
Chitosan: Properties and Its Application in Agriculture in Context of Molecular Weight
 , , ,
, , ,
Abstract
:1. Introduction
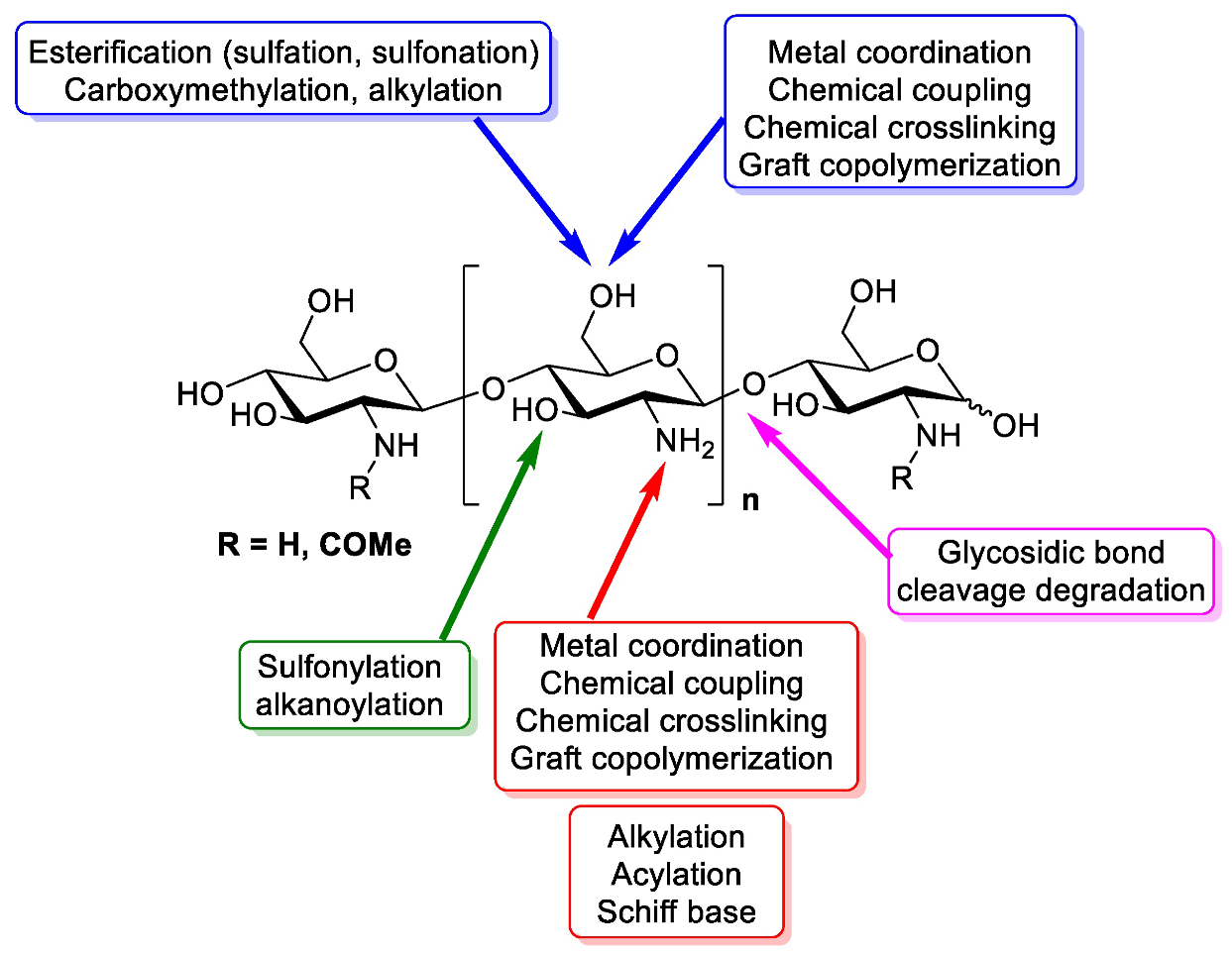
2. Chitosan: Structure and Natural Origins
3. General Characteristics of Chitosan
4. Properties
4.1. Low-Molecular-Weight Chitosan
4.1.1. Physicochemical Properties
4.1.2. Biological Properties
4.2. Medium-Molecular-Weight Chitosan
4.2.1. Physicochemical Properties
4.2.2. Biological Properties
4.3. High-Molecular-Weight Chitosan
4.3.1. Physicochemical Properties
4.3.2. Biological Properties
5. Applications in the Agricultural Sector
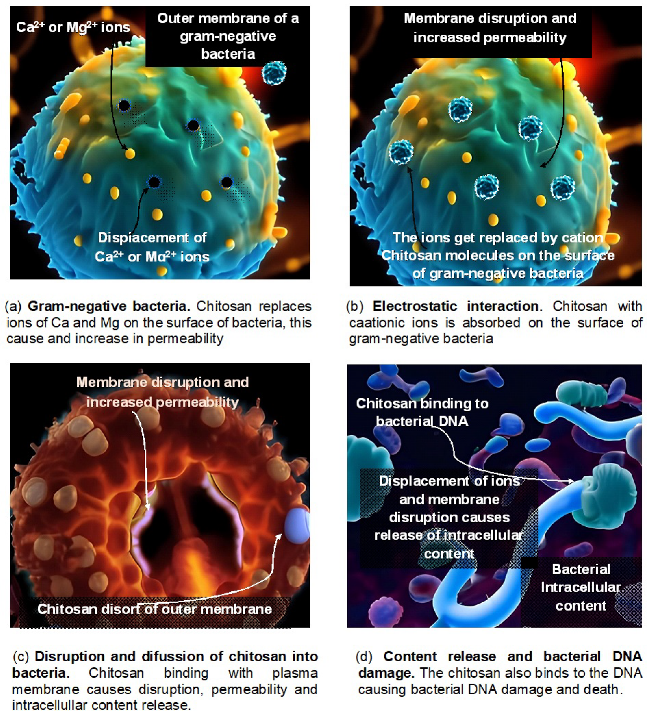
5.1. Antimicrobial Properties
5.1.1. Low-Molecular-Weight Chitosan
5.1.2. Medium-Molecular-Weight Chitosan
5.1.3. High-Molecular-Weight Chitosan
5.2. Antiviral Properties
5.2.1. Low-Molecular-Weight Chitosan
5.2.2. Medium-Molecular-Weight Chitosan
5.2.3. High-Molecular-Weight Chitosan
5.3. Antifungal Properties
5.3.1. Low-Molecular-Weight Chitosan
5.3.2. Medium-Molecular-Weight Chitosan
5.3.3. High Molecular Weight Chitosan
5.4. Chitosan’s Induction of Plant Protection Mechanisms
5.4.1. Low Molecular Weight Chitosan
5.4.2. Medium Molecular Weight Chitosan
5.4.3. High-Molecular-Weight Chitosan
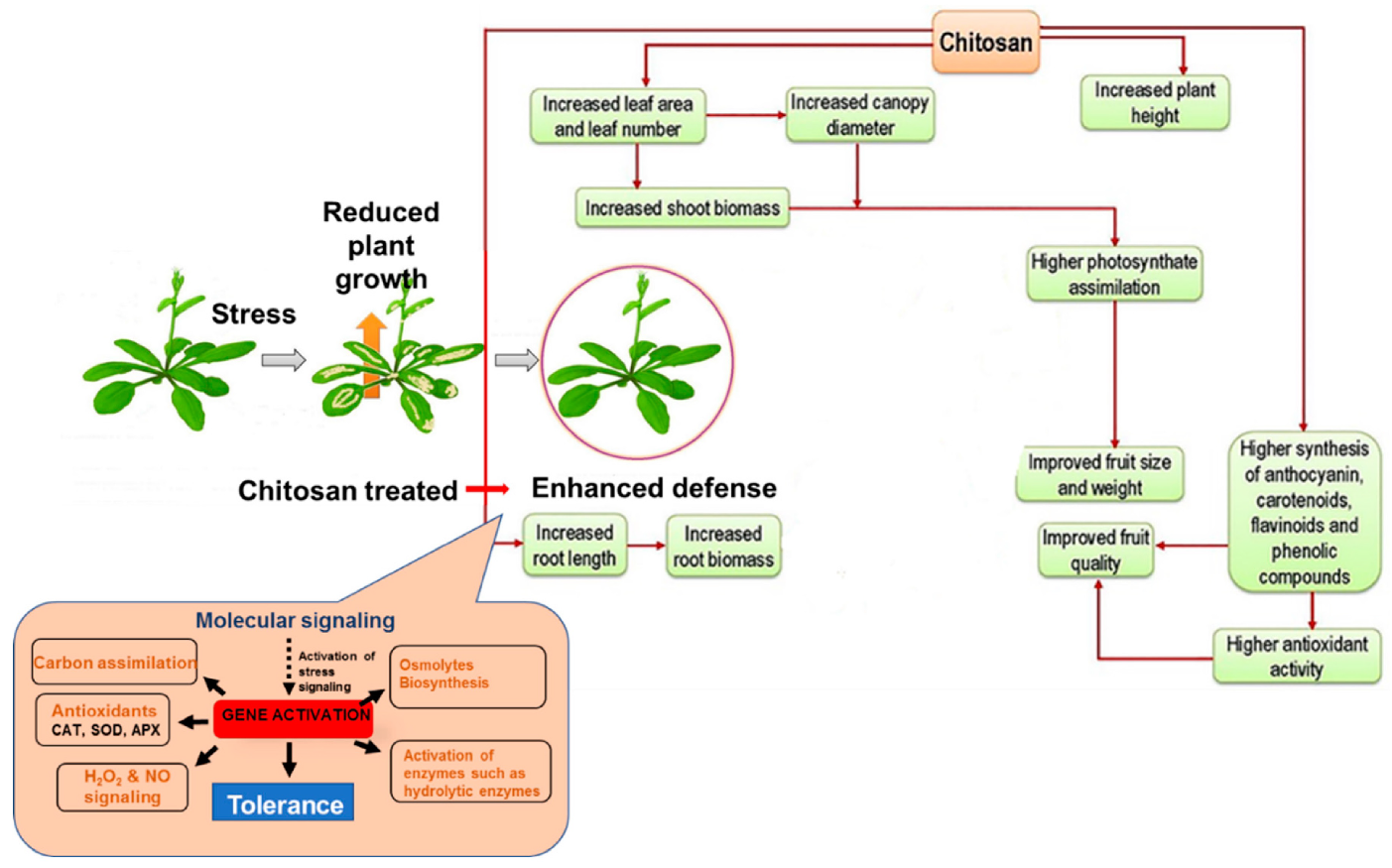
5.5. Growth Promoter
5.5.1. Low-Molecular-Weight Chitosan
5.5.2. Medium-Molecular-Weight Chitosan
5.5.3. High Molecular Weight Chitosan
6. Common Presentation
6.1. In Solution
6.2. Hydrogel
6.3. Chitosan-Based Nanocomposites
7. A Perspective of the Authors
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Davis, S. Chitosan: Manufacture, Properties, and Usage; Nova Science Publishers: New York, NY, USA, 2011. [Google Scholar]
- Huq, T.; Khan, A.; Brown, D.; Dhayagude, N.; He, Z.; Ni, Y. Sources, production and commercial applications of fungal chitosan: A review. J. Bioresour. Bioprod. 2022, 7, 85–98. [Google Scholar] [CrossRef]
- Crini, G.; Lichtfouse, E.; Crini, N.; Torri, G. Chitin and Chitosan: History, Fundamentals and Innovations. In Sustainable Agriculture Reviews; Crini, G., Lichtfouse, E., Eds.; Springer: Cham, Switzerland, 2019; Volume 35, pp. 49–123. [Google Scholar]
- Islam, M.M.; Shahruzzaman, M.; Biswas, S.; Sakib, M.N.; Rashid, T.U. Chitosan based bioactive materials in tissue engineering applications—A review. Bioact. Mater. 2020, 5, 164–183. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Vázquez, M.; Vega-Ruiz, B.; Ramos-Zúñiga, R.; Saldaña-Koppel, D.A.; Quiñones-Olvera, L.F. Chitosan and its potential use as a scaffold for tissue engineering in regenerative medicine. BioMed Res. Int. 2015, 2015, 821279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madni, A.; Kousar, R.; Naeem, N.; Wahid, F. Recent advancements in applications of chitosan-based biomaterials for skin tissue engineering. J. Bioresour. Bioprod. 2021, 6, 11–25. [Google Scholar] [CrossRef]
- Raza, Z.A.; Khalil, S.; Ayub, A.; Banat, I.M. Recent developments in chitosan encapsulation of various active ingredients for multifunctional applications. Carbohydr. Res. 2020, 492, 108004. [Google Scholar] [CrossRef]
- Ma, Z.; Garrido-Maestu, A.; Jeong, K.C. Application, mode of action, and in vivo activity of chitosan and its micro-and nanoparticles as antimicrobial agents: A review. Carbohydr. Polym. 2017, 176, 257–265. [Google Scholar] [CrossRef]
- Flórez, M.; Guerra-Rodríguez, E.; Cazón, P.; Vázquez, M. Chitosan for food packaging: Recent advances in active and intelligent films. Food Hydrocoll. 2022, 124, 107328. [Google Scholar] [CrossRef]
- Cazón, P.; Vázquez, M. Applications of chitosan as food packaging materials. Sustain. Agric. Rev. 2019, 6, 81–123. [Google Scholar]
- Saad, E.M.; Elshaarawy, R.F.; Mahmoud, S.A.; El-Moselhy, K.M. New ulva lactuca algae based chitosan bio-composites for bioremediation of Cd (II) ions. J. Bioresour. Bioprod. 2021, 6, 223–242. [Google Scholar] [CrossRef]
- Alsharari, S.F.; Tayel, A.A.; Moussa, S.H. Soil emendation with nano-fungal chitosan for heavy metals biosorption. Int. J. Biol. Macromol. 2018, 118, 2265–2268. [Google Scholar] [CrossRef]
- Rusdiarso, B.; Basuki, R. Stability improvement of humic acid as sorbent through magnetite and chitin modification. J. Kim. Sains Dan Apl. 2020, 23, 152–159. [Google Scholar] [CrossRef]
- Alterary, S.S.; Alshahrani, A.A.; Alsahli, S.A. Fabrication of novel buckypaper metal oxide nano-catalysis glycerol carbonate/MWCNTs membrane for efficient removal of heavy metals. Heliyon 2022, 8, e12633. [Google Scholar] [CrossRef]
- Wei, L.; Deng, W.; Li, S.; Wu, Z.; Cai, J.; Luo, J. Sandwich-like chitosan porous carbon Spheres/MXene composite with high specific capacitance and rate performance for supercapacitors. J. Bioresour. Bioprod. 2022, 7, 63–72. [Google Scholar] [CrossRef]
- Pandey, P.; Verma, M.; De, N. Chitosan in agricultural context—A review. Bull. Environ. Pharmacol. Life Sci. 2018, 3, 87–96. [Google Scholar]
- Sharif, R.; Mujtaba, M.; Ur Rahman, M.; Shalmani, A.; Ahmad, H.; Anwar, T.; Tianchan, D.; Wang, X. The Multifunctional Role of Chitosan in Horticultural Crops; A review. Molecules 2018, 23, 872. [Google Scholar] [CrossRef] [Green Version]
- Dima, J.B.; Sequeiros, C.; Zaritzky, N. Chitosan from marine crustaceans: Production, characterization and applications. In Biological Activities and Application of Marine Polysaccharides; IntechOpen: London, UK, 2017; pp. 39–56. [Google Scholar]
- Aranaz, I.; Alcántara, A.; Civera, M.; Arias, C.; Elorza, B.; Heras Caballero, A.; Acosta, N. Chitosan: An Overview of Its Properties and Applications. Polymers 2021, 13, 3256. [Google Scholar] [CrossRef]
- Crespo, M.; Martínez, M.; Hernández, J.; Lage Yusty, M. High-performance liquid chromatographic determination of chitin in the snow crab, Chionoecetes opilio. J. Chromatogr. 2006, 1116, 189–192. [Google Scholar] [CrossRef]
- Sunita, D. Extraction of Chitin from Trash Crabs (Podophthalmus vigil) by an Eccentric Method. Curr. Res. J. Biol. Sci. 2010, 2, 72–75. [Google Scholar]
- Sperstad, S.; Haug, T.; Paulsen, V.; Rode, T.; Strandskog, G.; Solem, S.; Styrvold, B.; Stensvåg, K. Characterization of crustins from the hemocytes of the spider crab, Hyas araneus, and the red king crab, Paralithodes camtschaticus. Dev. Comp. Immunol. 2009, 33, 583–591. [Google Scholar] [CrossRef]
- Hajji, S.; Younes, I.; Ghorbel-Bellaaj, O.; Hajji, R.; Rinaudo, M.; Nasri, M.; Jellouli, K. Structural differences between chitin and chitosan extracted from three different marine sources. Int. J. Biol. Macromol. 2014, 65, 298–306. [Google Scholar] [CrossRef]
- Abdou, E.; Nagy, K.; Elsabee, M. Extraction and characterization of chitin and chitosan from local sources. Bioresour. Technol. 2008, 99, 1359–1367. [Google Scholar] [CrossRef] [PubMed]
- Mahlous, M.; Tahtat, D.; Benamer, S.; Khodja, A. Gamma irradiation-aided chitin/chitosan extraction from prawn shells. Nucl. Instrum. Methods Phys. Res. Sect. B Beam Interact. Mater. Atoms 2007, 265, 414–417. [Google Scholar] [CrossRef]
- Kaya, M.; Cakmak, Y.; Baran, T.; Asan-Ozusaglam, M.; Menteş, A.; Tozak, K. New chitin, chitosan, and O-carboxymethyl chitosan sources from resting eggs of Daphnia longispina (Crustacea); with physicochemical characterization, and antimicrobial and antioxidant activities. Biotechnol. Bioprocess Eng. 2014, 19, 58–69. [Google Scholar] [CrossRef]
- Kaya, M.; Baran, T.; Menteş, A.; Asaroglu, M.; Sezen, G.; Tozak, K. Extraction and Characterization of α-Chitin and Chitosan from Six Different Aquatic Invertebrates. Food Biophys. 2014, 9, 145–157. [Google Scholar] [CrossRef]
- Chaussard, G.; Domard, A. New Aspects of the Extraction of Chitin from Squid Pens. Biomacromolecules 2014, 5, 559–564. [Google Scholar] [CrossRef]
- Fan, Y.; Saito, T.; Isogai, A. Preparation of Chitin Nanofibers from Squid Pen β-Chitin by Simple Mechanical Treatment under Acid Conditions. Biomacromolecules 2008, 9, 1919–1923. [Google Scholar] [CrossRef]
- Davies, G.; Knight, D.; Vollrath, F. Chitin in the Silk Gland Ducts of the Spider Nephila edulis and the Silkworm Bombyx mori. PLoS ONE 2013, 8, e73225. [Google Scholar] [CrossRef] [Green Version]
- John, C.C. The chemistry and Chemical Ecology of Octocorals (Coelenterata, Anthozoa, Octocorallia). Chem. Rev. 1992, 92, 613–631. [Google Scholar]
- Kaya, M.; Asan-Ozusaglam, M.; Erdogan, S. Comparison of antimicrobial activities of newly obtained low molecular weight scorpion chitosan and medium molecular weight commercial chitosan. J. Biosci. Bioeng. 2016, 121, 678–684. [Google Scholar] [CrossRef]
- Zhang, H.; Neau, S. In vitro degradation of chitosan by a commercial enzyme preparation: Effect of molecular weight and degree of deacetylation. Biomaterials 2001, 22, 1653–1658. [Google Scholar] [CrossRef]
- Liu, S.; Sun, J.; Yu, L.; Zhang, C.; Bi, J.; Zhu, F.; Qu, M.; Jiang, C.; Yang, Q. Extraction and Characterization of Chitin from the Beetle Holotrichia parallela Motschulsky. Molecules 2012, 17, 4604–4611. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Haga, A.; Sekiguchi, H.; Hirano, S. Structure of insect chitin isolated from beetle larva cuticle and silkworm (Bombyx mori) pupa exuvia. Int. J. Biol. Macromol. 2000, 27, 99–105. [Google Scholar] [CrossRef]
- Kaya, M.; Baran, T. Description of a new surface morphology for chitin extracted from wings of cockroach (Periplaneta americana). Int. J. Biol. Macromol. 2015, 75, 7–12. [Google Scholar] [CrossRef]
- Tanaka, K.; Katsura, N.; Saku, T.; Kasuga, S. Composite Texture of Chitin and Keratin in an Animal Organ, Lingula seta. Polym. J. 2015, 20, 119–123. [Google Scholar] [CrossRef] [Green Version]
- Synowiecki, J.; Al-Khateeb, N. Mycelia of Mucor rouxii as a source of chitin and chitosan. Food Chem. 1997, 60, 605–610. [Google Scholar] [CrossRef]
- Mathur, N.; Narang, C. Chitin and chitosan, versatile polysaccharides from marine animals. J. Chem. Educ. 1990, 67, 938. [Google Scholar] [CrossRef]
- Chobot, V.; Kremenák, J.; Opletal, L. Phytotherapeutic aspects of diseases of the circulatory system. 4. Chitin and chitosan. Ceska Slov. Farm. 1995, 44, 190–195. [Google Scholar]
- Kulikov, S.N.; Chirkov, S.N.; Il’Ina, A.V.; Lopatin, S.A.; Varlamov, V.P. Effect of the molecular weight of chitosan on its antiviral activity in plants. Appl. Biochem. Microbiol. 2006, 42, 200–203. [Google Scholar] [CrossRef]
- De Alvarenga, E.S.; de Oliveira, C.P.; Bellato, C.R. An approach to understanding the deacetylation degree of chitosan. Carbohydr. Polym. 2010, 80, 1155–1160. [Google Scholar] [CrossRef]
- Jaworska, M.; Sakurai, K.; Gaudon, P.; Guibal, E. Influence of chitosan characteristics on polymer properties. I: Crystallographic properties. Polym. Int. 2003, 52, 198–205. [Google Scholar] [CrossRef]
- Dotto, G.; Campana-Filho, S.; de Almeida Pinto, L.A. Frontiers in Biomaterials; Bentham Science Publishers: Bussum, The Netherlands, 2017. [Google Scholar]
- Verlee, A.; Mincke, S.; Stevens, C. Recent developments in antibacterial and antifungal chitosan and its derivatives. Carbohydr. Polym. 2017, 164, 268–283. [Google Scholar] [CrossRef] [PubMed]
- Minh, N.; van Hoa, N.; Trung, T. Preparation, properties, and application of low-molecular-weight chitosan. In Handbook of Chitin and Chitosan; Elsevier: Amsterdam, The Netherlands, 2020; Volume 1, pp. 453–471. [Google Scholar]
- Niazi, S. Production, Classification, Properties and Application of Chitosan. Int. J. Res. Agric. Sci. 2016, 3, 2358–3997. [Google Scholar]
- Zargar, V.; Asghari, M.; Dashti, A. A review on chitin and chitosan polymers: Structure, chemistry, solubility, derivatives, applications. ChemBioEng 2015, 2, 204–226. [Google Scholar] [CrossRef]
- Tishchenko, G.; Šimůnek, J.; Brus, J.; Netopilík, M.; Pekárek, M.; Walterová, Z.; Koppová, I.; Lenfeld, J. Low-molecular-weight chitosans: Preparation and characterization. Carbohydr. Polym. 2011, 86, 1077–1081. [Google Scholar] [CrossRef]
- Qin, C.; Zhou, B.; Zeng, L.; Zhang, Z.; Liu, Y.; Du, Y.; Xiao, L. The physicochemical properties and antitumor activity of cellulase-treated chitosan. Food Chem. 2004, 84, 107–115. [Google Scholar] [CrossRef]
- Thakur, V.; Thakur, M. Recent Advances in Graft Copolymerization and Applications of Chitosan: A Review. ACS Sustain. Chem. Eng. 2014, 2, 2637–2652. [Google Scholar] [CrossRef]
- Li, K.; Xing, R.; Liu, S.; Li, P. Advances in preparation, analysis and biological activities of single chitooligosaccharides. Carbohydr. Polym. 2016, 139, 178–190. [Google Scholar] [CrossRef]
- Lin, S.; Lin, Y.; Chen, H. Low molecular weight chitosan prepared with the aid of cellulase, lysozyme and chitinase: Characterisation and antibacterial activity. Food Chem. 2009, 1161, 47–53. [Google Scholar] [CrossRef]
- Kim, K.; Thomas, R. Antioxidative activity of chitosans with varying molecular weights. Food Chem. 2007, 101, 308–313. [Google Scholar] [CrossRef]
- Jae, Y.; Park, P.; Kim, S. Free radical scavenging properties of hetero-chitooligosaccharides using an ESR spectroscopy. Food Chem. Toxicol. 2004, 42, 381–387. [Google Scholar] [CrossRef]
- Gómez, B.; Duncan, R. Evaluation of the biological properties of soluble chitosan and chitosan microspheres. Int. J. Pharm. 1997, 148, 231–240. [Google Scholar] [CrossRef]
- Bof, M.; Bordagaray, V.; Locaso, D.; García, M. Chitosan molecular weight effect on starch-composite film properties. Food Hydrocoll. 2015, 51, 281–294. [Google Scholar] [CrossRef]
- Román-Doval, R.; Gómez-Sánchez, A.; Millán-Casarrubias, E.J.; Prokhorov, E.; Montejo-Alvaro, F.; de Luna Bugallo, A.; Mendoza, S. Physicochemical properties of pullulan/chitosan/graphene oxide composite films. Polym. Int. 2022, 71, 959–965. [Google Scholar] [CrossRef]
- Strupiechonski, E.; Moreno-Ríos, M.; Ávila-Dávila, E.O.; Román-Doval, R.; Prokhorov, E.; Kovalenko, Y.; Luna-Barcenas, G. Relaxation Phenomena in Chitosan-Au Nanoparticle Thin Films. Polymers 2021, 13, 3214. [Google Scholar] [CrossRef]
- Tokatli, K.; Demirdöven, A. Influences of chitosan coatings on functional compounds of sweet cherries. J. Food Sci. Technol. 2021, 58, 1808–1818. [Google Scholar] [CrossRef]
- Dotto, G.; Vieira, M.; Pinto, L. Use of chitosan solutions for the microbiological shelf life extension of papaya fruits during storage at room temperature. LWT-Food Sci. Technol. 2015, 64, 126–130. [Google Scholar] [CrossRef]
- Kumari, S.; Kishor, R. Chitin and chitosan: Origin, properties, and applications. In Handbook of Chitin and Chitosan; Elsevier: Amsterdam, The Netherlands, 2020; Volume 1, pp. 1–33. [Google Scholar]
- Wang, W.; Xue, C.; Mao, X. Chitosan: Structural modification, biological activity and application. Int. J. Biol. Macromol. 2020, 164, 4532–4546. [Google Scholar] [CrossRef]
- Kou, S.; Peters, L.; Mucalo, M. Chitosan: A review of sources and preparation methods. Int. J. Biol. Macromol. 2021, 169, 85–94. [Google Scholar] [CrossRef]
- Arancibia, M.; López-Caballero, M.; Gómez-Guillén, M.; Fernández-García, M.; Fernández-Martín, F.; Montero, P. Antimi-crobial and rheological properties of chitosan as affected by extracting conditions and humidity exposure. LWT-Food Sci. Technol. 2015, 60, 802–810. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, C.; Maciel, V.; Mendonça, M.; Franco, T. Chitosan biobased and intelligent films: Monitoring pH variations. LWT-Food Sci. Technol. 2014, 55, 83–89. [Google Scholar] [CrossRef]
- Muthu, M.; Gopal, J.; Chun, S.; Devadoss, A.J.P.; Hasan, N.; Sivanesan, I. Crustacean Waste-Derived Chitosan: Antioxidant Properties and Future Perspective. Antioxidants 2021, 10, 228. [Google Scholar] [CrossRef] [PubMed]
- Garcia, L.; Guedes, G.; da Silva, M.; Castelo-Branco, D.; Sidrim, J.; de Aguiar Cordeiro, R.; Rocha, M.; Vieira, R.; Brilhante, R. Effect of the molecular weight of chitosan on its antifungal activity against Candida spp. in planktonic cells and biofilm. Carbohydr. Polym. 2018, 195, 662–669. [Google Scholar] [CrossRef] [PubMed]
- Hosseinnejad, M.; Jafari, S. Evaluation of different factors affecting antimicrobial properties of chitosan. Int. J. Biol. Macromol. 2016, 85, 467–475. [Google Scholar] [CrossRef] [PubMed]
- Xing, K.; Zhu, X.; Peng, X.; Qin, S. Chitosan antimicrobial and eliciting properties for pest control in agriculture: A review. Agron. Sustain. Dev. 2015, 35, 569–588. [Google Scholar] [CrossRef] [Green Version]
- Rkhaila, A.; Chtouki, T.; Erguig, H.; el Haloui, N.; Ounine, K. Chemical Properties of Biopolymers (Chitin/Chitosan) and Their Synergic Effects with Endophytic Bacillus Species: Unlimited Applications in Agriculture. Molecules 2021, 26, 1117. [Google Scholar] [CrossRef]
- Torres, J.; Reyes, J.; Castellanos, T.; Angulo, C.; Quiñones, E.; Hernandez, L. A biopolymer with antimicrobial properties and plant resistance inducer against phytopathogens: Chitosan. Not. Bot. Horti Agrobot.-Cluj-Napoca 2021, 49, 12231. [Google Scholar] [CrossRef]
- Mansilla, A.; Albertengo, L.; Rodríguez, M.; Debbaudt, A.; Zúñiga, A.; Casalongué, C. Evidence on antimicrobial properties and mode of action of a chitosan obtained from crustacean exoskeletons on Pseudomonas syringae pv. tomato DC3000. Appl. Microbiol. Biotechnol. 2013, 97, 6957–6966. [Google Scholar] [CrossRef]
- Badawy, M.; Rabea, E.; Taktak, N. Antimicrobial and inhibitory enzyme activity of N-(benzyl) and quaternary N-(benzyl) chitosan derivatives on plant pathogens. Carbohydr. Polym. 2014, 11, 670–682. [Google Scholar] [CrossRef]
- Orzali, L.; Corsi, B.; Forni, C.; Riccioni, L. Chitosan in Agriculture: A New Challenge for Managing Plant Disease. In Biological Activities and Application of Marine Polysaccharides; InTech: London, UK, 2017. [Google Scholar]
- Rinaudo, M. Chitin and chitosan: Properties and applications. Prog. Polym. Sci. 2006, 31, 603–632. [Google Scholar] [CrossRef]
- Sharma, S.; Barman, K.; Siddiqui, M. Chitosan: Properties and roles in postharvest quality preservation of horticultural crops. In Eco-Friendly Technology for Postharvest Produce Quality; Academic Press: Cambridge, MA, USA, 2016; pp. 269–296. [Google Scholar]
- Abdellatef, M.A.; Elagamey, E.; Kamel, S.M. Chitosan Is the Ideal Resource for Plant Disease Management under Sustainable Agriculture. In Chitin-Chitosan—Isolation, Properties, and Applications; InTech Open: London, UK, 2022. [Google Scholar]
- Badawy, M. Structure and antimicrobial activity relationship of quaternary N-alkyl chitosan derivatives against some plant pathogens. J. Appl. Polym. Sci. 2010, 117, 960–969. [Google Scholar] [CrossRef]
- Ortega, H.; Gutiérrez, B.; Cadenas, G.; Jimenez, L. Antibacterial activity of chitosan and the interpolyelectrolyte complexes of poly(acrylic acid)-chitosan. Braz. Arch. Biol. Technol. 2010, 53, 623–628. [Google Scholar] [CrossRef] [Green Version]
- Marei, G.; Rabea, E.; Badawy, M. Preparation and Characterizations of Chitosan/Citral Nanoemulsions and their Antimi-crobial Activity. Appl. Food Biotechnol. 2018, 5, 69–78. [Google Scholar]
- Li, L.; Deng, J.; Deng, H.; Liu, Z.; Li, X. Preparation, characterization and antimicrobial activities of chitosan/Ag/ZnO blend films. Chem. Eng. J. 2010, 160, 378–382. [Google Scholar] [CrossRef]
- Hemantaranjan, A. A Future Perspective in Crop Protection: Chitosan and its Oligosaccharides. Adv. Plants Agric. Res. 2014, 1, 23–30. [Google Scholar] [CrossRef] [Green Version]
- Coqueiro, D.; di Piero, R. Antibiotic Activity against Xanthomonas Gardneri and Protection of Tomato Plants by Chitosan. J. Plant Pathol. 2011, 93, 337–344. [Google Scholar]
- Gonelimali, F.D.; Jiheng, L.; Wenhua, M.; Jinghu, X.; Fedrick, C.; Meiling, C.; Shaimaa, R. Antimicrbial Properties and Mechanism of Action of Some Plant Extracts against Food Pathogens and Spoilage Microorganisms. Front. Microbiol. 2018, 9, 1639. [Google Scholar] [CrossRef]
- Li, J.; Wu, Y.; Zhao, L. Antibacterial activity and mechanism of chitosan with ultra high molecular weight. Carbohydr. Polym. 2016, 148, 200–205. [Google Scholar] [CrossRef]
- Hernández, A.; Bautista, S.; Velázquez, M.; Méndez, M.; Sánchez, M.; Bello, L. Antifungal effects of chitosan with different molecular weights on in vitro development of Rhizopus stolonifer (Ehrenb.:Fr.) Vuill. Carbohydr. Polym. 2008, 73, 541–547. [Google Scholar] [CrossRef]
- Oliul, H.; Taehyun, C. Chitosan for Eco-friendly Control of Plant Disease. Asian J. Plant Pathol. 2017, 11, 53–70. [Google Scholar]
- Muñoz, A.; Cerrada, M.; Fernández, M. Antimicrobial Activity of Chitosan in Food. Agric. Biomed. 2013, 22–53. [Google Scholar]
- Faqir, Y.; Ma, J.; Chai, Y. Chitosan in modern agriculture production. Plant Soil Environ. 2021, 12, 667–679. [Google Scholar] [CrossRef]
- Chakraborty, M.; Mirza, H.; Rahman, M.; Khan, M.A.R.; Bhowmik, P.; Mahmud, N.U.; Tanveer, M.; Islam, T. Mechanism of Plant Growth Promotion and Disease Suprresion by Chitosan Biopolymer. Agriculture 2020, 10, 624. [Google Scholar] [CrossRef]
- Abd El-Aziz, M.; Khalil, M. Antiviral and Antinematodal potentials of chitosan: Review. J. Plant Sci. Phytopathol. 2020, 4, 55–59. [Google Scholar]
- Davydova, V.; Nagorskaya, V.; Gorbach, V.; Kalitnik, A.A.; Reunov, A.V.; Solov’Eva, T.F.; Ermak, I.M. Chitosan antiviral activity: Dependence on structure and depolymerization method. Appl. Biochem. Microbiol. 2011, 47, 103–108. [Google Scholar] [CrossRef]
- Gangireddygari, V.; Chung, B.; Cho, I.; Yoon, J. Inhibitory Effect of Chitosan and Phosphate Cross-linked Chitosan against Cucumber Mosaic Virus and Pepper Mild Mottle Virus. Plant Pathol. J. 2021, 37, 632–640. [Google Scholar] [CrossRef] [PubMed]
- El-Sayed, S.; Mahdy, M. Effect of chitosan on root-knot nematode, Meloidogyne javanica on tomato plants. Int. J. Chemtech Res. 2015, 7, 1985–1992. [Google Scholar]
- Nunes da Silva, M.; Cardoso, A.; Ferreira, D.; Brito, M.; Pintado, M.; Vasconcelos, M. Chitosan as a biocontrol agent against the pinewood nematode (Bursaphelenchus xylophilus). For. Pathol. 2014, 44, 420–443. [Google Scholar] [CrossRef]
- Iriti, M.; Varoni, E. Chitosan-induced antiviral activity and innate immunity in plants. Environ. Sci. Pollut. Res. 2015, 22, 2935–2944. [Google Scholar] [CrossRef]
- Badawy, M.; Rabea, E. A Biopolymer Chitosan and Its Derivatives as Promising Antimicrobial Agents against Plant Pathogens and Their Applications in Crop Protection. Hindawi Publ. Corp. Int. J. Carbohydr. Chem. 2011, 2011, 460381. [Google Scholar] [CrossRef]
- Meng, D.; Garba, B.; Ren, Y.; Yao, M.; Xia, X.; Li, M.; Wang, Y. Antifungal activity of chitosan against Aspergillus ochraceus and its possible mechanisms of action. Int. J. Biol. Macromol. 2020, 158, 1063–1070. [Google Scholar] [CrossRef]
- Feliziani, E.; Landi, L.; Romanazzi, G. Preharvest treatments with chitosan and other alternatives to conventional fungicides to control postharvest decay of strawberry. Carbohydr. Polym. 2015, 132, 111–117. [Google Scholar] [CrossRef]
- Reglinski, T.; Elmer, P.; Taylor, J.; Wood, P.N.; Hoyte, S. Inhibition of Botrytis cinerea growth and suppression of botrytis bunch rot in grapes using chitosan. Plant Pathol. 2010, 59, 882–890. [Google Scholar] [CrossRef]
- Zahid, N.; Maqbool, M.; Siddiqui, Y.; Manickam, S.; Ali, A. Regulation of inducible enzymes and suppression of anthracnose using submicron chitosan dispersions. Sci. Hortic. 2015, 193, 381–388. [Google Scholar] [CrossRef]
- Liu, H.; Tian, W.; Li, B.; Wu, G.; Ibrahim, M.; Tao, Z.; Wang, Y.; Xie, G.; Li, H.; Sun, G. Antifungal effect and mechanism of chitosan against the rice sheath blight pathogen, Rhizoctonia solani. Biotechnol. Lett. 2012, 34, 2291–2298. [Google Scholar] [CrossRef]
- Galván, I.; Akuaku, J.; Cruz, I.; Cheetham, J.; Golshani, A.; Smith, M. Disruption of protein synthesis as antifungal mode of action by chitosan. Int. J. Food Microbiol. 2013, 164, 108–112. [Google Scholar] [CrossRef]
- Ippólito, S.; Mendieta, J.; Terrile, M.; Tonón, C.; Mansilla, A.; Colman, S.; Albertengo, L.; Rodríguez, M.; Casalongué, C. Chitosan as Source for Pesticide Formulations. In Biological Activities and Application of Marine Polysaccharide; InTech: London, UK, 2017. [Google Scholar]
- Rahman, M.; Shovan, L.; Hjeljord, L.; Aam, B.; Eijsink, H.; Sørlie, M.; Tronsmo, A. Inhibition of Fungal Plant Pathogens by Synergistic Action of Chito-Oligosaccharides and Commercially Available Fungicides. PLoS ONE 2014, 9, e93192. [Google Scholar] [CrossRef]
- El Guilli, M.; Hamza, A.; Clément, C.; Ibriz, M.; Ait Barka, E. Effectiveness of Postharvest Treatment with Chitosan to Control Citrus Green Mold. Agriculture 2016, 6, 12. [Google Scholar] [CrossRef] [Green Version]
- Hernández, A. Current status of action mode and effect of chitosan against phytopathogens fungi. Afr. J. Microbiol. Res. 2011, 5, 4243–4247. [Google Scholar]
- Meng, X.; Yang, L.; Kennedy, J.; Tian, S. Effects of chitosan and oligochitosan on growth of two fungal pathogens and physiological properties in pear fruit. Carbohydr. Polym. 2010, 81, 70–75. [Google Scholar] [CrossRef]
- Hadwiger, L. Multiple effects of chitosan on plant systems: Solid science or hype. Plant Sci. 2013, 208, 42–49. [Google Scholar] [CrossRef]
- Nguyen, V.; Tran, T. Application of chitosan solutions for rice production in Vietnam. Afr. J. Biotechnol. 2013, 12, 382–384. [Google Scholar] [CrossRef] [Green Version]
- Hidangmayum, A.; Dwivedi, P.; Katiyar, D.; Hemantaranjan, A. Application of chitosan on plant responses with special reference to abiotic stress. Physiol. Mol. Biol. Plants 2019, 25, 313–326. [Google Scholar] [CrossRef] [PubMed]
- Lizárraga, E.; Pacheco, I.; Miranda, S. Chitosan application in Maize (Zea mays) to counteract the effects of abiotic stress at seedling level. Afr. J. Biotechnol. 2011, 10, 6349–6446. [Google Scholar]
- Lemke, P.; Jünemann, L.; Moerschbacher, B.M. Synergistic Antimicrobial Activities of Chitosan Mixtures and Chitosan–Copper Combinations. Int. J. Mol. Sci. 2022, 23, 3345. [Google Scholar] [CrossRef] [PubMed]
- Umoren, P.S.; Kavaz, D.; Nzila, A.; Sankaran, S.S.; Umoren, S.A. Biogenic Synthesis and Characterization of Chitosan-CuO Nanocomposite and Evaluation of Antibacterial Activity against Gram-Positive and-Negative Bacteria. Polymers 2022, 14, 1832. [Google Scholar] [CrossRef]
- Mahdavi, B.; Rahimi, A. Seed priming with chitosan improves the germination and growth performance of ajowan (Carum copticum) under salt stress. EurAsian J. BioSci. 2013, 7, 69–76. [Google Scholar] [CrossRef]
- Razavizadeh, R.; Adabavazeh, F.; Komatsu, S. Chitosan effects on the elevation of essential oils and antioxidant activity of Carum copticum L. seedlings and callus cultures under in vitro salt stress. J. Plant Biochem. Biotechnol. 2020, 29, 473–483. [Google Scholar] [CrossRef]
- Golkar, P.; Taghizadeh, M.; Yousefian, Z. The effects of chitosan and salicylic acid on elicitation of secondary metabolites and antioxidant activity of safflower under in vitro salinity stress. Plant Cell Tiss. Organ Cult. 2019, 137, 575–585. [Google Scholar] [CrossRef]
- Zong, H.; Li, K.; Liu, S.; Song, L.; Xing, R.; Chen, X.; Li, P. Improvement in cadmium tolerance of edible rape (Brassica rapa L.) with exogenous application of chitooligosaccharide. Chemosphere 2017, 181, 92–100. [Google Scholar] [CrossRef]
- Escudero, N.; Lopez, F.; Ghahremani, Z.; Zavala, E.; Alaguero, A.; Ros, C.; Lacasa, A.; Sorribas, F.; Lopez, L. Chitosan Increases Tomato Root Colonization by Pochonia chlamydosporia and Their Combination Reduces Root-Knot Nematode Damage. Front. Plant Sci. 2017, 8, 1415. [Google Scholar] [CrossRef] [Green Version]
- Guo, Z.; Xing, R.; Liu, S.; Zhong, Z.; Ji, X.; Wang, L.; Li, P. The influence of molecular weight of quaternized chitosan on antifungal activity. Carbohydr. Polym. 2008, 71, 694–697. [Google Scholar] [CrossRef]
- No, H. Antibacterial activity of chitosans and chitosan oligomers with different molecular weights. Int. J. Food Microbiol. 2002, 74, 65–72. [Google Scholar] [CrossRef]
- Vishu, B.; Varadaraj, M.; Tharanathan, R. Low Molecular Weight ChitosanPreparation with the Aid of Pepsin, Characteri-zation, and Its Bactericidal Activity. Biomacromolecule 2007, 8, 566–572. [Google Scholar] [CrossRef]
- Younes, I.; Sellimi, S.; Rinaudo, M.; Jellouli, K.; Nasri, M. Influence of acetylation degree and molecular weight of homoge-neous chitosans on antibacterial and antifungal activities. Int. J. Food Microbiol. 2014, 185, 57–63. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, S.; Sun, M.; Su, H.; Li, H.; Zhang, Y.; Chen, X.; Cao, Y.; Song, Y. Antibacterial activity of peptaibols from Trichoderma longibrachiatum SMF2 against gram-negative Xanthomonas oryzae pv. oryzae, the causal agent of bacterial leaf blight on rice. Front. Microbiol. 2022, 13, 1034779. [Google Scholar] [CrossRef]
- Lopez, F.; Lopez, L. Omics for Investigating Chitosan as an Antifungal and Gene Modulator. J. Fungi 2016, 2, 11. [Google Scholar] [CrossRef] [Green Version]
- Gong, B.; Wang, F.; Li, J. Hide-and-Seek: Chitin-Triggered Plant Immunity and Fungal Counterstrategies. Trends Plant Sci. 2020, 25, 805–816. [Google Scholar] [CrossRef]
- Guerra, M.; Vega, J.; Velázquez, M.; Hernández, A. Antifungal activity and release of compounds on Rhizopus stolonifer (Ehrenb.:Fr.) Vuill. by effect of chitosan with different molecular weights. Pestic. Biochem. Physiol. 2009, 93, 18–22. [Google Scholar] [CrossRef]
- Xing, K.; Li, T.; Liu, Y.; Zhang, J.; Zhang, Y.; Shen, X.; Li, X.; Miao, X.; Feng, Z.; Peng, X.; et al. Antifungal and eliciting properties of chitosan against Ceratocystis fimbriata in sweet potato. Food Chem. 2018, 268, 188–195. [Google Scholar] [CrossRef]
- Khayrova, A.; Lopatin, S.; Shagdarova, B.; Sinitsyna, O.; Sinitsyn, A.; Varlamov, V. Evaluation of Antibacterial and Antifungal Properties of Low Molecular Weight Chitosan Extracted from Hermetia illucens Relative to Crab Chitosan. Molecules 2022, 27, 577. [Google Scholar] [CrossRef]
- Wang, Q.; Zuo, J.; Wang, Q.; Na, Y.; Gao, L. Inhibitory effect of chitosan on growth of the fungal phytopathogen, Sclerotinia sclerotiorum, and sclerotinia rot of carrot. J. Integr. Agric. 2015, 14, 691–697. [Google Scholar] [CrossRef] [Green Version]
- Mejdoub, B.; Touihri, S.; Ammar, N.; Riahi, A.; Daami-Remadi, M. Effect of chitosan for the control of potato diseases caused by Fusarium species. J. Phytopathol. 2020, 168, 18–27. [Google Scholar] [CrossRef]
- Wang, Q.; Li, H.; Lei, Y.; Su, Y.; Long, Y. Chitosan as an Adjuvant to Improve Isopyrazam Azoxystrobin against Leaf Spot Disease of Kiwifruit and Enhance Its Photosynthesis, Quality, and Amino Acids. Agriculture 2022, 12, 373. [Google Scholar] [CrossRef]
- Wang, L.; Wu, H.; Qin, G.; Meng, X. Chitosan disrupts Penicillium expansum and controls postharvest blue mold of jujube fruit. Food Control. 2014, 41, 56–62. [Google Scholar] [CrossRef]
- Ruiz-García, Y.; Gómez-Plaza, E. Elicitors: A Tool for Improving Fruit Phenolic Content. Agriculture 2013, 3, 33–52. [Google Scholar] [CrossRef] [Green Version]
- Fan, Z.; Qin, Y.; Liu, S.; Xing, R.; Yu, H.; Li, P. Chitosan Oligosaccharide Fluorinated Derivative Control Root-Knot Nematode (Meloidogyne incognita) Disease Based on the Multi-Efficacy Strategy. Mar. Drugs 2020, 18, 273. [Google Scholar] [CrossRef]
- Guizhi, F.; Xiaocan, L.; Xiaodong, W.; Qiaoli, Z.; Yaguang, Z. Chitosan activates defense responses and triterpenoid production in cell suspension cultures of Betula platyphylla Suk. Afr. J. Biotechnol. 2010, 9, 2816–2820. [Google Scholar]
- Zong, H.; Liu, S.; Xing, R.; Chen, X.; Li, P. Protective effect of chitosan on photosynthesis and antioxidative defense system in edible rape (Brassica rapa L.) in the presence of cadmium. Ecotoxicol. Environ. Saf. 2017, 138, 271–278. [Google Scholar] [CrossRef]
- Stasińska, M.; Barbara, H. Protective, Biostimulating, and Eliciting Effects of Chitosan and Its Derivatives on Crop Plants. Molecules 2022, 27, 2801. [Google Scholar] [CrossRef]
- Mejía, L.; Durán, F.; Chapa, A.; Torres, I.; Cruz, A.; González, M.; Ocampo, R.; Guevara, R. Oxidative and Molecular Responses in Capsicum annuum L. after Hydrogen Peroxide, Salicylic Acid and Chitosan Foliar Applications. Int. J. Mol. Sci. 2013, 14, 10178–10196. [Google Scholar] [CrossRef] [Green Version]
- Rendina, N.; Nuzzaci, M.; Scopa, A.; Cuypers, A.; Sofo, A. Chitosan-elicited defense responses in Cucumber mosaic virus (CMV)-infected tomato plants. J. Plant Physiol. 2019, 234, 9–17. [Google Scholar] [CrossRef]
- Liu, J.; Zhang, X.; Kennedy, J.; Jiang, M.; Cai, Q.; Wu, X. Chitosan induces resistance to tuber rot in stored potato caused by Alternaria tenuissima. Int. J. Biol. Macromol. 2019, 140, 851–857. [Google Scholar] [CrossRef]
- Zheng, F.; Zheng, W.; Li, L.; Pan, S.; Liu, M.; Zhang, W.; Liu, H.; Zhu, C. Chitosan Controls Postharvest Decay and Elicits Defense Response in Kiwifruit. Food Bioprocess Technol. 2017, 10, 1937–1945. [Google Scholar] [CrossRef]
- El Amerany, F.; Rhazi, M.; Balcke, G.; Wahbi, S.; Meddich, A.; Taourirte, M.; Hause, B. The Effect of Chitosan on Plant Physiology, Wound Response, and Fruit Quality of Tomato. Polymers 2022, 14, 5006. [Google Scholar] [CrossRef]
- Khalil, M.; Badawy, M. Nematicidal activity of a biopolymer chitosan at different molecular weights against root-knot nematode, Meloidogyne incognita. Plant Protect. Sci. 2012, 48, 170–178. [Google Scholar] [CrossRef] [Green Version]
- Radwan, M.; Farrag, S.; Abu, M.; Ahmed, N. Extraction, characterization, and nematicidal activity of chitin and chitosan derived from shrimp shell wastes. Biol. Fertil. Soils 2012, 48, 463–468. [Google Scholar] [CrossRef]
- Pongprayoon, W.; Siringam, T.; Panya, A.; Roytrakul, S. Application of Chitosan in Plant Defense Responses to Biotic and Abiotic Stresses. Appl. Sci. Eng. Prog. 2020, 15, 3865. [Google Scholar] [CrossRef]
- Darwis, D.; Puspitasari, T.; Iramani, D.; Susilowati, S.; Pangerteni, D. Preparation of Low Molecular Weight Chitosan by Radiation and its Application for Plant Growth Promoter Chapter 10. In Proceedings of the Technical Meeting on Radiation Processed Materials in Products from Polymers for Agricultural Applications, Vienna, Austria, 8–12 July 2013. [Google Scholar]
- Falcón, A.; Costales, D.; González, D.; Morales, D.; Mederos, Y.; Jerez, E.; Cabrera, J. Chitosans of different molecular weight enhance potato (Solanum tuberosum L.) yield in a field trial. Span. J. Agric. Res. 2017, 15, 25. [Google Scholar]
- Kumar, V.; Kirubanandam, S.; Soundararajan, A.; Sudha, P. Chitin and Chitosan—The Defence Booster in Agricultural Field. In Handbook of Biopolymers: Advances and Multifaceted Application; CRC Press: Boca Raton, FL, USA, 2018; pp. 93–134. [Google Scholar]
- Chamnanmanoontham, N.; Pongprayoon, W.; Pichayangkura, R.; Roytrakul, S.; Chadchawan, S. Chitosan enhances rice seedling growth via gene expression network between nucleus and chloroplast. Plant Growth Regul. 2015, 75, 101–114. [Google Scholar] [CrossRef]
- Malerba, M.; Cerana, R. Chitosan Effects on Plant Systems. Int. J. Mol. Sci. 2016, 17, 996. [Google Scholar] [CrossRef]
- Quitadamo, F.; de Simone, V.; Beleggia, R.; Trono, D. Chitosan-Induced Activation of the Antioxidant Defense System Counteracts the Adverse Effects of Salinity in Durum Wheat. Plants 2021, 10, 1365. [Google Scholar] [CrossRef] [PubMed]
- Attia, M.S.; Osman, M.S.; Mohamed, A.S.; Mahgoub, H.A.; Garada, M.O.; Abdelmouty, E.S.; Abdel Latef, A.A.H. Impact of Foliar Application of Chitosan Dissolved in Different Organic Acids on Isozymes, Protein Patterns and Physio-Biochemical Characteristics of Tomato Grown under Salinity Stress. Plants 2021, 10, 388. [Google Scholar] [CrossRef] [PubMed]
- Chanratana, M.; Han, G.; Melvin, M.; Roy, A.; Sundaram, S.; Halim, M.; Sa, T. Evaluation of chitosan and alginate immobilized Methylobacterium oryzae CBMB20 on tomato plant growth. Arch. Agron. Soil Sci. 2018, 64, 1489–1502. [Google Scholar] [CrossRef]
- Salachna, P.; Zawadzińska, A. Effect of chitosan on plant growth, flowering and corms yield of potted freesia. J. Ecol. Eng. 2014, 15, 97–102. [Google Scholar]
- Chookhongkha, N.; Miyagawa, S.; Jirakiattikul, Y.; Photchanachai, S. Chilli growth and seed productivity as affected by chitosan. Int. Conf. Agric. Technol. Food Sci. 2012, 17–18, 146–149. [Google Scholar]
- Krupa, M.; Fornal, N. Application of Chitosan in vitro to Minimize the Adverse Effects of Salinity in Petunia × atkinsiana D. don. J. Ecol. Eng. 2018, 19, 143–149. [Google Scholar] [CrossRef]
- Safikhan, S.; Khoshbakht, K.; Chaichi, M.; Amini, A.; Motesharezadeh, B. Role of chitosan on the growth, physiological parameters and enzymatic activity of milk thistle (Silybum marianum (L.) Gaertn.) in a pot experiment. J. Appl. Res. Med. Aromat. Plants 2018, 10, 49–58. [Google Scholar] [CrossRef]
- Kweon, K.; Kang, D.W. Drug-release behavior of chitosan-g-poly(vinyl alcohol) copolymer matrix. J. Appl. Polym. Sci. 1999, 74, 458–464. [Google Scholar] [CrossRef]
- Zhang, M.; Li, X.H.; Gong, Y.D.; Zhao, N.M.; Zhang, X.F. Properties and biocompatibility of chitosan films modified by blending with PEG. Biomaterials 2002, 23, 2641. [Google Scholar] [CrossRef]
- Wang, H.; Qian, J.; Ding, F. Recent advances in engineered chitosan-based nanogels for biomedical applications. J. Mater. Chem. B 2017, 5, 6986–7007. [Google Scholar] [CrossRef]
- Wang, H.; Qian, J.; Ding, F. Emerging Chitosan-Based Films for Food Packaging Applications. J. Agric. Food Chem. 2018, 66, 395–413. [Google Scholar] [CrossRef]
- Elhefian, E. Chitosan-Based Polymer Blends: Current Status and Applications. Chem. Soc. Pak. 2014, 36, 11–27. [Google Scholar]
- Qu, B.; Luo, Y. Chitosan-based hydrogel beads: Preparations, modifications and applications in food and agriculture sectors—A review. Int. J. Biol. Macromol. 2020, 152, 437–448. [Google Scholar] [CrossRef]
- Saharan, V.; Pal, A. Chitosan Based Nanomaterials in Plant Growth and Protection; Springer: New Delhi, India, 2016; pp. 33–41. [Google Scholar]
- Mukarram, M.; Khan, M.M.A.; Choudhary, S.; Zehra, A.; Naeem, M.; Aftab, T. Natural polysaccharides: Novel plant growth regulators. In Plant Growth Regulators: Signalling under Stress Condition; Springer: Berlin/Heidelberg, Germany, 2021; pp. 335–354. [Google Scholar]
- Jing, W.; Yin, L.; Lin, X.; Yu, Y.; Lian, D.; Shi, Z.; Chen, P.; Tang, M.; Yang, C. Simultaneous Adsorption of Cu2+ and Cd2+ by a Simple Synthesis of Environmentally Friendly Bamboo Pulp Aerogels: Adsorption Properties and Mechanisms. Polymers 2022, 14, 4909. [Google Scholar] [CrossRef]
- Gonçalves, J.; Esquerdo, V.; Sant’Anna, T.; de Almeida Pinto, L. Chitosan-Based Hydrogels. In Sustainable Agriculture Reviews; Springer International Publishing: Berlin/Heidelberg, Germany, 2019; pp. 147–173. [Google Scholar]
- Kundu, R.; Payal, P. Antimicrobial Hydrogels: Promising Soft Biomaterials. ChemistrySelect 2020, 5, 14800–14810. [Google Scholar] [CrossRef]
- Yu, J.; Wang, D.; Geetha, N.; Khawar, K.; Jogaiah, S.; Mujtaba, M. Current trends and challenges in the synthesis and applications of chitosan-based nanocomposites for plants: A review. Carbohydr. Polym. 2021, 261, 117904. [Google Scholar] [CrossRef]
- Cruz-Luna, A.R.; Cruz-Martínez, H.; Vásquez-López, A.; Medina, D.I. Metal Nanoparticles as Novel Antifungal Agents for Sustainable Agriculture: Current Advances and Future Directions. J. Fungi 2021, 7, 1033. [Google Scholar] [CrossRef]
- Nesic, K.; Ivanovic, S.; Nesic, V. Fusarial toxins: Secondary metabolites of Fusarium fungi. Reviews of Environmental Contamination and Toxicology. Rev. Environ. Contam. Toxicol. 2014, 228, 101–120. [Google Scholar]
- Qin, C.; Li, H.; Xiao, Q.; Liu, Y.; Zhu, J.; Du, Y. Water-solubility of chitosan and its antimicrobial activity. Carbohydr. Polym. 2006, 63, 367–374. [Google Scholar] [CrossRef]
- Qin, Y.; Xing, R.; Liu, S.; Li, K.; Meng, X.; Li, R.; Cui, J.; Li, B.; Li, P. Novel thiosemicarbazone chitosan derivatives: Preparation, characterization, and antifungal activity. Carbohydr. Polym. 2012, 87, 2664–2670. [Google Scholar] [CrossRef]
- Sahab, A.; Waly, A.; Sabbour, M.; Nawar, L.S. Synthesis, antifungal and insecticidal potential of Chitosan (CS)-g-poly (acrylic acid) (PAA) nanoparticles against some seed borne fungi and insects of soybean. Int. J. Chemtech Res. 2015, 8, 589–598. [Google Scholar]
- Santo Pereira, A.E.; Silva, P.M.; Oliveira, J.L.; Oliveira, H.C.; Fraceto, L.F. Chitosan nanoparticles as carrier systems for the plant growth hormone gibberellic acid. Colloids Surfaces B Biointerfaces 2017, 150, 141–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sashiwa, H.; Aiba, S. Chemically modified chitin and chitosan as biomaterials. Prog. Polym. Sci. 2004, 29, 887–908. [Google Scholar]
- Sathiyabama, M.; Parthasarathy, R. Biological preparation of chitosan nanoparticles and its in vitro antifungal efficacy against some phytopathogenic fungi. Carbohydr. Polym. 2016, 151, 321–325. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Marine |
|---|
| Sellfish |
| Crab |
| Chionoecetes opilio [21] |
| Podophthalmus vigil [22] |
| Paralithodes camtschaticus [23] |
| Carcinus mediterraneus [24] |
| Water lobster |
| Crawfish [25] |
| Shrimp |
| Aristens antennatus [26] |
| Krill |
| Daphnia longispina [27] |
| Anax imperator [28] |
| Hyrophilus piceus [28] |
| Notonecta glauca [28] |
| Agabus bipustulatus [28] |
| Asellus aquaticus [28] |
| Molluscs |
| Squid pens |
| Loligo sp. [29] |
| Todarodes pacificus [30] |
| Terrestrial |
| Arthropods |
| Spyders |
| Geolycosa vultuosa [31] |
| Nephila edulis [31] |
| Scorpions |
| Mesobuthus gibbosus [32] |
| Beetles |
| Bombyx mori [33] |
| Holotrichia parallela [34] |
| Leptinotarsa decemlineata [35] |
| Cockroaches [36] |
| Brachiopods |
| Lingula seta [37] |
| Microorganisms |
| Fungus (cell walls) |
| Ascomydes |
| Mucor rouxii [38] |
| Blastomycota |
| Blastocladiaceae [39] |
| Chytridiomycota |
| Chytridiaceae |
| Protista |
| Brown algae [40] |
| Plantae |
| Green algae [40] |
| Chitosan | |||
|---|---|---|---|
| Properties | Low Molecular Weight (>150 kDa) | Medium Molecular Weight (150–700 kDa) | High Molecular Weight (>700 kDa) |
| Physicochemical | High nitrogen content | ||
| Weak base with deprotonated amino groups as nucleophiles (pKa = 6.3) | Medium nitrogen content | Low nitrogen content | |
| Great reactive groups for crosslinking and chemical activation | Limited solubility | Limited solubility | |
| Has chelating and complexing properties | Pseudoplastic behavior | Pseudoplastic behavior | |
| Ionic conductivity as polyelectrolytes (pH < 7) | Good reactive groups for crosslinking and chemical activation | Low reactive groups for crosslinking and chemical activation | |
| Great solubility | High tensile strength | Low tensile strength | |
| Low viscosity | Medium viscosity | High viscosity | |
| High permeability | Medium permeability | Low permeability | |
| Low melting point | |||
| High tensile strength | |||
| Biological | Biocompatible | ||
| Nontoxic | |||
| Biodegradable | |||
| Antimicrobial (fungi, bacteria, viruses) | Biocompatible | Biocompatible | |
| Mucoadhesive | Nontoxic | Nontoxic | |
| Antioxidant | Antimicrobial (fungi, bacteria, viruses) | Biodegradable | |
| Antitumor | Higher antioxidant | Antimicrobial (fungi, bacteria, viruses) | |
| Hypolipidemic | Lower antioxidant | ||
| Blood anticoagulants | |||
| Hypocholesteromic activities | |||
| Pathogen | Chitosan Molecular Weight | Chitosan Content in Acetic Acid | Research Strategy | Bacteriostatic Result | Reference |
|---|---|---|---|---|---|
| Botrytis cinerea | 97% of deacetylation degree and 7.6 × 10 | 0.5 and 1 mg/mL | Potato dextrose agar (PDA) | - | [121] |
| Escherichia coli | 82.33% of deacetylation degree and 1.21 × 10 | 1 mg/mL | Growth medium | 85% | [86] |
| Fusarium graminearum | 2.8 kDa | 1 mg/mL | Agarose culture | - | [122] |
| Magnaporthe grisea | 13.4 to 18.8% acetylation degree and average molecular weight of 10 kDa | 0.5 mg/mL | Agar (HiMedia) | 57% | [123] |
| Alternaria solani | 2 to 61% of deacetylation degree and average kDa of 42.5 to 135 | 25 mg/mL | PDA and Muller-Hinton agar | 24% | [124] |
| Xanthomonas oryzae | 607 kDa | 2 mg/mL | PDA | 76.47% | [125] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Román-Doval, R.; Torres-Arellanes, S.P.; Tenorio-Barajas, A.Y.; Gómez-Sánchez, A.; Valencia-Lazcano, A.A. Chitosan: Properties and Its Application in Agriculture in Context of Molecular Weight. Polymers 2023, 15, 2867. https://doi.org/10.3390/polym15132867
Román-Doval R, Torres-Arellanes SP, Tenorio-Barajas AY, Gómez-Sánchez A, Valencia-Lazcano AA. Chitosan: Properties and Its Application in Agriculture in Context of Molecular Weight. Polymers. 2023; 15(13):2867. https://doi.org/10.3390/polym15132867
Chicago/Turabian StyleRomán-Doval, Ramón, Sandra P. Torres-Arellanes, Aldo Y. Tenorio-Barajas, Alejandro Gómez-Sánchez, and Anai A. Valencia-Lazcano. 2023. "Chitosan: Properties and Its Application in Agriculture in Context of Molecular Weight" Polymers 15, no. 13: 2867. https://doi.org/10.3390/polym15132867