25. [Google Scholar] [CrossRef]
- Waldner, F.; Chen, Y.; Horan, H.; Hochan, Z. High temporal resolution of leaf area data improves empirical estimation of grain yield. Sci. Rep. 2019, 9, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Sørensen, H.K.; Fanourakis, D.; Tsaniklidis, G.; Bouranis, D.; Rezaei Nejad, A.; Ottosen, C.O. Using artificial lighting based on electricity price without a negative impact on growth, visual quality or stomatal closing response in Passiflora. Sci. Hortic. 2020, 267, 109354. [Google Scholar] [CrossRef]
- Montero, F.J.; de Juan, J.A.; Cuesta, A.; Brasa, A. Nondestructive methods to estimate leaf area in Vitis vinifera L. HortScience 2000, 35, 696–698. [Google Scholar] [CrossRef]
- Koubouris, G.; Bouranis, D.; Vogiatzis, E.; Rezaei Nejad, A.; Giday, H.; Tsaniklidis, G.; Ligoxigakis, E.K.; Blazakis, K.; Kalaitzis, P.; Fanourakis, D. Leaf area estimation by considering leaf dimensions in olive tree. Sci. Hortic. 2018, 240, 440–445. [Google Scholar] [CrossRef]
- Schwarz, D.; Kläring, H.P. Allometry to estimate leaf area of tomato. J. Plant Nutr. 2001, 24, 1291–1309. [Google Scholar] [CrossRef]
- Chitwood, D.H.; Ranjan, A.; Martinez, C.C.; Headland, L.R.; Thiem, T.; Kumar, R.; Covington, M.F.; Hatcher, T.; Naylor, D.T.; Zimmerman, S.; et al. A modern ampelography: A genetic basis for leaf shape and venation patterning in grape. Plant Physiol. 2014, 164, 259–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fanourakis, D.; Giday, H.; Li, T.; Kambourakis, E.; Ligoxigakis, E.K.; Papadimitriou, M.; Strataridaki, A.; Bouranis, D.; Fiorani, F.; Heuvelink, E.; et al. Antitranspirant compounds alleviate the mild-desiccation-induced reduction of vase life in cut roses. Postharvest Biol. Technol. 2016, 117, 110–117. [Google Scholar] [CrossRef]
- Backhaus, A.; Kuwabara, A.; Bauch, M.; Monk, N.; Sanguinetti, G.; Fleming, A. LEAFPROCESSOR: A new leaf phenotyping tool using contour bending energy and shape cluster analysis. New Phytol. 2010, 187, 251–261. [Google Scholar] [CrossRef] [PubMed]
- Wooten, J.W. Variations in leaf characteristics of six species of Sagittaria (Alismataceae) caused by various water levels. Aquatic Bot. 1986, 23, 321–327. [Google Scholar] [CrossRef]
- Tsukaya, H.; Kozuka, T.; Kim, G.T. Genetic control of petiole length in Arabidopsis thaliana. Plant Cell Physiol. 2002, 43, 1221–1228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poorter, H.; Fiorani, F.; Pieruschka, R.; Wojciechowski, T.; van der Putten, W.; Kleyer, M.; Schurr, U.; Postma, J. Pampered inside, pestered outside? Differences and similarities between plants growing in controlled conditions and in the field. New Phytol. 2016, 212, 838–855. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Cultivar | Length (cm) | Width (cm) | Petiole Length (cm) | Petiole Length/Leaf Length (%) | Leaf Area (cm2) | Perimeter (cm) | Circularity | Aspect Ratio | Roundness | Solidity |
|---|---|---|---|---|---|---|---|---|---|---|
| Alamos yellow | 13.01 b (±0.09) | 7.31 a (±0.08) | 2.37 c (±0.03) | 18.2 e (±0.2) | 41.7 b (±0.6) | 62.5 b (±0.6) | 0.136 c,d (±0.002) | 1.69 c (±0.01) | 0.596 b (±0.004) | 0.665 b (±0.004) |
| Baltica | 12.75 b,c (±0.08) | 5.98 c (±0.05) | 2.60 b (±0.03) | 20.4 d (±0.2) | 34.4 c (±0.4) | 56.6 c (±0.5) | 0.137 c,d (±0. 002) | 1.93 b (±0.01) | 0.522 c (±0.003) | 0.671 b (±0.002) |
| Baltica pink | 12.31 d (±0.08) | 5.62 d (±0.05) | 2.45 c (±0.02) | 20.0 d (±0.2) | 29.3 d (±0.4) | 49.2 e (±0.4) | 0.153 b (±0.001) | 2.04 a (±0.01) | 0.497 d (±0.004) | 0.628 c (±0.003) |
| Baltica salmon | 12.54 c,d (±0.09) | 5.74 c,d (±0.05) | 2.70 b (±0.03) | 21.6 c (±0.2) | 31.1 d (±0.5) | 52.9 d (±0.6) | 0.141 c (±0.002) | 2.09 a (±0.02) | 0.485 d (±0.004) | 0.635 c (±0.003) |
| Botempi red | 11.73 e (±0.10) | 6.31 b (±0.05) | 2.70 b (±0.04) | 22.9 b (±0.2) | 35.1 c (±0.5) | 50.6 d,e (±0.5) | 0.173 a (±0.002) | 1.51 d (±0.01) | 0.667 a (±0.003) | 0.706 a (±0.002) |
| Amethyst yellow | 14.09 a (±0.13) | 7.22 a (±0.07) | 3.35 a (±0.04) | 23.8 a (±0.3) | 46.4 a (±0.9) | 66.8 a (±0.8) | 0.133 d (±0.002) | 1.71 c (±0.03) | 0.609 b (±0.006) | 0.665 b (±0.006) |
| p | 0.001 | 0.001 | 0.001 | 0.001 | 0.001 | 0.001 | 0.001 | 0.001 | 0.001 | 0.001 |
| Cultivar | Length (cm) | Width (cm) | Petiole Length (cm) | Petiole Length/Leaf Length (%) | Leaf Area (cm2) | Perimeter (cm) | Circularity | Aspect Ratio | Roundness | Solidity |
|---|---|---|---|---|---|---|---|---|---|---|
| VIP | 12.83 b,c (±0.15) | 6.42 a,b (±0.11) | 3.50 a (±0.06) | 27.3 a (±0.4) | 37.5 a,b,c (±1.1) | 49.7 b (±0.8) | 0.190 b,c (±0.003) | 1.77 b,c (±0.02) | 0.571 c (±0.005) | 0.692 b,c (±0.004) |
| Chili pepper | 12.46 c,d (±0.17) | 6.54 a,b (±0.11) | 2.58 c (±0.06) | 20.7 c,d (±0.4) | 38.6 a,b,c (±1.1) | 51.7 b (±1.0) | 0.187 c (±0.004) | 1.64 d (±0.01) | 0.614 b (±0.005) | 0.701 b,c (±0.003) |
| Britain pink | 11.35 e,f (±0.14) | 5.97 c,d (±0.09) | 2.63 c (±0.04) | 23.4 b (±0.4) | 30.3 d,e (±0.9) | 42.2 c (±0.8) | 0.216 a (±0.004) | 1.69 c,d (±0.02) | 0.603 b (±0.007) | 0.689 b,c (±0.004) |
| Euro white | 14.09 a (±0.21) | 6.48 a,b (±0.12) | 3.33 a (±0.08) | 23.5 b (±0.4) | 39.0 a,b (±1.2) | 56.0 a (±1.0) | 0.156 d (±0.002) | 1.95 a (±0.02) | 0.517 d (±0.005) | 0.663 d (±0.003) |
| Euro yellow | 13.36 b (±0.16) | 6.47 a,b (±0.10) | 2.96 b (±0.05) | 22.2 b,c (±0.3) | 35.7 b,c (±0.8) | 58.7 a (±1.0) | 0.135 e (±0.003) | 1.94 a (±0.02) | 0.523 d (±0.006) | 0.634 e (±0.003) |
| Podolsk purple | 12.83 b,c (±0.14) | 6.30 a,b,c (±0.11) | 2.68 c (±0.05) | 21.0 c,d (±0.4) | 35.7 b,c (±1.0) | 59.5 a (±1.1) | 0.131 e (±0.003) | 1.82 b (±0.03) | 0.561 c (±0.007) | 0.663 d (±0.005) |
| Podolsk red | 12.83 b,c (±0.15) | 6.10 b,c (±0.09) | 2.53 c (±0.04) | 19.8 d (±0.3) | 34.5 c,d (±0.8) | 58.8 a (±1.1) | 0.129 e (±0.003) | 1.78 b (±0.02) | 0.570 c (±0.006) | 0.685 c (±0.004) |
| Veronica | 10.89 f (±0.16) | 5.57 d (±0.09) | 2.27 d (±0.03) | 21.0 c,d (±0.3) | 28.6 e (±0.8) | 42.9 c (±0.8) | 0.197 b,c (±0.003) | 1.81 b (±0.01) | 0.557 c (±0.004) | 0.702 b (±0.002) |
| Zenhya white | 12.01 d,e (±0.14) | 6.70 a (±0.09) | 3.29 a (±0.04) | 27.5 a (±0.3) | 40.5 a (±1.1) | 50.4 b (±0.8) | 0.204 a,b (±0.004) | 1.46 e (±0.01) | 0.692 a (±0.005) | 0.720 a (±0.004) |
| p | 0.001 | 0.001 | 0.001 | 0.001 | 0.001 | 0.001 | 0.001 | 0.001 | 0.001 | 0.001 |
| Model | Fitted Coefficient and Constant | R2, z | MSE z | PRESS z | SSE z | ||
|---|---|---|---|---|---|---|---|
| a | b | ||||||
| 1 | LA = a + b · L | −33.107 | 5.449 | 0.706 | 35.29 | 5.94 | 52858 |
| 2 | LA = a + b · W | 15.202 | 8.099 | 0.732 | 32.24 | 5.68 | 48300 |
| 3 | LA = a + b · L2 | 0.142 | 0.219 | 0.724 | 33.12 | 5.76 | 49614 |
| 4 | LA = a + b · W2 | 11.011 | 0.605 | 0.718 | 33.87 | 5.82 | 50741 |
| 5 | LA = a + b · L · W | 1.684 | 0.420 | 0.840 | 19.25 | 4.39 | 28841 |
| 6 | LA = a + b (L + W) | −35.515 | 3.761 | 0.827 | 20.76 | 4.56 | 31102 |
| 7 | LA = a + b (L + W)2 | −0.443 | 0.099 | 0.839 | 19.29 | 4.39 | 28899 |
| 8 | LA = a (L + W)3 | 11.874 | 0.003 | 0.833 | 20.09 | 4.48 | 30090 |
| Model | Fitted Coefficient and Constant | R2, z | MSE z | PRESS z | SSE z | ||
|---|---|---|---|---|---|---|---|
| a | b | ||||||
| 1 | LA = a + b · L | −33.107 | 5.449 | 0.763 | 30.87 | 5.56 | 30803 |
| 2 | LA = a + b · W | 15.202 | 8.099 | 0.824 | 22.92 | 4.79 | 22870 |
| 3 | LA = a + b · L2 | 0.142 | 0.219 | 0.764 | 30.82 | 5.55 | 30762 |
| 4 | LA = a + b · W2 | 11.011 | 0.605 | 0.816 | 24.06 | 4.91 | 24011 |
| 5 | LA = a + b · L · W | 1.684 | 0.420 | 0.879 | 15.74 | 3.97 | 15706 |
| 6 | LA = a + b (L + W) | −35.515 | 3.761 | 0.860 | 18.21 | 4.27 | 18168 |
| 7 | LA = a + b (L + W)2 | −0.443 | 0.099 | 0.864 | 17.78 | 4.22 | 17745 |
| 8 | LA = a (L + W)3 | 11.874 | 0.003 | 0.845 | 20.19 | 4.49 | 20153 |
| Model | Fitted Coefficient and Constant | R2, z | MSE z | PRESS z | SSE z | ||
|---|---|---|---|---|---|---|---|
| a | b | ||||||
| 1 | LA = a + b · Lb | −25.912 | 6.194 | 0.652 | 41.73 | 6.46 | 62466 |
| 3 | LA = a + b · Lb2 | 3.881 | 0.315 | 0.666 | 40.08 | 6.33 | 59997 |
| 5 | LA = a + b · Lb · W | 2.391 | 0.522 | 0.832 | 20.12 | 4.49 | 30119 |
| 6 | LA = a + b (Lb + W) | −32.272 | 4.180 | 0.817 | 21.91 | 4.68 | 32802 |
| 7 | LA = a + b (Lb + W)2 | 1.326 | 0.127 | 0.826 | 20.83 | 4.56 | 31186 |
| 8 | LA = a (Lb + W)3 | 13.115 | 0.005 | 0.817 | 22.00 | 4.69 | 32939 |
| Model | Fitted Coefficient and Constant | R2, z | MSE z | PRESS z | SSE z | ||
|---|---|---|---|---|---|---|---|
| a | b | ||||||
| 1 | LA = a + b · Lb | −25.912 | 6.194 | 0.601 | 54.21 | 7.36 | 60818 |
| 3 | LA = a + b · Lb2 | 3.881 | 0.315 | 0.596 | 54.92 | 7.41 | 61623 |
| 5 | LA = a + b · Lb · W | 2.391 | 0.522 | 0.809 | 25.99 | 5.10 | 29159 |
| 6 | LA = a + b (Lb + W) | −32.272 | 4.180 | 0.785 | 29.26 | 5.41 | 32831 |
| 7 | LA = a + b (Lb + W)2 | 1.326 | 0.127 | 0.781 | 29.72 | 5.45 | 33351 |
| 8 | LA = a (Lb + W)3 | 13.115 | 0.005 | 0.759 | 32.83 | 5.73 | 36839 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fanourakis, D.; Kazakos, F.; Nektarios, P.A. Allometric Individual Leaf Area Estimation in Chrysanthemum. Agronomy 2021, 11, 795. https://doi.org/10.3390/agronomy11040795
Fanourakis D, Kazakos F, Nektarios PA. Allometric Individual Leaf Area Estimation in Chrysanthemum. Agronomy. 2021; 11(4):795. https://doi.org/10.3390/agronomy11040795
Chicago/Turabian StyleFanourakis, Dimitrios, Filippos Kazakos, and Panayiotis A. Nektarios. 2021. "Allometric Individual Leaf Area Estimation in Chrysanthemum" Agronomy 11, no. 4: 795. https://doi.org/10.3390/agronomy11040795