Variation of Phenotypic Traits in Twelve Bambara Groundnut (Vigna subterranea (L.) Verdc.) Genotypes and Two F2 Bi-Parental Segregating Populations

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growing Conditions
2.2. Traits Recorded
2.3. Data Analysis
3. Results
3.1. Phenotypic Trait Variation in Twelve Bambara Groundnut Genotypes
3.2. Principal Components Analysis for Twelve Genotypes Based on Phenotypic Traits
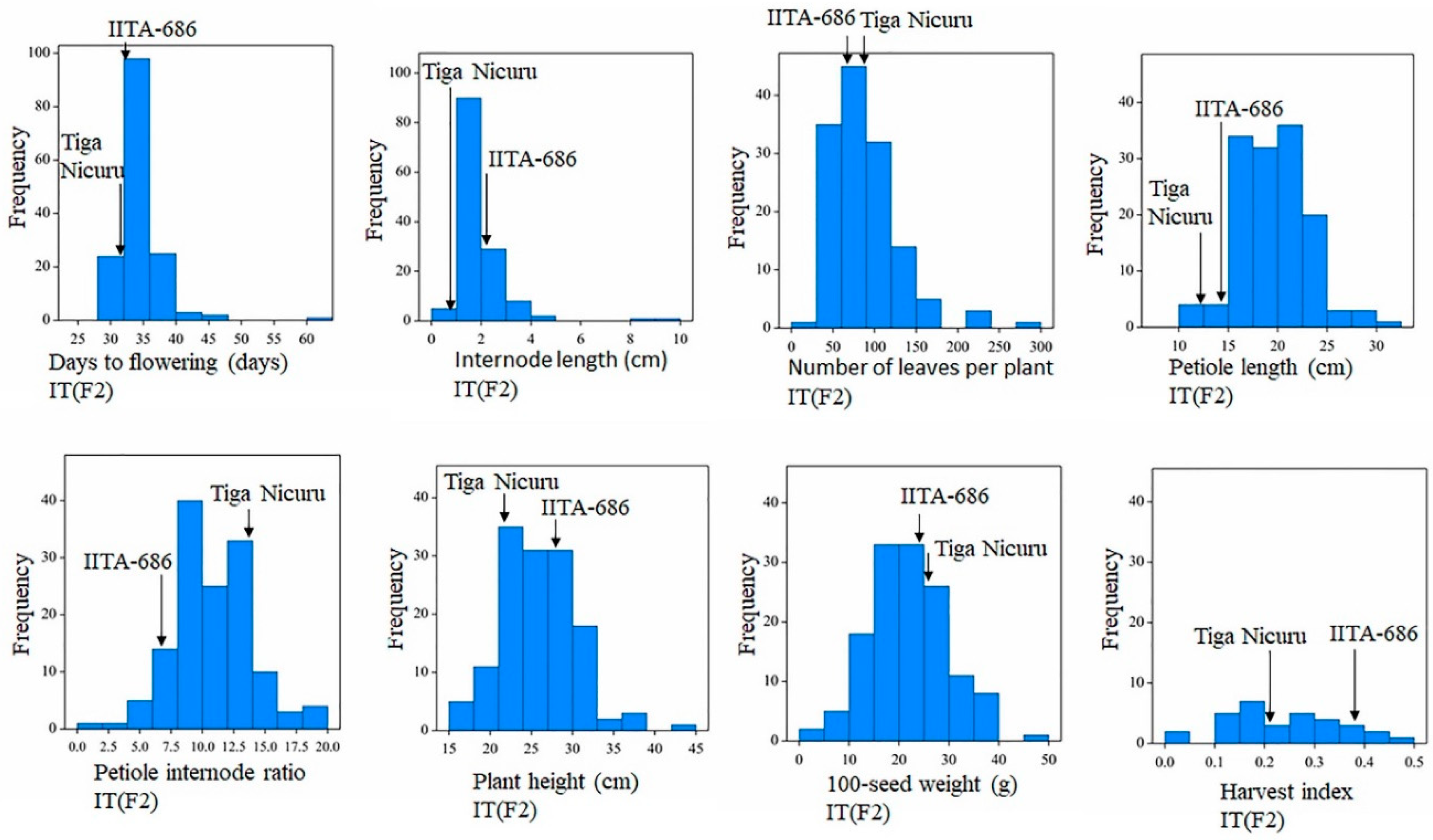
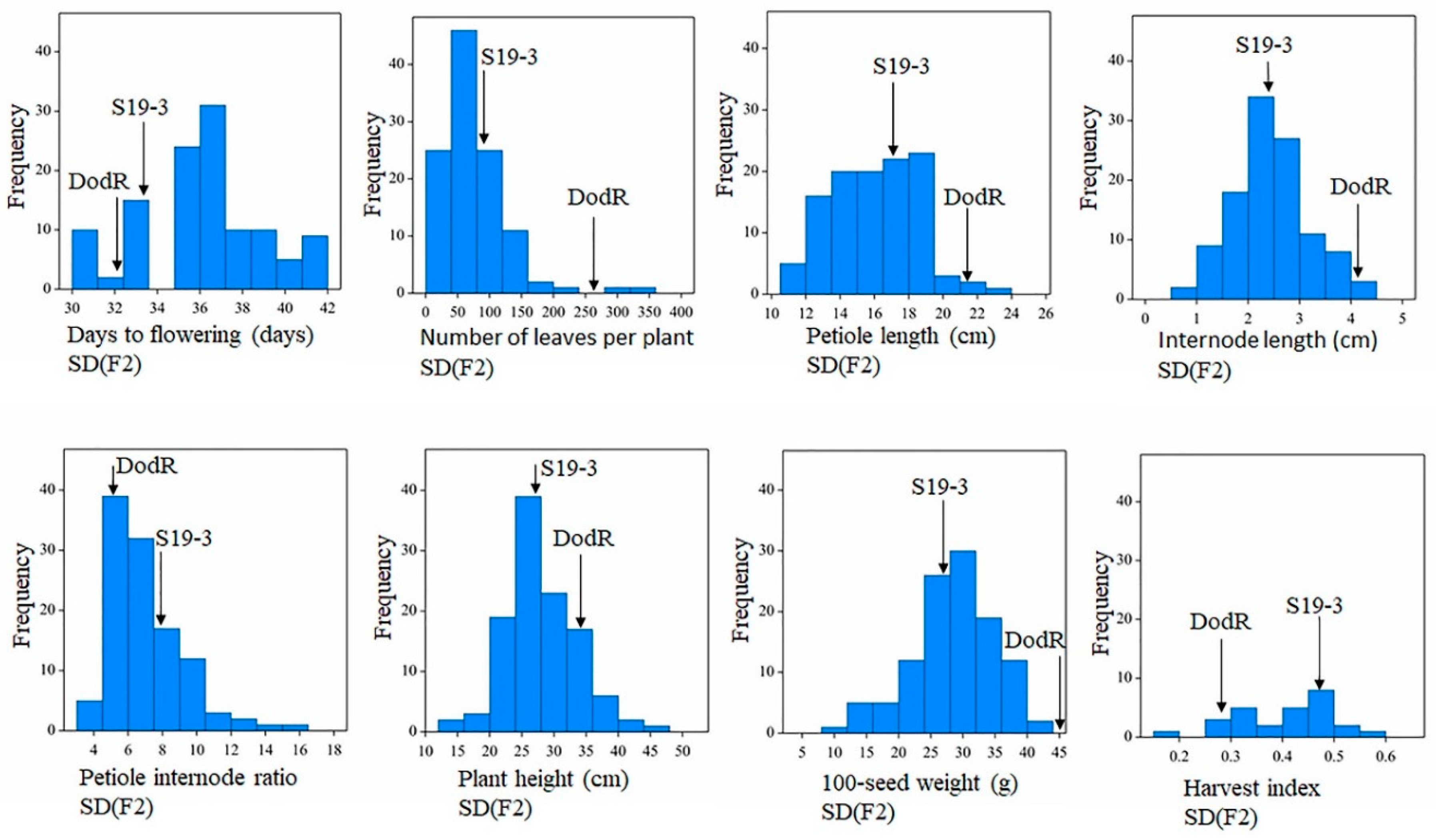
3.3. Phenotypic Trait Variations in the F2 bi-Parental Segregating Populations
3.4. Correlation Coefficient Analysis of Phenotypic Traits in the F2 bi-Parental Segregating Populations
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Azam-Ali, S.N.; Sesay, A.; Karikari, S.K.; Massawe, F.J.; Aguilar-Manjarrez, J.; Bannayan, M.; Hampson, K.J. Assessing the potential of an underutilized crop–A case study using bambara groundnut. Exp. Agric. 2001, 37, 433–472. [Google Scholar] [CrossRef]
- Massawe, F.; Mayes, S.; Cheng, A. Crop diversity: An unexploited treasure trove for food security. Trends Plant Sci. 2016, 21, 365–368. [Google Scholar] [CrossRef] [PubMed]
- Atoyebi, J.O.; Oyatomi, O.; Osilesi, O.; Adebawo, O.; Abberton, M. Morphological characterisation of selected african accessions of bambara groundnut (Vigna subterranea (L.) Verdc.). Int. J. Plant Res. 2017, 7, 29–35. [Google Scholar] [CrossRef]
- Mabhaudhi, T.; Chibarabada, T.P.; Chimonyo, V.G.P.; Modi, A.T. Modelling climate change impact: A Case of bambara groundnut (Vigna subterranea). Phys. Chem. Earth 2018, 105, 25–31. [Google Scholar] [CrossRef]
- Blay, E.T.; Offei, S.K.; Danquah, E.Y. Genetic diversity in cocoyam as revealed by random amplified polymorphic DNA (RAPD) markers. In Proceedings of the Final Research Coordination Meeting on Genetic Improvement Of Under-utilized And Neglected Crops In low Income Food Defict Countires Through Irradiation And Related Techiques, Pretoria, South Africa, 19–23 May 2003; pp. 131–142. [Google Scholar]
- Padulosi, S.; Hoeschle-Zeledon, I. Underutilized Plant Species: What Are They? Leisa Mag. 2004, 20, 5–6. [Google Scholar]
- Jaenicke, H.; Höschle-Zeledon, I. (Eds.) Strategic Framework for Underutilized Plant Species Research and Development, with Special Reference to Asia and the Pacific, and to Sub-Saharan Africa; International Centre for Underutilised Crops, Colombo, Sri Lanka and Global Facilitation Unit for Underutilized Species: Rome, Italy, 2006; p. 33. [Google Scholar]
- Mayes, S.; Massawe, F.J.; Alderson, P.G.; Roberts, J.A.; Azam-Ali, S.N.; Hermann, M. The potential for underutilized crops to improve security of food production. J. Exp. Bot. 2012, 63, 1075–1079. [Google Scholar] [CrossRef] [Green Version]
- Mayes, S.; Ho, W.K.; Chai, H.H.; Gao, X.; Kundy, A.C.; Mateva, K.I.; Zahrulakmal, M.; Hahiree, M.K.I.M.; Kendabie, P.; Licea, L.C.S.; et al. Bambara groundnut: An exemplar underutilised legume for resilience under climate change. Planta 2019, 250, 803–820. [Google Scholar] [CrossRef] [Green Version]
- Williams, J.T.; Haq, N. Global Research on Underutilised Crops: An Assessment of Current Activities and Proposals for Enhanced Cooperation; International Centre for Underutilised Crops: Southampton, UK, 2000; Volume 50. [Google Scholar]
- Tanzi, A.S.; Ho, W.K.; Massawe, F.; Mayes, S. Development and interaction between plant architecture and yield-related traits in winged bean (Psophocarpus Tetragonolobus (L.) DC.). Euphytica 2019, 215, 1–14. [Google Scholar] [CrossRef]
- Sarkar, A.; Sensarma, S.R.; VanLoon, G.W. Sustainable Solutions for Food Security: Combating Climate Change by Adaptation; Springer: Cham, Switzerland, 2019. [Google Scholar] [CrossRef]
- Aremu, M.O.; Olaofe, O.; Akintayo, E.T. Chemical composition and physicochemical characteristics of two varieties of bambara groundnut (Vigna Subterrenea) flours. J. Appl. Sci. 2006, 6, 1900–1903. [Google Scholar] [CrossRef] [Green Version]
- Massawe, F.; Mwale, S.S.; Azam-Ali, S.; Roberts, J. Breeding in Bambara Groundnut (Vigna subterranea (L.) Verdc.): Strategic Considerations. Afr. J. Biotechnol. 2005, 4, 463–471. [Google Scholar]
- Oyeyinka, S.A.; Singh, S.; Adebola, P.O.; Gerrano, A.S.; Amonsou, E.O. Physicochemical Properties of starches with variable amylose contents extracted from bambara groundnut genotypes. Carbohydr. Polym. 2015, 133, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Massawe, F.; Schenkel, W.; Basu, S.; Temba, E.M. Artificial hybridisation in bambara groundnut (Vigna subterranea (L.) Verdc.). In Proceedings of the International Bambara Groundnut Symposium, Gaborone, Botswana, 5–12 September 2003; Botswana Colleage of Agriculture: Francistown, Botswana, 2003; pp. 1193–1209. [Google Scholar]
- Basu, S.; Mayes, S.; Davey, M.; Roberts, J.A.; Azam-Ali, S.N.; Mithen, R.; Pasquet, R.S. Inheritance of “domestication” traits in bambara groundnut (Vigna subterranea (L.) Verdc.). Euphytica 2007, 157, 59–68. [Google Scholar] [CrossRef]
- Berchie, J.N.; Sarkodie-Addo, J.; Adu-Dapaah, H.; Agyemang, A.; Addy, S.; Asare, E.; Donkor, J. Yield evaluation of three early maturing bambara groundnut (Vigna subterranea L. Verdc) Landraces at the CSIR-Crops research institute, Fumesua-Kumasi, Ghana. J. Agron. 2010, 9, 175–179. [Google Scholar] [CrossRef]
- Victoria, L.; Benson, O.; Beatrice, A.; Regina, N.; Koech, P.K.; Skilton, R.A.; Francesca, S. Morphological, genetic and symbiotic characterization of Root Nodule Bacteria Isolated from Bambara Groundnuts (Vigna subterranea L. Verdc) from soils of Lake Victoria Basin, Western Kenya. J. Appl. Biol. Biotechnol. 2015. [Google Scholar] [CrossRef] [Green Version]
- Suwanprasert, J.; Toojinda, T.; Srinives, P.; Chanprame, S. Hybridization Technique for Bambara Groundnut. Breed. Sci. 2006, 56, 125–129. [Google Scholar] [CrossRef] [Green Version]
- Okpuzor, J.; Ogbunugafor, H.A.; Okafor, U.; Sofidiya, M.O. Identification of Protein Types in Bambara Nut Seeds: Perspectives for Dietary Protein Supply in Developing Countries. EXCLI J. 2010, 9, 17–28. [Google Scholar] [CrossRef]
- Azman Halimi, R.; Barkla, B.J.; Mayes, S.; King, G.J. The potential of the underutilized pulse bambara groundnut (Vigna subterranea (L.) Verdc.) for nutritional food security. J. Food Compos. Anal. 2019, 77, 47–59. [Google Scholar] [CrossRef]
- Basu, S.; Roberts, J.A.; Azam-Ali, S.N.; Mayes, S. Development of microsatellite markers for bambara groundnut (Vigna subterranea L. Verdc.)—An underutilized african legume crop species. Mol. Ecol. Notes 2007, 7, 1326–1328. [Google Scholar] [CrossRef]
- Olukolu, B.A.; Mayes, S.; Stadler, F.; Ng, N.Q.; Fawole, I.; Dominique, D.; Azam-Ali, S.N.; Abbott, A.G.; Kole, C. Genetic Diversity in bambara groundnut (Vigna subterranea (L.) Verdc.) as Revealed by phenotypic descriptors and DArT marker analysis. Genet. Resour. Crop EVol 2012, 59, 347–358. [Google Scholar] [CrossRef]
- Aliyu, S.; Massawe, F.; Mayes, S. Beyond Landraces: Developing improved germplasm resources for underutilized species—A case for bambara groundnut. Biotechnol. Genet. Eng. Rev. 2015, 30, 127–141. [Google Scholar] [CrossRef]
- Mayes, S.; Ho, W.K.; Kendabie, P.; Chai, H.H.; Aliyu, S.; Feldman, A.; Halimi, R.A.; Massawe, F.; Azam-Ali, S. Applying molecular genetics to underutilised species—Problems and opportunities. Malays. Appl. Biol. 2015, 44, 1–9. [Google Scholar]
- Kendabie, P.; Jorgensen, S.T.; Massawe, F.; Fernandez, J.; Azam-Ali, S.; Mayes, S. Photoperiod control of yield and sink capacity in Bambara groundnut (Vigna subterranea) genotypes. Food Energy Secur. 2020, e240. [Google Scholar] [CrossRef]
- Camberlin, P.; Martiny, N.; Philippon, N.; Richard, Y. Determinants of the interannual relationships between remote sensed photosynthetic activity and rainfall in tropical africa. Remote Sens. Environ. 2007, 106, 199–216. [Google Scholar] [CrossRef] [Green Version]
- Siebert, A. Hydroclimate extremes in africa: Variability, observations and modeled projections. Geogr. Compass 2014, 8, 351–367. [Google Scholar] [CrossRef]
- Redjeki, E.S.; Mayes, S.; Azam-Ali, S. Evaluating the stability and adaptability of bambara groundnut (Vigna subterranea (L.) Verd.) landraces in different agro-ecologies. Acta Hortic. 2013, 979, 389–400. [Google Scholar] [CrossRef]
- Mateva, K.I.; Chai, H.H.; Mayes, S.; Massawe, F. Root foraging capacity in bambara groundnut (Vigna subterranea (L.) Verdc.) core parental lines depends on the root system architecture during the pre-flowering stage. Plants 2020, 9, 645. [Google Scholar] [CrossRef] [PubMed]
- Jørgensen, S.T.; Liu, F.; Ouédraogo, M.; Ntundu, W.H.; Sarrazin, J.; Christiansen, J.L. Drought responses of two bambara groundnut (Vigna subterranea L. Verdc.) landraces collected from a dry and a humid area of africa. J. Agron. Crop Sci. 2010, 196, 412–422. [Google Scholar] [CrossRef]
- Mwale, S.S.; Azam-Ali, S.N.; Massawe, F.J. Growth and development of bambara groundnut (Vigna subterranea) in response to soil moisture. 2. resource capture and conversion. Eur. J. Agron. 2007, 26, 354–362. [Google Scholar] [CrossRef]
- Mwale, S.S.; Azam-Ali, S.N.; Massawe, F.J. Growth and development of bambara groundnut (Vigna subterranea) in response to soil moisture: 1. dry matter and yield. Eur. J. Agron. 2007, 26, 345–353. [Google Scholar] [CrossRef]
- IPGRI/IITA/BAMNET. Descriptors for Vigna subterranea bambara groundnut (Vigna subterranea). In International Plant Genetic Resources Institute, Rome; International Institute of Tropical Agriculture, Ibadan; The International Bambara Groundnut Network: Hamburg, Germany, 2000. [Google Scholar]
- Molosiwa, O.O.; Aliyu, S.; Stadler, F.; Mayes, K.; Massawe, F.; Kilian, A.; Mayes, S. SSR marker development, genetic diversity and population structure analysis of bambara groundnut [Vigna subterranea (L.) Verdc.] landraces. Genet. Resour. Crop EVol 2015, 62, 1225–1243. [Google Scholar] [CrossRef]
- Aliyu, S.; Massawe, F.; Mayes, S. Genetic diversity and population structure of bambara groundnut (Vigna subterranea (L.) Verdc.): Synopsis of the past two decades of analysis and implications for crop improvement programmes. Genet. Resour. Crop EVol 2016, 63, 925–943. [Google Scholar] [CrossRef] [Green Version]
- Karikari, S.K.; Tabona, T.T. Constitutive traits and selective indices of bambara groundnut (Vigna subterranea (L) Verdc) landraces for drought tolerance under botswana conditions. Phys. Chem. Earth 2004, 29, 1029–1034. [Google Scholar] [CrossRef]
- Pedercini, M.; Kanamaru, H.; Derwisch, S. Potential Impacts of Climate Change on Food Security in Mali Potential Impacts of Climate Change on Food Security in Mali; Natural Resources Management and Environment Department, Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 2012. [Google Scholar]
- Chai, H.H.; Massawe, F.; Mayes, S. Effects of mild drought stress on the morpho-physiological characteristics of a bambara groundnut segregating population. Euphytica 2016, 208, 225–236. [Google Scholar] [CrossRef]
- Klimek-Kopyra, A.; Zajac, T.; Oleksy, A.; Kulig, B.; Slizowska, A. The value of different vegetative indices (NDVI, GAI) for the assessment of yield potential of pea (Pisum Sativum L.) at different growth stages and under varying management practices. Acta Agrobot. 2018, 71. [Google Scholar] [CrossRef]
- Xing, Y.; Zhang, Q. Genetic and molecular bases of rice yield. Annu. Rev. Plant Biol. 2010, 61, 421–442. [Google Scholar] [CrossRef] [PubMed]
- Mathan, J.; Bhattacharya, J.; Ranjan, A. Enhancing crop yield by optimizing plant developmental features. Development 2016, 143, 3283–3294. [Google Scholar] [CrossRef] [Green Version]
- Mabhaudhi, T.; Modi, A.T. Growth, phenological and yield responses of a bambara groundnut (Vigna subterranea (L.) Verdc.) landrace to imposed water stress under field conditions. S. Afr. J. Plant Soil 2013, 30, 69–79. [Google Scholar] [CrossRef]
- Ntundu, W.H.; Shillah, S.A.; Marandu, W.Y.F.; Christiansen, J.L. Morphological diversity of bambara groundnut [Vigna subterranea (L.) Verdc.] landraces in Tanzania. Genet. Resour. Crop EVol 2006, 53, 367–378. [Google Scholar] [CrossRef]
- Mickelbart, M.V.; Hasegawa, P.M.; Bailey-Serres, J. Genetic mechanisms of abiotic stress tolerance that translate to crop yield stability. Nat. Rev. Genet. 2015, 16, 237–251. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Geographical Origin | Landraces | Collected Country | Annual Rainfall (mm) | Distinctive Characteristics |
|---|---|---|---|---|
| East Africa | DodR | Tanzania | 1000 [28,29] | Quantitative for short days, high 100-seed weight and yield [27,30] |
| IITA-686 | Tanzania | 1000 [28,29] | Quantitative for long days, shallow and highly branched root growth habit [27,31] | |
| TAN385 | Tanzania | 1000 [28,29] | - | |
| West Africa | LunT | Sierra Leone | ˃2000 [31] | Quantitative for short days, shallow and highly branched root growth habit [27,31] |
| Tiga Nicuru | Mali | 450 [28,32] | Quantitative for short days, bunchy growth habit, early maturity [27,33,34] | |
| Ankpa4 | Nigeria | >2000 [29,31] | Qualitative for short days [27] | |
| Getso | Nigeria | >2000 [29,31] | Quantitative for short days [27] | |
| Southern Africa | S19-3 | Namibia | 365 [29,32] | Quantitative for long days, early maturity, drought tolerant long taproots and great root length distribution [27,31,32,34] |
| Uniswa red | Kingdom of Eswatini | 1390 [32] | Quantitative for long days, long growth cycle [27,34] | |
| DipC | Botswana | 500 [31] | Quantitative for long days, long taproots and great root length distribution [27,31] | |
| AHM | Namibia | 365 [29,32] | - | |
| Southeast Asia | Gresik | Indonesia | ˃2000 [31] | Quantitative for short days, shallow and highly branched root growth habit [27,31] |
| Traits | DTF | NL | PH | PL | IL | P/I | HI | 100SW | SP |
|---|---|---|---|---|---|---|---|---|---|
| (days) | (cm) | (cm) | (cm) | (ratio) | (g) | (%) | |||
| AHM | 35.50 cd | 532.80 a | 35.20 a | 26.13 ab | 3.76 ab | 7.08 d | 0.09 cd | 21.45 efg | 77.70 a |
| Ankpa4 | 42.40 b | 198.20 b | 33.79 ab | 23.49 abc | 2.86 c | 8.23 cd | 0.04 d | 20.09 fg | 39.58 d |
| DipC | 38.57 bc | 250.40 b | 34.50 a | 27.04 a | 2.49 cde | 10.89 ab | 0.14 cd | 29.03 cdef | 75.10 a |
| DodR | 32.10 de | 266.50 b | 34.35 a | 21.60 c | 4.04 a | 5.55 e | 0.28 abc | 45.49 cdef | 78.01 a |
| Getso | 30.80 e | 82.10 c | 31.05 abc | 15.23 de | 1.83 ef | 8.49 cd | 0.16 bcd | 52.07 a | 62.74 bc |
| Gresik | 51.19 a | 159.60 b | 31.30 abc | 15.23 de | 2.11 de | 7.20 d | 0.26 abc | 33.32 cd | 74.54 a |
| IITA-686 | 32.50 de | 66.00 c | 28.15 bcd | 14.79 de | 2.09 de | 7.15 d | 0.38 ab | 24.68 def | 75.73 a |
| LunT | 33.00 de | 184.00 b | 34.44 a | 25.36 abc | 2.60 cd | 9.65 bc | 0.26 abc | 37.26 bc | 71.53 ab |
| S19-3 | 33.40 de | 84.80 c | 27.33 cd | 17.44 d | 2.46 cde | 7.30 d | 0.47 a | 27.47 cdef | 77.87 a |
| TAN385 | 33.50 de | 768.10 a | 34.50 a | 23.07 bc | 3.15 bc | 7.42 d | 0.03 d | 13.04g | 61.74 c |
| Tiga Nicuru | 27.13 f | 81.10 c | 22.71 d | 12.52 e | 0.96 g | 13.36 a | 0.21 bcd | 24.58 def | 74.08 a |
| Uniswa Red | 33.44 de | 80.30 c | 28.33 bcd | 14.64 de | 1.13 fg | 13.24 a | 0.18 bcd | 31.05 cde | 73.21 a |
| Mean | 35.30 | 229.42 | 31.31 | 19.71 | 2.46 | 8.80 | 0.21 | 29.95 | 70.15 |
| F pr | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
| Traits | DTF | NL | PH | PL | IL | P/I | HI | 100SW | SP |
|---|---|---|---|---|---|---|---|---|---|
| DTF | - | - | - | - | - | - | - | - | - |
| NL | 0.05 | - | - | - | - | - | - | - | - |
| PH | 0.37 | 0.62 * | - | - | - | - | - | - | - |
| PL | 0.17 | 0.61 * | 0.86 ** | - | - | - | - | - | - |
| IL | 0.15 | 0.64 * | 0.79 ** | 0.74 ** | - | - | - | - | - |
| P/I | −0.28 | −0.35 | −0.50 | −0.27 | −0.77 ** | - | - | - | - |
| HI | −0.15 | −0.60 * | −0.49 | −0.45 | −0.18 | −0.18 | - | - | - |
| 100SW | −0.13 | −0.50 | 0.04 | −0.21 | −0.07 | −0.07 | 0.31 | - | - |
| SP | −0.23 | −0.12 | −0.25 | −0.18 | −0.04 | 0.03 | 0.59 * | 0.22 | - |
| IITA-686 | Tiga Nicuru | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Traits | Mean | Min | Max | SD | Variance | Normality | Min | Max | Min | Max |
| DTF | 34.98 | 31.00 | 61.00 | 3.54 | 12.51 | <0.001 | 29.00 | 34.00 | 27.00 | 28.00 |
| NL | 87.24 | 17.00 | 297.00 | 42.56 | 1811.00 | <0.001 | 28.00 | 114.00 | 43.00 | 116.00 |
| PH (cm) | 26.18 | 16.00 | 44.00 | 4.69 | 22.01 | 0.03 | 18.00 | 34.50 | 19.50 | 27.00 |
| PL (cm) | 19.72 | 11.83 | 30.67 | 3.50 | 12.23 | 0.13 | 10.10 | 19.75 | 10.60 | 15.57 |
| IL (cm) | 2.03 | 0.85 | 4.90 | 0.69 | 0.48 | <0.001 | 1.53 | 2.75 | 0.73 | 1.13 |
| P/I | 11.22 | 5.21 | 22.90 | 3.24 | 10.48 | 0.09 | 5.52 | 8.85 | 9.71 | 16.91 |
| HI | 0.24 | 0.01 | 0.46 | 0.11 | 0.01 | 0.45 | 0.06 | 0.76 | 0.06 | 0.36 |
| 100SW (g) | 21.85 | 4.69 | 45.56 | 7.70 | 59.31 | 0.57 | 15.67 | 29.50 | 19.90 | 35.21 |
| S19-3 | DodR | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Traits | Mean | Min | Max | SD | Variance | Normality | Min | Max | Min | Max |
| DTF | 36.41 | 31.00 | 42.00 | 2.81 | 7.90 | <0.001 | 29.00 | 36.00 | 29.00 | 36.00 |
| NL | 77.67 | 15.00 | 325.00 | 50.61 | 2561.00 | <0.001 | 6.00 | 133.00 | 95.00 | 396.00 |
| PH (cm) | 28.34 | 13.30 | 46.50 | 5.66 | 32.02 | 0.02 | 18.30 | 33.00 | 27.00 | 44.00 |
| PL (cm) | 16.20 | 10.83 | 23.33 | 2.54 | 6.43 | 0.44 | 7.17 | 24.50 | 18.10 | 24.50 |
| IL (cm) | 2.48 | 0.73 | 4.50 | 0.77 | 0.59 | 0.55 | 1.17 | 3.55 | 3.00 | 5.00 |
| P/I | 7.09 | 3.89 | 16.36 | 2.15 | 4.64 | <0.001 | 5.63 | 9.37 | 3.81 | 7.50 |
| HI | 0.40 | 0.16 | 0.56 | 0.10 | 0.01 | 0.33 | 0.32 | 0.85 | 0.16 | 0.40 |
| 100SW (g) | 28.72 | 10.79 | 41.75 | 6.40 | 40.91 | 0.05 | 16.70 | 39.58 | 37.88 | 52.70 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, X.; Bamba, A.S.A.; Kundy, A.C.; Mateva, K.I.; Chai, H.H.; Ho, W.K.; Musa, M.; Mayes, S.; Massawe, F. Variation of Phenotypic Traits in Twelve Bambara Groundnut (Vigna subterranea (L.) Verdc.) Genotypes and Two F2 Bi-Parental Segregating Populations. Agronomy 2020, 10, 1451. https://doi.org/10.3390/agronomy10101451
Gao X, Bamba ASA, Kundy AC, Mateva KI, Chai HH, Ho WK, Musa M, Mayes S, Massawe F. Variation of Phenotypic Traits in Twelve Bambara Groundnut (Vigna subterranea (L.) Verdc.) Genotypes and Two F2 Bi-Parental Segregating Populations. Agronomy. 2020; 10(10):1451. https://doi.org/10.3390/agronomy10101451
Chicago/Turabian StyleGao, Xiuqing, Aliyu Siise Abdullah Bamba, Aloyce Callist Kundy, Kumbirai Ivyne Mateva, Hui Hui Chai, Wai Kuan Ho, Mukhtar Musa, Sean Mayes, and Festo Massawe. 2020. "Variation of Phenotypic Traits in Twelve Bambara Groundnut (Vigna subterranea (L.) Verdc.) Genotypes and Two F2 Bi-Parental Segregating Populations" Agronomy 10, no. 10: 1451. https://doi.org/10.3390/agronomy10101451