
A Review of Development and Utilization for Edible Fungal Polysaccharides: Extraction, Chemical Characteristics, and Bioactivities
Abstract
:1. Introduction
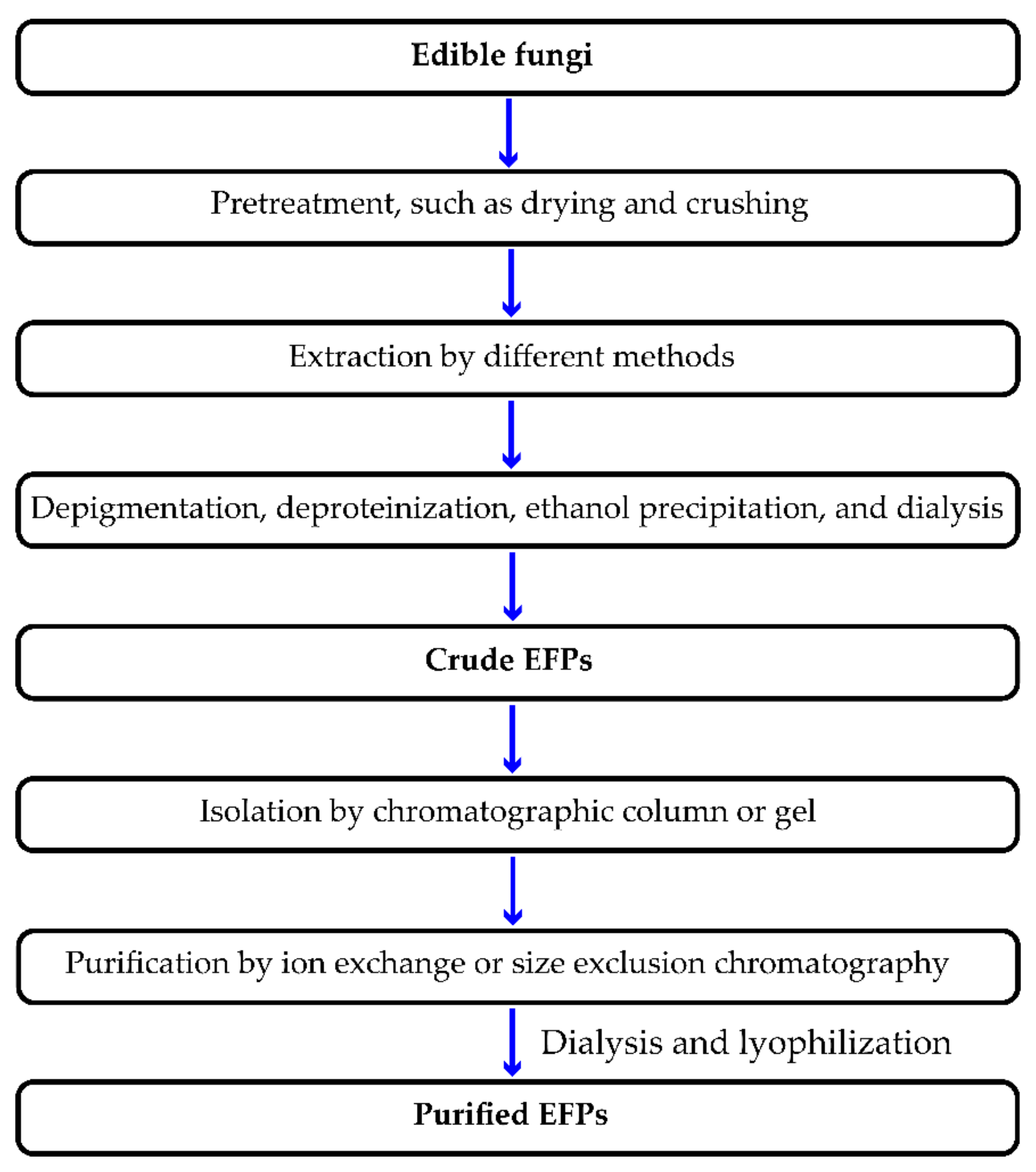
2. Extraction Method of EFPs
2.1. HWE
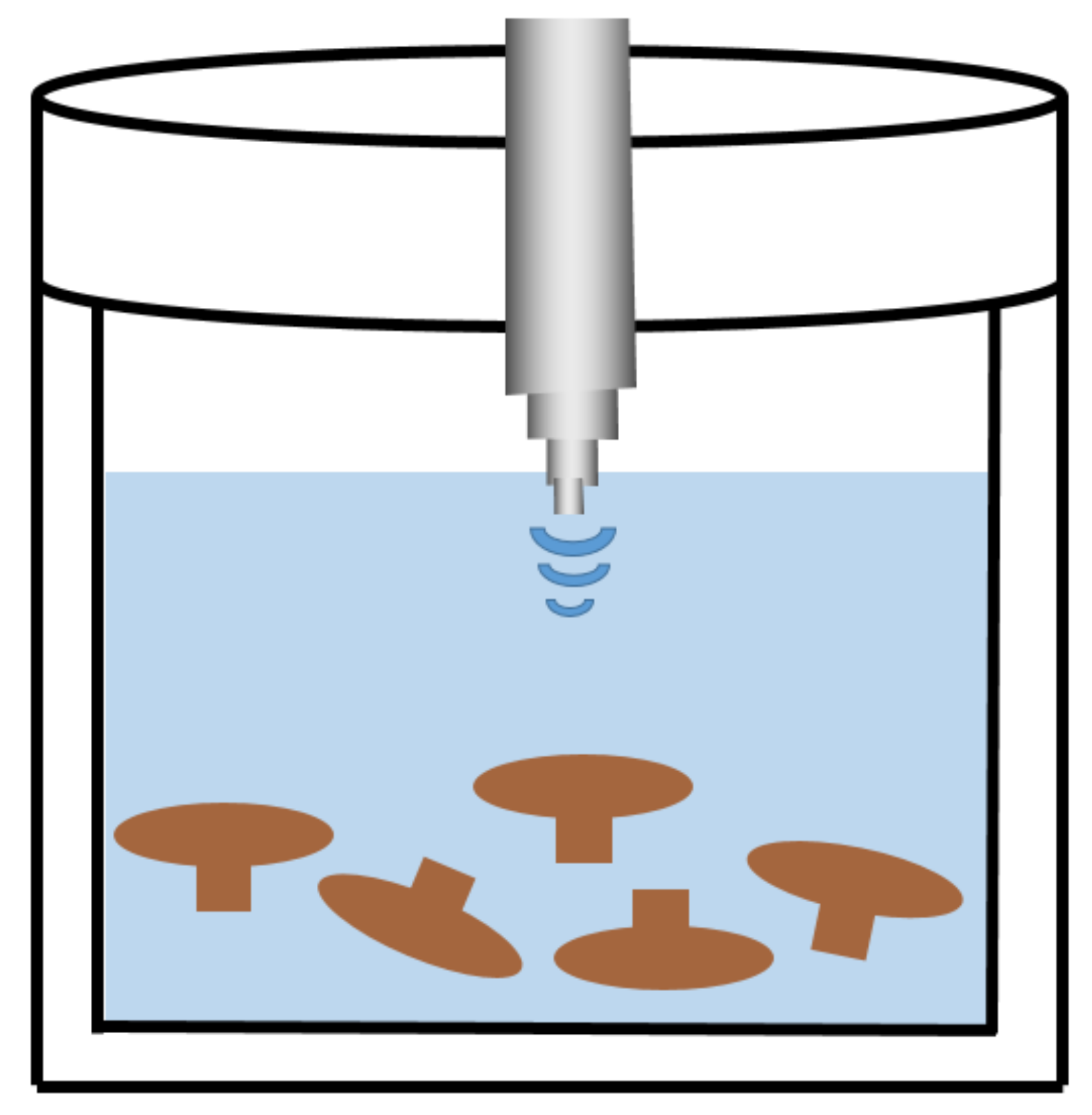
2.2. UAE
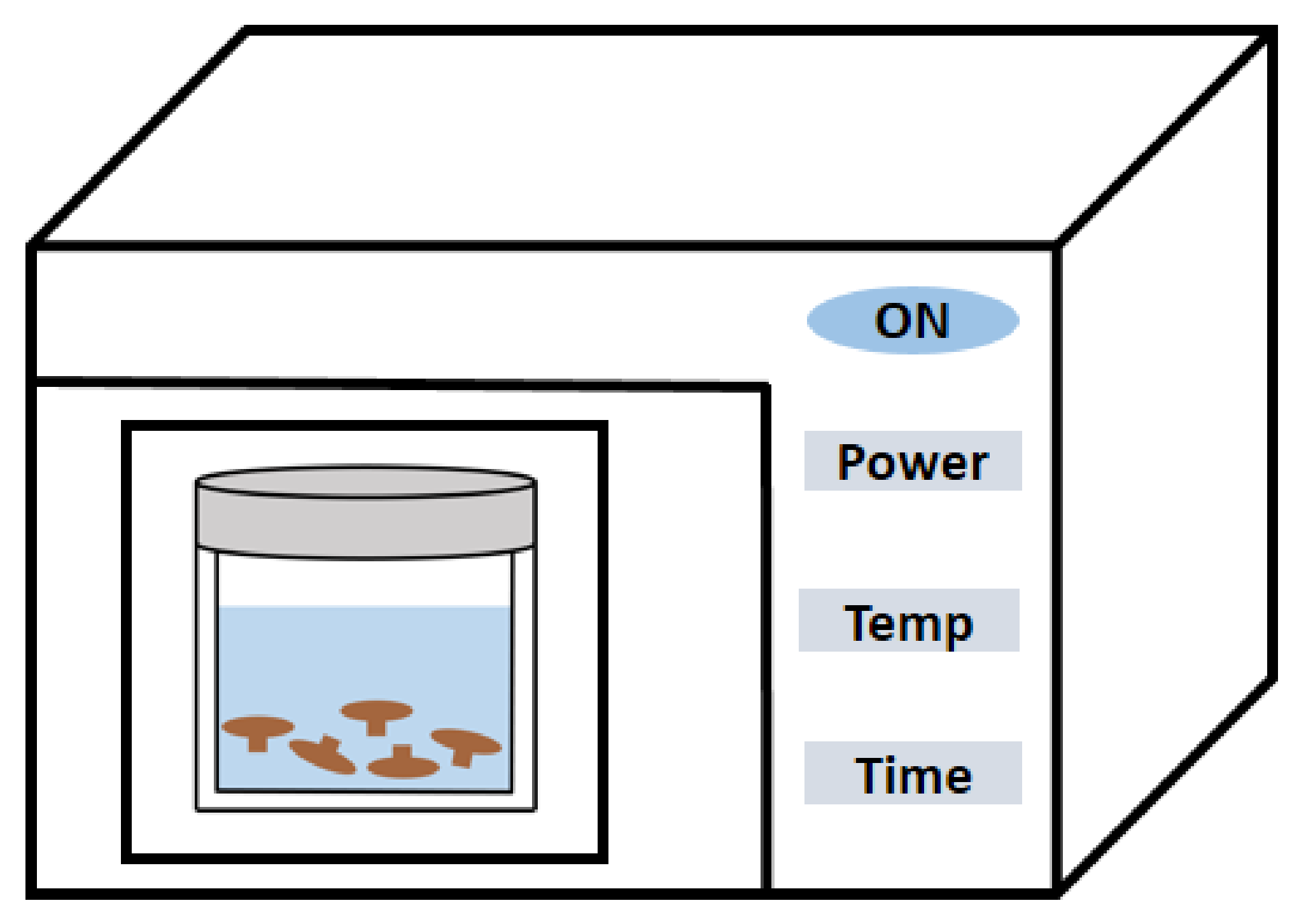
2.3. MAE
2.4. EAE
2.5. Alkali/Acid-Assisted Extraction (AAE)
2.6. Fermentation-Assisted Extraction (FAE)
2.7. Others

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Extraction Method | Edible Fungus Origin | Extraction Step | Yield | Ref |
|---|---|---|---|---|
| Hot water extraction | Cordyceps sinensis | Extraction in boiling water; repeated 2 times for 2 h each time | / | [21] |
| Flammulina velutipes mycelium | Degreasing with 95% ethanol, extraction with hot water at 95 °C for 4 h (repeated twice), rotary evaporation under reduced pressure at 50 °C, precipitation with 98% ethanol, deproteinization with cetyltrimethylammonium bromide (CTAB), decolorization by macroporous ion exchange resin, and precipitation and separation with 95% ethanol | 2.2% | [22] | |
| Lentinus edodes | Extraction in hot water at 98 °C for 1 h (50 g/L, w/v), cross-flow microfiltration, and reverse osmosis (nanofiltration) | 5% | [23] | |
| Pleurotus florida | Extraction in boiling water for 6 h | / | [24] | |
| Sarcodon aspratus | Pretreatment with 95% ethanol for 1.5 h and extraction with boiling water for 1.5 h (repeated 3 times) | / | [25] | |
| Lentinus edodes | Pretreatment with 95% ethanol for 24 h, extraction in boiling water twice, 1 h each time; precipitation with 95% alcohol, decolorization with H2O2, and protein removal | 0.9% | [26] | |
| Dictyophora indusiata | Extraction in boiling water for 2 h (1:30, w/w) and precipitation with absolute ethanol | 13.2% | [27] | |
| Ganoderma applanatum | Extraction in boiling water for 3 h and precipitation with 95% ethanol (1:3, v/v) at 4 °C for 24 h | 2.14% | [28] | |
| Poria cocos Wolf | Degreasing with petroleum ether for 24 h (repeated twice), extraction in boiling water for 2 h (20:1 mL/g, w/v), precipitation with ethanol overnight (4 °C), and protein removal by Sevag method | / | [29] | |
| Ultrasonic-assisted extraction | Medicinal Fungi | Extraction by ultrasonic (20 kHz, 130 W) at 45–50 °C for 30 min with a solid–liquid ratio of 1:3, w/v | 18% | [39] |
| Flammulina velutipes Stipe | Ultrasonic extraction (680 W) for 19.8 min at a liquid-to-solid ratio of 28 mL/g, w/v | 16.20% | [40] | |
| Poria cocos Wolf | Degreasing with petroleum ether for 24 h and then extraction in a 200 W ultrasonic bath at a liquid–solid ratio of 20:1 (mL/g) for 1 h | / | [29] | |
| Grifola frondosa | Extraction by sonication (1:30 g/mL, w/v) at 50 kHz for 30 min, and then extraction in a water bath (90 °C) for 3 h and precipitation with absolute ethanol | / | [41] | |
| Phellinus linteus mycelia | Ultrasonic extraction (5.0%, w/v) at different powers (200, 400 and 600 W) in a 40 °C water bath for 30, 60, 90, 120, 150 and 180 min. | / | [42] | |
| Microwave-assisted extraction | Poria cocos Wolf | Degreasing pretreatment and microwave-assisted extraction for 2 min (20:1 mL/g, w/v, 2450 MHz, 800 W) | 9.95% | [29] |
| Mushroom | Microwave-assisted extraction (1:30, w/v, 2455 MHz, 30 bar) and precipitation with ethanol | 12.1–44.2% | [49] | |
| Cordyceps gunnii mycelia | Microwave-assisted extraction at 70 °C for 5 min (280 W) at a solid-to-liquid ratio of 1:20, w/v, repeated 3 times, followed by precipitation with ethanol at 4 °C for 12 h | 30.35% | [50] | |
| Enzyme-assisted extraction | Poria cocos Wolf | Degreasing with petroleum ether for 24 h, followed by extraction with 1.0% cellulase at a liquid-solid ratio of 20:1 (mL/g) for 1 h | / | [29] |
| Tricholoma mongolicum Imai | Degreasing pretreatment, extraction with 20 g/kg cellulase in water bath for 127 min (50 g/L, w/v, pH = 4.0, 50 °C), and then precipitated with ethanol | 18.96% | [8] | |
| Cordyceps gunnii mycelia | Extraction with 2% solution containing 0.5% cellulase (1:20, w/v, pH 5.0) at 55 °C for 80 min | 32.50% | [50] | |
| Armillaria mellea mushroom | Extraction (30:1 mL/g, w/v) with 1.9% composite enzyme (cellulose and papain, 2:1) at 50 °C for 138 min, accompanied by 360 W ultrasonic for 20 min | 40.56% | [55] | |
| Alkali/acid-assisted extraction | Lentinus edodes | Extraction in boiling water, repeated 2 times for 1 h each time; treatment with 2% NaOH and 0.05% NaBH for 24 h; precipitation with 95% ethanol | 4.8% | [26] |
| Mushroom fruiting bodies | Degreasing with 95% ethanol (1:10, w/v); extraction in boiling water (1:20, w/v), repeated 3 times for 4 h each time; extraction with 0.5 M NaOH and a trace amount of NaBH for 3 h ×2 and 2 h ×1 at 80 °C (1:20, w/v), respectively; precipitation with 95% ethanol | 1.03–16.6%, | [62] | |
| Russula albonigra (Krombh.) Fr. | Boiling in 4% NaOH for 1 h; precipitation with 1:5 (v/v) EtOH; purification by gel permeation chromatography | / | [63] | |
| Phellinus linteus | Extraction in hot water at 95 °C for 8 h, repeated twice; repeating the last step 2 times under the same conditions with 1% (NH4)2C2O4; extraction with 1.25 M NaOH/0.05% NaBH4 at room temperature, repeated twice for 3 h each time | 9.12–19.49% | [60] | |
| Pleurotus sajor-caju | Extraction with 1% ammonium oxalate at 98 °C for 3–5 h; precipitation with ethanol; extraction with a mixture of 5% NaOH and 0.05% NaBH at 30 °C for 12–20 h | / | [64] | |
| Pleurotus ostreatus | Extraction with 3.8% HCl at 30 °C for 300 min | / | [65] | |
| Fermentation extraction | Fermentation broth of Morchella conica | Precipitation with 95% ethanol after centrifugation, deproteinization, and degreasing | 5.09% | [70] |
| Pleurotus geesteranus 5(#) | Production of exopolysaccharide (EPS) in optimally defined medium (per liter) of 60.0 g maltose, 5.0 g tryptone, 1 mM NaCl, 5 mM KH2PO4, initial pH 6.0 (28 °C) | 16.97 g/L | [71] | |
| Pleurotus geesteranus 5(#) | Growing in glucose peptone medium (GP) medium (0.3% peptone, 3% glucose) for 4 d (150 rpm, 26 °C); fermented in a 5 L stirrer using 4% (v/v) mycelium suspension | 11.09 g/L | [72] | |
| Agaricus bisporus MJ-0811 | Culture in potato dextrose agar (PDA) medium for 4 d (30 °C) and then inoculation into liquid medium (150 rpm, 4 d); precipitation with 95% ethanol at 4 °C for 24 h | 2.69 g/L | [73] | |
| Coprinus comatus | Extraction in boiling water for 1 h, repeated 3 times; precipitation with 95% ethanol for 12 h | 4.50–41.27% | [74] | |
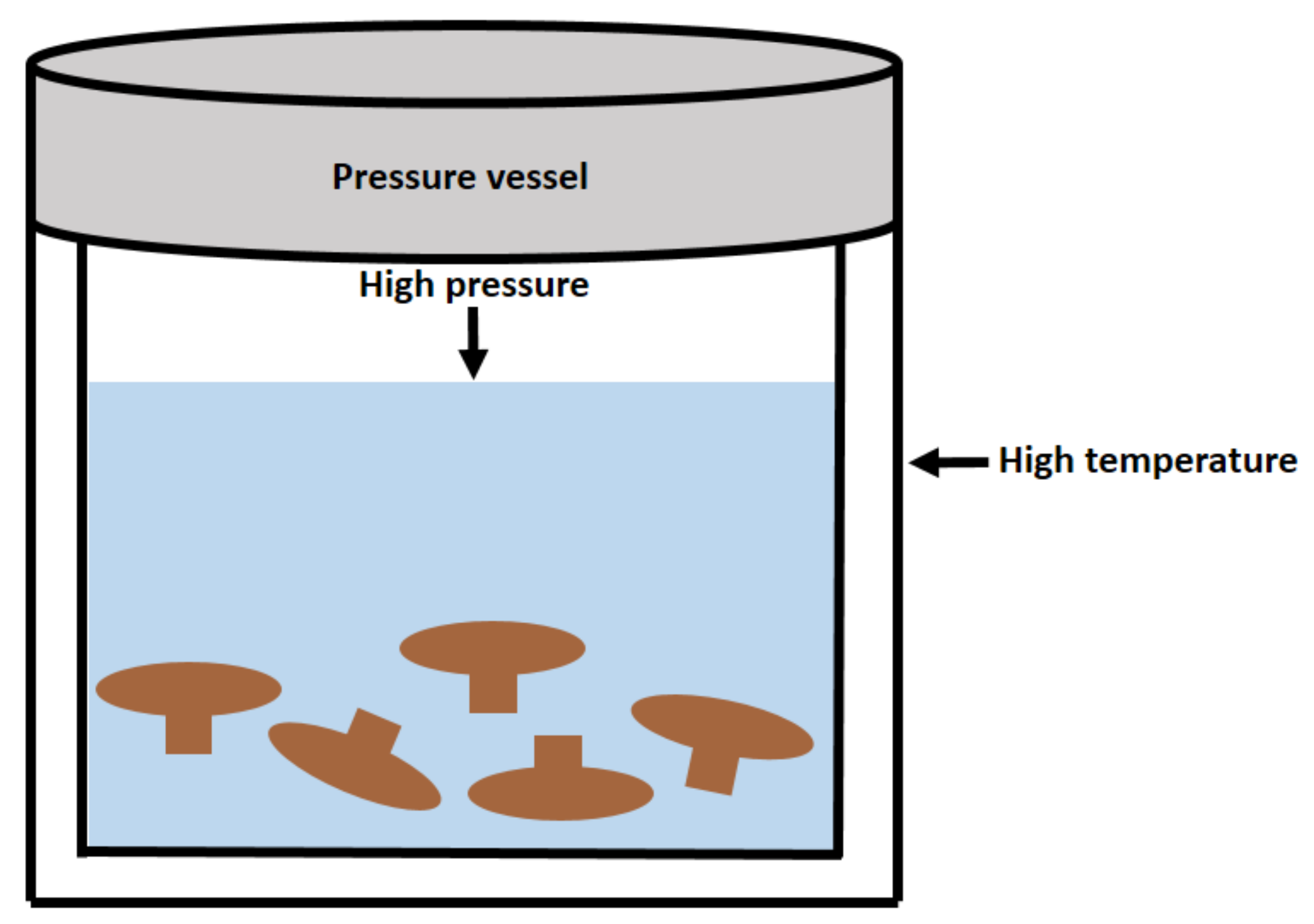
| Subcritical water extraction | Lentinus edodes | Subcritical water extraction at 5 MPa | 11.03–14.11% | [33] |
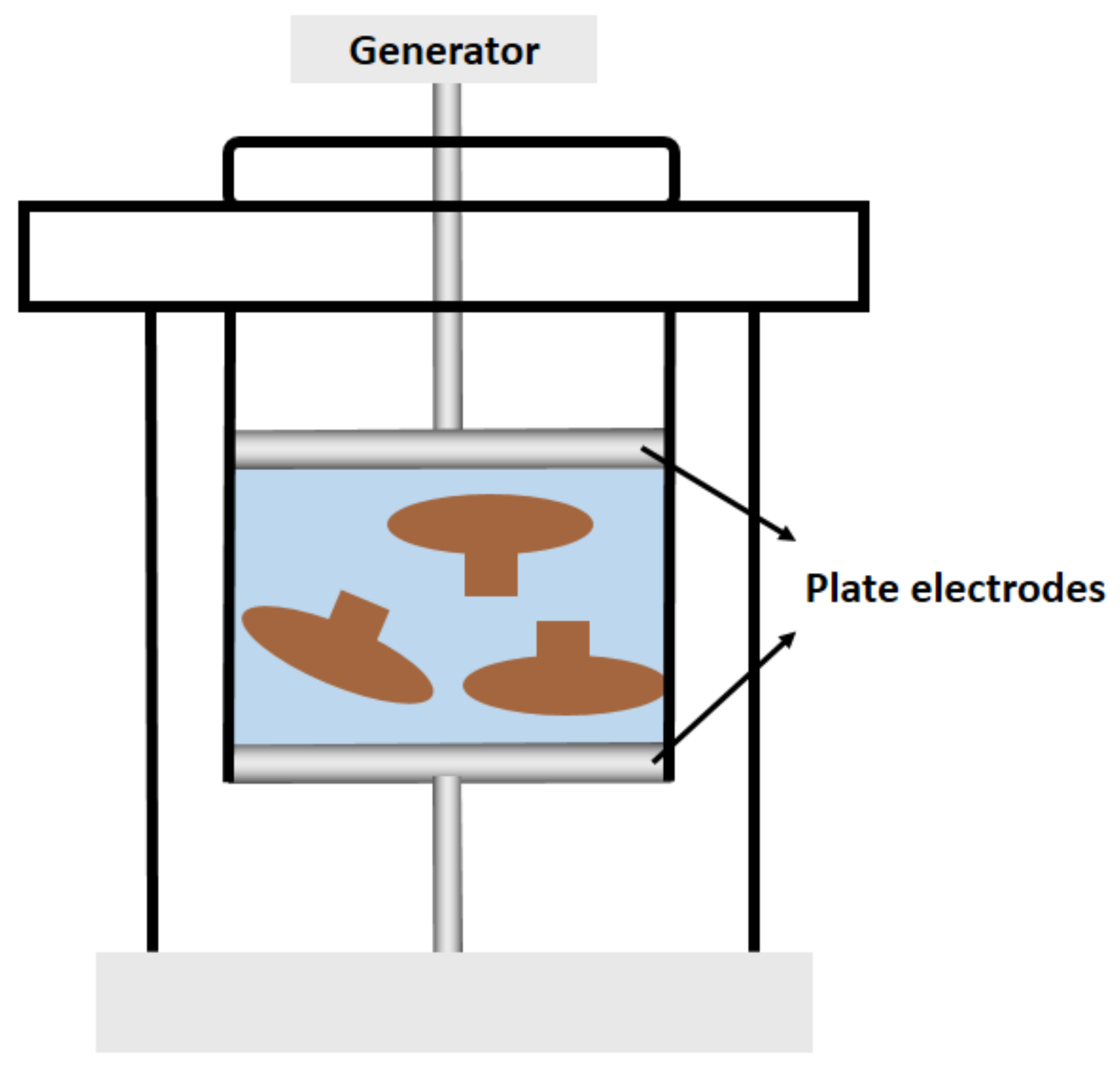
| High voltage pulsed electric field-assisted extraction | Morchella esculenta | Extraction (27 mL/g, w/v) by high-voltage pulsed electric field (18 kV/cm electric field strength, number of pulses 7) | 56.03 μg/mL | [18] |
| Complex extraction | Lentinus edodes | Extraction by microwave (440 W, 10 min) and enzyme (48 °C, pH = 5.0) | 5.42% | [79] |
| Homogenate extraction | Lentinus edodes | Extraction for 66 s (30 mL/g, w/v, pH = 10) | 13.2% | [81] |
| Vacuum extraction | Lentinus edodes | Extraction for 25 min (26 mL/g, w/v, 0.08 MPa, 62 °C) | 4.28% | [82] |
| Nanoparticle extraction | Sparassis crispa and Phellinus linteus | Extraction for 2 h (20 mL/g w/v, pH = 10), emulsification with 20% nanoparticles under high pressure at 30 °C for 40 min, and then HWE at 95 °C for 30 h | 54.2% | [83] |
3. Preliminary Chemical Composition of EFPs
3.1. MW
3.2. Monosaccharide Composition
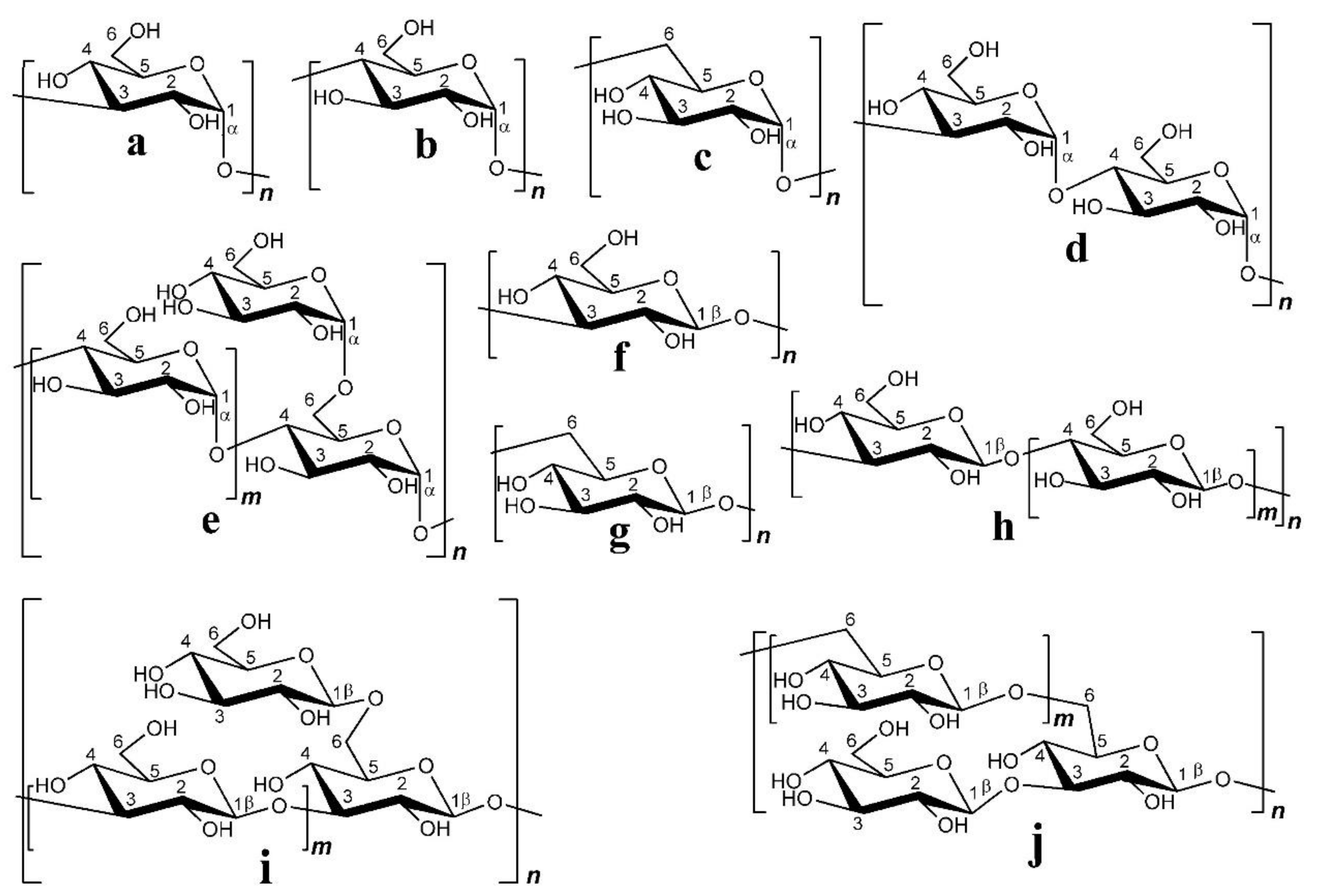
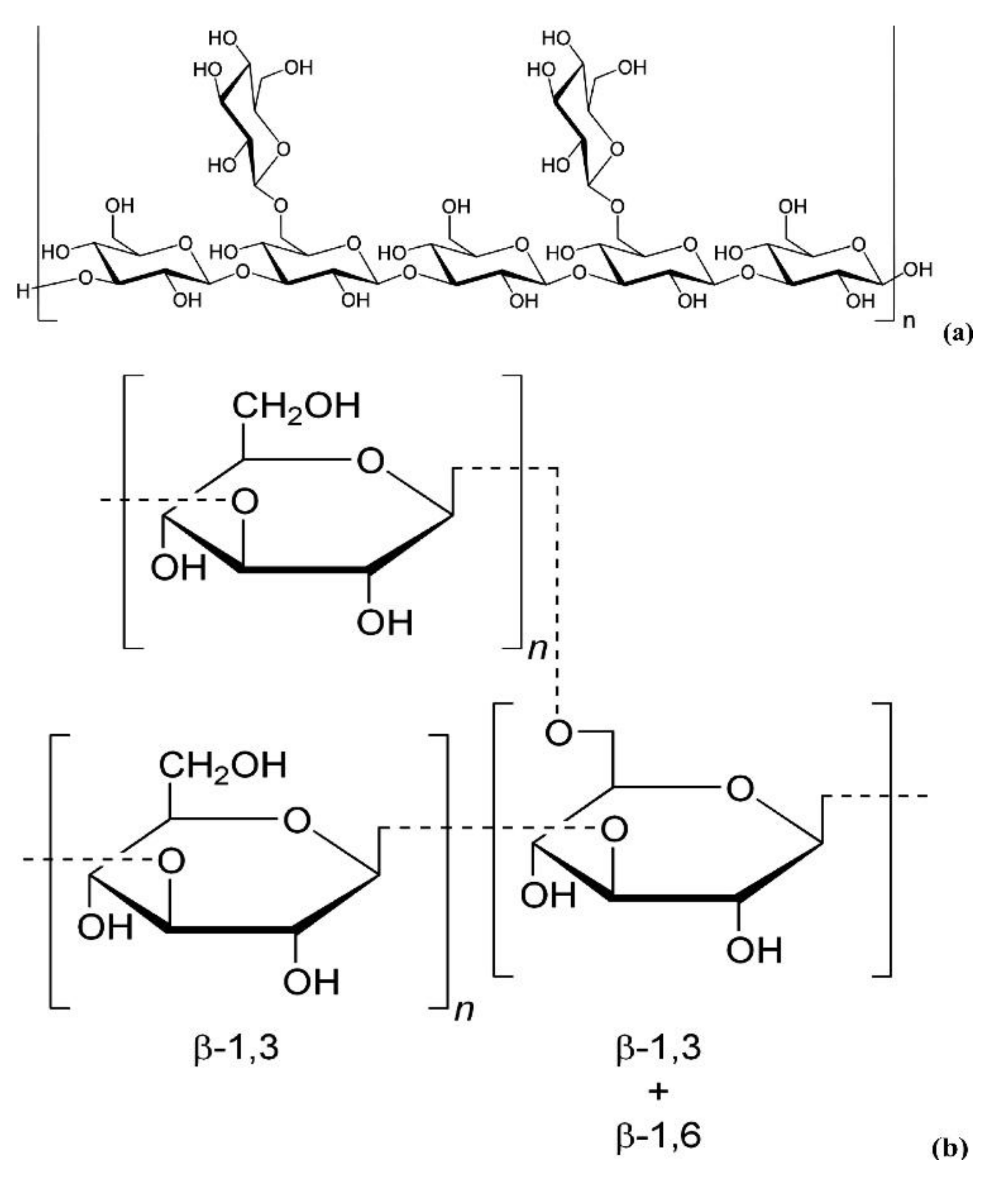
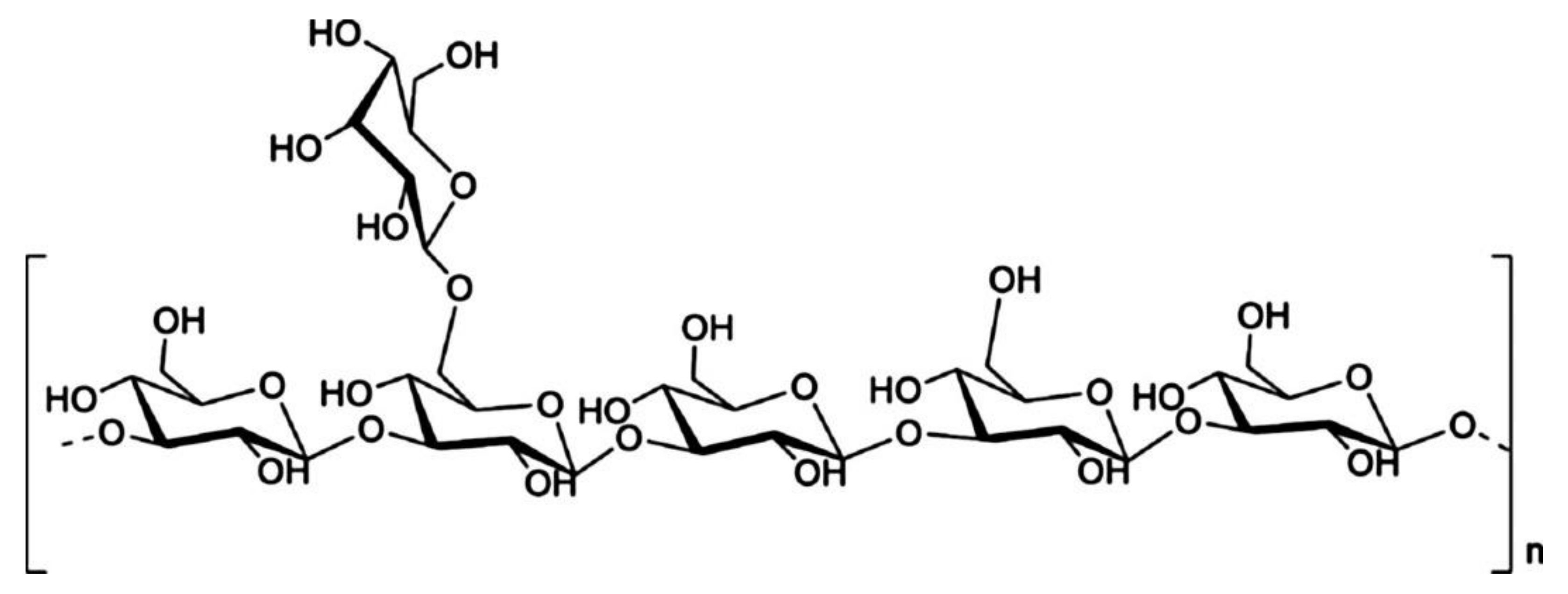
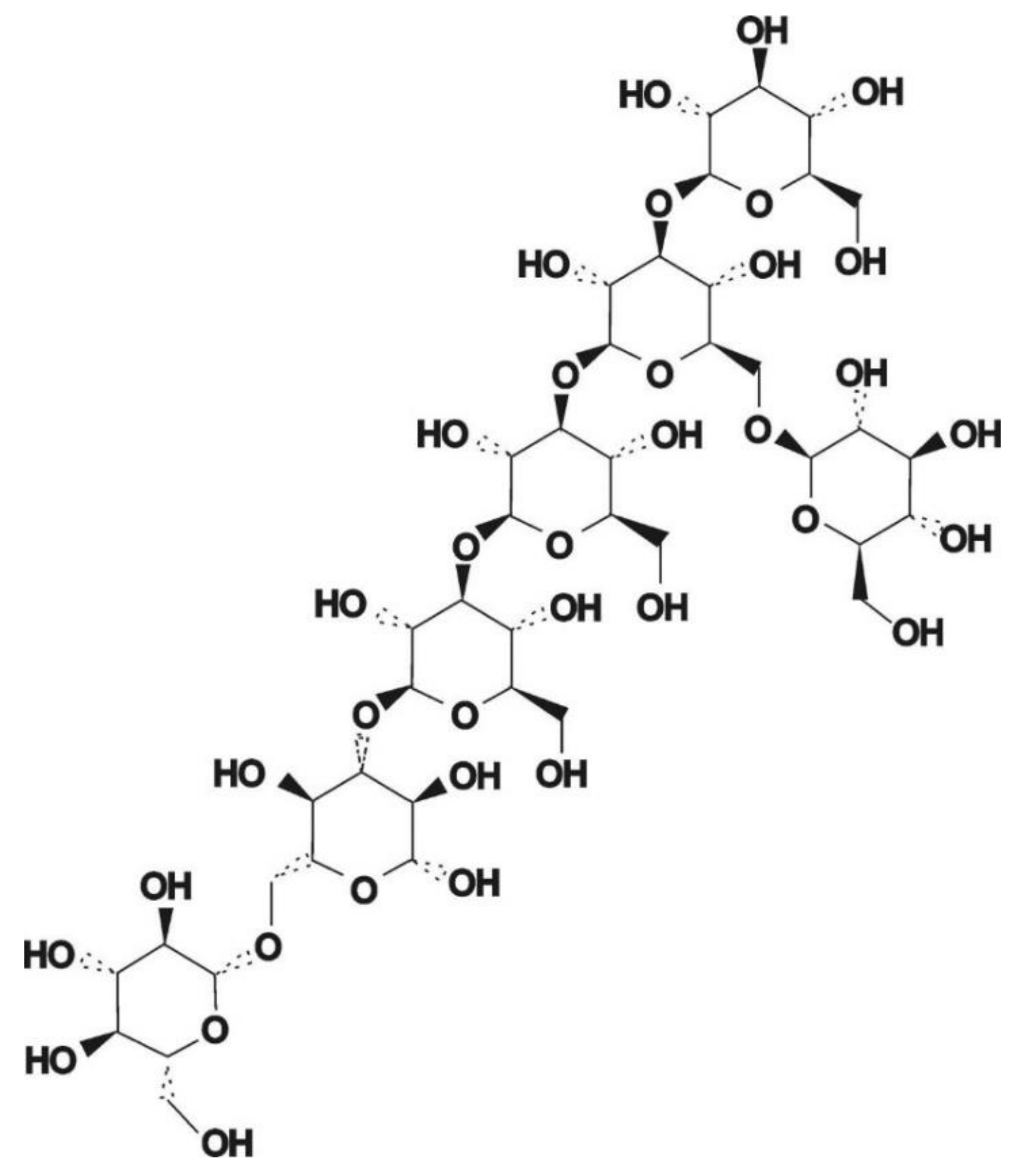
3.3. Glycosidic Bond Type and Backbone Structure
4. In Vitro Bioactivity of EFPs
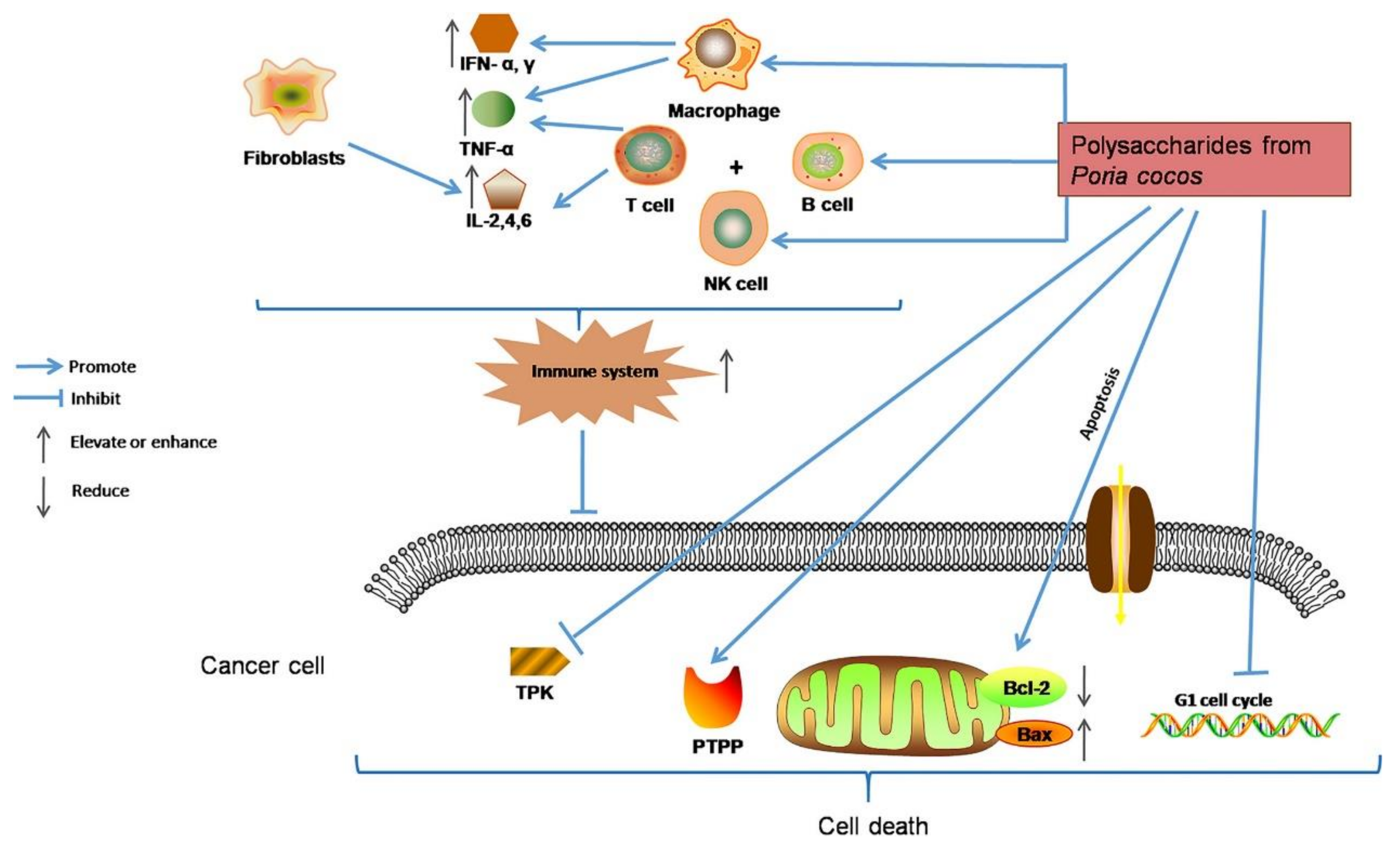
4.1. Antineoplastic Activity
4.2. Immunomodulatory Activity
4.3. Antioxidant Activity
4.4. Glycosidase Inhibitory Activity
| Bioactivity | Edible Fungus Origin | Regulatory Mechanism | Ref |
|---|---|---|---|
| Antioxidant activity | Poria cocos Wolf | Favorable in vitro Fe2+ chelation, DPPH scavenging, and hydroxyl radical scavenging activities | [29] |
| Agaricus bisporus | Strong in vitro scavenging activity of hydroxyl radicals (OH−) and superoxide radicals (O2−) | [73] | |
| Coprinus comatus | Excellent in vitro Fe2+ chelation, 1,1-diphenyl-2-picrylhydrazyl (DPPH), and OH− scavenging activities | [74] | |
| Oudemansiella radicata | Strong DPPH, 2,2-azino-bis (3-ethylbenzothiazoline-6-sulphonic acid) (ABTS), OH−, ·O2− scavenging activity, and Fe2+ chelation activity | [31] | |
| Russula griseocarnosa | Strong DPPH, ABTS, OH−, and ·O2− scavenging activities | [79] | |
| Paxillus involutus | Significant hydroxyl, DPPH, ABTS, and ·O2− radical scavenging activities | [32] | |
| Lepista nuda | Strong capability to chelate iron ions, scavenge DPPH and ·O2− radicals | [89] | |
| Immunostimulatory activity | Cordyceps sinensis | Enhancing SOD activity, reducing MDA concentration, and regulating the secretion of cytokines IL-4, IL-5, and IL-17 in serum | [21] |
| Flammulina velutipes mycelium | Significantly promoting NO production, interleukin-1 (IL-1) production, and tumor necrosis factor-α (TNF-α) secretion in macrophages | [22] | |
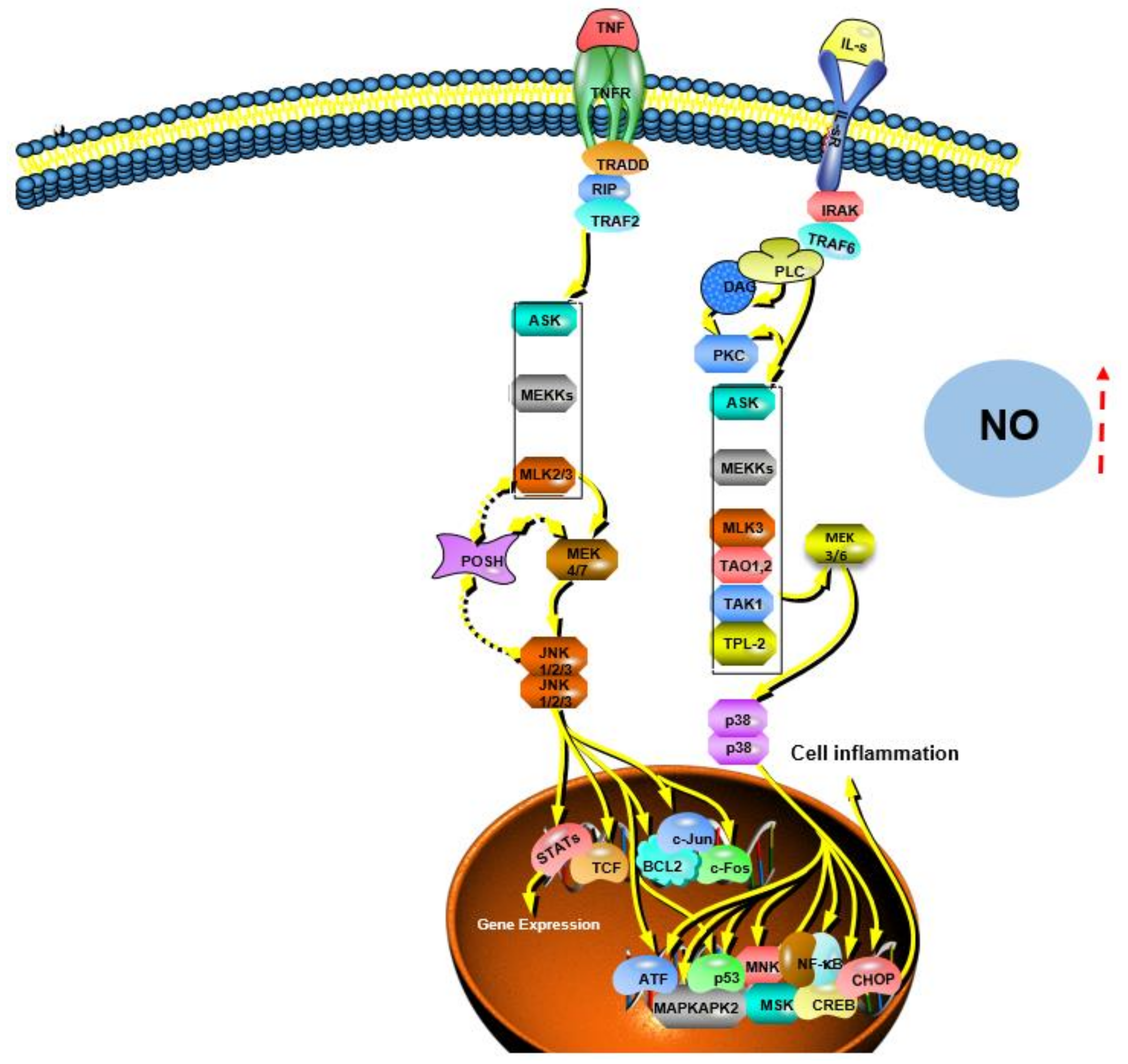
| Inonotus obliquus | Promoting NO/ROS production, TNF-α secretion, and phagocytic uptake in macrophages; promoting the proliferation and complexation of mouse splenocytes; effectively promoting macrophage activation through MAPK and NF-κB signaling pathways | [119] | |
| Morchella conica | Regulating NO production in macrophages, modulating innate immune responses within a specific concentration range, and promoting splenocyte proliferation | [70] | |
| Phellinus linteus | Inhibiting MyD88-dependent and MAPK signaling pathways, resulting in intracellular inhibition of ROS formation, cytokine (TNF-α, IL-1α, IL-1β, and IL-4) production, VCAM-1 expression, JNK, and p38 activation and phosphorylation, and NF-κB activation | [86] | |
| Coriolus versicolor mushroom | Directly binding mIg with repetitive epitopes, allowing efficient cross-linking of mIg on the B cell surface and driving B cell proliferation and Ig production, interacting with TLR4 to stimulate B cell activation | [120] | |
| Cordyceps taii | Increasing thymus weight and enhancing immune function of pancreatic β cells in STZ-induced diabetic mice | [142] | |
| Antineoplastic activity | Lentinus edodes | Directly inhibiting the in vitro proliferation of tumor cells H22, HepG2, and SMMC-7721; significantly inhibiting the growth of tumors in mice and greatly increasing the levels of IL-2 and TNF-α in serum; inducing tumor cell apoptosis | [26] |
| Lentinus edodes | Inhibiting the proliferation of MCF-7 cells; triggering the mitochondrial apoptosis pathway by activating caspase-7 and activating autophagy in MCF-7 cells, thereby accelerating cell apoptosis | [105] | |
| Russula griseocarnosa | Inhibiting the proliferation of Hela and Siha cervical cancer cells | [79] | |
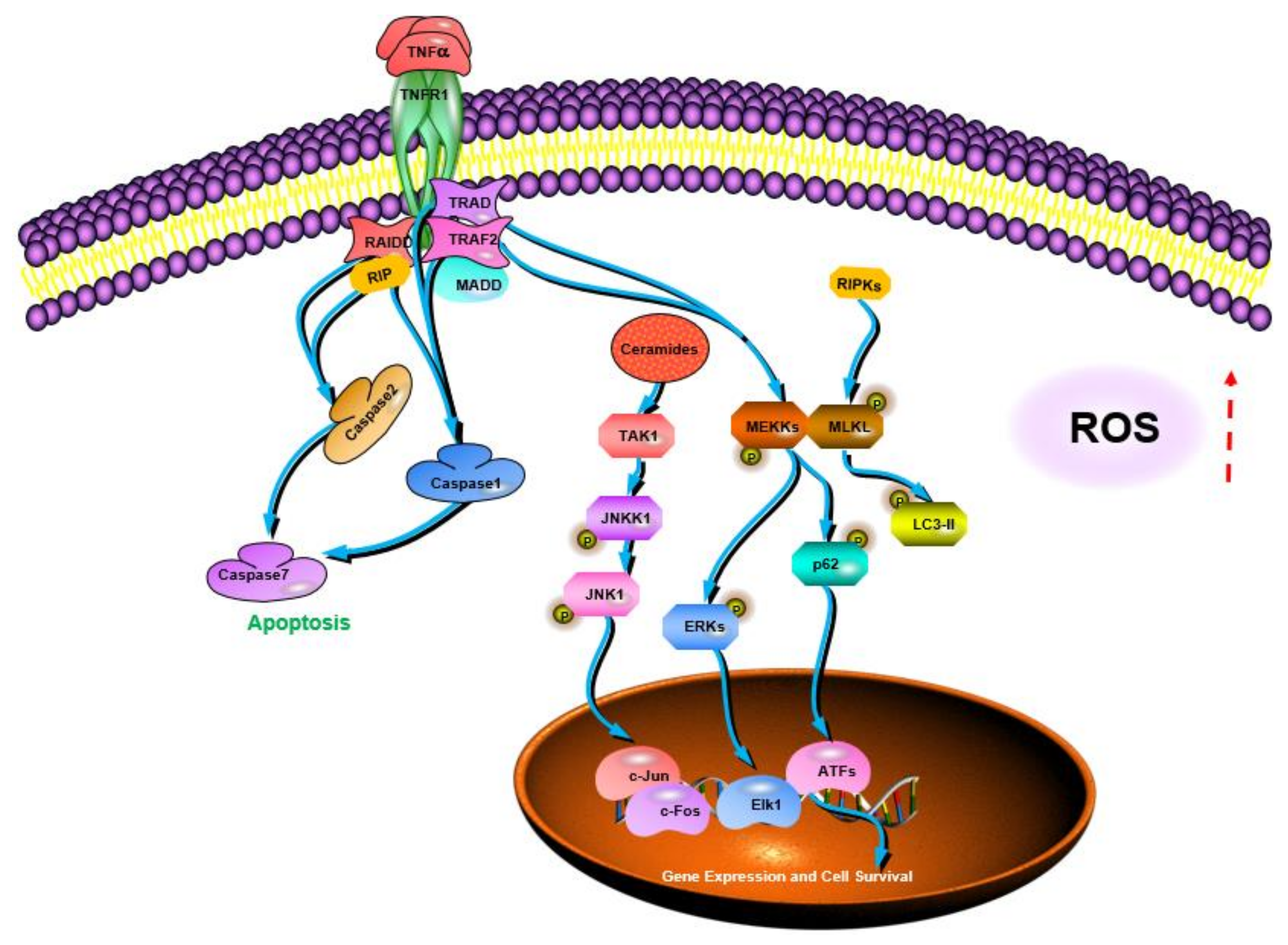
| Coriolus versicolor | Mediating the necroptosis of cancer cells by activating the RIPK1/RIPK3/MLKL pathway; inducing the activation of the TNF-α/TNFR1 pathway in breast cancer cells; inhibiting the expression of TNF-α in melanoma cells and triggering the production of ROS | [106] | |
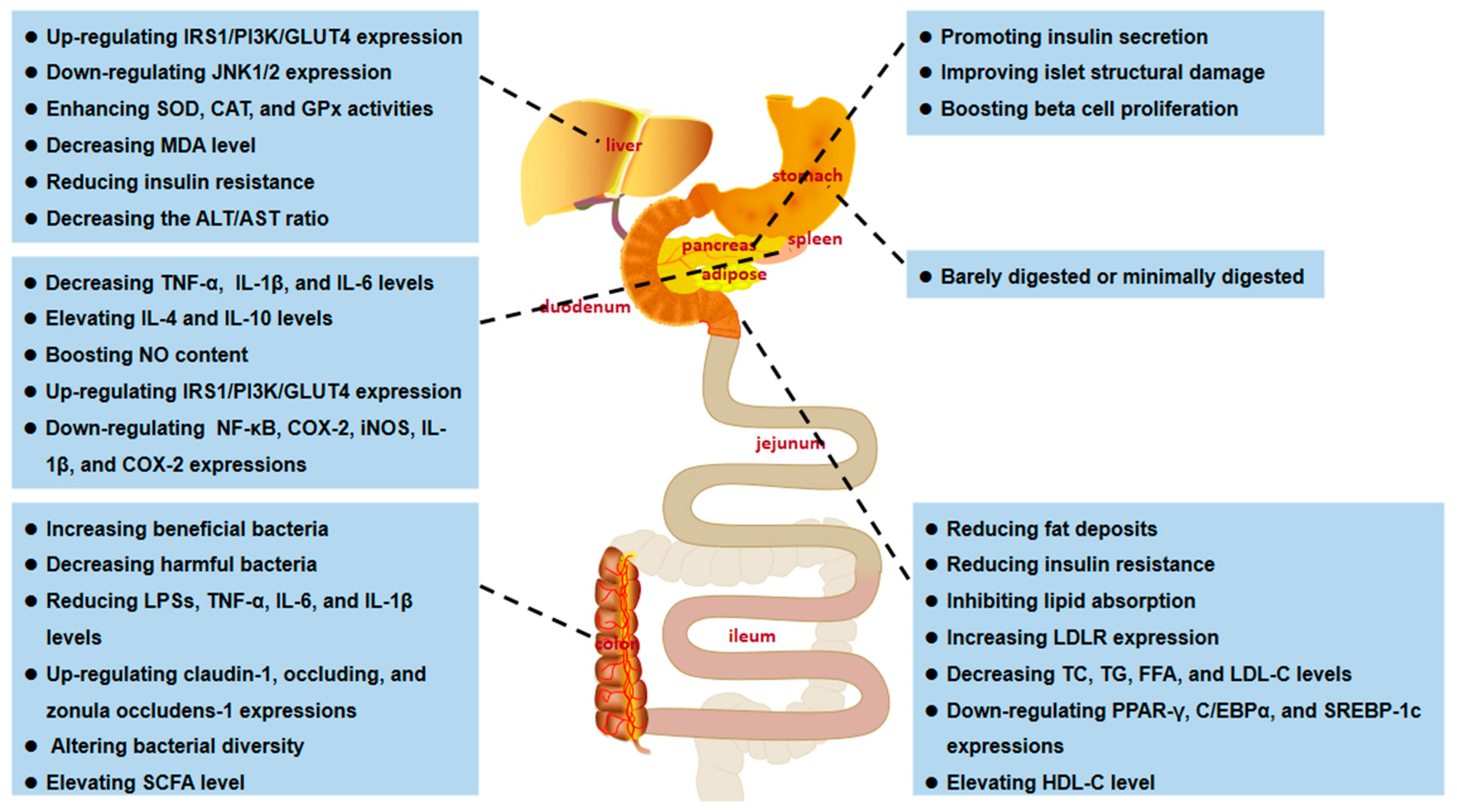
| Anti-obesity activity | Pleurotus eryngii | Inhibiting lipid absorption and increasing mesenteric fat, reducing body weight, promoting LDLR expression, and enhancing serum LDL cholesterol uptake | [143] |
| Dictyophora indusiata | Reversing HFD-induced changes in obesity-related parameters, lowering body weight, and reducing the degree of lipid accumulation; decreasing the expression of lipid genes (PPAR-γ, C/EBPα, and SREBP-1c) | [144] | |
| Pleurotus geesteranus 5(#) | Lowering total cholesterol, triacylglycerol, LDL cholesterol, HDL cholesterol, and atherogenic index | [72] | |
| Ganoderma applanatum | Slowing the rate of food intake in obese rats; significantly reducing TC, TG, LDL-C levels, and atherosclerosis index | [28] | |
| Cordyceps taii | Regulating lipid metabolism parameters (serum levels of TC, TG, LDL-C, and HDL-C) in diabetic mice and effectively improving lipid disorders | [142] | |
| Hypoglycemic activity | Pleurotus geesteranus 5(#) | Improving pancreatic β-cell damage, promoting insulin synthesis, and lowering blood glucose | [72] |
| Grifola frondosa | Decreasing fasting blood glucose (FBG) levels; regulating blood biochemical parameter levels and inflammatory responses in the liver and kidneys; improving glucose consumption and alleviating the insulin resistance (IR) by increasing IRS1/PI3K/GLUT4 gene/protein expression and decreasing JNK1/2 gene/protein expression | [41] | |
| Suillellus luridus | Increasing serum insulin levels, enhancing antioxidant enzyme (SOD, CAT, and GPx) activity in the liver and kidneys, and decreasing MDA levels | [88] | |
| Cordyceps taii | Reducing FBG level and IR, increasing insulin level, significantly improving islet structure damage, and promoting β-cell proliferation | [142] | |
| Coprinus comatus | Significant in vitro alpha-amylase inhibitory activity | [74] | |
| Mushroom fruiting bodies | Activating autophagy to reduce IR and fat deposition; effectively lowering blood glucose and protecting islets from damage caused by hyperglycemia | [62] | |
| Intestinal homeostasis-regulating activity | Dictyophora indusiata | Reversing gut dysbiosis and increasing beneficial flora, including Lactobacillus and Ruminococaceae, decreasing endotoxemia by decreasing lipopolysaccharides (LPSs) and pro-inflammatory cytokines levels, such as TNF-α, IL-6, and interleukin-1β (IL-1β), and increasing the expression of claudin-1, occludin, and zonula occludens-1 | [27] |
| Pleurotus eryngii | Altering the gut environment to increase the abundance of short-chain fatty acid (SCFA)-producing bacteria (Anaerostipes and Closterridium) and decrease the abundance of Roseburia | [143] | |
| Dictyophora indusiata | Restoring the altered bacterial flora caused by the HED diet and increasing bacterial diversity; decreasing the Firmicutes to Bacteroidetes ratio | [144] | |
| Ganoderma lucidum and Poria cocos | Remodeling gut microbiota composition, reducing species richness, reducing Firmicutes/Bacteroidetes ratio, and significantly promoting lactic acid-producing (LAP) and SCFA-producing bacterial growth | [145] | |
| Grifola frondosa | Increasing the abundance of Bacteroidetes in the gut while decreasing the numbers of Firmicutes and Proteus; increasing the relative abundance of SCFA-producing bacteria, including Alloprevotella and Blautia | [41] | |
| Anti-inflammatory activity | Dictyophora indusiata | Reducing pro-inflammatory cytokine levels (TNF-α, IL-1β, and IL-6) in adipose and liver tissue while enhancing anti-inflammatory cytokines ((IL-4 and IL-10) expression | [144] |
| Inonotus obliquus | Inhibiting the NF-κB, COX-2, and iNOS signaling pathways | [146] | |
| Agaricus bisporus | Inhibiting the expression of pro-inflammatory genes, including IL-1β and COX-2 | [147] | |
| Anti-aging activity | Agaricus bisporus | Strong OH- and DPPH free radical scavenging and iron ion chelation capability, significantly improving D-galactose-induced liver damage, kidney damage, and early dysregulation of lipid metabolism | [148] |
| Flammulina velutipes | Strong DPPH, OH- radical scavenging, and Fe2+ chelating activity; enhancing antioxidant enzyme (SOD, GSH-Px, CAT, and T-AOC) activities; reducing lipid peroxidation (MDA), and improving inflammatory response (reducing Ach E and NOS activities) | [149] |
5. In Vivo Bioactivity of TFPs
5.1. Antidiabetic Activity
5.2. Anti-Obesity Activity
5.3. Antitumor Activity
5.4. Intestinal Homeostasis-Regulating Activity
6. Others
7. Conclusions and Prospectives
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Carbonero, E.R.; Gracher, A.H.P.; Smiderle, F.R.; Rosado, F.R.; Sassaki, G.L.; Gorin, P.A.J.; Iacomini, M. A β-glucan from the fruit bodies of edible mushrooms Pleurotus eryngii and Pleurotus ostreatoroseus. Carbohydr. Polym. 2006, 66, 252–257. [Google Scholar] [CrossRef]
- Dubey, S.K.; Chaturvedi, V.K.; Mishra, D.; Bajpeyee, A.; Tiwari, A.; Singh, M.P. Role of edible mushroom as a potent therapeutics for the diabetes and obesity. 3 Biotech. 2019, 9, 450. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Shi, X.; Yin, J.; Nie, S. Bioactive polysaccharide from edible Dictyophora spp.: Extraction, purification, structural features and bioactivities. Bioact. Carbohydr. Diet. Fibre 2018, 14, 25–32. [Google Scholar] [CrossRef]
- Lu, X.K.; Brennan, M.A.; Narciso, J.; Guan, W.Q.; Zhang, J.; Li, Y.; Serventi, L.; Brennan, C.S. Correlations between the phenolic and fibre composition of mushrooms and the glycaemic and textural characteristics of mushroom enriched extruded products. LWT-Food Sci. Technol. 2020, 118, 108730. [Google Scholar] [CrossRef]
- Leong, Y.K.; Yang, F.C.; Chang, J.S. Extraction of polysaccharides from edible mushrooms: Emerging technologies and recent advances. Carbohydr. Polym. 2021, 251, 117006. [Google Scholar] [CrossRef]
- Steimbach, L.; Borgmann, A.V.; Gomar, G.G.; Hoffmann, L.V.; Rutckeviski, R.; de Andrade, D.P.; Smiderle, F.R. Fungal beta-glucans as adjuvants for treating cancer patients-A systematic review of clinical trials. Clin. Nutr. 2021, 40, 3104–3113. [Google Scholar] [CrossRef]
- Smiderle, F.R.; Morales, D.; Gil-Ramírez, A.; de Jesus, L.I.; Gilbert-López, B.; Iacomini, M.; Soler-Rivas, C. Evaluation of microwave-assisted and pressurized liquid extractions to obtain β-d-glucans from mushrooms. Carbohydr. Polym. 2017, 156, 165–174. [Google Scholar] [CrossRef]
- Zhao, Y.M.; Song, J.H.; Wang, J.; Yang, J.M.; Wang, Z.B.; Liu, Y.H. Optimization of cellulase-assisted extraction process and antioxidant activities of polysaccharides from Tricholoma mongolicum Imai. J. Sci. Food Agric. 2016, 96, 4484–4491. [Google Scholar] [CrossRef]
- Zhang, Z.F.; Lv, G.Y.; Song, T.T.; Jin, Q.L.; Huang, J.B.; Fan, L.F.; Cai, W.M. Comparison of the preliminary characterizations and antioxidant properties of polysaccharides obtained from Phellinus baumii growth on different culture substrates. Carbohydr. Polym. 2015, 132, 397–399. [Google Scholar] [CrossRef]
- Luan, F.; Ji, Y.; Peng, L.; Liu, Q.; Cao, H.; Yang, Y.; He, X.; Zeng, N. Extraction, purification, structural characteristics and biological properties of the polysaccharides from Codonopsis pilosula: A review. Carbohydr. Polym. 2021, 261, 117863. [Google Scholar] [CrossRef] [PubMed]
- Miao, J.; Regenstein, J.M.; Qiu, J.; Zhang, J.; Zhang, X.; Li, H.; Zhang, H.; Wang, Z. Isolation, structural characterization and bioactivities of polysaccharides and its derivatives from Auricularia-A review. Int. J. Biol. Macromol. 2020, 150, 102–113. [Google Scholar] [CrossRef] [PubMed]
- Moradi, Z.; Kalanpour, N. Kefiran, a branched polysaccharide: Preparation, properties and applications: A review. Carbohydr. Polym. 2019, 223, 115100. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Chen, Z.; Zhou, H.; Yu, C.; Han, Z.; Shao, S.; Hu, X.; Wei, X.; Wang, Y. Effects of extraction methods on physicochemical properties and hypoglycemic activities of polysaccharides from coarse green tea. Glycoconj. J. 2020, 37, 241–250. [Google Scholar] [CrossRef]
- Udchumpisai, W.; Bangyeekhun, E. Purification, Structural Characterization, and Biological Activity of Polysaccharides from Lentinus velutinus. Mycobiology 2020, 48, 51–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, X.Y.; Liu, R.L.; Ma, X.; Zhang, Z.Q. Polyethylene glycol as a novel solvent for extraction of crude polysaccharides from Pericarpium granati. Carbohydr. Polym. 2014, 101, 886–889. [Google Scholar] [CrossRef] [PubMed]
- Teng, C.; Qin, P.; Shi, Z.; Zhang, W.; Ren, G. Structural characterization and antioxidant activity of alkali-extracted polysaccharides from quinoa. Food Hydrocoll. 2020, 113, 106392. [Google Scholar] [CrossRef]
- Morales, D.; Smiderle, F.R.; Villalva, M.; Abreu, H.; Rico, C.; Santoyo, S.; Iacomini, M.; Soler-Rivas, C. Testing the effect of combining innovative extraction technologies on the biological activities of obtained β-glucan-enriched fractions from Lentinula edodes. J. Funct. Foods 2019, 60, 103446. [Google Scholar] [CrossRef]
- Liu, C. Extraction, separation and purification of acidic polysaccharide from Morchella esculenta by high voltage pulsed electric field. Int. J. Bioaut. 2019, 23, 193–202. [Google Scholar] [CrossRef] [Green Version]
- Parniakov, O.; Lebovka, N.I.; Hecke, E.V.; Vorobiev, E. Pulsed electric field assisted pressure extraction and solvent extraction from mushroom (Agaricus Bisporus). Food Bioprocess. Technol. 2014, 7, 174–183. [Google Scholar] [CrossRef]
- Xujie, H.; Wei, C. Optimization of extraction process of crude polysaccharides from wild edible BaChu mushroom by response surface methodology. Carbohydr. Polym. 2008, 72, 67–74. [Google Scholar] [CrossRef]
- Zhang, J.; Yu, Y.; Zhang, Z.; Ding, Y.; Dai, X.; Li, Y. Effect of polysaccharide from cultured Cordyceps sinensis on immune function and anti-oxidation activity of mice exposed to 60Co. Int. Immunopharmacol. 2011, 11, 2251–2257. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Wang, Y.; Wang, Y.; Chen, T.; Tang, H.; Wang, M. Purification, characterization and immuno-modulating properties of polysaccharides isolated from Flammulina velutipes mycelium. Am. J. Chin. Med. 2010, 38, 191–204. [Google Scholar] [CrossRef]
- Morales, D.; Smiderle, F.R.; Piris, A.J.; Soler-Rivas, C.; Prodanov, M. Production of a β-D-glucan-rich extract from Shiitake mushrooms (Lentinula edodes) by an extraction/microfiltration/reverse osmosis (nanofiltration) process. Innov. Food Sci. Emerg. Technol. 2019, 51, 80–90. [Google Scholar] [CrossRef]
- Dey, B.; Bhunia, S.K.; Maity, K.K.; Patra, S.; Mandal, S.; Maiti, S.; Maiti, T.K.; Sikdar, S.R.; Islam, S.S. Chemical analysis of an immunoenhancing water-soluble polysaccharide of an edible mushroom, Pleurotus florida blue variant. Carbohydr. Res. 2010, 345, 2736–2741. [Google Scholar] [CrossRef] [PubMed]
- Han, X.Q.; Wu, X.M.; Chai, X.Y.; Chen, D.; Dai, H.; Dong, H.L.; Ma, Z.Z.; Gao, X.M.; Tu, P.F. Isolation, characterization and immunological activity of a polysaccharide from the fruit bodies of an edible mushroom, Sarcodon aspratus (Berk.) S. Ito. Food Res. Int. 2011, 44, 489–493. [Google Scholar] [CrossRef]
- Wang, K.P.; Zhang, Q.L.; Liu, Y.; Wang, J.; Cheng, Y.; Zhang, Y. Structure and inducing tumor cell apoptosis activity of polysaccharides isolated from Lentinus edodes. J. Agric. Food Chem. 2013, 61, 9849–9858. [Google Scholar] [CrossRef]
- Kanwal, S.; Joseph, T.P.; Owusu, L.; Xiaomeng, R.; Meiqi, L.; Yi, X. A polysaccharide isolated from Dictyophora indusiata promotes recovery from antibiotic-driven intestinal dysbiosis and improves gut epithelial barrier function in a mouse model. Nutrients 2018, 10, 1003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mfopa, A.; Mediesse, F.K.; Mvongo, C.; Nkoubatchoundjwen, S.; Lum, A.A.; Sobngwi, E.; Kamgang, R.; Boudjeko, T. Antidyslipidemic potential of water-soluble polysaccharides of Ganoderma applanatum in MACAPOS-2-induced obese rats. Evid. Based Complement. Altern. Med. 2021, 2021, 2452057. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Zhang, Y.; Wang, X.; Huang, X.; Fei, Y.; Yu, Y.; Shou, D. Antioxidant property of water-soluble polysaccharides from Poria cocos Wolf using different extraction methods. Int. J. Biol. Macromol. 2016, 83, 103–110. [Google Scholar] [CrossRef]
- Xue, D.; Farid, M.M. Pulsed electric field extraction of valuable compounds from white button mushroom (Agaricus bisporus). Innov. Food Sci. Emerg. Technol. 2015, 29, 178–186. [Google Scholar] [CrossRef]
- Wang, Y.; Jia, J.; Ren, X.; Li, B.; Zhang, Q. Extraction, preliminary characterization and in vitro antioxidant activity of polysaccharides from Oudemansiella radicata mushroom. Int. J. Biol. Macromol. 2018, 120, 1760–1769. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhou, Y.; Liu, M.; Wang, Q.; Li, Y. Extraction optimization, characterization, antioxidant and immunomodulatory activities of a novel polysaccharide from the wild mushroom Paxillus involutus. Int. J. Biol. Macromol. 2018, 112, 326–332. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wen, C.; Gu, J.; Ji, C.; Duan, Y.; Zhang, H. Effects of subcritical water extraction microenvironment on the structure and biological activities of polysaccharides from Lentinus edodes. Int. J. Biol. Macromol. 2019, 123, 1002–1011. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Huang, X.; Peng, Q.; Shan, Y.; Xue, F. Physicochemical properties of peanut protein isolate-glucomannan conjugates prepared by ultrasonic treatment. Ultrason. Sonochem. 2014, 21, 1722–1727. [Google Scholar] [CrossRef]
- Ke, L.Q. Optimization of ultrasonic extraction of polysaccharides from Lentinus edodes based on enzymatic treatment. J. Food Process. Preserv. 2015, 39, 254–259. [Google Scholar] [CrossRef]
- Alzorqi, I.; Sudheer, S.; Lu, T.J.; Manickam, S. Ultrasonically extracted β-D-glucan from artificially cultivated mushroom, characteristic properties and antioxidant activity. Ultrason. Sonochem. 2017, 35, 531–540. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Wang, W.P.; Zhang, H.S.; Huang, Q. Optimization of ultrasonic-assisted extraction of water-soluble polysaccharides from Boletus edulis mycelia using response surface methodology. Carbohydr. Polym. 2012, 87, 614–619. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Yong, F.; Liang, J.; Hu, Q. Optimization of ultrasonic extraction of Flammulina velutipes polysaccharides and evaluation of its acetylcholinesterase inhibitory activity. Food Res. Int. 2011, 44, 1269–1275. [Google Scholar] [CrossRef]
- Cheung, Y.C.; Siu, K.C.; Wu, J.Y. Kinetic models for ultrasound-assisted extraction of water-soluble components and polysaccharides from medicinal fungi. Food Bioprocess. Technol. 2013, 6, 2659–2665. [Google Scholar] [CrossRef]
- Zhao, L.Y.; Fu, Y.X.; Chen, C.; Yang, W.J.; Hu, Q.H. Ultrasonic-assisted extraction and chromatography separation of polysaccharides from the base of Flammulina velutipes Stipe. Sep. Sci. Technol. 2014, 50, 824–832. [Google Scholar] [CrossRef]
- Chen, Y.; Liu, D.; Wang, D.; Lai, S.; Zhong, R.; Liu, Y.; Yang, C.; Liu, B.; Sarker, M.R.; Zhao, C. Hypoglycemic activity and gut microbiota regulation of a novel polysaccharide from Grifola frondosa in type 2 diabetic mice. Food Chem. Toxicol. 2019, 126, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.K.; Wang, Y.Y.; Ma, H.L.; Wang, Z.B. Ultrasonic effects on the degradation kinetics, preliminary characterization and antioxidant activities of polysaccharides from Phellinus linteus mycelia. Ultrason. Sonochem. 2016, 29, 251–257. [Google Scholar] [CrossRef] [PubMed]
- Rombaut, N.; Tixier, A.S.; Bily, A.; Chemat, F. Green extraction processes of natural products as tools for biorefinery. Biofuels Bioprod. Biorefin. 2014, 8, 530–544. [Google Scholar] [CrossRef]
- Yan, S.; Pan, C.; Yang, X.; Chen, S.; Qi, B.; Huang, H. Degradation of Codium cylindricum polysaccharides by H2O2-Vc-ultrasonic and H2O2-Fe2+-ultrasonic treatment: Structural characterization and antioxidant activity. Int. J. Biol. Macromol. 2021, 182, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wang, L. Effect of extraction method on structure and antioxidant activity of Hohenbuehelia serotina polysaccharides. Int. J. Biol. Macromol. 2016, 83, 270–276. [Google Scholar] [CrossRef]
- Delazar, A.; Nahar, L.; Hamedeyazdan, S.; Sarker, S.D. Microwave-assisted extraction in natural products isolation. Methods Mol. Biol. 2012, 864, 89–115. [Google Scholar]
- Xu, N.; Sun, Y.H.; Guo, X.L.; Liu, C.; Qian, M.; Hou, J.M. Optimization of ultrasonic-microwave synergistic extraction of polysaccharides from Morchella conica. J. Food Process. Preserv. 2018, 42, 13423. [Google Scholar] [CrossRef]
- Chen, J.; Lai, P.; Shen, H.; Zhen, H.; Fang, R. Effect of extraction methods on polysaccharide of Clitocybe maxima stipe. Adv. J. Food Sci. Technol. 2013, 5, 370–373. [Google Scholar] [CrossRef]
- Gil-Ramírez, A.; Smiderle, F.R.; Morales, D.; Iacomini, M.; Soler-Rivas, C. Strengths and weaknesses of the aniline-blue method used to test mushroom (1→3)-β-D-glucans obtained by microwave-assisted extractions. Carbohydr. Polym. 2019, 217, 135–143. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Z.Y.; Dong, F.; Liu, X.; Lv, Q.; Liu, F.; Chen, L.; Wang, T.; Wang, Z.; Zhang, Y. Effects of extraction methods on the yield, chemical structure and anti-tumor activity of polysaccharides from Cordyceps gunnii mycelia. Carbohydr. Polym. 2016, 140, 461–471. [Google Scholar] [CrossRef] [PubMed]
- You, Q.; Yin, X.; Zhang, S.; Jiang, Z. Extraction, purification, and antioxidant activities of polysaccharides from Tricholoma mongolicum Imai. Carbohydr. Polym. 2014, 99, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Cui, J.; Chen, A.H.; Zong, Z.M.; Wei, X.Y. Optimization of ultrasonic-microwave assisted extraction and hepatoprotective activities of polysaccharides from Trametes orientalis. Molecules 2019, 24, 147. [Google Scholar] [CrossRef] [Green Version]
- Marić, M.; Grassino, A.N.; Zhu, Z.; Barba, F.J.; Brnčić, M.; Rimac Brnčić, S. An overview of the traditional and innovative approaches for pectin extraction from plant food wastes and by-products: Ultrasound-, microwaves-, and enzyme-assisted extraction. Trends Food Sci. Technol. 2018, 76, 28–37. [Google Scholar] [CrossRef]
- Liang, L.; Xiaoyu, Y.; Lei, P.; Yue, S.; Yan, W. Comparing three methods of extraction of Auricularia auricula polysaccharides. Curr. Top. Nutraceutical Res. 2019, 17, 7–10. [Google Scholar] [CrossRef]
- Yu, G.P.; Yue, C.H.; Zang, X.D.; Chen, C.; Dong, L.W.; Liu, Y.Q. Purification, characterization and in vitro bile salt-binding capacity of polysaccharides from Armillaria mellea mushroom. Czech. J. Food Sci. 2019, 37, 51–56. [Google Scholar] [CrossRef]
- Zhao, Y.M.; Wang, J.; Wu, Z.G.; Yang, J.M.; Li, W.; Shen, L.X. Extraction, purification and anti-proliferative activities of polysaccharides from Lentinus edodes. Int. J. Biol. Macromol. 2016, 93 Pt A, 136–144. [Google Scholar] [CrossRef]
- Fan, Y.; Wu, X.; Zhang, M.; Zhao, T.; Zhou, Y.; Han, L.; Yang, L. Physical characteristics and antioxidant effect of polysaccharides extracted by boiling water and enzymolysis from Grifola frondosa. Int. J. Biol. Macromol. 2011, 48, 798–803. [Google Scholar] [CrossRef]
- Yi, Y.; Xu, W.; Wang, H.X.; Huang, F.; Wang, L.M. Natural polysaccharides experience physiochemical and functional changes during preparation: A review. Carbohydr. Polym. 2020, 234, 115896. [Google Scholar] [CrossRef] [PubMed]
- Klaus, A.; Kozarski, M.; Niksic, M.; Jakovljevic, D.; Todorovic, N.; Van Griensven, L. Antioxidative activities and chemical characterization of polysaccharides extracted from the basidiomycete Schizophyllum commune. LWT-Food Sci. Technol. 2011, 44, 2005–2011. [Google Scholar] [CrossRef]
- Wang, Z.-B.; Pei, J.-J.; Ma, H.-L.; Cai, P.-F.; Yan, J.-K. Effect of extraction media on preliminary characterizations and antioxidant activities of Phellinus linteus polysaccharides. Carbohydr. Polym. 2014, 109, 49–55. [Google Scholar] [CrossRef]
- Baeva, E.; Bleha, R.; Lavrova, E.; Sushytskyi, L.; Čopíková, J.; Jablonsky, I.; Klouček, P.; Synytsya, A. Polysaccharides from Basidiocarps of cultivating mushroom Pleurotus ostreatus: Isolation and structural characterization. Molecules 2019, 24, 2740. [Google Scholar] [CrossRef]
- Yang, S.; Yan, J.; Yang, L.; Meng, Y.; Wang, N.; He, C.; Fan, Y.; Zhou, Y. Alkali-soluble polysaccharides from mushroom fruiting bodies improve insulin resistance. Int. J. Biol. Macromol. 2019, 126, 466–474. [Google Scholar] [CrossRef] [PubMed]
- Nandi, A.K.; Samanta, S.; Maity, S.; Sen, I.K.; Islam, S.S. Antioxidant and immunostimulant β-glucan from edible mushroom Russula albonigra (Krombh.) Fr. Carbohydr. Polym. 2014, 99, 774–782. [Google Scholar] [CrossRef] [PubMed]
- Sermwittayawong, D.; Patninan, K.; Phothiphiphit, S.; Boonyarattanakalin, S.; Sermwittayawong, N.; Hutadilok-Towatana, N. Purification, characterization, and biological activities of purified polysaccharides extracted from the gray oyster mushroom [Pleurotus sajor-caju (Fr.) Sing.]. J. Food Biochem. 2018, 42, 12606. [Google Scholar] [CrossRef]
- Szwengiel, A.; Stachowiak, B. Deproteinization of water-soluble ß-glucan during acid extraction from fruiting bodies of Pleurotus ostreatus mushrooms. Carbohydr. Polym. 2016, 146, 310–319. [Google Scholar] [CrossRef]
- Zhang, B.; Li, Y.; Zhang, F.; Linhardt, R.J.; Zeng, G.; Zhang, A. Extraction, structure and bioactivities of the polysaccharides from Pleurotus eryngii: A review. Int. J. Biol. Macromol. 2020, 150, 1342–1347. [Google Scholar] [CrossRef]
- Xu, Y.; Shen, M.; Chen, Y.; Lou, Y.; Luo, R.; Chen, J.; Zhang, Y.; Li, J.; Wang, W. Optimization of the polysaccharide hydrolysate from Auricularia auricula with antioxidant activity by response surface methodology. Int. J. Biol. Macromol. 2018, 113, 543–549. [Google Scholar] [CrossRef]
- Gu, J.; Li, Q.; Liu, J.; Ye, Z.; Feng, T.; Wang, G.; Wang, W.; Zhang, Y. Ultrasonic-assisted extraction of polysaccharides from Auricularia auricula and effects of its acid hydrolysate on the biological function of Caenorhabditis elegans. Int. J. Biol. Macromol. 2021, 167, 423–433. [Google Scholar] [CrossRef]
- Yan, J.K.; Ding, Z.C.; Gao, X.; Wang, Y.Y.; Yang, Y.; Wu, D.; Zhang, H.N. Comparative study of physicochemical properties and bioactivity of Hericium erinaceus polysaccharides at different solvent extractions. Carbohydr. Polym. 2018, 193, 373–382. [Google Scholar] [CrossRef]
- Su, C.A.; Xu, X.Y.; Liu, D.Y.; Ming, W. Isolation and characterization of exopolysaccharide with immunomodulatory activity from fermentation broth of Morchella conica. Daru J. Pharm. Sci. 2013, 21, 5. [Google Scholar] [CrossRef] [Green Version]
- He, P.; Geng, L.; Mao, D.; Wang, Z.; Xu, C. Production, preliminary characterization, and bioactivities of exopolysaccharides from Pleurotus geesteranus 5(#). Prep. Biochem. Biotechnol. 2013, 43, 108–122. [Google Scholar]
- Mao, D.B.; Ma, Y.P.; Geng, L.J.; Zhang, A.J.; Zheng, J.Q.; Xu, C.P. Fermentation characteristics in stirred-tank reactor of exopolysaccharides with hypolipidemic activity produced by Pleurotus geesteranus 5#. An. Acad. Bras. Cienc. 2013, 85, 1473–1481. [Google Scholar]
- Mao, Y.; Mao, J.; Meng, X. Extraction optimization and bioactivity of exopolysaccharides from Agaricus bisporus. Carbohydr. Polym. 2013, 92, 1602–1607. [Google Scholar] [CrossRef]
- Cao, H.; Ma, S.; Guo, H.; Cui, X.; Wang, S.; Zhong, X.; Wu, Y.; Zheng, W.; Wang, H.; Yu, J.; et al. Comparative study on the monosaccharide compositions, antioxidant and hypoglycemic activities in vitro of intracellular and extracellular polysaccharides of liquid fermented Coprinus comatus. Int. J. Biol. Macromol. 2019, 139, 543–549. [Google Scholar] [CrossRef]
- Rodríguez-Seoane, P.; Díaz-Reinoso, B.; González-Muñoz, M.J.; Fernández de Ana Portela, C.; Domínguez, H. Innovative technologies for the extraction of saccharidic and phenolic fractions from Pleurotus eryngii. LWT-Food Sci. Technol. 2019, 101, 774–782. [Google Scholar] [CrossRef]
- Huamán-Leandro, L.R.; González-Muñoz, M.J.; Fernández-de-Ana, C.; Rodríguez-Blanco, A.; Torres, M.D.; Domínguez, H. Autohydrolysis of Lentinus edodes for obtaining extracts with antiradical properties. Foods 2020, 9, 74. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Sun, Y.; Mao, Q.; Guo, X.; Li, P.; Liu, Y.; Xu, N. Characteristics and antitumor activity of Morchella esculenta polysaccharide extracted by pulsed electric field. Int. J. Mol. Sci. 2016, 17, 986. [Google Scholar] [CrossRef] [Green Version]
- Parniakov, O.; Barba, F.J.; Grimi, N.; Marchal, L.; Jubeau, S.; Lebovka, N.; Vorobiev, E. Pulsed electric field assisted extraction of nutritionally valuable compounds from microalgae Nannochloropsis spp. using the binary mixture of organic solvents and water. Innov. Food Sci. Emerg. Technol. 2015, 27, 79–85. [Google Scholar] [CrossRef]
- Yuan, Y.; Liu, Y.; Liu, M.; Chen, Q.; Jiao, Y.; Liu, Y.; Meng, Z. Optimization extraction and bioactivities of polysaccharide from wild Russula griseocarnosa. Saudi Pharm. J. 2017, 25, 523–530. [Google Scholar] [CrossRef]
- Lin, Y.; Zeng, H.; Wang, K.; Lin, H.; Li, P.; Huang, Y.; Zhou, S.; Zhang, W.; Chen, C.; Fan, H. Microwave-assisted aqueous two-phase extraction of diverse polysaccharides from Lentinus edodes: Process optimization, structure characterization and antioxidant activity. Int. J. Biol. Macromol. 2019, 136, 305–315. [Google Scholar] [CrossRef]
- Ke, L.; Chen, H. Homogenate extraction of crude polysaccharides from Lentinus edodes and evaluation of the antioxidant activity. Food Sci. Technol. 2016, 36, 533–539. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Wang, A.; Liu, L.; Tian, G.; Xu, F. Extraction of polysaccharides under vacuum condition from Lentinus edodes stipe and their antioxidant activities in vitro. Food Sci. Biotechnol. 2019, 28, 759–767. [Google Scholar] [CrossRef]
- Park, H.G.; Shim, Y.Y.; Choi, S.O.; Park, W.M. New Method Development for Nanoparticle Extraction of Water-Soluble beta-(1 -> 3)-D-Glucan from Edible Mushrooms, Sparassis crispa and Phellinus linteus. J. Agric. Food Chem. 2009, 57, 2147–2154. [Google Scholar] [CrossRef]
- Yin, C.; Fan, X.; Fan, Z.; Shi, D.; Gao, H. Optimization of enzymes-microwave-ultrasound assisted extraction of Lentinus edodes polysaccharides and determination of its antioxidant activity. Int. J. Biol. Macromol. 2018, 111, 446–454. [Google Scholar] [CrossRef]
- Maity, G.N.; Maity, P.; Khatua, S.; Acharya, K.; Dalai, S.; Mondal, S. Structural features and antioxidant activity of a new galactoglucan from edible mushroom Pleurotus djamor. Int. J. Biol. Macromol. 2021, 168, 743–749. [Google Scholar] [CrossRef]
- Wu, S.J.; Liaw, C.C.; Pan, S.Z.; Yang, H.C.; Ng, L.T. Phellinus linteus polysaccharides and their immunomodulatory properties in human monocytic cells. J. Funct. Foods 2013, 5, 679–688. [Google Scholar] [CrossRef]
- Liao, W.; Luo, Z.; Liu, D.; Ning, Z.; Yang, J.; Ren, J. Structure characterization of a novel polysaccharide from Dictyophora indusiata and its macrophage immunomodulatory activities. J. Agric. Food Chem. 2015, 63, 535–544. [Google Scholar] [CrossRef]
- Zhang, L.; Liu, Y.; Ke, Y.; Liu, Y.; Luo, X.; Li, C.; Zhang, Z.; Liu, A.; Shen, L.; Chen, H.; et al. Antidiabetic activity of polysaccharides from Suillellus luridus in streptozotocin-induced diabetic mice. Int. J. Biol. Macromol. 2018, 119, 134–140. [Google Scholar] [CrossRef]
- Shu, X.; Zhang, Y.; Jia, J.; Ren, X.; Wang, Y. Extraction, purification and properties of water-soluble polysaccharides from mushroom Lepista nuda. Int. J. Biol. Macromol. 2019, 128, 858–869. [Google Scholar] [CrossRef]
- Xie, J.; Zou, L.; Xie, Y.; Wu, X.; Wang, L. Analysis of the monosaccharide composition of water-soluble Coriolus versicolor polysaccharides by ultra-performance liquid chromatography/photodiode array detector. Chromatographia 2021, 84, 615–622. [Google Scholar] [CrossRef]
- Liang, J.; Zhang, M.; Wang, X.; Ren, Y.; Yue, T.; Wang, Z.; Gao, Z. Edible fungal polysaccharides, the gut microbiota, and host health. Carbohydr. Polym. 2021, 273, 118558. [Google Scholar] [CrossRef]
- Maity, P.; Sen, I.K.; Chakraborty, I.; Mondal, S.; Bar, H.; Bhanja, S.K.; Mandal, S.; Maity, G.N. Biologically active polysaccharide from edible mushrooms: A review. Int. J. Biol. Macromol. 2021, 172, 408–417. [Google Scholar] [CrossRef]
- Synytsya, A.; Novák, M. Structural diversity of fungal glucans. Carbohydr. Polym. 2013, 92, 792–809. [Google Scholar] [CrossRef] [PubMed]
- Bao, Z.; Yao, L.; Zhang, X.; Lin, S. Isolation, purification, characterization, and immunomodulatory effects of polysaccharide from Auricularia auricula on RAW264.7 macrophages. J. Food Biochem. 2020, 44, 13516. [Google Scholar] [CrossRef]
- Biscaia, S.M.P.; Carbonero, E.R.; Bellan, D.L.; Borges, B.S.; Costa, C.R.; Rossi, G.R.; Gonçalves, J.P.; Melo, C.M.; Lívero, F.A.R.; Ruthes, A.C.; et al. Safe therapeutics of murine melanoma model using a novel antineoplasic, the partially methylated mannogalactan from Pleurotus eryngii. Carbohydr. Polym. 2017, 178, 95–104. [Google Scholar] [CrossRef]
- Liu, G.; Ye, J.; Li, W.; Zhang, J.; Wang, Q.; Zhu, X.A.; Miao, J.Y.; Huang, Y.H.; Chen, Y.J.; Cao, Y. Extraction, structural characterization, and immunobiological activity of ABP Ia polysaccharide from Agaricus bisporus. Int. J. Biol. Macromol. 2020, 162, 975–984. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.K.; Li, N.; Song, S.Y.; Zhang, Y.J.; Wang, J.R. Three exopolysaccharides from the liquid fermentation of Polyporus umbellatus and their bioactivities. Int. J. Biol. Macromol. 2019, 132, 629–640. [Google Scholar] [CrossRef]
- Li, X.; Xu, W. TLR4-mediated activation of macrophages by the polysaccharide fraction from Polyporus umbellatus(pers.) Fries. J. Ethnopharmacol. 2011, 135, 1–6. [Google Scholar] [CrossRef]
- He, P.; Zhang, A.; Zhang, F.; Linhardt, R.J.; Sun, P. Structure and bioactivity of a polysaccharide containing uronic acid from Polyporus umbellatus sclerotia. Carbohydr. Polym. 2016, 152, 222–230. [Google Scholar] [CrossRef] [PubMed]
- He, P.F.; Zhang, A.Q.; Wang, X.L.; Qu, L.; Li, G.L.; Li, Y.P.; Sun, P.L. Structure elucidation and antioxidant activity of a novel polysaccharide from Polyporus umbellatus sclerotia. Int. J. Biol. Macromol. 2016, 82, 411–417. [Google Scholar] [CrossRef]
- He, P.F.; He, L.; Zhang, A.Q.; Wang, X.L.; Qu, L.; Sun, P.L. Structure and chain conformation of a neutral polysaccharide from sclerotia of Polyporus umbellatus. Carbohydr. Polym. 2017, 155, 61–67. [Google Scholar] [CrossRef]
- Yan, J.; Meng, Y.; Zhang, M.; Zhou, X.; Cheng, H.; Sun, L.; Zhou, Y. A 3-O-methylated heterogalactan from Pleurotus eryngii activates macrophages. Carbohydr. Polym. 2019, 206, 706–715. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Yong, Y.; Xia, X.; Wang, Z.; Liang, Y.; Zhang, S.; Lu, L. The excreted polysaccharide of Pleurotus eryngii inhibits the foam-cell formation via down-regulation of CD36. Carbohydr. Polym. 2014, 112, 16–23. [Google Scholar] [CrossRef]
- Maity, P.; Sen, I.K.; Maji, P.K.; Paloi, S.; Devi, K.S.; Acharya, K.; Maiti, T.K.; Islam, S.S. Structural, immunological, and antioxidant studies of β-glucan from edible mushroom Entoloma lividoalbum. Carbohydr. Polym. 2015, 123, 350–358. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Wang, J.; Hu, H.; Li, Q.; Liu, Y.; Wang, K. Functional polysaccharide Lentinan suppresses human breast cancer growth via inducing autophagy and caspase-7-mediated apoptosis. J. Funct. Foods 2018, 45, 75–85. [Google Scholar] [CrossRef]
- Pan, H.; Wang, Y.; Na, K.; Wang, Y.; Wang, L.; Li, Z.; Guo, C.; Guo, D.; Wang, X. Autophagic flux disruption contributes to Ganoderma lucidum polysaccharide-induced apoptosis in human colorectal cancer cells via MAPK/ERK activation. Cell. Death Dis. 2019, 10, 456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.M.; Zhong, R.F.; Chen, J.; Luo, Z.G. Structural characterization, anticancer, hypoglycemia and immune activities of polysaccharides from Russula virescens. Int. J. Biol. Macromol. 2021, 184, 380–392. [Google Scholar] [CrossRef]
- Ma, Q.; Jiang, J.G.; Yuan, X.; Qiu, K.; Zhu, W. Comparative antitumor and anti-inflammatory effects of flavonoids, saponins, polysaccharides, essential oil, coumarin and alkaloids from Cirsium japonicum DC. Food Chem. Toxicol. 2019, 125, 422–429. [Google Scholar] [CrossRef] [PubMed]
- Pawlikowska, M.; Jędrzejewski, T.; Brożyna, A.A.; Wrotek, S. Protein-bound polysaccharides from Coriolus versicolor induce RIPK1/RIPK3/MLKL-mediated necroptosis in ER-positive breast cancer and amelanotic melanoma cells. Cell. Physiol. Biochem. 2020, 54, 591–604. [Google Scholar] [PubMed]
- Pawlikowska, M.; Piotrowski, J.; Jędrzejewski, T.; Kozak, W.; Slominski, A.T.; Brożyna, A.A. Coriolus versicolor-derived protein-bound polysaccharides trigger the caspase-independent cell death pathway in amelanotic but not melanotic melanoma cells. Phytother. Res. 2020, 34, 173–183. [Google Scholar] [CrossRef]
- Li, W.; Wang, J.; Chen, Z.; Gao, X.; Chen, Y.; Xue, Z.; Guo, Q.; Ma, Q.; Chen, H. Physicochemical properties of polysaccharides from Lentinus edodes under high pressure cooking treatment and its enhanced anticancer effects. Int. J. Biol. Macromol. 2018, 115, 994–1001. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; He, Y.; Zeng, P.; Liu, Y.; Zhang, M.; Hao, C.; Wang, H.; Lv, Z.; Zhang, L. Molecular basis for Poria cocos mushroom polysaccharide used as an antitumour drug in China. J. Cell. Mol. Med. 2019, 23, 4–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalmo, R.A.; Bøgwald, J. Beta-glucans as conductors of immune symphonies. Fish Shellfish Immunol. 2008, 25, 384–396. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.O.; Park, H.W.; Kim, J.H.; Lee, J.Y.; Moon, S.H.; Shin, C.S. Anti-cancer effect and structural characterization of endo-polysaccharide from cultivated mycelia of Inonotus obliquus. Life Sci. 2006, 79, 72–80. [Google Scholar] [CrossRef]
- Su, Z.; Yang, Z.; Xu, Y.; Chen, Y.; Yu, Q. Apoptosis, autophagy, necroptosis, and cancer metastasis. Mol. Cancer 2015, 14, 48. [Google Scholar] [CrossRef] [Green Version]
- Carini, M.; Aldini, G.; Orioli, M.; Piccoli, A.; Rossoni, G.; Maffei Facino, R. Nitric oxide release and distribution following oral and intraperitoneal administration of nitroaspirin (NCX 4016) in the rat. Life Sci. 2004, 74, 3291–3305. [Google Scholar] [CrossRef] [Green Version]
- El Enshasy, H.A.; Hatti-Kaul, R. Mushroom immunomodulators: Unique molecules with unlimited applications. Trends Biotechnol. 2013, 31, 668–677. [Google Scholar] [CrossRef] [PubMed]
- Muszyńska, B.; Grzywacz-Kisielewska, A.; Kała, K.; Gdula-Argasińska, J. Anti-inflammatory properties of edible mushrooms: A review. Food Chem. 2018, 243, 373–381. [Google Scholar] [CrossRef] [PubMed]
- Won, D.P.; Lee, J.S.; Kwon, D.S.; Lee, K.E.; Shin, W.C.; Hong, E.K. Immunostimulating activity by polysaccharides isolated from fruiting body of Inonotus obliquus. Mol. Cells 2011, 31, 165–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, S.F.; Zhuang, T.F.; Si, Y.M.; Qi, K.Y.; Zhao, J. Coriolus versicolor mushroom polysaccharides exert immunoregulatory effects on mouse B cells via membrane Ig and TLR-4 to activate the MAPK and NF-κB signaling pathways. Mol. Immunol. 2015, 64, 144–151. [Google Scholar] [CrossRef]
- Jeong, J.B.; Seo, E.W.; Jeong, H.J. Effect of extracts from pine needle against oxidative DNA damage and apoptosis induced by hydroxyl radical via antioxidant activity. Food Chem. Toxicol. 2009, 47, 2135–2141. [Google Scholar] [CrossRef] [PubMed]
- Ji, X.; Ku, T.; Zhu, N.; Ning, X.; Wei, W.; Li, G.; Sang, N. Potential hepatic toxicity of buprofezin at sublethal concentrations: ROS-mediated conversion of energy metabolism. J. Hazard Mater. 2016, 320, 176–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, X.; Wang, Q.; Wang, Y.; Du, L.; Xu, C.; Liu, Q. Brusatol enhances the radiosensitivity of A549 cells by promoting ROS production and enhancing DNA damage. Int. J. Mol. Sci. 2016, 17, 997. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Zhou, H.; Zhang, J.; Li, F.; Wei, K.; Wei, X.; Wang, Y. Valorization of polysaccharides obtained from dark tea: Preparation, physicochemical, antioxidant, and hypoglycemic properties. Foods 2021, 10, 2276. [Google Scholar] [CrossRef]
- Sun, Y.-X.; Liu, J.-C.; Kennedy, J.F. Purification, composition analysis and antioxidant activity of different polysaccharide conjugates (APPs) from the fruiting bodies of Auricularia polytricha. Carbohydr. Polym. 2010, 82, 299–304. [Google Scholar] [CrossRef]
- Su, C.H.; Lai, M.N.; Ng, L.T. Effects of different extraction temperatures on the physicochemical properties of bioactive polysaccharides from Grifola frondosa. Food Chem. 2017, 220, 400–405. [Google Scholar] [CrossRef]
- Akram, K.; Shahbaz, H.M.; Kim, G.R.; Farooq, U.; Kwon, J.H. Improved extraction and quality characterization of water-soluble polysaccharide from gamma-irradiated Lentinus edodes. J. Food Sci. 2017, 82, 296–303. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Chen, X.; Li, F.; Wei, K.; Chen, J.; Wei, X.; Wang, Y. Preparation, physicochemical and hypoglycemic properties of natural selenium-enriched coarse tea glycoproteins. Plant Foods Hum. Nutr. 2022, 77, 258–264. [Google Scholar] [CrossRef] [PubMed]
- Shi, M.; Zhang, Z.; Yang, Y. Antioxidant and immunoregulatory activity of Ganoderma lucidum polysaccharide (GLP). Carbohydr. Polym. 2013, 95, 200–206. [Google Scholar] [CrossRef]
- Luo, A.; He, X.; Zhou, S.; Fan, Y.; Luo, A.; Chun, Z. Purification, composition analysis and antioxidant activity of the polysaccharides from Dendrobium nobile Lindl. Carbohydr. Polym. 2010, 79, 1014–1019. [Google Scholar] [CrossRef]
- Wang, J.; Hu, S.; Nie, S.; Yu, Q.; Xie, M. Reviews on mechanisms of in vitro antioxidant activity of polysaccharides. Oxid. Med. Cell. Longev. 2016, 2016, 1–13. [Google Scholar]
- Bisht, S.; Kant, R.; Kumar, V. α-D-Glucosidase inhibitory activity of polysaccharide isolated from Acacia tortilis gum exudate. Int. J. Biol. Macromol. 2013, 59, 214–220. [Google Scholar] [CrossRef]
- Wu, J.; Shi, S.; Wang, H.; Wang, S. Mechanisms underlying the effect of polysaccharides in the treatment of type 2 diabetes: A review. Carbohydr. Polym. 2016, 144, 474–494. [Google Scholar] [CrossRef] [PubMed]
- Khursheed, R.; Singh, S.K.; Wadhwa, S.; Gulati, M.; Awasthi, A.; Kumar, R.; Ramanunny, A.K.; Kapoor, B.; Kumar, P.; Corrie, L. Exploring role of probiotics and Ganoderma lucidum extract powder as solid carriers to solidify liquid self-nanoemulsifying delivery systems loaded with curcumin. Carbohydr. Polym. 2020, 250, 116996. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Yu, C.; Zhou, H.; Wei, X.; Wang, Y. Comparative evaluation for phytochemical composition and regulation of blood glucose, hepatic oxidative stress and insulin resistance in mice and HepG2 models of four typical Chinese dark teas. J. Sci. Food Agric. 2021, 101, 6563–6577. [Google Scholar] [CrossRef]
- Zhu, J.; Yu, C.; Han, Z.; Chen, Z.; Wei, X.; Wang, Y. Comparative analysis of existence form for selenium and structural characteristics in artificial selenium-enriched and synthetic selenized green tea polysaccharides. Int. J. Biol. Macromol. 2020, 154, 1408–1418. [Google Scholar] [CrossRef]
- Oboh, G.; Ademiluyi, A.O.; Akinyemi, A.J.; Henle, T.; Saliu, J.A.; Schwarzenbolz, U. Inhibitory effect of polyphenol-rich extracts of jute leaf (Corchorus olitorius) on key enzyme linked to type 2 diabetes (α-amylase and α-glucosidase) and hypertension (angiotensin I converting) in vitro. J. Funct. Foods 2012, 4, 450–458. [Google Scholar] [CrossRef]
- Feng, S.; Luan, D.; Ning, K.; Shao, P.; Sun, P. Ultrafiltration isolation, hypoglycemic activity analysis and structural characterization of polysaccharides from Brasenia schreberi. Int. J. Biol. Macromol. 2019, 135, 141–151. [Google Scholar] [CrossRef]
- Shen, M.; Fang, Z.; Chen, Y.; Chen, Y.; Xiao, B.; Guo, L.; Xu, Y.; Wang, G.; Wang, W.; Zhang, Y. Hypoglycemic Effect of the Degraded Polysaccharides from the Wood Ear Medicinal Mushroom Auricularia auricula-judae (Agaricomycetes). Int. J. Med. Mushrooms 2019, 21, 1033–1042. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Xia, Y.; Liu, D.; He, Y.; Mu, T.; Huo, Y.; Liu, J. Inhibitory effects of Lentinus edodes mycelia polysaccharide on α-glucosidase, glycation activity and high glucose-induced cell damage. Carbohydr. Polym. 2020, 246, 116659. [Google Scholar] [CrossRef]
- Wang, B.H.; Cao, J.J.; Zhang, B.; Chen, H.Q. Structural characterization, physicochemical properties and α-glucosidase inhibitory activity of polysaccharide from the fruits of wax apple. Carbohydr. Polym. 2019, 211, 227–236. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.M.; Dai, R.; Luo, Y.; Xiao, J.H. Glucose-lowering and hypolipidemic activities of polysaccharides from Cordyceps taii in streptozotocin-induced diabetic mice. BMC Complement. Altern. Med. 2019, 19, 230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakahara, D.; Nan, C.; Mori, K.; Hanayama, M.; Kikuchi, H.; Hirai, S.; Egashira, Y. Effect of mushroom polysaccharides from Pleurotus eryngii on obesity and gut microbiota in mice fed a high-fat diet. Eur. J. Nutr. 2020, 59, 3231–3244. [Google Scholar] [CrossRef]
- Kanwal, S.; Aliya, S.; Xin, Y. Anti-obesity effect of Dictyophora indusiata mushroom polysaccharide (DIP) in high fat diet-induced obesity via regulating inflammatory cascades and intestinal microbiome. Front. Endocrinol. 2020, 11, 558874. [Google Scholar] [CrossRef]
- Khan, I.; Huang, G.; Li, X.; Leong, W.; Xia, W.; Hsiao, W.L.W. Mushroom polysaccharides from Ganoderma lucidum and Poria cocos reveal prebiotic functions. J. Funct. Foods 2018, 41, 191–201. [Google Scholar] [CrossRef]
- Ma, L.; Chen, H.; Dong, P.; Lu, X. Anti-inflammatory and anticancer activities of extracts and compounds from the mushroom Inonotus obliquus. Food Chem. 2013, 139, 503–508. [Google Scholar] [CrossRef] [PubMed]
- Smiderle, F.R.; Alquini, G.; Tadra-Sfeir, M.Z.; Iacomini, M.; Wichers, H.J.; Van Griensven, L.J.L.D. Agaricus bisporus and Agaricus brasiliensis (1→6)-β-d-glucans show immunostimulatory activity on human THP-1 derived macrophages. Carbohydr. Polym. 2013, 94, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Liu, H.; Wang, W.; Wang, X.; Zhang, C.; Zhang, J.; Jing, H.; Ren, Z.; Gao, Z.; Song, X.; et al. Antioxidant and anti-aging effects of acidic-extractable polysaccharides by Agaricus bisporus. Int. J. Biol. Macromol. 2018, 106, 1297–1306. [Google Scholar] [CrossRef] [PubMed]
- Yuan, F.; Gao, Z.; Liu, W.; Li, H.; Jia, L. Characterization, antioxidant, anti-aging and organ protective effects of sulfated polysaccharides from Flammulina velutipes. Molecules 2019, 24, 3517. [Google Scholar] [CrossRef] [Green Version]
- Khursheed, R.; Singh, S.K.; Wadhwa, S.; Gulati, M.; Awasthi, A. Enhancing the potential preclinical and clinical benefits of quercetin through novel drug delivery systems. Drug Discov. Today 2020, 25, 209–222. [Google Scholar] [CrossRef]
- Zhu, J.; Wu, M.; Zhou, H.; Cheng, L.; Wei, X.; Wang, Y. Liubao brick tea activates the PI3K-Akt signaling pathway to lower blood glucose, metabolic disorders and insulin resistance via altering the intestinal flora. Food Res. Int. 2021, 148, 110594. [Google Scholar] [CrossRef]
- Kelly, S.D.; Neary, S.L. Ominous octet and other scary diabetes stories: The overview of pathophysiology of type 2 diabetes mellitus. Physician Assist. Clin. 2020, 5, 121–133. [Google Scholar] [CrossRef]
- Zhang, C.; Zhang, L.; Liu, H.; Zhang, J.; Hu, C.; Jia, L. Antioxidation, anti-hyperglycaemia and renoprotective effects of extracellular polysaccharides from Pleurotus eryngii SI-04. Int. J. Biol. Macromol. 2018, 111, 219–228. [Google Scholar] [CrossRef] [PubMed]
- Hruby, A.; Hu, F.B. The Epidemiology of Obesity: A Big Picture. Pharmacoeconomics 2015, 33, 673–689. [Google Scholar] [CrossRef] [PubMed]
- Jiang, T.; Gao, X.; Wu, C.; Tian, F.; Lei, Q.; Bi, J.; Xie, B.; Wang, H.Y.; Chen, S.; Wang, X. Apple-derived pectin modulates gut microbiota, improves gut barrier function, and attenuates metabolic endotoxemia in rats with diet-induced obesity. Nutrients 2016, 8, 126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, G.; Kimatu, B.M.; Zhao, L.; Yang, W.; Pei, F.; Hu, Q. In vivo fermentation of a Pleurotus eryngii polysaccharide and its effects on fecal microbiota composition and immune response. Food Funct. 2017, 8, 1810–1821. [Google Scholar] [CrossRef]
- Mosikanon, K.; Arthan, D.; Kettawan, A.; Tungtrongchitr, R.; Prangthip, P. Yeast β-glucan modulates inflammation and Waist circumference in overweight and obese subjects. J. Diet. Suppl. 2017, 14, 173–185. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Li, W.; Huang, X.; Liu, Y.; Li, Q.; Zheng, Z.; Wang, K. A polysaccharide from Lentinus edodes inhibits human colon cancer cell proliferation and suppresses tumor growth in athymic nude mice. Oncotarget 2017, 8, 610–623. [Google Scholar] [CrossRef] [Green Version]
- Harhaji, L.; Mijatović, S.; Maksimović-Ivanić, D.; Stojanović, I.; Momcilović, M.; Maksimović, V.; Tufegdzić, S.; Marjanović, Z.; Mostarica-Stojković, M.; Vucinić, Z.; et al. Anti-tumor effect of Coriolus versicolor methanol extract against mouse B16 melanoma cells: In vitro and in vivo study. Food Chem. Toxicol. 2008, 46, 1825–1833. [Google Scholar] [CrossRef]
- Niu, L.; Wu, Y.; Liu, H.; Wang, Q.; Li, M.; Jia, Q. The structural characterization of a novel water-soluble polysaccharide from edible mushroom Leucopaxillus giganteus and its antitumor activity on H22 tumor-bearing mice. Chem. Biodivers. 2021, 18, 2001010. [Google Scholar] [CrossRef]
- Yu, M.; Jia, H.M.; Zhou, C.; Yang, Y.; Sun, L.L.; Zou, Z.M. Urinary and fecal metabonomics study of the protective effect of Chaihu-Shu-Gan-San on antibiotic-induced gut microbiota dysbiosis in rats. Sci. Rep. 2017, 7, 46551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, C.J.; Sears, C.L.; Maruthur, N. Gut microbiome and its role in obesity and insulin resistance. Ann. N. Y. Acad Sci. 2020, 1461, 37–52. [Google Scholar] [CrossRef] [PubMed]
- Khursheed, R.; Singh, S.K.; Wadhwa, S.; Kapoor, B.; Gulati, M.; Kumar, R.; Ramanunny, A.K.; Awasthi, A.; Dua, K. Treatment strategies against diabetes: Success so far and challenges ahead. Eur. J. Pharmacol. 2019, 862, 172625. [Google Scholar] [CrossRef] [PubMed]
- Yin, C.; Noratto, G.D.; Fan, X.; Chen, Z.; Yao, F.; Shi, D.; Gao, H. The impact of mushroom polysaccharides on gut microbiota and its beneficial effects to host: A review. Carbohydr. Polym. 2020, 250, 116942. [Google Scholar] [CrossRef]
- Turnbaugh, P.J.; Ley, R.E.; Mahowald, M.A.; Magrini, V.; Mardis, E.R.; Gordon, J.I. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature 2006, 444, 1027–1031. [Google Scholar] [CrossRef]
- Larsen, N.; Vogensen, F.K.; van den Berg, F.W.; Nielsen, D.S.; Andreasen, A.S.; Pedersen, B.K.; Al-Soud, W.A.; Sørensen, S.J.; Hansen, L.H.; Jakobsen, M. Gut microbiota in human adults with type 2 diabetes differs from non-diabetic adults. PLoS ONE 2010, 5, e9085. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Sun, W.; Yu, N.; Sun, J.; Yu, X.; Li, X.; Xing, Y.; Yan, D.; Ding, Q.; Xiu, Z.; et al. Anti-diabetic effect of baicalein is associated with the modulation of gut microbiota in streptozotocin and high-fat-diet induced diabetic rats. J. Funct. Foods 2018, 46, 256–267. [Google Scholar] [CrossRef]
- Moro, C.; Palacios, I.; Lozano, M.; D’Arrigo, M.; Guillamón, E.; Villares, A.; Martínez, J.A.; García-Lafuente, A. Anti-inflammatory activity of methanolic extracts from edible mushrooms in LPS activated RAW 264.7 macrophages. Food Chem. 2012, 130, 350–355. [Google Scholar] [CrossRef]
- Enns, L.C.; Wiley, J.C.; Ladiges, W.C. Clinical relevance of transgenic mouse models for aging research. Crit. Rev. Eukaryot. Gene Expr. 2008, 18, 81–91. [Google Scholar] [CrossRef]
- Johansen, J.S.; Harris, A.K.; Rychly, D.J.; Ergul, A. Oxidative stress and the use of antioxidants in diabetes: Linking basic science to clinical practice. Cardiovasc. Diabetol. 2005, 4, 5. [Google Scholar] [CrossRef] [PubMed]















| Edible Fungus Origin | Monosaccharide Composition and Molar Ratio | Ref |
|---|---|---|
| Cordyceps sinensis | CP-PS, Man: Glc: Gal: xyl: Rha: Ara = 38.37: 27.44: 24.25: 5.22: 2.51:2.21, respectively | [21] |
| Flammulina velutipes | FVP2C, Glc: Gal: Man: Fuc = 100: 14: 7: 4, respectively | [22] |
| Phellinus linteus | PSCP, Glc: Man: Gal: N-Acetyl Glu: unidentified monosaccharide = 37.4: 2.2: 12.6: 29.5: 18.3, respectively | [86] |
| Lentinus edodes | SLNT1, Glc: Gly = 3.4: 1, SLNT2, Glc: Gly = 0.9: 1, JLNT1, Glc: Gly = 2.8: 1, JLNT2, Glc: Gly = 2.7: 1, JLNT2, JLNT3, Glc: Gly = 3: 1, respectively | [26] |
| Dictyophora indusiate | DP, Glc: Gal: Man = 56.2: 14.1: 29.7, respectively | [87] |
| Poria cocos Wolf | PCP-H, Man: Glc: Gal: Ara = 0.92: 0.18: 86.88: 12.01, PCP-M, Man: Glc: Gal: Ara = 4.02: 4.93: 79.48: 11.57, PCP-E, Man: Glc: Gal: Ara = 1.98: 0.36: 81.72: 15.93, PCP-U, Man: Glc: Gal: Ara = 2.18: 2.36: 87.27: 8.18, respectively | [29] |
| Grifola frondosa | GFP, L-Ara: D-Man: D-Glc = 3.79: 1.00: 49.70, respectively | [41] |
| Suillellus luridus | Suilu.A, Ara: xyl: Man: Glc: Gal = 2.06: 19.66: 46.07: 21.53, Suilu.C, Ara: xyl: Man: Glc: Gal = 2.71: 19.89: 41.90: 24.77, Suilu.S, Ara: xyl: Man: Glc: Gal = 4.36: 20.94: 49.09: 17.25, respectively | [88] |
| Coprinus comatus | EP, Man: Glu: Gal: Xyl: Ara = 6.45: 8.95: 9.64: 10.33: 11.34, IP, Man: Gal: Xyl = 6.46: 9.65: 10.35, respectively | [74] |
| Oudemansiella radicata | ORP-1, Man: Rib: Glc: Ga: xyl = 2.46: 2.63: 3.89: 3.22: 2.78, ORP-2, Rib: Glc: xyl = 2.63: 3.38: 2.65, ORP-3, Gl: xyl = 3.38: 2.65, respectively | [31] |
| Paxillus involutus | PIP2-1, Man: Glc: Gal: Fuc = 2.8: 62.2: 25.4: 9.6, respectively | [32] |
| Lepista nuda | LNP-1, Man: Glc: Gal: xyl: Ara: Fuc = 19.0: 33.5: 18.0: 4.6: 21.0: 3.9, LNP-2, Man: Glc: Gal: Ara: Fuc = 23.5: 11.4: 34.2: 21.4: 9.5, respectively | [89] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, Y.; He, H.; Wang, Q.; Yang, X.; Jiang, S.; Wang, D. A Review of Development and Utilization for Edible Fungal Polysaccharides: Extraction, Chemical Characteristics, and Bioactivities. Polymers 2022, 14, 4454. https://doi.org/10.3390/polym14204454
Sun Y, He H, Wang Q, Yang X, Jiang S, Wang D. A Review of Development and Utilization for Edible Fungal Polysaccharides: Extraction, Chemical Characteristics, and Bioactivities. Polymers. 2022; 14(20):4454. https://doi.org/10.3390/polym14204454
Chicago/Turabian StyleSun, Yujun, Huaqi He, Qian Wang, Xiaoyan Yang, Shengjuan Jiang, and Daobing Wang. 2022. "A Review of Development and Utilization for Edible Fungal Polysaccharides: Extraction, Chemical Characteristics, and Bioactivities" Polymers 14, no. 20: 4454. https://doi.org/10.3390/polym14204454