
Combining QTL Mapping and Gene Expression Analysis to Elucidate the Genetic Control of ‘Crumbly’ Fruit in Red Raspberry (Rubus idaeus L.)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Genomic Analysis: Quantitative Trait Loci (QTL)
2.3. Isolation of RNA
2.4. Microarray Experimental Design and Data Analysis
2.5. Microarray Analysis
2.5.1. Selection of Microarray Probes Mapped Inside the Crumbly QTLs
2.5.2. Analysis of Variance (ANOVA)
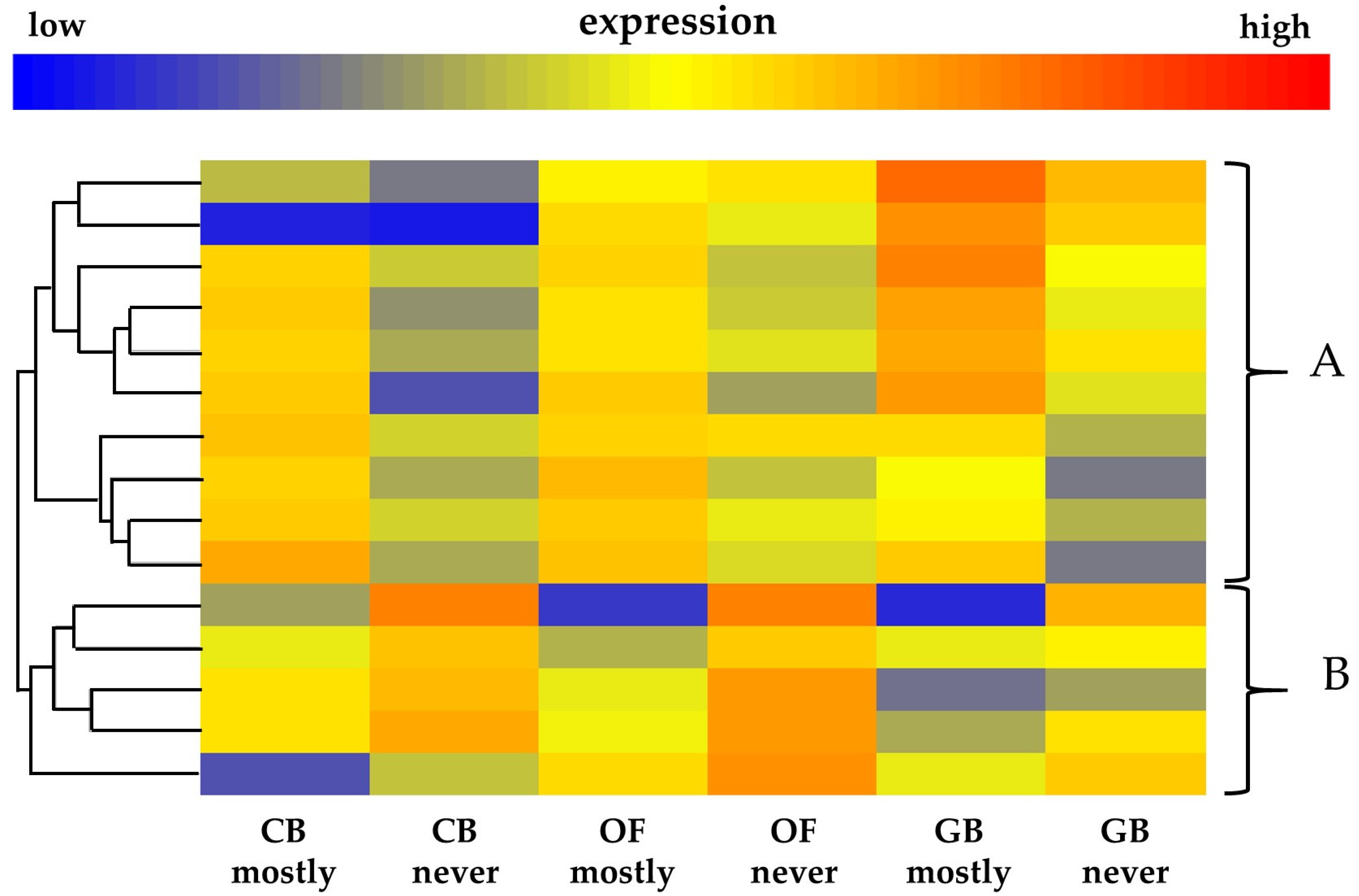
2.5.3. Heatmap Tree Clustering
2.5.4. BLAST Search
2.5.5. Functional Homology
3. Results
3.1. Crumbly Fruit QTL
3.2. Gene Ontology Analysis of Differentially Expressed Genes
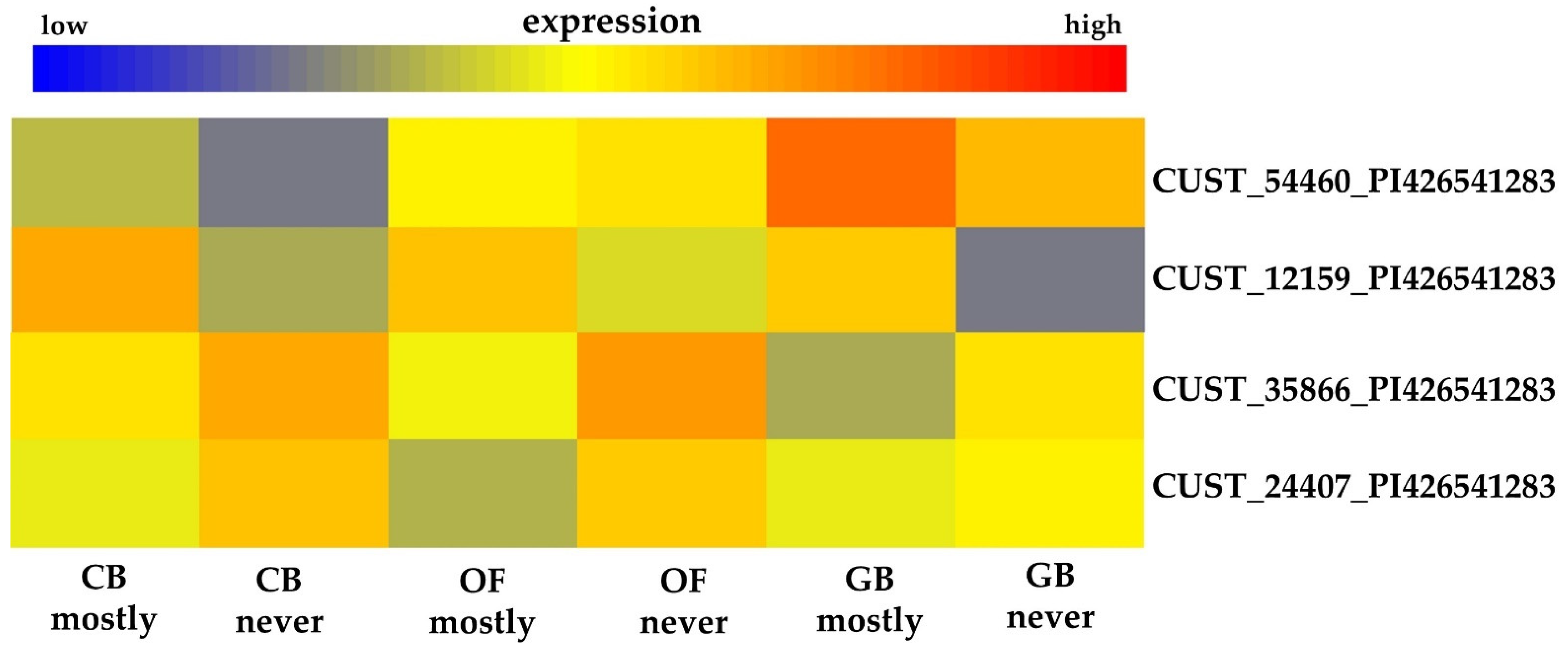
3.3. Selection of A. thaliana Ortholog Genes Matching Those Probes That Are Significantly Differentially Expressed Having GO Terms Other Than Flower Development, Hormones, or Pollen
3.4. Relationship Between Crumbly Fruit and Ripening
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Graham, J.; Smith, K.; McCallum, S.; Hedley, P.E.; Cullen, D.W.; Dolan, A.; Milne, L.; McNicol, J.W.; Hackett, C.A. Towards an understanding of the control of ‘crumbly’ fruit in red raspberry. SpringerPlus 2015, 4, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scolari, L.M. Understanding the Genetic and Physiology Controls of ‘Crumbly’ Fruit in Red Raspberry (Rubus Idaeus). Ph.D Thesis, Heriot-Watt University, Edinburgh, UK, February 2021. [Google Scholar]
- Quito-Avila, D.F.; Lightle, D.; Martin, R.R. Effect of Raspberry bushy dwarf virus, Raspberry leaf mottle virus, and Raspberry latent virus on Plant Growth and Fruit Crumbliness in ‘Meeker’ Red Raspberry. Plant Dis. 2014, 98, 176–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hackett, C.A.; Milne, L.; Smith, K.; Hedley, P.; Morris, J.; Simpson, C.G.; Preedy, K.; Graham, J. Enhancement of Glen Moy x Latham raspberry linkage map using GbS to further understand control of developmental processes leading to fruit ripening. BMC Genet. 2018, 19, 59. [Google Scholar] [CrossRef] [PubMed]
- Hackett, C.A.; McLean, K.; Bryan, G.J. Linkage Analysis and QTL Mapping Using SNP Dosage Data in a Tetraploid Potato Mapping Population. PLoS ONE 2013, 8, e63939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Ooijen, J.W. MapQTL ® 5 Software for the mapping of quantitative trait loci in experimental populations of diploid species. Wageningen, 2004.
- van Ooijen, J.W. Accuracy of mapping quantitative trait loci in autogamous species. Theor. Appl. Genet. 1992, 84, 803–811. [Google Scholar] [CrossRef] [PubMed]
- Kloosterman, B.; Oortwijn, M.; Uitdewilligen, J.; America, T.; De Vos, R.; Visser, R.G.; Bachem, C.W. From QTL to candidate gene: Genetical genomics of simple and complex traits in potato using a pooling strategy. BMC Genom. 2010, 11, 158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- The James Hutton Institute Private Resource. Available online: http://camel.hutton.ac.uk/raspberry/ (accessed on 9 April 2021).
- The Arabidopsis Information Resource (TAIR). Available online: https://www.webcitation.org/query?id=1392515205347100&date=%400&fromform=1 (accessed on 16 February 2014).
- The Arabidopsis Information Resource (TAIR). Gene Ontology at TAIR. Available online: https://www.webcitation.org/5ElORcKdS (accessed on 16 November 2013).
- Jennings, D.L. Raspberries and Blackberries: Their Breeding, Diseases and Growth. Academic Press Limited: London, UK, 1988; p. 230. [Google Scholar]
- Ruan, Y.-L.; Patrick, J.W.; Bouzayen, M.; Osorio, S.; Fernie, A.R. Molecular regulation of seed and fruit set. Trends Plant Sci. 2012, 17, 656–665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, R.; Khurana, A.; Sharma, A.K. Role of plant hormones and their interplay in development and ripening of fleshy fruits. J. Exp. Bot. 2013, 65, 4561–4575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azzi, L.; Deluche, C.; Gévaudant, F.; Frangne, N.; Delmas, F.; Hernould, M.; Chevalier, C. Fruit growth-related genes in tomato. J. Exp. Bot. 2015, 66, 1075–1086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.-H.; Xu, Z.-H.; Xue, H.-W. Arabidopsis Membrane Steroid Binding Protein 1 Is Involved in Inhibition of Cell Elongation. Plant Cell 2005, 17, 116–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Marzo, M.; Herrera-Ubaldo, H.; Caporali, E.; Novák, O.; Strnad, M.; Balanzà, V.; Ezquer, I.; Mendes, M.A.; de Folter, S.; Colombo, L. SEEDSTICK Controls Arabidopsis Fruit Size by Regulating Cytokinin Levels and FRUITFULL. Cell Rep. 2020, 30, 2846–2857.e3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wada, T.; Tachibana, T.; Shimura, Y.; Okada, K. Epidermal Cell Differentiation inArabidopsisDetermined by aMybHomolog, CPC. Science 1997, 277, 1113–1116. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.; Huang, S. ArabidopsisACT11 modifies actin turnover to promote pollen germination and maintain the normal rate of tube growth. Plant J. 2015, 83, 515–527. [Google Scholar] [CrossRef] [PubMed]
- Gutkowska, M.; Wnuk, M.; Nowakowska, J.; Lichocka, M.; Stronkowski, M.M.; Swiezewska, E. Rab geranylgeranyl transferase β subunit is essential for male fertility and tip growth in Arabidopsis. J. Exp. Bot. 2015, 66, 213–224. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Jiang, Y.; Zhang, X.; Wang, X.; Wang, Y.; Han, Y.; Coupland, G.; Jin, J.B.; Searle, I.; Fu, Y.-F.; et al. Two SUMO Proteases SUMO PROTEASE RELATED TO FERTILITY1 and 2 Are Required for Fertility in Arabidopsis. Plant Physiol. 2017, 175, 1703–1719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; Li, Y.; Liu, F.; Tan, X.; Rui, Q.; Tong, Y.; Qiao, L.; Gao, R.; Li, G.; Shi, R.; et al. Overexpressed Tomosyn Binds Syntaxins and Blocks Secretion during Pollen Development. Plant Physiol. 2019, 181, 1114–1126. [Google Scholar] [CrossRef] [PubMed]
- Mo, P.; Zhu, Y.; Liu, X.; Zhang, A.; Yan, C.; Wang, D. Identification of two phosphatidylinositol/phosphatidylcholine transfer protein genes that are predominately transcribed in the flowers of Arabidopsis thaliana. J. Plant Physiol. 2007, 164, 478–486. [Google Scholar] [CrossRef] [PubMed]
- Hála, M.; Soukupová, H.; Synek, L.; Žárský, V. Arabidopsis RAB geranylgeranyl transferase β-subunit mutant is constitutively photomorphogenic, and has shoot growth and gravitropic defects. Plant J. 2010, 62, 615–627. [Google Scholar] [CrossRef] [PubMed]
- Edlund, A.F.; Swanson, R.; Preuss, D. Pollen and Stigma Structure and Function: The Role of Diversity in Pollination. Plant Cell 2004, 16, S84–S97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scofield, S.; Dewitte, W.; Murray, J.A.H. The KNOX gene SHOOT MERISTEMLESS is required for the development of reproductive meristematic tissues in Arabidopsis. Plant J. 2007, 50, 767–781. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Year | LG | POS. (cM) | Mean | %Var | LOD | S.E. | Detected Previously | Key Parent | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ac | ad | bc | bd | ac | ad | bc | bd | |||||||
| 2007 a | 1 | 6 | 0.0325 | 0.1717 | 0.44 | 0.382 | 10.09 | 5.05 | 0.0735 | 0.0547 | 0.0693 | 0.0549 | yes | Latham |
| 2009 a | 1 | 7 | 0.0589 | 0.1541 | 0.4517 | 0.5365 | 16.62 | 8.12 | 0.0728 | 0.0556 | 0.0687 | 0.0562 | yes | Latham |
| 2010 a | 1 | 0 | 0.3444 | 0.1595 | 0.4439 | 0.5468 | 9.07 | 4.7 | 0.0768 | 0.06 | 0.0784 | 0.064 | yes | Latham |
| 2011 a | 1 | 9 | 0.7709 | 0.7415 | 1.511 | 1.5776 | 14.56 | 6.45 | 0.1678 | 0.1279 | 0.1673 | 0.1271 | yes | Latham |
| 2012 a | 1 | 10 | 0.4466 | 0.4391 | 0.8235 | 0.8139 | 13.46 | 5.16 | 0.0894 | 0.0696 | 0.084 | 0.0643 | yes | Latham |
| 2012 b | 1 | 9 | 0.6913 | 0.6321 | 1.5116 | 1.6296 | 15.91 | 6.05 | 0.2079 | 0.1584 | 0.1898 | 0.148 | yes | Latham |
| 2007 a | 3 | 59 | 0.206 | 0.2014 | 0.5333 | 0.1075 | 11.8 | 6.05 | 0.0525 | 0.0704 | 0.0602 | 0.0638 | no | c both |
| 2009 a | 3 | 59 | 0.2639 | 0.2443 | 0.5952 | 0.129 | 12.32 | 6.31 | 0.055 | 0.0737 | 0.0631 | 0.0668 | no | c both |
| 2010 a | 3 | 57 | 0.2591 | 0.3517 | 0.5919 | 0.1572 | 10.33 | 5.25 | 0.0602 | 0.0782 | 0.0614 | 0.0803 | no | c both |
| 2011 a | 3 | 107 | 0.7845 | 0.948 | 1.5999 | 1.7028 | 13.43 | 6.04 | 0.1227 | 0.1382 | 0.1502 | 0.1868 | yes | Latham |
| 2012 a | 3 | 54 | 0.5463 | 0.5677 | 0.8801 | 0.4645 | 10.19 | 4.44 | 0.0681 | 0.0877 | 0.0671 | 0.0906 | no | c both |
| 2012 b | 3 | 106 | 0.7324 | 0.9398 | 1.627 | 1.6786 | 11.96 | 4.75 | 0.1512 | 0.1712 | 0.1795 | 0.2165 | yes | Latham |
| Microarray Probe | Significant Stage*Phenotype Interaction | A. thaliana Gene ID | QTL | Scaffold | Gene Ontology (GO) Term |
|---|---|---|---|---|---|
| CUST_54460_PI426541283 | e | AT1G62360.1 | cr_JHI_3-15 | 4 | GO:0009691 (cytokinin biosynthetic process) GO:0048440 (carpel development) GO:0010582 (floral meristem determinacy) |
| CUST_38171_PI426541283 | a | AT3G57540.1 | cr_JHI_3-20 | 419 | GO:0009737 (response to abscisic acid) |
| CUST_38657_PI426541283 | c | AT2G46410.1 | cr_JHI_3-15 | 4 | GO:0009753 (response to jasmonic acid) GO:0009751 (response to salicylic acid) |
| ƚ CUST_27324_PI426541283 | e | AT3G63440.1 | cr_JHI_3-20 | 1500 | GO:0019139 (cytokinin dehydrogenase activity) GO:0009690 (cytokinin metabolic process) |
| ƚ CUST_10154_PI426541283 | e | AT3G63440.1 | cr_JHI_3-20 | 1500 | GO:0019139 (cytokinin dehydrogenase activity) GO:0009690 (cytokinin metabolic process) |
| CUST_22099_PI426541283 | a | AT5G52240.1 | cr_JHI_1-15 | 260 | GO:0005496 (steroid binding) |
| CUST_33454_PI426541283 | e | AT1G05180.1 | cr_JHI_3-15 | 4 | GO:0009734 (auxin activated signalling pathway) GO:0010252 (auxin homeostasis) GO:0009735 (response to cytokinin) |
| CUST_28007_PI426541283 | c | AT3G29770.1 | cr_JHI_3-15 | 509 | GO:0080030 (methyl indole-3-acetate esterase activity) GO:0080031 (methyl salicylate esterase activity) GO:0080032 (methyl jasmonate esterase activity) GO:0009694 (jasmonic acid metabolic process) GO:0009696 (salicylic acid metabolic process) |
| CUST_20489_PI426541283 | e | AT1G11910.1 | cr_JHI_3-15 | 4 | GO:0009735 (response to cytokinin) |
| CUST_12159_PI426541283 | c | AT2G19130.1 | cr_JHI_3-15 | 858 | GO:0048544 (recognition of pollen) |
| CUST_44619_PI426541283 | e | AT3G12110.1 | cr_JHI_3-20 | 3486 | GO:0010583 (response to cyclopentenone) |
| CUST_24407_PI426541283 | b | AT5G12210.1 | cr_JHI_3-15 | 734 | GO:0009555 (pollen development) |
| CUST_37835_PI426541283 | b | AT1G30330.2 | cr_JHI_3-15 | 664 | GO:0009733 (response to auxin) GO:0009734 (auxin activated signalling pathway) |
| CUST_35866_PI426541283 | f | AT4G28210.1 | cr_JHI_3-15 | 65 | GO:0009793 (embryo development ending in seed dormancy) |
| CUST_13398_PI426541283 | d | AT2G04240.1 | cr_JHI_3-15 | 826 | GO:0009687 (abscisic acid metabolic process) GO:0009739 (response to gibberellin) |
| Trait | ||||||||
|---|---|---|---|---|---|---|---|---|
| Crumbly | Fruit Ripening | |||||||
| Microarray Probe | A. thaliana Gene ID | Position (cM) | LG | Scaffold | QTLs | GO Terms | ||
| Crumbly | Fruit Set | Green Fruit | ||||||
| CUST_54460_PI426541283 | AT1G62360.1 | 99.4 | 3 | 4 | X | X | X | GO:0048440 GO:0009691 |
| CUST_12159_PI426541283 | AT2G19130.1 | 105 | 3 | 858 | X | X | GO:0048544 | |
| CUST_35866_PI426541283 | AT4G28210.1 | 103.3 | 3 | 65 | X | X | X | GO:0009793 GO:0009409 |
| CUST_24407_PI426541283 | AT5G12210.1 | 102 | 3 | 734 | X | X | X | GO:0009555 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Scolari, L.M.; Hancock, R.D.; Hedley, P.E.; Morris, J.; Smith, K.; Graham, J. Combining QTL Mapping and Gene Expression Analysis to Elucidate the Genetic Control of ‘Crumbly’ Fruit in Red Raspberry (Rubus idaeus L.). Agronomy 2021, 11, 794. https://doi.org/10.3390/agronomy11040794
Scolari LM, Hancock RD, Hedley PE, Morris J, Smith K, Graham J. Combining QTL Mapping and Gene Expression Analysis to Elucidate the Genetic Control of ‘Crumbly’ Fruit in Red Raspberry (Rubus idaeus L.). Agronomy. 2021; 11(4):794. https://doi.org/10.3390/agronomy11040794
Chicago/Turabian StyleScolari, Luca M., Robert D. Hancock, Pete E. Hedley, Jenny Morris, Kay Smith, and Julie Graham. 2021. "Combining QTL Mapping and Gene Expression Analysis to Elucidate the Genetic Control of ‘Crumbly’ Fruit in Red Raspberry (Rubus idaeus L.)" Agronomy 11, no. 4: 794. https://doi.org/10.3390/agronomy11040794